1 Department of Perinatology and Gynecology, University of Medical Sciences in Poznań, Poland 2 Human Reproductive Sciences Unit, Medical Research Council, Edinburgh, Scotland 3 Reproductive and Developmental Sciences, Simpson Centre for Reproductive Health, Edinburgh, Scotland Archives of Perinatal Medicine 15(2), 83-88, 2009 ORIGINAL PAPER Endocrine gland-derived vascular endothelial growth factor in etiology of pre-eclampsia – our experiences MICHAŁ SZUBER 1 , WIESŁAW MARKWITZ 1 , HENRY N. JABBOUR 2 , FIONA DENISON 3 , GRZEGORZ H. BRĘBOROWICZ 1 Abstract Background: Pre-eclampsia (PE) is a hypertensive complication of 7-10% of all pregnancies worldwide. Endo- crine gland-derived vascular endothelial growth factor (EG-VEGF, prokineticin 1 – PK-1) and its receptor (PKR1) has been identified. The main function of EG-VEGF is angiogenesis, and therefore it may play a crucial role in impaired endothelial functioning in pre-eclamptic pregnancies. Materials and methods: We designed two study groups: pregnant PE-patients and healthy controls. The samples were obtained during elective caesa- rean sections of the consented patients. We have collected myometrial and placental biopsies from 40 patients (19 were PE-patients). Tissue was collected and fixed or frozen. RNA was extracted and subjected to quan- titative PCR and fixed tissue was used for immunohistochemistry. A sample of every tissue was cultured in vitro, following stimulation with EG-VEGF and the activation of the intracellular pathways was assessed. Results: There is a peak response in 30 minutes after adding ligand (EG-VEGF) to the pre-incubated tissue. This pattern is alike in myometrium and placenta of the controls whereas in the PE-patients the placental response is blurred. The response to the ligand is stronger in placentas of the controls than in their myo- metriums whereas in PE-patients it is completely opposite. In immunohistochemistry slides greater presence of PK1 and PKR1 was seen in endothelial cells of the controls. RT-PCR shows greater expression of the PK1 in placentas of the controls than those of the PE-patients. Conclusions: From this stage of our research we can conclude that signaling and expression of EG-VEGF in PE-patients are disturbed. Key words: pre-eclampsia, prokineticin-1 (PK1), endocrine gland-derived vascular endothelial growth factor Background Pre-eclampsia (PE) is a hypertensive complication of 7-10% of all pregnancies worldwide, is responsible for approximately 40% of iatrogenic preterm deliveries and is associated with significant perinatal morbidity and mortality of the newborns. In Europe, hypertensive disorders of pregnancy are the second most common cause of maternal mortality whereas PE increases the future risk of developing hypertension, coronary heart disease and cerebro-vascular disease in mothers. In spite of recent findings, the patophysiology of PE still remains unclear. The pathogenesis of PE is thought to act at three levels: defective placentation, placental ischemia and endothelial cell dysfunction of which the latest is considered to be a key factor associated with PE. Endo- thelial cell dysfunction is believed to be responsible for several changes, such as increased endothelial-mediated vasoconstriction, increased vascular permeability and in- creased endothelial-mediated platelet aggregation lead- ing to maternal hypertension, proteinuria and thrombo- cytopenia. Prokineticin-1 (PK1) [also known as endocrine gland derived vascular endothelial growth factor (EG-VEGF)] is a recently described protein with a range of physiolo- gical and pathological functions [1]. The cognate recep- tors for PK1 are two closely related G protein-coupled receptors, PK receptor (PKR)-1 and PKR2. These couple to either Gi or Gq [2-4], activating downstream signaling pathways that include calcium mobilization, stimulation of phosphoinositide turnover, and activation of MAPK. PK1 is expressed in steroidogenic tissues including the placenta [5, 6], ovary [7, 8], and adrenal [3]. It mediates tissue specific vascular effects, which include capillary endothelial cell survival, proliferation, differentiation, and induction of fenestrae [3]. In contrast to VEGF, PK1 has no effect on endothelial cells derived from aorta, um- bilical vein, or cornea [5]. More recently, mouse studies have demonstrated that PKR1 gene activation promotes angiogenesis in cultured cardiac endothelial cells with- out increasing VEGF levels [9]. PK1 is also thought to have a role in immune regulation, affecting differentia- tion of human bone marrow cells into a distinct mono-
Transcript
1 Department of Perinatology and Gynecology, University of Medical Sciences in Poznań, Poland2 Human Reproductive Sciences Unit, Medical Research Council, Edinburgh, Scotland3 Reproductive and Developmental Sciences, Simpson Centre for Reproductive Health, Edinburgh, Scotland
Archives of Perinatal Medicine 15(2), 83-88, 2009 ORIGINAL PAPER
MICHAŁ SZUBER1, WIESŁAW MARKWITZ1, HENRY N. JABBOUR2, FIONA DENISON3, GRZEGORZ H. BRĘBOROWICZ1
AbstractBackground: Pre-eclampsia (PE) is a hypertensive complication of 7-10% of all pregnancies worldwide. Endo-crine gland-derived vascular endothelial growth factor (EG-VEGF, prokineticin 1 – PK-1) and its receptor(PKR1) has been identified. The main function of EG-VEGF is angiogenesis, and therefore it may play a crucialrole in impaired endothelial functioning in pre-eclamptic pregnancies. Materials and methods: We designedtwo study groups: pregnant PE-patients and healthy controls. The samples were obtained during elective caesa-rean sections of the consented patients. We have collected myometrial and placental biopsies from 40 patients(19 were PE-patients). Tissue was collected and fixed or frozen. RNA was extracted and subjected to quan-titative PCR and fixed tissue was used for immunohistochemistry. A sample of every tissue was cultured invitro, following stimulation with EG-VEGF and the activation of the intracellular pathways was assessed.Results: There is a peak response in 30 minutes after adding ligand (EG-VEGF) to the pre-incubated tissue.This pattern is alike in myometrium and placenta of the controls whereas in the PE-patients the placentalresponse is blurred. The response to the ligand is stronger in placentas of the controls than in their myo-metriums whereas in PE-patients it is completely opposite. In immunohistochemistry slides greater presenceof PK1 and PKR1 was seen in endothelial cells of the controls. RT-PCR shows greater expression of the PK1in placentas of the controls than those of the PE-patients. Conclusions: From this stage of our research wecan conclude that signaling and expression of EG-VEGF in PE-patients are disturbed.
BackgroundPre-eclampsia (PE) is a hypertensive complication
of 7-10% of all pregnancies worldwide, is responsible forapproximately 40% of iatrogenic preterm deliveries andis associated with significant perinatal morbidity andmortality of the newborns. In Europe, hypertensivedisorders of pregnancy are the second most commoncause of maternal mortality whereas PE increases thefuture risk of developing hypertension, coronary heartdisease and cerebro-vascular disease in mothers. In spiteof recent findings, the patophysiology of PE still remainsunclear. The pathogenesis of PE is thought to act atthree levels: defective placentation, placental ischemiaand endothelial cell dysfunction of which the latest isconsidered to be a key factor associated with PE. Endo-thelial cell dysfunction is believed to be responsible forseveral changes, such as increased endothelial-mediatedvasoconstriction, increased vascular permeability and in-creased endothelial-mediated platelet aggregation lead-ing to maternal hypertension, proteinuria and thrombo-cytopenia.
Prokineticin-1 (PK1) [also known as endocrine glandderived vascular endothelial growth factor (EG-VEGF)]is a recently described protein with a range of physiolo-gical and pathological functions [1]. The cognate recep-tors for PK1 are two closely related G protein-coupledreceptors, PK receptor (PKR)-1 and PKR2. These coupleto either Gi or Gq [2-4], activating downstream signalingpathways that include calcium mobilization, stimulationof phosphoinositide turnover, and activation of MAPK.PK1 is expressed in steroidogenic tissues including theplacenta [5, 6], ovary [7, 8], and adrenal [3]. It mediatestissue specific vascular effects, which include capillaryendothelial cell survival, proliferation, differentiation,and induction of fenestrae [3]. In contrast to VEGF, PK1has no effect on endothelial cells derived from aorta, um-bilical vein, or cornea [5]. More recently, mouse studieshave demonstrated that PKR1 gene activation promotesangiogenesis in cultured cardiac endothelial cells with-out increasing VEGF levels [9]. PK1 is also thought tohave a role in immune regulation, affecting differentia-tion of human bone marrow cells into a distinct mono-
M. Szuber, W. Markwitz, H.N. Jabbour, F. Denison, G.H. Bręborowicz84
cyte-derived cell population primed for release of pro-inflammatory cytokines [10]. On stimulation with LPS,PK1-primed monocytes/macrophages demonstrate anexaggerated release of IL-12 and TNF-α and down-re-gulated production of IL-10 [10], thus favoring a pro-inflammatory T-helper cell type 1 response. During thefirst trimester of pregnancy, PK1 and PKR1 are predo-minantly expressed in syncytiotrophoblast, with expres-sion peaking from 8-10 wk of gestation. PK1 is also ex-pressed in specialized macrophages called Hofbauercells in the placental villi from 6 wk of gestation. Incontrast, PK2 is not expressed in trophoblast and ex-pression of PKR2 is 80 times less than that of PKR1 [6].Supported by studies in the mouse [11], it has thereforebeen suggested that PK1 may have a role in trophoblastdifferentiation and placental angiogenesis during earlypregnancy. Less is known about placental expression ofPK1 and PKR1 in later pregnancy. Although PK1 ex-pression has been demonstrated in normal third-trimes-ter placenta, its immunolocalization and putative fun-ctions are not known [12]. Moreover, the expression andimmunolocalization of PKR1 in third trimester placentaremain to be investigated. There are also some studiesshowing no difference in EG-VEGF mRNA levelsbetween normal and PE placentas [12], however nearlyno studies investigating EG-VEGF protein expressionnor PKR1 and PKR2 mRNAs and proteins expressionwere conducted so far. The myometrium is also the tis-sue that has never been tested worldwide so far.
Materials and methods
The study had taken place in MRC in Edinburgh andwas subsequently followed in Department of Perina-tology and Gynaecology, University of Medical Sciencesin Poznan, Poland. We designed two study groups: preg-nant PE-patients (PE is defined as an onset of hyper-tension during late pregnancy, from 20th week of ges-tation until the 2nd week of puerperium) with systolic anddiastolic BP more than 140/90 mm Hg on at least twooccasions and urinary protein more than ++ on dipstick(or more than 0.5 g/24 H, or more than 300 mg/l, orACR > 30 mg/mmol) and otherwise healthy, pregnantwomen. The samples were obtained during the electivecaesarean sections of the consented patients from bothgroups (third trimester only). Collection of placentas andmyometrium samples was approved by the Ethics Com-mittee by the University of Medical Sciences in Poznań,Poland, and followed the recommended guidelines forusing human subjects. We have collected and processedmyometrial and placental biopsies from 40 patients so
far. Among those 19 were PE-patients. Each sample wasapproximately 3 by 2 cm in size. Tissue was collectedand divided into three samples. One part of every samplewas fixed overnight at 4EC in 4% paraformaldehyde in 10mM PBS and wax embedded for immunohistochemicalanalysis. Another sample was placed in RNA later, andRNA was extracted using an RNeasy Kit (Qiagen, Va-lencia, CA) according to the manufacturer’s instructionsand subjected to quantitative PCR. The tissue was ho-mogenized until the samples were uniformly homoge-neous. The concentration of RNA was measured usinga spectrophotometer, and RNA quality was confirmed onagarose gels. Real-time PCR was performed by moni-toring in real time the increase in fluorescence of theSYBR Green using 7900 HT Fast real-time PCR Systemby Iscience. In addition, a third sample of every tissuewas placed in RPMI (Sigma, Poole, UK) (containing2 mm l-glutamine, 100 IU penicillin, and 100 g/ml strep-tomycin) and transported to the laboratory for in vitroculture. Tissue was cultured in vitro, following stimu-lation with EG-VEGF and the activation of the intra-cellular pathways was assessed. After stimulation with li-gand (EG-VEGF) (time course – 0, 5, 10, 20, 30 minutesafter stimulation respectively) the tissues were homo-genized in lysis buffer [50 mM Tris-HCl (pH 7.5), 150mM NaCl, 1% sodium deoxycholate, 0.1% sodium dode-cyl sulfate, 1% Triton X-100, 1 mM phenylmethylsulfo-nylfluoride, 5 μg/ml leupeptin, 5 μg/ml aprotinin] byusing a Tissue lyser Qiagen by Retsch, until the sampleswere uniformly homogeneous. After centrifugation, pro-tein concentrations of the supernatant were determinedwith BSA (fraction V; Sigma, St. Louis, MO) as stan-dards. Placental and myometrial proteins were electro-phoretically separated on NuPAGE 4-12% Novex Bis-TrisGels by Invitrogen and electrically transferred onto0.25 μm polyvinylidene difluoride membranes. The blotswere washed with PBS-Tween 20 0.1% (PBS-T) andblocked in Odyssey Blocking Buffer for 1 hour. Then theblots were incubated with a mixture of antibodies againstpERK (pERK rabbit phospho p44/42 MAP Kinase byCell Signalling®) and tERK (tERK mouse p42 MAPKinase by Cell Signalling®) for 60 minutes. Afterwardsthe membranes were washed in PBS Tween and incu-bated in the mixture of secondary antibodies againstprimary antibodies (IRDye 800 Conjugated Affinity Pu-rified Anti-Mouse IgG (H&L) (Goat) by Rockland; AlexaFluor 680 goat anti-rabbit IgG (H+L) highly cross ab-sorbed 2 mg/ml by Molecular Probes) for 60 minutes.After several washes in PBS Tween and PBS the mem-branes were scanned using LI-COR Biosciences Odyssey
Endocrine gland-derived vascular endothelial growth factor in etiology of pre-eclampsia 85
Infrared Imaging System. The intensities of immunore-active bands were measured and the image analyzed ona desktop computer using LI-COR Biosciences Odysseysoftware. The mean pixel density for each band was ana-lyzed to obtain relative OD units for phosphorylation ofERK signalling proteins (Fig. 1, Fig. 2)
Fig. 1. Signalling of the pERK
Fig. 2. Signalling of the tERK
ResultsThe tissue is still being processed, and so far the
signalling processes are not completed, yet. In samplesthat are ready (about 50% of all samples) there is a pro-minent pattern with peak response (fold in pERK/tERKratio to minute “0”) in 30 minutes after adding ligand(EG-VEGF) to the pre-incubated tissue. This pattern isvery much alike in both myometrium and placenta of thecontrol group (Fig. 3, Fig. 4) whereas in the PE-patientsthe placental response is blurred (Fig. 5). We also dis-covered that the response to ligand is stronger in placen-tas of the control patients than in their myometriumswhereas in PE-patients it is completely opposite (Fig. 6).
Apart from signalling the immunohistochemistryexamination was performed. In those slides abundantpresence of PKR1 but not PK1 was seen in endothelialcells of the control (Fig. 8) while in PE-patients theamount of staining was slightly lower for the receptor
(PKR1) as well as for the ligand (PK1) (Fig. 7). In myo-metrium samples of both groups it was difficult to visua-lize even the receptor, not mentioning the ligand (EG-VEGF). Negative controls confirmed the physiologicalstructures of placentas and myometriums in both controland PE-patients.
Fig. 3. Myometriums’ pERK/tERK fold – normals
Fig. 4. Placentas’ pERK/tERK fold – normals
Fig. 5. Myometriums’ pERK/tERK fold – PE-group
Fig. 6. Placentas’ pERK/tERK fold – PE- group
M. Szuber, W. Markwitz, H.N. Jabbour, F. Denison, G.H. Bręborowicz86
Fig. 7. PKR1 (top) and PK1 (bottom) in preeclamptic placentas
As for the RNA assessment, table 1 shows the values ofPK1 and PKR1 expression in placenta and myometrium(myo), in PE-group and in controls (CTRL). The numbervalues of the expression correspond to the referencevalue which was the expression of PK1 and PKR1 in nor-mal, eutopic, second-phase endometrium.
Fig. 8. PKR1 (top) and PK1 (bottom) in normal placenta
In tables below (Tab. 2-5) the PE and control groupsare compared in terms of the statistic significance (P ) ofthe difference in the expression of PK1 and PKR1 show-ed as median.
Table 1. Values of the PK1 and PKR1 expression in placenta and myometrium (myo)in the PE-group and in the controls (CTRL)
Group n Mean Std Dev Std. Error Max Min Median 25% 75%
Table 2. Placenta PKR1 controls versus PE. No statistically significant differences were detected (P = 0.471)
Mann-Whitney Rank Sum Test
Normality Test: Failed (P < 0.050)
Group N Missing Median 25% 75%
Placenta CTRLPKR1 20 0 0.475 0.114 1.626
Placenta PKR1 12 0 0.934 0.39 1.019
Endocrine gland-derived vascular endothelial growth factor in etiology of pre-eclampsia 87
Table 3. Myometrium PKR1 controls versus PE. No statistically significant differences were detected (P = 0.487)
Mann-Whitney Rank Sum Test
Normality Test: Failed (P < 0.050)
Group N Missing Median 25% 75%
MyoCTRLPKR1 13 0 0.286 0.134 2.646
MyoPKR1 11 1 0.218 0.109 0.371
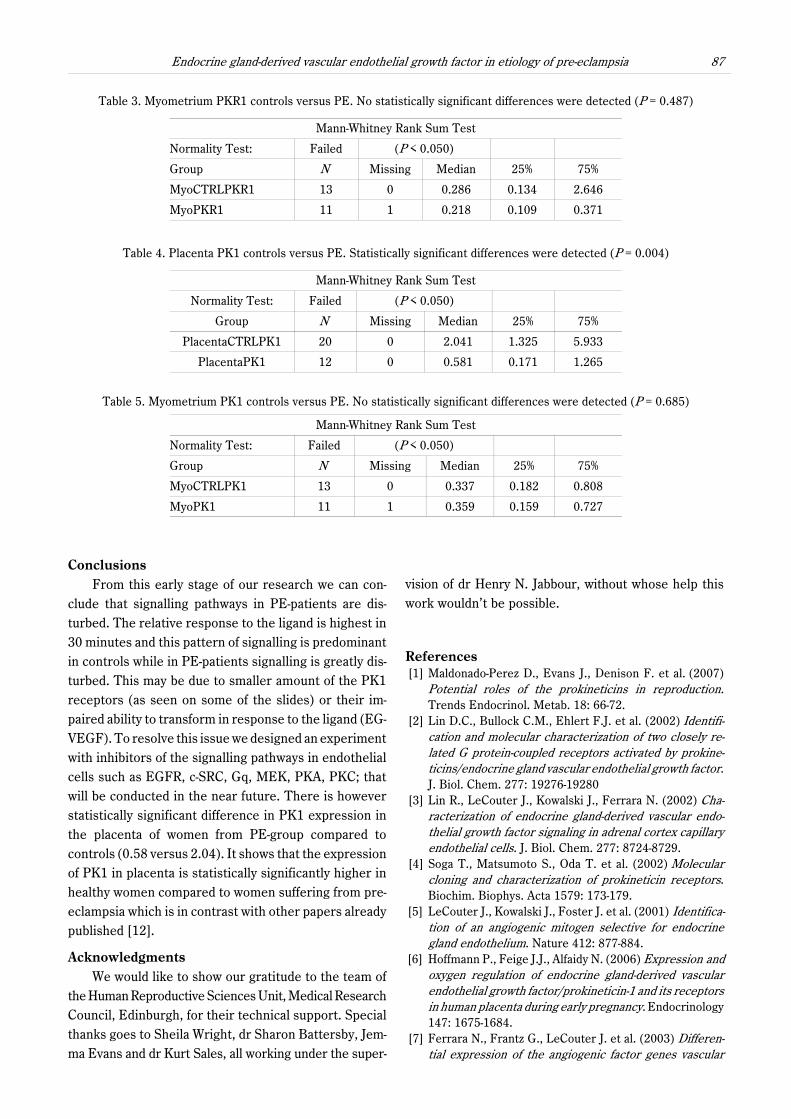
Table 4. Placenta PK1 controls versus PE. Statistically significant differences were detected (P = 0.004)
Mann-Whitney Rank Sum Test
Normality Test: Failed (P < 0.050)
Group N Missing Median 25% 75%
PlacentaCTRLPK1 20 0 2.041 1.325 5.933
PlacentaPK1 12 0 0.581 0.171 1.265
Table 5. Myometrium PK1 controls versus PE. No statistically significant differences were detected (P = 0.685)
Mann-Whitney Rank Sum Test
Normality Test: Failed (P < 0.050)
Group N Missing Median 25% 75%
MyoCTRLPK1 13 0 0.337 0.182 0.808
MyoPK1 11 1 0.359 0.159 0.727
ConclusionsFrom this early stage of our research we can con-
clude that signalling pathways in PE-patients are dis-turbed. The relative response to the ligand is highest in30 minutes and this pattern of signalling is predominantin controls while in PE-patients signalling is greatly dis-turbed. This may be due to smaller amount of the PK1receptors (as seen on some of the slides) or their im-paired ability to transform in response to the ligand (EG-VEGF). To resolve this issue we designed an experimentwith inhibitors of the signalling pathways in endothelialcells such as EGFR, c-SRC, Gq, MEK, PKA, PKC; thatwill be conducted in the near future. There is howeverstatistically significant difference in PK1 expression inthe placenta of women from PE-group compared tocontrols (0.58 versus 2.04). It shows that the expressionof PK1 in placenta is statistically significantly higher inhealthy women compared to women suffering from pre-eclampsia which is in contrast with other papers alreadypublished [12].
AcknowledgmentsWe would like to show our gratitude to the team of
the Human Reproductive Sciences Unit, Medical ResearchCouncil, Edinburgh, for their technical support. Specialthanks goes to Sheila Wright, dr Sharon Battersby, Jem-ma Evans and dr Kurt Sales, all working under the super-
vision of dr Henry N. Jabbour, without whose help thiswork wouldn’t be possible.
References [1] Maldonado-Perez D., Evans J., Denison F. et al. (2007)
Potential roles of the prokineticins in reproduction.Trends Endocrinol. Metab. 18: 66-72.
[2] Lin D.C., Bullock C.M., Ehlert F.J. et al. (2002) Identifi-cation and molecular characterization of two closely re-lated G protein-coupled receptors activated by prokine-ticins/endocrine gland vascular endothelial growth factor.J. Biol. Chem. 277: 19276-19280
[3] Lin R., LeCouter J., Kowalski J., Ferrara N. (2002) Cha-racterization of endocrine gland-derived vascular endo-thelial growth factor signaling in adrenal cortex capillaryendothelial cells. J. Biol. Chem. 277: 8724-8729.
[4] Soga T., Matsumoto S., Oda T. et al. (2002) Molecularcloning and characterization of prokineticin receptors.Biochim. Biophys. Acta 1579: 173-179.
[5] LeCouter J., Kowalski J., Foster J. et al. (2001) Identifica-tion of an angiogenic mitogen selective for endocrinegland endothelium. Nature 412: 877-884.
[6] Hoffmann P., Feige J.J., Alfaidy N. (2006) Expression andoxygen regulation of endocrine gland-derived vascularendothelial growth factor/prokineticin-1 and its receptorsin human placenta during early pregnancy. Endocrinology147: 1675-1684.
[7] Ferrara N., Frantz G., LeCouter J. et al. (2003) Differen-tial expression of the angiogenic factor genes vascular
M. Szuber, W. Markwitz, H.N. Jabbour, F. Denison, G.H. Bręborowicz88
endothelial growth factor (VEGF) and endocrine gland-derived VEGF in normal and polycystic human ovaries.Am. J. Pathol. 162: 1881-1893.
[8] Fraser H.M., Bell J., Wilson H., Taylor P.D. et al. (2005)Localization and quantification of cyclic changes in theexpression of endocrine gland vascular endothelial growthfactor in the human corpus luteum. J. Clin. Endocrinol.Metab. 90: 427-434
[9] Urayama K., Guilini C., Messaddeq N. et al. (2007) Theprokineticin receptor-1 (GPR73) promotes cardiomyocytesurvival and angiogenesis. FASEB J. 21: 2980-2993.
[10] Dorsch M., Qiu Y., Soler D. et al. (2005) PK1/EG-VEGFinduces monocyte differentiation and activation. J. Leu-koc. Biol. 78: 426-434.
[11] Hoffmann P., Feige J.J., Alfaidy N. (2007) Placental ex-pression of EG-VEGF and its receptors PKR1 (prokine-
ticin receptor-1) and PKR2 throughout mouse gestation.Placenta 28: 1049-1058.
[12] Chung J.Y., Song Y., Wang Y. et al. (2004) Differential ex-pression of vascular endothelial growth factor (VEGF),endocrine gland derived-VEGF, and VEGF receptors inhuman placentas from normal and preeclamptic pregnan-cies. J. Clin. Endocrinol. Metab. 89: 2484-2490.
J Michał SzuberDepartment of Perinatology and GynecologyUniversity of Medical Sciences in PoznańPolna 33, 60-535 Poznań, Poland