Scholars' Mine Scholars' Mine Masters Theses Student Theses and Dissertations 1968 Endogenous metabolism of Polyporus sulfureus. : Utilization of Endogenous metabolism of Polyporus sulfureus. : Utilization of exogenous carbohydrates and fatty acids exogenous carbohydrates and fatty acids Shu-Mei Wang Follow this and additional works at: https://scholarsmine.mst.edu/masters_theses Part of the Chemistry Commons Department: Department: Recommended Citation Recommended Citation Wang, Shu-Mei, "Endogenous metabolism of Polyporus sulfureus. : Utilization of exogenous carbohydrates and fatty acids" (1968). Masters Theses. 6816. https://scholarsmine.mst.edu/masters_theses/6816 This thesis is brought to you by Scholars' Mine, a service of the Missouri S&T Library and Learning Resources. This work is protected by U. S. Copyright Law. Unauthorized use including reproduction for redistribution requires the permission of the copyright holder. For more information, please contact [email protected].
Transcript
Scholars' Mine Scholars' Mine
Masters Theses Student Theses and Dissertations
1968
Endogenous metabolism of Polyporus sulfureus. : Utilization of Endogenous metabolism of Polyporus sulfureus. : Utilization of
exogenous carbohydrates and fatty acids exogenous carbohydrates and fatty acids
Shu-Mei Wang
Follow this and additional works at: https://scholarsmine.mst.edu/masters_theses
Part of the Chemistry Commons
Department: Department:
Recommended Citation Recommended Citation Wang, Shu-Mei, "Endogenous metabolism of Polyporus sulfureus. : Utilization of exogenous carbohydrates and fatty acids" (1968). Masters Theses. 6816. https://scholarsmine.mst.edu/masters_theses/6816
This thesis is brought to you by Scholars' Mine, a service of the Missouri S&T Library and Learning Resources. This work is protected by U. S. Copyright Law. Unauthorized use including reproduction for redistribution requires the permission of the copyright holder. For more information, please contact [email protected].
1. Standard Curve for Total Organic Solids • • • • • • 17
2. Standard Curve for Reducing Sugar • • • • • • • • • 20
3. Self-absorption Correction Curve for 14c in the Gas Flow Proportional Counter {Riol, Model 40-9B, 2-9 and 49-30) ~ o ~ • • • • • • • • • • • • • • • • • 24
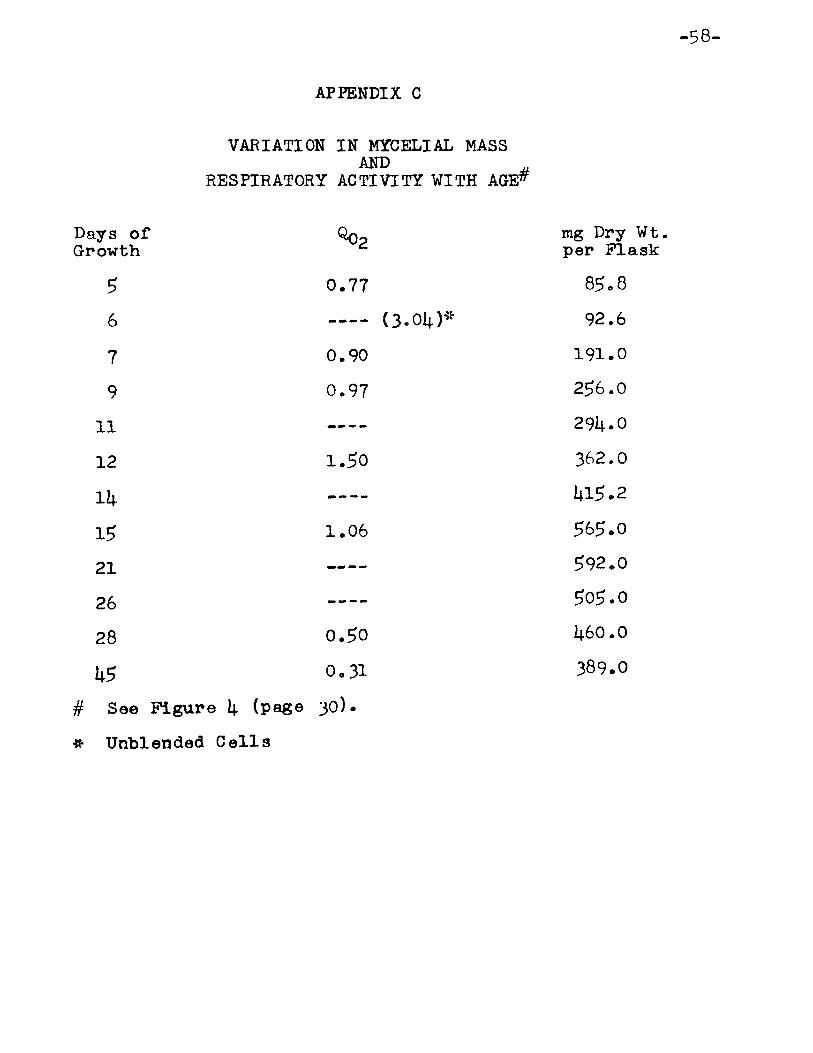
4. Variation in Mycelial Mass, Respiration or Mycelium and Total Organic Solids and Reducing SUgar in the Medium with Age of Mycelium • • • • • • • • • • • • 30
~. The Influence of pH on Glucose Oxidation by 7 Day Old Cells of ~· sulfureus • • • • • • • • • • • • • 31
6. Effect of Exogenous Substrate on the Respiration of
7.
7 Day Old !:· sulfureus • • • • • • • • • • • • • • 35
Recovery of 14c as co2 with Various 14c-labeled Exogenous Substrates • • • • • • • • • • • • • • • 39
LIST OF TABLES
Table
1.
2.
3.
4.
Compositions of Media •• • • • • • • • • • • • • •
Growth or P. sulfureus on Different Media • • • • •
Change or Medium pH with Growth of P. sulfureus on 4 '?& Malt Medium • • • • • • • • • • -. • • • • • • •
Total Organic Solids and Reducing Sugar at Various Times During the Growth of P. sulfureus on 4% Malt Medium • • • • • • • • • • -. • • • • • • • • • • •
-vii-
page
13
26
27
29
S. Respiratory Quotient forE· sulfureus with Age • • 32
6. Influence of Carbohydrates on the Respiration of P. sul fureus • • • • • • • • • • • • • • • • • • • 33
?. ~2 Values in the Presence of Organic Acids • • • • 34
8. The Utilization or Glucose-u-1 4c (50pM, 1850 CPM/pM carbon) by the Intact Cells 6r P. sulfureus 36
9. Utilization of 14c-labeled Substrates by Cells of P. sulfureus • • • • • • • • • • • • • • • • • • • 38
-1-
I. INTRODUCTION
Polyporus sulfureus has the unusual property that almost
thirty per cent of the dry weight of the mycelium grown either
in surface or submerged culture is a mixture of triterpenoid
acids, of which three fourths is eburicoic acid (Gascoigne et al
1950; Pan et al, 1961; and Fried et al, 1964).
Since there is an inverse relationship between storage
lipid and carbohydrate of cells, i.e., individual classes of
organisms have a tendency to synthesize either but not both,
it is of interest to determine the type of control exercised
by the organism for selecting the synthesis of its storage
material.
P. sulfureus is an example of an organism which stores
lipids rather than carbohydrates. A study of its metabolism
was thought to be of interest since such a study might shed
light on the control of the "shunt" between carbohydrate and
lipid metabolism in this organism.
The determination of actual metabolic controls is a long
and difficult task. Therefore one aspect of ~· sulfureus
metabolism was selected for the present investigation, namely,
the general aspects of growth of P. sulfur~ and its endogenous
and exogenous respiration.
-2-
I I • LITERATURE
Ao GENERAL SURVEY OF POLYPORUS SULFUREU::> AND EBURICOIC ACID
Po1yporus sulfureus is a fungus within the class or basidiomycetes. This filamentous rungus has been known to
produce in good yield eburicoic acid, a triterpenoid compound
with the structure below: (Gascoigne et al, 1950, 1951, 1953)
COOH
HO
Eburicoic acid was first isolated from the £ruiting
bodies of Fornes officinalis ~., by Kurono and Kariyone who
gave the melting point as 28)°C and reported an empirical
formula of c 30~8o3 for the material they isolated. Through
a series ot• studies (Gascoigne et al 1950, 1951, 1953; Lahey
and Strasse~ 1951; and Holker et al 1953), it was recognized
that this acid has an empirical formula of c31 HS0 o3
which
corresponds to the structure shown above. The single carbon
unit attached to the side chain was found to originate from
formate, one of the members of the one-carbon metabolic pool
(Dauben and Richards, 1956; Dauben, Ban and Richards, 1957;
Dauben, Fonken and Boswell, 1957).
In addition to F. o1'ficinalis Fr., eburicoic acid has - -also been isolated from the mycelium or several Polyporus
species; !• anthracophi1us, !· eucalyptorum, P. hispidus
and !• su1fureus. Poria species such as !· cocos and Lentinus
-3-
A patent (Pan et al, 1961) indicates that organisms producing
eburicoic acid have been investigated for use as a commercial
source of steroids.
Efimenko (1960) reported that eburicoic acid extracted
from ~· betulinus Karst stimulated yeast fermentation by
17-46% and also stimulated mycelial growth of fungi by 20-26%.
Recently there has been interest in this acid because of its
selective anti-a.ndrogenic activity. It selectively inhibits
the androgenic effect on ventral prostate glands, seminal
vesicles and coagulating glands but does not have the androgenic
effect on total body growth. This acid also is reported as
an antibiotic (Bates, 1967). It is one of the two anti-
fungal steroid antibiotics (another is Viridan produced by
Trichoderma iride) reported by Harvey et al (1967).
Through an extensive study on the biosynthesis of steroid
compounds (Dauben and Richards, 1956; Bloch, 1965; Richards
and Henderickson, 1964; Gascoigne et al, 1950), it has been
shown that acetate in its active form, acetyl CoA, serves as
a precursor for the biosynthesis of eburicoic acid through
squalene to lanosterol. This is the sgme mechanism as that
for the biosynthesis o1' steroids (Dauben and Richards, 19.56;
Lawrie et al, 196.5).
Another triterpenoid, 15d-hydroxyeburicoic acid also
has been found in the triterpenoid acids extracted from~·
sulfureus and because of its occurrence this new acid has
been named sulphure.nic acid. Thirty per cent o!" the dry
weight of the mycelium of this fungus grown either in surface
-4-
or submerged culture is triterpenoid acids, three fourths o1'
which was accounted for as eburicoic acid and the remainder
as sulphurenic acid (Fried et al, 1964).
The mycelium of' this fungus also has been reported to
produce volatile amines such as methylamine, dimethylamine,
n-propylamine, iso-amylamine, colamine and phenylethylamine
as a result of the decarboxylation of various amino acids
(Rainbow and Rose, 1963).
B. BIOCHEMISTRY OF THE BASIDIOMYCETE GENUS, POLYPORUS
The biochemistry of' basidiomycetes has been studied very
little to date. The following discussion contains information
about the basidiomycetes genus, Polyporus, to which the
organism dealt with in this thesis belongs. The literature
search was not exhaustive but it is felt that the information
recorded here is nearly complete in so far as it describes
the biochemical activities that have been examined and report
ed in the literature.
The intracellular enzyme, phenolase, from the mycelium
of~· sulfureus was studied by Roesch (1965). He reported
a rapid enzymic oxidation o1' tyrosine and P-cresol and a slow
attack on pyrocatechol at pH 4-6 from an extract of' the mycelium
or this fungus. This first demonstration of such an activity
in this species is or importance for the detoxication of
heartwood toxins.
Some other investigations have been made of the metabolism
of the polyporus species, but not necessarily P. sulfureus.
The enzymes of the Embden-Meyerhof and pentose phosphate
-5-
pathways in P. brumali s extracts have been investigated by
Casselton (1966). In his work, cell-free extracts were
prepared from the mycelium o1· ~· brumalis by grinding .first
with sand for five minutes followed by centrifuging at
26,000 x g for twenty minutes at a temperature between 2-6°C.
From the measurement of the oxidation or reduction of pyridine
nucleotides in the presence or absence of different substrates,
the activities of the enzymes were assayed. From this investi
gation, Casselton was able to show that the cell-.free extracts
of ~· brumalis contained all the enzymes required for the
metabolism of glucose by both the Embden-Meyerho.f pathway
and the hexose monophosphate pathway.
The respiration o.f non-proliferating cells of ~· 2a1ustris
was studied by Newcomb and Jennison (1962). The oxygen and
carbon dioxide evolution were measured by co.nventional
manometric techniques in their study. They found that the
respiratio.n of ~· palustris mycelium was sensitive to mechani
cal disintegration. The changes in respiration with cell age,
the effect of starvation on endogenous respiration and on
glucose oxidation as well as the effect o.f buffer systems and
pH on respiration were i.nvestigated. They found that maximum
endogenous and exogenous respiration occurred in 12-hour old
pellets from shake cultures, and was associated with the lag
phase and early phase of accelerated growth. They found
that there was a decrease in exogenous and endogenous %2
with starvation and also that the respiring mycelium was
relatively insensitive to changes in pH betwee.n 4 and 6.
-6-
The Q02 was maximum at pH 5.0 - 5.5, which is the range of
the optimum for the growth or most of the wood-rotting
basidiomycetes.
The carbohydrate metabolism of ~· circinatus was studied
by Avigad et al (1961). They reported that this fungus
possesses a galactose oxidase that oxidizes the carbon-6
position of galactose, yielding galactodialdose, instead of
oxidation at the carbon-1 position as with glucose oxidase.
This oxidation of a hexose at carbon-6 was thought to occur
through a glucuronate-xylulose pathway. The reaction scheme,
which uses part of the hexose monophosphate pathway, is as
follows:
D-glucose ~ D-glucuronate --+ L-gulonate ~ xyli tol ~
* For complete description or the assay methods see page 21; these data are summarized in Table 8 and Table 9 (pages 36 and 3B>. Also see Figure 7 (page 39).
-61-
Data for the Utilization of Glucose-l-14c -Assay Medium: 2 ml of cell suspension in pH 6 and 0.067 M 1 phosphate Ruffer + 0.5 ml of 0.1 M glucose-l- 4c
(5.55 X 10 · CPM/50 pM Glucose).
Dry weight of cell suspension used in Run II are 23.6 mg and 23.7 mg in Run III.
Run II Run III Time p.l 02 CPM in % recovery )11 02 CPM in % recovery (Min.) co of' radio- co of radio-
evofved acti vi tx evofved activi ti
5 7 8
10 13 11
20 27 26
30 36 41
40 42 51
so 64 64
60 74 82 0.15 76 78 0.14
70 92 90
80 102 102
90 121 118
100 133 132
110 --- 145
120 167 137 0.25 159 144 0.26
150 224 199
180 276 199 o. 36 240 273 0.49
210 355 280
240 387 246 0.44 321 367 0.66
-62-
Data for the Utilization of Acetate-l-1 4c
Assay l"edium: 2 ml of cell suspension in pH 6 and 0.067 M , phosphate gurfer + O.t) ml of 0.1 M acetate-1-1 ~c (1.11 X 10 CPM/SO pM Acetate).
Dry weight of cell suspension used i n Run IV are 2 S • 4 mg and 23.6 mg in Run v.
Run IV Run V Time p-1 02 CPM in % recovery ?1 02 CPM in % recovery
(Min. ) co! of radio- co of radio-evo ved activit:l evofved activit~
c; 4 4
10 8 13
20 20 2S
30 31 43
40 39 47
so t)O 56
60 5'5 1780 0.16 66 1965 0.18
70 67 69
80 74 76
90 86 104
100 88 105
110 100 106
120 108 4740 0.43 121 4510 0.41
1SS 13S 1S8
180 176 S400 0.48 174 5440 0.49
210 198 203
240 240 8390 0.76 232 8000 0.72
-63-
Data for the Utilization of Glucose-u-14c
Assay Medium: 2 ml of cell suspension in pH 6 and 0.067 M 1 phosphate ~uffer + o.S ml of 0.1 M glucose-u-1 4c
( S. 55 X 10 CPM/50 fM Glucose).
Dry weight of cell suspension used in Run VI are 24.1 mg and 25.5 mg in Run VII.
Time ul 0 (Min.) 1 2
10
20
30
40
so 60
70
80
90
100
110
120
150
180
210
240
2
8
11
16
36
41
46
50 60
66
75
96
124
lSl
175
Run VI CPM in
C02 evolved
324
527
900
1240
% recovery pl o2 of radioactivity
0.06
0.10
0.16
0.22
8
13
25
31
38
52
63
69
78
82
92
103
115
143
175
192
229
Run VII CPM in % recovery
C02 of radio-evoived activtti
320 0.06
644 0.12
111S 0.20
1540 0.28
-64-
Data for the Utilization of Glucose-6-14c
Assay Medium: 2 ml of cell suspension in pH 6 and 0.067 M 1 phosphate euffer + 0.5 ml of 0.1 M glucose-6- 4c (5 .55 X 10 · CPM/50 JIM Glucose).
Dry weight of' cell suspension used in Run VIII are 23.5 mg and 23.4 mg in Run IX.
Run VIII Run IX Time pl 02 CPM in % recovery '1 02 CPM in % recovery
(Min.) C02 of radio- cof of' radio-evolved activitz evo ved activity
5 12 6
10 14 14
20 20 26
30 36 29
40 47
50 57
60 40 31 0.06 70 23 0.04
70 42 75
80 46 88
90 .53 97
100 .55 105
110 .57 116
120 62 43 0.08 128 55 0.10
150 78 177
180 102 66 0.12 202 70 0.13
210 114 215
240 133 43 o.o8 242 59 0.12
Wei tht of Ba.CO ~ per Square Centimeter o!' Planchet in Determinati6n of Radioactivity in Evolved co2*
Substrate Warburg Run. Time Weight o1' Baco 3 (Hour)
G1ucose-1-14c 1 .34 .5
2 45.0
3 48.0
4 50.0
Glucose-6-14c 1 53.2
2 55.4
3 51.6
4 50.3
Glucose-u-14c 1 51.0
2 53.0
3 54 6
4 54.9
Acetate-l-14c 1 68.2
2 60.8
3 68.3
4 73.7
-65-
* For complete description o1' the method o1' determination
See pages 22-23.
APPENDIX F
DATA FOR THE PREPARATION OF SELF-ABSORPTION CORRECTION CURVEH
# For complete description ot• the method or preparation of' the curve see page 22.; these data are plotted in Figure 3 (page 24).
~~ The respective apparent specif'ic activities were plotted against the sample thickness, this curve was extrapolated to zero thickness and this value ()27.0 CPM/mg BaCO ) considered as the actual specific activity. The pe~centage o1· relative specif'ic ac ti vi ty at each sample thickness was obtained by using the value o!' the actual specif'ic activity as 100 per cent.
-67-
BIBLIOGRAPHY
Avigad, G., Asensio, c., Amaral D. and Horecker, B. L. (1961). "Ge.lactodialdose Production with an Enzyme from the Mold Po~orus circinatus", Biochem. Biophys. Res. Commun. , 474-477.
Bates, R. B. {1967}. "Terpenoid Antibiotics" in "Antibiotics, vol. II, Biosynthesis", P. 134, PP. 146-147. Ed. by Gottlieb, D.and Shaw, P. D., Springer-Verlag New York Inc.
Bloch, K. (1965). "The Biological Synthesis of Chol a sterol", Science 150, 19-28.
Bloom, B. and Stet ten, D. Jr. (195 3). "Pathways or Glucose Metabolism", J. Amer. Chem. Soc. 72., 5446.
Blumenthal, H. J. (196 3). "Endogenous Metabolism or Filamentous Fungi", Ann. N.Y. Acad. Sci. 102, 688-706. -
Blumenthal, H. J. (1965). "Carbohydrate Metabolism. I. Glycolysis n 1 n "The Fungi", Vol. I, P. 258. Ed. by Ainsworth, Ge c. and Sussman, A. s., Academic Press, N.Y.
Cassel ton, P. J. (1966). "Enzymes of the Embden-Meyerhof and Pentose Phosphate Pathways in Pol{~orus brumalis Extracts", .J. Exptl. Botany !1, 2) 579-589.
Cheldelin, V. H. (1960). "Metabolic Pathways in Microorganisms", .John Wiley and Sons, Inc. N.Y.
Cochrane, v. w. and Peck, H. D., Jr. (1953). "The Metabolisn of Species or Streptomyces. VI. Tricarboxylic Acid Cycle Reactions i.n Streptomyces coelicolor", .r. Bacteriol. §2, 37-44.
Cochrane, v. w. (1958). "Physiology of Fungi". P. 19, John Wiley and Sons, N.Y.
Da.uben, W, Go and Richards, .J. H. (1956). "The Biosynthesis of the Triterpene, Eburicoic Acid", J. Amer. Chern. Soc. 1§_, 5 329-5 336. ·
Daub en, W. G., Fonken, G. J. and Boswell, G. A. (1957). "The Biosynthetic Precursor of the Extra Carbon Atom in the Side-Chain of Steroids", J. Amer. Chern. Soc. 12• 1000-1001.
-68-
Daub en, W. G., Ban, Y. and Richards, J. H. (1957) • "The Biosynthesis ot• the Triterpene, Eburicoic Acid: The Utilization ot· Methyl-labeled Acetate", J. Amer. Chem. Soc. 12, 968-970.
deFiebre, C. w. and Knight s. G. (1953). "The Oxidation of Glucose by Penicillium chrysogenum", J. Bacteriol. 66, 17o-172.
Efimenko, 0. M. (1960). "Physiologically Active Compounds of the Wood-rot Fungus Poly.Eorus betulinus, Karst", Mikrobiologiya ~' 548-556. Chem. Abstr. 2i' 2807c.
Foster, J. W. (1949). "Chemical Activities of Fungi" Academic Press Inc., Publishers, N.Y.
Foulkes, E. c. (1955). "Cell Permeability-The Uptake o1' pyruvate by Yeast", J. Gen. Physiol. ~' 425-431. Chem. Abstr. ~' 15714b.
Fried, J., Grabowich, P., Sabo, E. F., and Cohen, A. I. (1964) "The Structure ot' Sulphurenic Acid, A New Triterpenoid from Polyporus sulfureus 11
, Tetrahedron 20 ( 10)' 2297-2312.
Gascoigne, R. M., Holker, J. s. E., Ralph, B. J., and Robertson, A. {1950). "Occurence of Eburicoic Acid", Nature 166, 652.
Gascoigne, R. M., Holker, J. S. E., Ralph, B. J., and Robertson, A. (1951). "The Chemistry of Fungi. Part XVI. Eburicoic Acid", J. Chem. Soc., 2.346-2352.
Gascoigne, R. M., Robertson, A., and Simes, J. J. H. (1953). "The Chemistry ot' Fungi. Part XVII. Dehydroeburic oi c Ac i d 11
, J. C hem. ::> oc • 18 30 -18 3 7 •
Halliwell, G. (1960). "A Micro-determination of Carbohydrates and Proteins", Biochem. J. ]±, 457-462.
Harvey, C. L., Sihr, c. J., and Knight, s. G. (1967). 11Fucidin 1 in "Antibiotics, Vol. I. Mechanism of Action", P. 404. Ed. by Gottlieb, D. and Shaw, P. D., Springer-Verlag New York Inc.
Hodge, J. E. and Hofreiter, B. T. (1962). "The Determination o1' Reducing Sugars and Carbohydrates", in "Methods in Carbohydrate Chemistry", Vol. I. PP. 385-387. Ed. by Whistler, R. L. and Wolfrom, M. L., Academic Press, New York.
-69-
Holker, J. s. E., Powell, A. D. G., Robertson, A., Simes, J. J. H., and 'Yright R. s. (1953). "The Chemistry of Fungi. Part XVIII. The cyclic system or Eburicoic Acid", J. Chem. Soc., 2414-2422.
Holker, J. S. E., Powell, A. D. G., Robertson, A., Simes, J. J. H., Wright, R. s., and (in part) Gascoigne, R. M. (1953). "The Chemistry of Fungi. Part XIX. The Structure of Eburicoic Acid", J. Cham. Soc., 2422-2429.
Kuo, M. H. and Blumenthal, H. J. (1961 ). "Purirication and Properties of an Acid Phosphomonoesterase From Neurospora crassa.", Biochim. Biophys. Acta 2_, 13-29.
Lahey, F. N. and Strasser, P. H. A. {1951). "Eburicoic Acidu, J. Cham. Soc., 873.
Lawrie, W.f Mclean, J., Pauson, P. L., and Watson, J. (1965). 'Biosynthesis of Eburicoic Acid", Cham. Commun.,
623-624.
Newcomb, H. R. and Jennison, M. W. (1962). 'Physiology of Wood-rotting Basidiomycetes, IV. Respiration or non-proliferating cells of' Polyporus palustris", Can. J. Microbiol. ~, 145-156o
Pan, s. C., Metuchen and Lerner, L. J. (1961). "Process for the Production or Eburicoic Acid", U. S. Patent 3,010,878; Patented Nov. 28, 1961.
Pan, s. c., Laskin, A. I. and Principe, P. (1962). "Paper Chronmatography or Tri terpenoic and steroid acids", J. Chromatogr. ~' 32-36.
Rainbow, c. and Rose, A. H. {196)). "Biochemistry of Industrial Microorganisms", Academic Press, N.Y.
Ralph, Bo J. and V. J. Bender (1965). "Isolation of Two New Polysaccharides from the Cell Wall of polyporus tumulosus", Cham. Ind. (London), 1181.
Ramachandran, s. and Gottlieb, D. {1963). "Pathways of Glucose Catabolism in Caldariom~ces furnago (Ill.) 11
,
Biochim. Biophys. Acta 69, 74-8 • -Rauen, H. M. (1965). "Biochemisches Tashenbuch", Springer
Verlag, o. H. G., Berlin, Germany.
Reese, E. T. and Mandel s, M. (1962). "Enzr.nic JY.drolysi s of Cellulose and Its Derivatives', in 'Methods in Carbohydrate Chemistry", Vol. III. PP. 139-150. Ed. by Whistler, R. L. and Wolfrom M. L., Academic Press, N.Y.
-70-
Richards, J. H. and Henderickson, J. B. (1964). "The Biosynthesis of Steroids, Terpanes, and Acetogenins", W. A. Benjamin, Inc., N.Y.
Robbins, W. J. (1950). "A Surverr o:f the Growth Requirements of Some Basidiomycetes', Mycologia 42, 470-476.
Robbins,
Roesch,
W. J. and Hervey, A. (1958). "Wood, Tomato and Malt extracts and Growth o:r Some Basidiomycetes", Mycologia 2Q, 745-752.
R. (1965). nintracellular Phenolasses of the Sul.fur pore Fungus Pol~porus suli'ureus", Naturwissenschaften ~' 3~ -351. Cham. A6str. ~' 120Jlc.
Umbreit, w. w., Burris, R. H., and Stauffer (1964). "Manometric Techniques", 4th Ed. Burgess Publishing Company, Minneapolis, Minnesota.
Wang, c. H. and Willis, D. L. (1965). "Radiotra.cer Methodology in Biological Science", PrenticeHall, Inc., Englewood Cli:ffs, N.J.
ACKNOWLEDGEMENT:::>
The author wishes to express her sincere appreciation
to her advisor, Dr. Donald J. Siehr, ror his guidance and
encouragement during this research project.
The author also wishes to thank the Chemistry Department
or the University or Missouri at Rolla ror providing racili
ties, equipments and financial assistance which made this
thesis possible.
The loving help and encouragement or her parents and her
husband made her graduate study possible and it is to them
that this thesis is at•t•ectionately dedicated.
VITA
The author was born on March 21, 1942, in Taichung,
Taiwan, Republic of' China. She received her high school
education at the Provincial Taichung Girls Middle School
and graduated in June 1959.
-72-
She received a Bachelor of Science degree in Chemistry
from the Chung-Hsing University in Taichung, Taiwan, in
June, 1964.
She entered the Graduate ~chool at The University of
Missouri at Rolla in ~eptember, 1965. She served as a
research assistant in the Chemistry Department for 1966-1967