Energy advantages of orientation to solar radiation in three African ruminants Robyn S. Hetem a,n , W. Maartin Strauss a,b , Bert G. Heusinkveld c , Steven de Bie d , Herbert H.T. Prins d , Sipke E. van Wieren d a Brain Function Research Group, School of Physiology, Medical School, University of the Witwatersrand, 7 York Road, Parktown 2193, South Africa b Department of Environmental Science, University of South Africa, Private Bag X6, Florida 1710, South Africa c Department of Meteorology and Air Quality, Wageningen University, Droevendaalsesteeg 3a, 6708 PB, Wageningen, The Netherlands d Resource Ecology Group, Wageningen University, Droevendaalsesteeg 3a, 6708 PB, Wageningen, The Netherlands article info Article history: Received 25 February 2011 Accepted 28 July 2011 Available online 3 August 2011 Keywords: Solar radiation Cylindrical model Endotherm Behaviour abstract Animal orientation relative to incident solar radiation allows an animal to effectively adjust the amount of radiant heat gained from an environment. Yet recent literature found ruminants to primarily orientate north/south and proposed magnetic alignment as the most parsimonious explanation. To test whether such northerly orientation has an energy advantage, we used heated cylindrical models to estimate energy costs of thermoregulation associated with north and east orientations of three species of African ruminants under cool winter conditions. Concurrent behavioural observations revealed that eland, blue wildebeest and impala did not preferentially orientate north/south during warm summer or cool winter conditions. Instead, all three species preferred to orientate perpendicular to incident solar radiation during winter and parallel to incident solar radiation during summer, throughout the day. On clear winter days with little wind, more than 60% of animal orientation preference could be accounted for by the energy savings associated with that orientation. Thus energy demands are likely to be the primary driver of animal orientation preferences. & 2011 Elsevier Ltd. All rights reserved. 1. Introduction Behavioural thermoregulation is an animal’s primary defence against changes in the thermal environment. The advantages of behavioural thermoregulation were hypothesised nearly 50 years ago (Bartholomew, 1964) and have been well-documented in many ectotherms, which rely primarily on behaviour to regulate body temperature (for review see Stevenson, 1985). Orientation relative to incident solar radiation is one such thermoregulatory behaviour, which allows an animal to effectively adjust the amount of radiant heat gained from an environment. In cold environments, animals can facilitate heat uptake by presenting the long axis of their bodies perpendicular to the incident solar radiation. Conversely, in hot environments, animals can reduce the surface area exposed to solar radiation by orientating the long axis of their bodies parallel to the sun’s rays, thereby minimising the heat load. For example, spiders (Humphreys, 1991; Suter, 1981), butterflies (Ohsaki, 1986; Shelly and Ludwig, 1985), dragonflies (May, 1976; Tracy et al., 1979), wasps (Coelho, 2001), flies (O’Neill et al., 1990), lizards (Muth, 1977; Waldschmidt, 1980), snakes (Lillywhite, 1980) and tortoises (Coe, 2004) all adjust their orientation relative to incident solar radiation to facilitate or minimise radiant heat gain in order to maintain a body temperature within a preferred or optimal range. The contribution of solar radiation to the energy budget of endothermic animals is more difficult to predict because the animal’s heat production and evaporative water loss allow for the maintenance of body temperatures independent of radiant heat loads (Walsberg, 1992). Nevertheless, orientation to solar radiation has been proposed to reduce energy costs associated with the endothermy in birds (Fortin et al., 2000; Lustick, 1969; Walsberg, 1982) and small mammals (Brown and Downs, 2007; Chappell and Bartholomew, 1981). Comparatively few studies have quantified the effects of orientation to solar radiation in large mammals. Early studies on cattle (Riemerschmid, 1943) and sheep (Clapperton et al., 1965) showed that these ruminants can reduce heat gain by orientating parallel to the sun’s rays thereby reducing the surface area presented to incident solar radiation. Since then a handful of studies have computed energy savings of orientation to solar radiation in ruminants, namely, cattle (Keren and Olson, 2007; Olson and Wallander, 2001), black wildebeest (Connochaetes gnou; Maloney et al., 2005) and springbok (Antidorcas marsupialis; Hetem et al., 2009; Hofmeyr and Louw, 1987). Although orientation to solar radiation plays an important role in thermoregulation of ruminants, it is not the only factor which may influence an animal’s orientation. Begall et al. (2008) Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/jtherbio Journal of Thermal Biology 0306-4565/$ - see front matter & 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.jtherbio.2011.07.012 n Corresponding author. Tel.: þ2711 717 2163; fax: þ2711 643 2765. E-mail addresses: [email protected], [email protected] (R.S. Hetem). Journal of Thermal Biology 36 (2011) 452–460
Energy advantages of orientation to solar radiation in threeAfrican ruminants
Robyn S. Hetem a,n, W. Maartin Strauss a,b, Bert G. Heusinkveld c, Steven de Bie d,Herbert H.T. Prins d, Sipke E. van Wieren d
a Brain Function Research Group, School of Physiology, Medical School, University of the Witwatersrand, 7 York Road, Parktown 2193, South Africab Department of Environmental Science, University of South Africa, Private Bag X6, Florida 1710, South Africac Department of Meteorology and Air Quality, Wageningen University, Droevendaalsesteeg 3a, 6708 PB, Wageningen, The Netherlandsd Resource Ecology Group, Wageningen University, Droevendaalsesteeg 3a, 6708 PB, Wageningen, The Netherlands
a r t i c l e i n f o
Article history:
Received 25 February 2011
Accepted 28 July 2011Available online 3 August 2011
Animal orientation relative to incident solar radiation allows an animal to effectively adjust the amount
of radiant heat gained from an environment. Yet recent literature found ruminants to primarily
orientate north/south and proposed magnetic alignment as the most parsimonious explanation. To test
whether such northerly orientation has an energy advantage, we used heated cylindrical models to
estimate energy costs of thermoregulation associated with north and east orientations of three species
of African ruminants under cool winter conditions. Concurrent behavioural observations revealed that
eland, blue wildebeest and impala did not preferentially orientate north/south during warm summer or
cool winter conditions. Instead, all three species preferred to orientate perpendicular to incident solar
radiation during winter and parallel to incident solar radiation during summer, throughout the day. On
clear winter days with little wind, more than 60% of animal orientation preference could be accounted
for by the energy savings associated with that orientation. Thus energy demands are likely to be the
primary driver of animal orientation preferences.
& 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Behavioural thermoregulation is an animal’s primary defenceagainst changes in the thermal environment. The advantages ofbehavioural thermoregulation were hypothesised nearly 50 yearsago (Bartholomew, 1964) and have been well-documented in manyectotherms, which rely primarily on behaviour to regulate bodytemperature (for review see Stevenson, 1985). Orientation relativeto incident solar radiation is one such thermoregulatory behaviour,which allows an animal to effectively adjust the amount of radiantheat gained from an environment. In cold environments, animalscan facilitate heat uptake by presenting the long axis of their bodiesperpendicular to the incident solar radiation. Conversely, in hotenvironments, animals can reduce the surface area exposed to solarradiation by orientating the long axis of their bodies parallel to thesun’s rays, thereby minimising the heat load. For example, spiders(Humphreys, 1991; Suter, 1981), butterflies (Ohsaki, 1986; Shellyand Ludwig, 1985), dragonflies (May, 1976; Tracy et al., 1979),wasps (Coelho, 2001), flies (O’Neill et al., 1990), lizards (Muth,1977; Waldschmidt, 1980), snakes (Lillywhite, 1980) and tortoises
ll rights reserved.
: þ2711 643 2765.
(Coe, 2004) all adjust their orientation relative to incident solarradiation to facilitate or minimise radiant heat gain in order tomaintain a body temperature within a preferred or optimal range.
The contribution of solar radiation to the energy budget ofendothermic animals is more difficult to predict because theanimal’s heat production and evaporative water loss allow forthe maintenance of body temperatures independent of radiantheat loads (Walsberg, 1992). Nevertheless, orientation to solarradiation has been proposed to reduce energy costs associated withthe endothermy in birds (Fortin et al., 2000; Lustick, 1969;Walsberg, 1982) and small mammals (Brown and Downs, 2007;Chappell and Bartholomew, 1981). Comparatively few studies havequantified the effects of orientation to solar radiation in largemammals. Early studies on cattle (Riemerschmid, 1943) and sheep(Clapperton et al., 1965) showed that these ruminants can reduceheat gain by orientating parallel to the sun’s rays thereby reducingthe surface area presented to incident solar radiation. Since then ahandful of studies have computed energy savings of orientation tosolar radiation in ruminants, namely, cattle (Keren and Olson,2007; Olson and Wallander, 2001), black wildebeest (Connochaetes
gnou; Maloney et al., 2005) and springbok (Antidorcas marsupialis;Hetem et al., 2009; Hofmeyr and Louw, 1987).
Although orientation to solar radiation plays an importantrole in thermoregulation of ruminants, it is not the only factorwhich may influence an animal’s orientation. Begall et al. (2008)
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460 453
recently suggested that cattle, red deer (Cervus elaphus), and roedeer (Capreolus capreolus) preferentially orientate north/southand proposed magnetic alignment as the most parsimoniousexplanation. A subsequent paper suggested that even low-fre-quency magnetic fields generated by high-voltage power linescould disrupt such magnetic alignment (Burda et al., 2009). Wetherefore investigated whether three different-sized species ofAfrican ruminants were preferentially selecting a north/southorientation, under both hot summer and cool winter conditions.Secondly, we used heated cylindrical models to estimate theenergy costs of homeothermy when the model was orientatednorth and east to test whether there was an energy advantage tonortherly orientation under cool winter conditions when thermo-regulatory theory might predict otherwise.
2. Material and methods
2.1. Sensor construction
Fig. 1 shows the model design. Models were constructed fromtwo different sized galvanised iron cylinders (1 mm wall thickness,Ferrocal b.v. Lucht- en milieutechniek, Hazenberg, the Netherlands)to represent both large (315 mm diameter, 780 mm length) andmedium (250 mm diameter, 620 mm length) sized ruminants. Toprevent heat leakage, both ends of the cylinder were equippedwith lids, made of the same material, which were pressed into thecylinder and sealed with insulation tape. Each model was mountedon a wooden pole 1 m above the ground.
Silver coated copper wire with PTFE ribbon insulation(HP26SBA.16100 Belden cable, Farnell, Maarssen, the Netherlands)
+
-
H
L
HL
GNC1
GND
GN12V
Data logger
Power supply 12V
Power supply 48V
DIV 10:1
insulation
Power to fans
Thermocouples
220V
220V
Fig. 1. Schematic representation of the heated model which we used to estimate the
(1 mm wall thickness). The large cylinder (315 mm diameter, 780 mm length) represen
620 mm length) represented the impala.
was used for the heating coil and was glued with aluminium tapeto the inner surface of the cylinders. The wire for the large andsmall models was doubled up and looped regularly at �15 mmintervals. A copper frame was placed inside the model to supporttwo fans, which ensured uniform heat distribution within thecylinders. Fans (VAPO bearing axial fan, 50�50 mm with air flow184 m3 h�1, KDE1205PFV2.11.MS.A.GN, RS Components, Haarlem,the Netherlands) were mounted next to each other in the centre ofthe copper frame and blew in opposite directions.
Four thermocouples (type K, alumel/chromel, Tex/Sitw-24f-KK,Thermoshop, the Netherlands) were insulated and glued to thewall of the cylinder, two per side, with insulating aluminium tape.The surface temperature of the cylinder was calculated as thearithmetic average of the four measured surface temperaturevalues and was maintained at 37 1C. A power supply (400 W,48 V, SMQ400PS48-C, Farnell, Maarssen, the Netherlands) ensuredthe models could maintain their temperature at 37 1C whenexposed to ambient temperatures below 5 1C. Once the modelreached 37 1C, a shunt (3 A, 75 mV ammeter plate, SHR3A75, RSComponents, Haarlem, the Netherlands) and solid state relay (10 A,60 V, CMX60D10, Farnell, Maarssen, the Netherlands) rerouted thecurrent to prevent further heating.
Measurements of model surface temperatures, current andvoltage were recorded on a data logger (CR1000, CampbellScientific Inc., Loughborough, UK). Data were recorded everysecond and stored as an average every minute.
2.2. Testing and calibrating
Model design was tested in a climatic chamber to optimisethermal homogeneity and ensure an adequate power supply
1234
Shunt
Solid state relay
- - ++
Heating wires
Galvanized iron cylinder
energy cost of thermoregulation. We used two sizes of galvanised iron cylinders
ted the eland and blue wildebeest, whereas the small cylinder (250 mm diameter,
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460454
under cold ambient temperatures. Models were constructed to beidentical, so that the amount of power required to maintain thedifferent models at 37 1C was comparable under different envir-onmental conditions. Such assumptions were confirmed in aclimatic chamber over a temperature range of 5–35 1C. Allthermocouples were compared individually, in an insulated waterbath, against a high-accuracy thermometer (Quat 100, Heraeus,Hanau, Germany) and were accurate to �0.3 1C. We do not reportstandard operative temperature since accurate calculations ofstandard operative temperature require measurement of meta-bolic heat production and evaporative heat loss for calibration(for review see Dzialowski, 2005) and such measurements wereunfeasible for our large ruminants in the field. Instead we simplycalculated the energy required to maintain the cylinders at 37 1Cunder cool winter conditions, since the model was not designedto operate under hot summer conditions.
2.3. Field experiment
In order to measure the energy cost of thermoregulation undercool winter conditions, where our study species naturally occur,we set up the models in Asante Sana Private Game Reserve(321150S, 25100E), in the Eastern Cape, South Africa. The field siteis a 10,697 ha reserve with long cool winters. The vegetation, atlow altitudes (ca. 1000 m) in the valley floor, where eland(Tragelaphus oryx) and impala (Aepyceros melampus) wereobserved, is classified as Southern Karoo Riviere vegetation type(Mucina and Rutherford, 2006) and is dominated by Acacia karroo
trees and shrubs. The vegetation at higher altitudes of theSneeuberg (1200–1600 m above sea level), where herds of bluewildebeest (Connochaetes taurinus) were observed, is denser andmay best be classified as ‘‘temperate thicket’’ or ‘‘incipient forest’’(Clark et al., 2009).
Two pelt samples of each species, namely eland and bluewildebeest, were mounted on the large-sized models. On aseparate occasion, two impala pelts were mounted on themedium-sized models. The pelts were sliced and tanned accord-ing to international standards (Woods Tanning, Uitenhage, SouthAfrica). After processing, pelt thickness averaged 1.270.3 mm forthe eland, 1.670.2 mm for the blue wildebeest and 1.370.1 mmfor the impala pelts. Vaseline petroleum jelly was used to ensuregood contact between the model and the pelt.
For comparison of the energy consequences of orientation,models were placed in the open, with one model positioned toface north and another model positioned to face east. After 24 h ofdata collection the models were rotated by 901, so that the modelwhich faced north faced east and vice versa. Hence, two days ofdata were collected from the models covered in pelts from each ofthe experimental species. Behavioural observations of the experi-mental species in winter coincided with model data collection.
2.4. Behavioural observations
We made behavioural observation of animal orientation for aminimum of two days per species in August 2009, austral winter,and December 2010, austral summer. Two of us (RH andWMS) made all the observations, using binoculars, either on footor from a vehicle within o1 km of the animals. Scan samples(Altmann, 1974) of behaviour were made at 5 min intervals,throughout �11 h observation periods from sunrise to sunset.An instantaneous sampling method (Altmann, 1974) was used torecord the behaviour of each individual at each scan. Individualsfrom up to four herds were observed simultaneously. Werecorded the direction each animal was facing in relation tocompass orientation, based on eight 451 arcs relative to north,their orientation to solar radiation (long axis of body parallel,
perpendicular or oblique to the sun), their orientation to winddirection (long axis of body parallel, perpendicular or oblique tothe wind), whether animals were in sunshine or shade andstanding or lying. The herds under observation consisted ofbetween 5 and 50 individuals. The median number of adultsunder observation at each scan was 18 for eland, seven for bluewildebeest and nine for impala.
2.5. Meteorological data measurements
We collected climatic data from a portable weather station(Hobo Weather Station, Onset Computer Corporation, Pocasset,MA, USA) located within 100 m of the models. We monitoredwind speed (m s�1), solar radiation (W m�2), dry-bulb tempera-ture (1C), relative humidity (%) and standard (150 mm diameter)black globe temperature (1C) for the duration of the study.
2.6. Data analysis
The number of individuals orientated in each compass arc wassummed for each hour. The proportion of individuals orientatedin each arc was then calculated as the total number of individualsin each arc divided by the total number of animals observed inthat hour. Similar procedures were used to calculate the propor-tion of animals orientated parallel, perpendicular or oblique toincident solar radiation and wind direction. Orientation to windwas incorporated into analysis only when wind speeds exceeded1 m s�1. Since the expected random values for the obliqueorientation relative to the incident solar radiation and winddirection are twice that expected for parallel and perpendicularorientations, observed values for oblique orientation were halvedfor simpler analysis. Within each species, average daily orienta-tion relative to compass direction, incident solar radiation andwind direction was compared using a one-way analysis ofvariance (ANOVA). Chi-squared analysis of original frequencydata was used to test for association between each behaviouralcategory and the different species.
We compared power output, per unit surface area of themodel, for both the model facing north and the model facing eastper day in winter. Assuming that our ruminants had roughly thesame shape, we scaled the power output to account for thedifference in size between our study species. Although the radiantheat transfer coefficient does not change with body size, theconvective heat transfer is inversely proportional to approxi-mately the square root of the effective diameter of the animal(Mitchell et al., 1969). Hence the power output was convertedbased on the equation
Panimal ¼ Pcylinder qmanimal
mcylinder
� �0:67
þð1�qÞmanimal
mcylinder
� �0:27" #
where P is the power output (J s�1) of the cylinder/animal, m isthe body mass (kg) of the cylinder/animal and q is the proportionof heat generated within the cylinder/animal that is lost to theenvironment by the cylinder/animal via radiant heat transfer. Themass of the model was estimated based on its volume by theequation V¼m/r, where r¼density (0.8 kg L�1); q depends onthe ambient conditions to which the animal is exposed anddecreases with increasing radiation and wind speed. Webracketed our power scaling within q¼0 (minimum) and q¼0.7(maximum), as estimated from human models in still environ-ments with no radiation (Fanger, 1970).
We calculated the energy costs of maintaining homeothermyper hour for the models facing both north and east. As aconservative estimate of energy savings associated with orienta-tion, we correlated, per day, the minimum energy cost of
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460 455
orientating north, as represented by the energy differencebetween the north-facing and east-facing model, per hour, aftercorrection for animal mass, against the ratio of animals orientatednorth/south versus east/west within that hour. By summing theabsolute energy difference between the north-facing and east-facing models over 11 h of sunlight, we could estimate energysaving of orientating north and east at different times of dayduring winter. The energy savings were then converted to apercentage of the Basal Metabolic Rate (BMR) of each species asrepresented by the equation BMR¼293m0.75 where BMR is in kJper day and m is the body mass (kg; Kleiber, 1961).
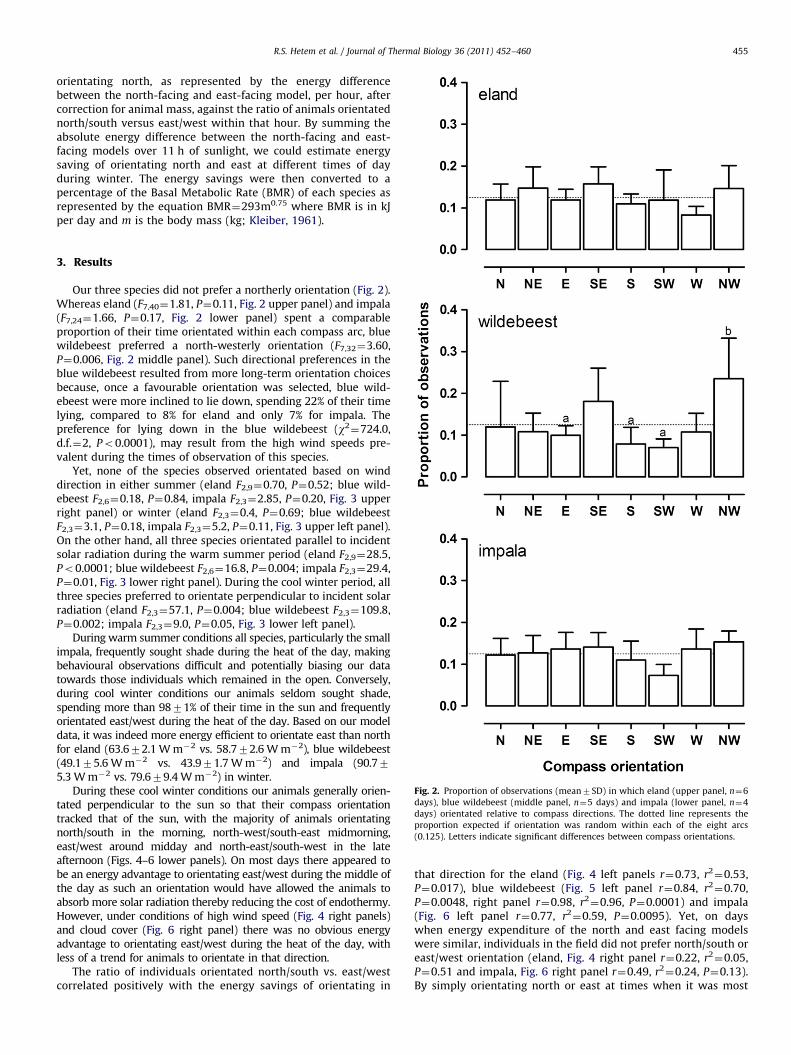
Fig. 2. Proportion of observations (mean7SD) in which eland (upper panel, n¼6
days), blue wildebeest (middle panel, n¼5 days) and impala (lower panel, n¼4
days) orientated relative to compass directions. The dotted line represents the
proportion expected if orientation was random within each of the eight arcs
(0.125). Letters indicate significant differences between compass orientations.
3. Results
Our three species did not prefer a northerly orientation (Fig. 2).Whereas eland (F7,40¼1.81, P¼0.11, Fig. 2 upper panel) and impala(F7,24¼1.66, P¼0.17, Fig. 2 lower panel) spent a comparableproportion of their time orientated within each compass arc, bluewildebeest preferred a north-westerly orientation (F7,32¼3.60,P¼0.006, Fig. 2 middle panel). Such directional preferences in theblue wildebeest resulted from more long-term orientation choicesbecause, once a favourable orientation was selected, blue wild-ebeest were more inclined to lie down, spending 22% of their timelying, compared to 8% for eland and only 7% for impala. Thepreference for lying down in the blue wildebeest (w2
¼724.0,d.f.¼2, Po0.0001), may result from the high wind speeds pre-valent during the times of observation of this species.
Yet, none of the species observed orientated based on winddirection in either summer (eland F2,9¼0.70, P¼0.52; blue wild-ebeest F2,6¼0.18, P¼0.84, impala F2,3¼2.85, P¼0.20, Fig. 3 upperright panel) or winter (eland F2,3¼0.4, P¼0.69; blue wildebeestF2,3¼3.1, P¼0.18, impala F2,3¼5.2, P¼0.11, Fig. 3 upper left panel).On the other hand, all three species orientated parallel to incidentsolar radiation during the warm summer period (eland F2,9¼28.5,Po0.0001; blue wildebeest F2,6¼16.8, P¼0.004; impala F2,3¼29.4,P¼0.01, Fig. 3 lower right panel). During the cool winter period, allthree species preferred to orientate perpendicular to incident solarradiation (eland F2,3¼57.1, P¼0.004; blue wildebeest F2,3¼109.8,P¼0.002; impala F2,3¼9.0, P¼0.05, Fig. 3 lower left panel).
During warm summer conditions all species, particularly the smallimpala, frequently sought shade during the heat of the day, makingbehavioural observations difficult and potentially biasing our datatowards those individuals which remained in the open. Conversely,during cool winter conditions our animals seldom sought shade,spending more than 9871% of their time in the sun and frequentlyorientated east/west during the heat of the day. Based on our modeldata, it was indeed more energy efficient to orientate east than northfor eland (63.672.1 W m�2 vs. 58.772.6 W m�2), blue wildebeest(49.175.6 W m�2 vs. 43.971.7 W m�2) and impala (90.775.3 W m�2 vs. 79.679.4 W m�2) in winter.
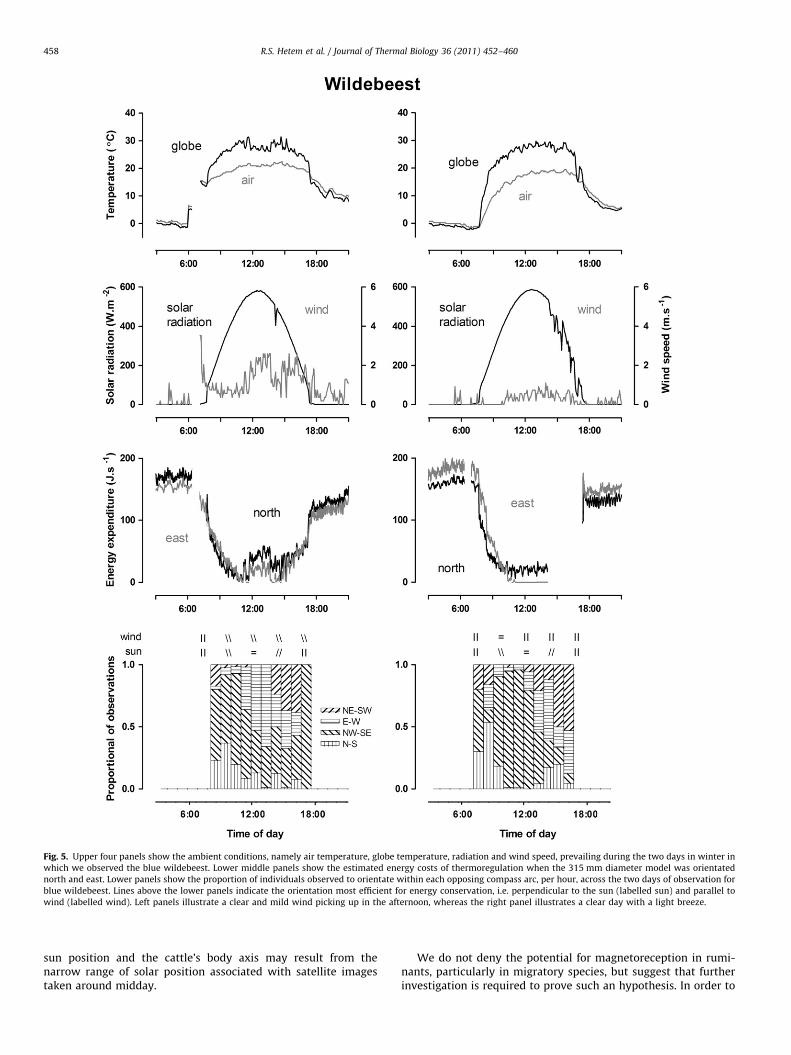
During these cool winter conditions our animals generally orien-tated perpendicular to the sun so that their compass orientationtracked that of the sun, with the majority of animals orientatingnorth/south in the morning, north-west/south-east midmorning,east/west around midday and north-east/south-west in the lateafternoon (Figs. 4–6 lower panels). On most days there appeared tobe an energy advantage to orientating east/west during the middle ofthe day as such an orientation would have allowed the animals toabsorb more solar radiation thereby reducing the cost of endothermy.However, under conditions of high wind speed (Fig. 4 right panels)and cloud cover (Fig. 6 right panel) there was no obvious energyadvantage to orientating east/west during the heat of the day, withless of a trend for animals to orientate in that direction.
The ratio of individuals orientated north/south vs. east/westcorrelated positively with the energy savings of orientating in
that direction for the eland (Fig. 4 left panels r¼0.73, r2¼0.53,
P¼0.017), blue wildebeest (Fig. 5 left panel r¼0.84, r2¼0.70,
P¼0.0048, right panel r¼0.98, r2¼0.96, P¼0.0001) and impala
(Fig. 6 left panel r¼0.77, r2¼0.59, P¼0.0095). Yet, on days
when energy expenditure of the north and east facing modelswere similar, individuals in the field did not prefer north/south oreast/west orientation (eland, Fig. 4 right panel r¼0.22, r2
¼0.05,P¼0.51 and impala, Fig. 6 right panel r¼0.49, r2
¼0.24, P¼0.13).By simply orientating north or east at times when it was most
Fig. 3. Proportion of observations (mean7SD) in which eland (open bars), blue wildebeest (hatched bars) and impala (black bars) orientated parallel, perpendicular and
oblique to the wind direction (upper panels) and incident solar radiation (lower panels) in winter (left panels) and summer (right panels). The dotted line represents the
proportion expected if orientation was random (0.25); significant differences between orientations indicated by *Po0.05, **Po0.01, ***Po0.001.
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460456
energetically efficient to do so, individuals could substantiallyreduce the energy cost of thermoregulation, with the eland saving1.0–3.4 MJ day�1 (equivalent to �11% BMR), the blue wildebeest0.7–1.4 MJ day�1 (equivalent to �9% BMR) and the impala0.2–0.7 MJ day�1 (equivalent to �16% BMR).
4. Discussion
We showed no preference for north/south orientation in threespecies of African ruminants exposed to both warm summer andcool winter conditions in South Africa (Fig. 2). Instead, we showeda preference for perpendicular orientation to incident solar radia-tion during winter and a preference for parallel orientation toincident solar radiation in summer, throughout the day, in all threespecies (Fig. 3, lower panels). Perpendicular orientation to incidentsolar radiation would have increased the surface area exposed tothe sun’s rays, thereby facilitating the absorption of radiant heatunder cool sunny conditions. On clear days with little wind, morethan 60% of animal orientation preference could be accounted forby the energy savings associated with that orientation. Easterlyorientation to incident solar radiation during the heat of the daysignificantly reduced metabolic costs, particularly for the smallimpala, which was able to conserve 16% of its basal metabolic coststhrough appropriate orientation. It is clear from these results thatanimal orientation in our study was primarily driven by animalsattempting to reduce the energy demands of endothermy.
Although our sample size in terms of the number of days ofreplicates was small, it was sufficient to show significant andinteresting differences. Furthermore, we acknowledge there are
limitations associated with the use of heated models, but believeour heated cylinders provided a reasonable estimate of theamount of energy required to maintain homeothermy under thecool ambient conditions experienced by our animals in winter.Our estimate of energy savings associated with perpendicularorientation to incident solar radiation for the impala was similarto that previously calculated for springbok (0.2 MJ day�1; Hetemet al., 2009). However, our estimate of energy savings for bluewildebeest was slightly less than that calculated for black wild-ebeest exposed to cool winter conditions (Maloney et al., 2005).Their estimate of energy saving (7 MJ day�1) may more accu-rately reflect total energy savings associated with continualadjustments in orientation so that the animals remained perpen-dicular to incident solar radiation throughout the day, whereasour models simply accounted for energy savings associated withnortherly and easterly orientation throughout the day.
The energy advantage of orientating perpendicular to incidentsolar radiation would be greatest during cool conditions with highlevels of solar radiation, such as those experienced during winterin our study. Similarly, cattle orientate perpendicular to solarradiation during cool sunny days (Gonyou and Stricklin, 1981;Kennedy and Day, 1989; Olson and Wallander, 2001) because ofthe associated energy advantages of such thermoregulatorybehaviour (Keren and Olson, 2006a, 2006b, 2007).
However, under conditions of heat stress during warm sum-mer months, our three species, and other ruminants (Berry et al.,1984; Hofmeyr and Louw, 1987; Maloney et al., 2005; Vrahimisand Kok, 1992), orientated parallel to incident solar radiation inan attempt to reduce radiant heat gain. During the heat of the day,when such parallel orientation would be most advantageous,
Fig. 4. Upper four panels show the ambient conditions, namely air temperature, globe temperature, radiation and wind speed, prevailing during the two days in winter in
which we observed the eland. Lower middle panels show the estimated energy costs of thermoregulation when the 315 mm diameter model was orientated north and
east. Lower panels show the proportion of individuals observed to orientate within each opposing compass arc, per hour, across the two days of observation for eland. Lines
above the lower panels indicate the orientation most efficient for energy conservation, i.e. perpendicular to the sun (labelled sun) and parallel to wind (labelled wind) at
different times throughout the day. Left panels illustrate a clear day with a strong northerly wind, whereas the right panels illustrates a clear day with gusty winds that
frequently changed direction.
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460 457
a north/south orientation minimised the surface area exposedto incident solar radiation. It was under these conditions,around midday on cloudless summer days, that Begall et al.
(2008) showed a preference for north/south orientation incattle, based on satellite images from 14 countries, includingSouth Africa. That these authors found no correlation between
Fig. 5. Upper four panels show the ambient conditions, namely air temperature, globe temperature, radiation and wind speed, prevailing during the two days in winter in
which we observed the blue wildebeest. Lower middle panels show the estimated energy costs of thermoregulation when the 315 mm diameter model was orientated
north and east. Lower panels show the proportion of individuals observed to orientate within each opposing compass arc, per hour, across the two days of observation for
blue wildebeest. Lines above the lower panels indicate the orientation most efficient for energy conservation, i.e. perpendicular to the sun (labelled sun) and parallel to
wind (labelled wind). Left panels illustrate a clear and mild wind picking up in the afternoon, whereas the right panel illustrates a clear day with a light breeze.
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460458
sun position and the cattle’s body axis may result from thenarrow range of solar position associated with satellite imagestaken around midday.
We do not deny the potential for magnetoreception in rumi-nants, particularly in migratory species, but suggest that furtherinvestigation is required to prove such an hypothesis. In order to
Fig. 6. Upper four panels show the ambient conditions, namely air temperature, globe temperature, radiation and wind speed, prevailing during the two days in winter in
which we observed the impala. Lower middle panels show the estimated energy costs of thermoregulation when the 250 mm diameter model was orientated north and
east. Lower panels show the proportion of individuals observed to orientate within each opposing compass arc, per hour, across the two days of observation for impala.
Lines above the lower panels indicate the orientation most efficient for energy conservation, i.e. perpendicular to the sun (labelled sun) and parallel to wind (labelled
wind). Left panels illustrate a clear and cool day with gusty wind picking up in the afternoon, whereas the right panel illustrates a partly cloudy day with a warm and gusty
wind and thundershowers in the afternoon indicated by the rain cloud.
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460 459
confirm the existence of magnetic alignment in ruminants,orientation preferences would need to be investigated undercontrolled conditions in which animal orientation is not influ-enced by thermoregulatory demands. Until such indisputable
evidence is provided, we maintain, based on evidence providedin this paper, that energy demand overrides potential magneticalignment cues and is the primary driver of animal orientationpreferences.
R.S. Hetem et al. / Journal of Thermal Biology 36 (2011) 452–460460
Acknowledgements
We thank Herman van Oeveren and Anna Michta for thedesign and testing of the model and Duncan Mitchell for adviceregarding heat transfer equations. We also thank David andMarlene McCay for allowing the study to take place on theirprivate game reserve, Asante Sana. We are grateful to Richard andKitty Viljoen for their hospitality and on site support. This studywas funded by Shell International, Netherlands, and the FacultyResearch Committee of the University of the Witwatersrand.
References
Altmann, J., 1974. Observational study of behaviour: sampling methods. Behaviour49, 227–267.
Bartholomew, G., 1964. The roles of physiology and behaviour in the maintenanceof homeostasis in the desert environment. In: Hughes, G. (Ed.), Symposia ofthe Society of Experimental Biology, vol. 18: Homeostasis and feedbackmechanisms. Cambridge University Press, Cambridge, pp. 7–29.
Begall, S., Cerveny, J., Neef, J., Vojtech, O., Burda, H., 2008. Magnetic alignment ingrazing and resting cattle and deer. Proc. Natl. Acad. Sci. USA 105,13451–13455.
Berry, H.H., Siegfried, W.R., Crowe, T.M., 1984. Orientation of wildebeest in relationto sun angle and wind direction. Madoqua 13, 297–301.
Brown, K.J., Downs, C.T., 2007. Basking behaviour in the rock hyrax (Procaviacapensis) during winter. Afr. Zool. 42, 70–79.
Burda, H., Begall, S., Cerveny, J., Neef, J., Nemec, P., 2009. Extremely low-frequencyelectromagnetic fields disrupt magnetic alignment of ruminants. Proc. Natl.Acad. Sci. USA 106, 5708–5713.
Chappell, M.A., Bartholomew, G.A., 1981. Standard operative temperatures andthermal energetics of the antelope ground squirrel Ammospermophilus leu-curus. Physiol. Zool. 54, 81–93.
Clapperton, J.L., Joyce, J.P., Blaxter, K.L., 1965. Estimates of the contribution of solarradiation to the thermal exchanges of sheep at a latitude of 551 north. J. Agric.Sci. 64, 37–49.
Clark, V.R., Barker, N.P., Mucina, L., 2009. The Sneeuberg: a new centre of floristicendemism on the Great Escarpment, South Africa. S. Afr. J. Bot. 75, 196–238.
Coe, M., 2004. Orientation, movement and thermoregulation in the giant tortoises(Testudo (Geochelone) gigantea) of Aldabra Atoll, Seychelles. Trans. R. Soc. S.Afr. 59, 73–77.
Coelho, J.R., 2001. Behavioral and physiological thermoregulation in male cicadakillers (Sphecius speciosus) during territorial behavior. J. Therm. Biol. 26,109–116.
Dzialowski, E.M., 2005. Use of operative temperature and standard operativetemperature models in thermal biology. J. Therm. Biol. 30, 317–334.
Fanger, P.O., 1970. Thermal Comfort. Danish Technical Press, Copenhagen.Fortin, D., Larochelle, J., Gauthier, G., 2000. The effect of wind, radiation and body
orientation on the thermal environment of Greater Snow goose goslings.J. Therm. Biol. 25, 227–238.
Gonyou, H.W., Stricklin, W.R., 1981. Orientation of feedlot bulls with respect to thesun during periods of high solar radiation in winter. Can. J. Anim. Sci. 61,809–816.
Hetem, R.S., de Witt, B.A., Fick, L.G., Fuller, A., Kerley, G.I.H., Meyer, L.C.R., Mitchell,D., Maloney, S.K., 2009. Body temperature, thermoregulatory behaviour andpelt characteristics of three colour morphs of springbok (Antidorcas marsupia-lis). Comp. Biochem. Physiol. A Comp. Physiol. 152, 379–388.
Hofmeyr, M.D., Louw, G.N., 1987. Thermoregulation, pelage conductance and renalfunction in the desert-adapted springbok, Antidorcas marsupialis. J. AridEnviron. 13, 137–151.
Humphreys, W., 1991. Thermal behaviour of a small spider (Araneae: araneidae :Araneinae) on horizontal webs in semi-arid Western Australia. Behav. Ecol.Sociobiol. 28, 47–54.
Kennedy, A.D., Day, A.C., 1989. Orientation to the sun of British and exotic breedcross cows during the winter. Appl. Anim. Behav. Sci. 24, 115–125.
Keren, E.N., Olson, B.E., 2006a. Thermal balance of cattle grazing winter range:model application. J. Anim. Sci. 84, 1238–1247.
Keren, E.N., Olson, B.E., 2006b. Thermal balance of cattle grazing winter range:model development. J. Therm. Biol. 31, 371–377.
Keren, E.N., Olson, B.E., 2007. Applying thermal imaging software to cattle grazingwinter range. J. Therm. Biol. 32, 204–211.
Kleiber, M., 1961. The Fire of Life: An Introduction to Animal Energetics. Wiley,New York.
Lillywhite, H.B., 1980. Behavioral thermoregulation in Australian Elapid snakes.Copeia 3, 452–458.
Lustick, S., 1969. Bird energetics: effects of artificial radiation. Science 163, 387–390.Maloney, S.K., Moss, G., Mitchell, D., 2005. Orientation to solar radiation in black
May, M.L., 1976. Thermoregulation and adaptation to temperature in dragonflies(Odonata: Anisoptera). Ecol. Monogr. 46, 1–32.
Mitchell, D., Wyndham, C.H., Vermeulen, A.J., Hodgson, T., Atkins, A.R., Hofmeyr,H.S., 1969. Radiant and convective heat transfer of nude men in dry air. J. Appl.Physiol. 26, 111–118.
Mucina, L., Rutherford, M.C., 2006. The vegetation of South Africa, Lesotho andSwaziland. Strelitzia 19, 1–807.
Muth, A., 1977. Thermoregulatory postures and orientation to the sun: a mechan-istic evaluation for the zebra-tailed lizard, Callisaurus draconoides. Copeia 4,710–720.
O’Neill, K.M., Kemp, W.P., Johnson, K.A., 1990. Behavioural thermoregulation inthree species of robber flies (Diptera, Asilidae: Efferia). Anim. Behav. 39,181–191.
Ohsaki, N., 1986. Body temperatures and behavioural thermoregulation strategiesof three Pieris butterflies in relation to solar radiation. J. Ethol. 4, 1–9.
Olson, B.E., Wallander, R.T., 2001. Orientation of beef cattle grazing foothill winterrange in Montana In: Proceedings of the American Society of Animal Science,Western Section.
Riemerschmid, G., 1943. Some aspects of solar radiation in its relation to cattle inSouth Africa and Europe. Onderstepoort J. Vet. Res. 18, 327–353.
Shelly, T.E., Ludwig, D., 1985. Thermoregulatory behavior of the butterfly Calistonubila (Satyridae) in a Puerto Rican forest. Oikos 44, 229–233.
Stevenson, R.D., 1985. The relative importance of behavioral and physiologicaladjustments controlling body temperature in terrestrial ectotherms. Am. Nat.126, 362–386.
Suter, R.B., 1981. Behavioral thermoregulation: solar orientation in Frontinellacommunis (Linyphiidae), a 6-mg spider. Behav. Ecol. Sociobiol. 8, 77–81.
Tracy, C.R., Tracy, B.J., Dobkin, D.S., 1979. The role of posturing in behavioralthermoregulation by black dragons (Hagenius brevistylus Selys; Odonata).Physiol. Zool. 52, 565–571.
Vrahimis, S., Kok, O.B., 1992. Body orientation of black wildebeest in a semi-aridenvironment. Afr. J. Ecol. 30, 169–175.
Waldschmidt, S., 1980. Orientation to the sun by the iguanid lizardsUta stansburiana and Sceloporus undulatus: hourly and monthly variations.Copeia, 3458–3462.
Walsberg, G.E., 1982. Coat color, solar heat gain, and conspicuousness in thePhainopepla. Auk 99, 495–502.
Walsberg, G.E., 1992. Quantifying radiative heat gain in animals. Am. Zool. 32,217–223.