Division of Microbiology, Indian Agricultural Research Institute (IARI), New Delhi, IndiaInstitute of Microbial Technology, Chandigarh, IndiaDivision of Agricultural Chemicals, Indian Agricultural Research Institute (IARI), New Delhi, IndiaNational Bureau of Agricultural Important Microorganisms, Mau Nath Bhanjan, Uttar Pradesh, IndiaDepartment of Biotechnology, Kurukshetra University, Kurukshetra, India
r t i c l e i n f o
rticle history:eceived 14 March 2013ccepted 7 April 2013vailable online 12 April 2013
eywords:nabaenaungicidal activity
a b s t r a c t
An investigation was undertaken to optimize the physiological conditions and characterize the bioac-tive compound responsible for fungicidal activity in Anabaena laxa. Fungicidal activity against Pythiumdebaryanum in the A. laxa cultures increased, when grown under continuous light (CL) at the stationaryphase (28 d), with further enhancement on doubling phosphorus levels and high pH (9.0) in the growthmedium. Preparatory thin layer chromatography analyses revealed the peptide nature of the fungicidalmetabolite, with highest activity in a spot with Rf 0.13. The bioactive fraction (in terms of fungicidalactivity) was isolated and identified using HPLC with a retention time of 14.7. Fourier transform infrared
T-IRC–MSPLC, NMR
(FT-IR) spectrum of the purified bioactive fraction indicated the cyclic peptide nature of the antifungalcompound. The structural elucidation using 1H, and 13C NMR analyses revealed the same number and typeof carbons as present in previously reported majusculamide C from Lyngbya majuscula. GC–MS indicateda similar major mass ion fragment spectra, with peaks as previously obtained from majusculamide C. Thisrepresents a first report on biosynthesis of a fungicidal compound in A. laxa, with structural similarities
to majusculamide C.
. Introduction
Fungal diseases are one of the most important causes that drasti-ally reduce the agricultural crop production worldwide, for whichynthetic fungicides are the major means of control. The exces-ive use of fungicides causes environmental pollution, toxicity touman beings, besides the development of fungicide-resistance inhese phytopathogens [1]. Toward this endeavor, an eco-friendlyay to reduce the agricultural losses is to use microbes, as biocon-
rol agents, which is simple, cost–effective and user friendly. In thisontext, cyanobacteria are one of the possible candidates becausef their promise as a biofertilizer and ability to produce a diverse
ange of bioactive metabolites [2–6].
Cyanobacteria constitute a group of organisms belonging tohe Bacteria domain, capable of producing a number of secondary
∗ Corresponding author at: Division of Microbiology, Indian Agricultural Researchnstitute (IARI), New Delhi 110012, India. Tel.: +91 011 25848431;ax: +91 011 25846420.
E-mail address: [email protected] (R. Prasanna).1 Centre for Cellular and Molecular Biology (CCMB), Council of Scientific and
ndustrial Research (CSIR), Hyderabad-500 007, Andhra Pradesh, India.
metabolites, exhibiting antibiotic, algicidal, antifungal, cytotoxic,immuno-suppressive and enzyme inhibitory activities [7–9]. Sec-ondary metabolites produced by cyanobacteria such as fischerellinA [10], calophycin [11], laxaphycins [12], scytonemin A [13,14],tanikolide [15], tolytoxin and schizotrin A [16], majusculamide C[17], AK-3 [18] and benzoic acid derivative [19] have been reportedfor their fungicidal activity. Besides serving as allelochemicals andregulators of the succession of algal and bacterial populations inthe aquatic bodies, such chemicals can also serve as herbicide orbio-control agents [5,6,20].
Our earlier investigations have revealed the tremendousgenetic and biochemical diversity within the genus Anabaena andmicrocystin, chitosanase and endoglucanase homologues wereidentified, but no allelochemicals were identified [4,21–25]. Thepresent study was therefore undertaken to explore the possibilityof fungicidal compound (s) and identify, characterize and evaluatethe potential of such metabolite(s) from Anabaena laxa.
2. Materials and methods
2.1. Organism and growth conditions
The axenized culture of A. laxa exhibiting fungicidal activity was selected fromprevious investigations [22,24]. A. variabilis was selected as a negative control for all
V. Gupta et al. / Process Biochemistry 48 (2013) 768–774 769
Table 1List of treatments and fungicidal activity in A. laxa grown under different P levels and pH values under the optimized condition of continuous light (CL) and growth periodof 28 d.
Treatmenta (T) Environmental/nutritional condition Zone of inhibitionb (mm)
a Treatment numbers were also similar in A. variabilis (negative control).b Values presented are means (±SEM) of three replicates.c Control (normal growth conditions). ND, not detectable.
he experiments. The Anabaena strains were grown and maintained in BG11 medium26] at 27 ± 1 ◦C under 16:8 h light:dark cycles. The intensity of white light used was0–55 �mol photons m−2 s−2. The phytopathogenic fungi namely Fusarium oxyspo-um (ITCC 4998), Rhizoctonia solani (ITCC 4578) and Pythium debaryanum (ITCC 95)sed for evaluating fungicidal activity, were obtained from the Indian Type Cultureollection, Division of Plant Pathology, IARI, New Delhi, India. The fungal strainsere grown on Potato Dextrose Agar (PDA) medium and maintained at 28 ± 2 ◦C in
n incubator.
.2. Influence of environmental/nutritional factors on fungicidal activity
An experiment was set up to study the effect of different environmen-al/nutritional factors such as light–dark conditions, pH and phosphorus (P) onungicidal activity. The 14, 21, 28 and 42 d old cultures of both Anabaena strains wererown independently under different light:dark conditions-continuous light (CL)nd dark (CD); L:D-8:16; L:D-16:8. The culture grown under L:D-16:8 was treated as
control. Further, the fungicidal activity in both Anabaena strains grown under dif-erent pH (4.0–12.0) and P levels (43–344 �M) at the optimized growth period (28 d)nd CL condition was evaluated (Table 1). The culture grown under continuous lightCL), in phosphorus concentration (172 �M) and pH (7.5) was treated as control. The
levels were maintained by addition of KCl to provide equivalent balancing counterons in the BG-11 medium [27]. An inoculum level of 10% from 20–25 d old culturesf both Anabaena strains was used uniformly in all the treatments. The extracellularltrates of all the samples were obtained by centrifugation at 4000 × g for 5 min.he fungicidal activity was evaluated by disc diffusion assay using the extracellularltrates [4]. Nystatin (100 U), a commercial antifungal compound (HiMedia) wassed as a positive control against P. debaryanum.
.3. Mass culture of cyanobacteria and extraction of secondary metabolites
Both the Anabaena strains were mass multiplied in 3 L flasks and incubated underhe optimized conditions of T6 (344 �M of P + pH 7.5) and T9 (344 �M of P + pH 9.0)Table 1) along with the controls. The 28 d old cultures were harvested by pelletingn a high speed centrifuge (8000 × g) for 20 min. The cell free culture filtrate extracts
ere pooled and extracted with dichloromethane. The dichloromethane layer waseparated from the aqueous layer and dried over anhydrous sodium sulphate. Theolvent was distilled under reduced pressure on a rotary evaporator. The residuehus obtained was dissolved in minimum quantity of methanol and stored at 4 ◦Cntil further analyses.
.4. Chemical characterization of fungicidal compound
Thin layer chromatography (TLC) was performed on Silica gel 60 (Pre-coatederck) TLC plates using chloroform:ethyl acetate:methanol:water (5:16.5:1) as sol-
ent system. Spots on TLC plates were visualized either by iodine or spraying with solution of ninhydrin (G Biosciences, India). Bands with different Rf values wereeparated and purified with methanol. The fungicidal activity of the partially puri-ed metabolites was observed (as described above) against P. debaryanum. The TLC
ands exhibiting highest fungicidal activity were further purified by preparatoryLC.
The crude dichloro methane (DCM) extracts from both Anabaena strains wereractionated using high-performance liquid chromatography (HPLC), followed byesting for fungicidal activity to identify the bioactive fraction. HPLC was performed
on a Waters 2996 instrument with a photodiode array detector (PAD) system with515 series dual pump system fitted with a PrincetonspherTM reverse phase column(C18, 100 A, 5 �m, 4.6 mm × 250 mm), using a linear gradient of 5–95% of acetoni-trile containing 0.1% (v/v) trifluoroacetic acid (TFA) and water at a flow rate of0.4 ml min−1 at �max 270 nm.
The functional groups present in the biologically active fraction were evaluatedusing FT-IR Synthesis Monitoring System (Perkin Elmer) with sample dispersed inthe KBr pellet. The proton nuclear magnetic resonance spectra (1H NMR) and 13CNMR were recorded on a Bruker 400 AC (400 MHz) NMR. The sample was ana-lyzed by dissolving in a deuteriated CDCl3 and tetramethylsilane (TMS) was usedas an internal standard. The bioactive fraction was analyzed on the Micromass GCTequipped with EI operating at 70 eV and using a source temperature of 250 ◦C, withmass analyzer of TOF and capillary column of HP-5 ms (30 m × 0.32 mm × 0.25 �m).The column oven temperature started at 80 ◦C and kept for 3 min, then reached to240 ◦C at a rate of 8 ◦C/min.
2.5. Statistical analyses
All the data of fungicide al activity was recorded in triplicates. The ANOVA (Anal-ysis of variance) was performed using MSTAT-C statistical package to evaluate thesignificant difference among the means.
3. Results
3.1. Measurement of fungicidal activity under the differentgrowth conditions
A. laxa showed fungicidal activity against P. debaryanum at allstages of growth (14, 21, 28 and 42 d old cultures) under theconditions employed. A two–three fold enhancement in activ-ity was observed from 14 to 28 d of incubation (Fig. 1 andSupplementary Fig. 1A and B). At 28 d, an enhancement of14% in fungicidal activity was observed in CL, as compared tocontrol (L:D-16:8). However, at 28 d, in CD (continuous dark)and L:D-8:16 conditions, fungicidal activity decreased by 67 and58%, respectively as compared to control (Fig. 1). In A. vari-abilis, no fungicidal activity was recorded (Supplementary Fig. 1Aand B).
Supplementary material related to this article found, in theonline version, at http://dx.doi.org/10.1016/j.procbio.2013.04.002.
At the optimized CL condition and growth period of 28 d,in A. laxa measured under different nutritional conditions, anincrease in fungicidal activity of 10 and 40% against P. debaryanum
in treatments T6 (344 �M of P + pH 7.5) and T9 (344 �M ofP + pH 9.0), respectively was recorded, as compared to control(Table 1). In T9 treatment, an increase of 20% in fungicidal activ-ity was observed against F. oxysporum and R. solani, respectively,
770 V. Gupta et al. / Process Biochemistry 48 (2013) 768–774
Fig. 1. Measurement of fungicidal activity of 14 d, 28 d and 42 d old cultures underdifferent light–dark conditions. CL (continuous phase of light); CD (continuous phaseof dark); L:D-16:8 (16 h of light and 8 h of dark, respectively) and L:D-8:16 (8 h oflight and 16 h of dark, respectively). N indicates nystatin (100 U), positive controlfp
a9t
3
tvF
o
rcstp(tpwar
mtd(
Fig. 3. Reverse phase high-performance liquid chromatography of the partially puri-
Fol
or antifungal activity. The differences in the mean were statistically significant at < 0.01 in one way ANOVA test.
s compared to control. However, in T3 treatment (172 �M P + pH.0), fungicidal activity was at par with control against all fungiested.
.2. Isolation and partial purification of the fungicidal compound
The application of crude DCM extracts of A. laxa from T9 showedhe largest zone of inhibition (14 mm). The DCM extracts of A.ariabilis did not show any fungicidal activity (Supplementaryig. S2).
Supplementary material related to this article found, in thenline version, at http://dx.doi.org/10.1016/j.procbio.2013.04.002.
Thin layer chromatography of the crude DCM extracts of A. laxaevealed four spots (Rf values of 0.13, 0.30, 0.42 and 0.55) in theontrol, followed by three (Rf values of 0.13, 0.30 and 0.42) and twopots (Rf values of 0.13 and 0.30) in T6 and T9 treatments, respec-ively, after visualization with iodine (Fig. 2A). The metaboliteurified from a spot with Rf of 0.13 in control and both treatmentsT6 and T9) showed fungicidal activity against P. debaryanum, buthe inhibition zone (14 mm) was largest with the T9 treatment sam-le (Fig. 2B). TLC of crude extract of A. variabilis showed two spotsith Rf values of 0.42 and 0.55 (corresponding to A. laxa) in control
nd both treatments, but no corresponding fungicidal activity wasecorded (data not shown).
In order to evaluate the peptide nature of the fungicidaletabolite, the partially purified extracts of control and both
reatments (T6 and T9) of A. laxa were sprayed with the ninhy-rin which revealed two spots with Rf values of 0.13 and 0.25Fig. 2C).
ig. 2. (A) Thin layer chromatography (TLC) of the partially purified extract of A. laxa witf P + pH 9.0, respectively (Table 1). (B) Fungicidal activity (against Pythium debaryanum)
axa and in A. variabilis, T9 as a negative control. (C) Thin layer chromatography (TLC) of t
fied methanol extracts from T9 treatment (344 �M (P) + pH 9.0) of (A) A. laxa and(B) A. variabilis. (C) Fungicidal activity (against Pythium debaryanum) of the differentHPLC purified fractions of A. laxa.
3.3. HPLC analysis of the crude extract
The crude DCM extracts of control and treatments – T6 and T9 ofA. laxa and A. variabilis were fractioned using reverse phase HPLC.The crude compound belonging to A. laxa and A. variabilis resolved
h iodine vapors. T6 and T9 indicate treatments of 344 �M of P + pH 7.5 and 344 �M of the spot 1 (Rf 0.13) (large circle) in control and both treatments T6 and T9 of A.he partially purified extract of A. laxa with ninhydrin spraying.
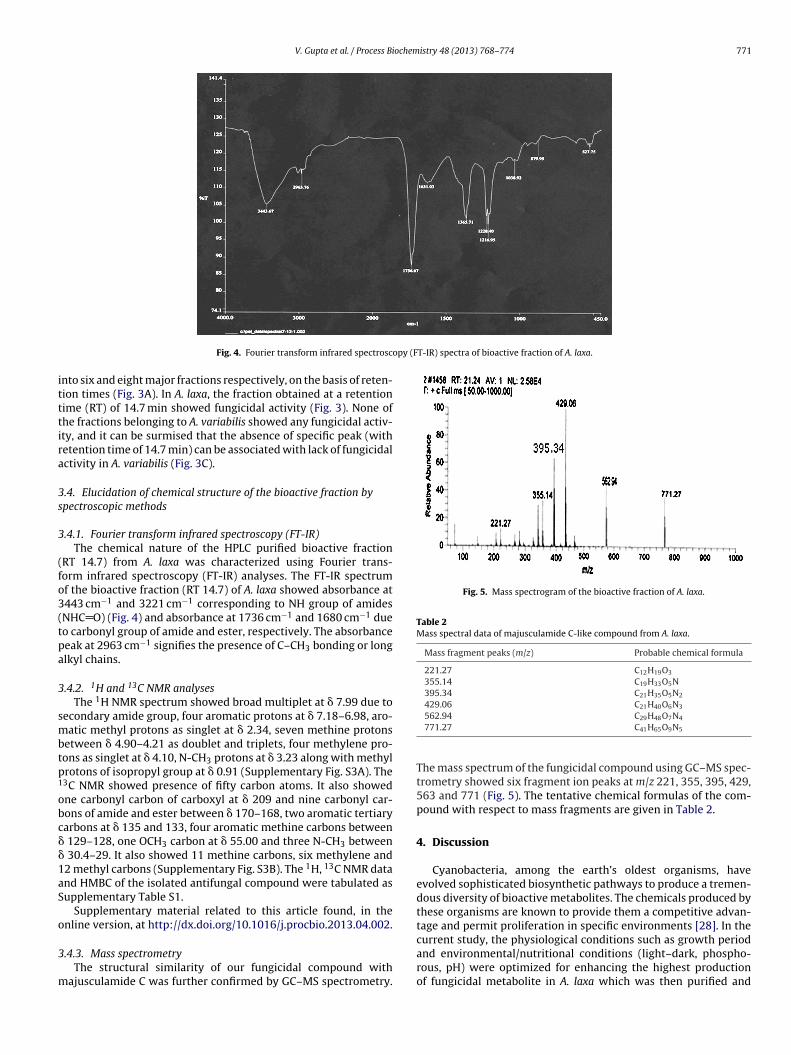
V. Gupta et al. / Process Biochemistry 48 (2013) 768–774 771
opy (FT-IR) spectra of bioactive fraction of A. laxa.
itttira
3s
3
(fo3(tpa
3
smbtp1
obc��1aS
o
3
m
Fig. 5. Mass spectrogram of the bioactive fraction of A. laxa.
Table 2Mass spectral data of majusculamide C-like compound from A. laxa.
Mass fragment peaks (m/z) Probable chemical formula
221.27 C12H19O3
355.14 C19H33O5N395.34 C21H35O5N2
429.06 C21H40O6N3
Fig. 4. Fourier transform infrared spectrosc
nto six and eight major fractions respectively, on the basis of reten-ion times (Fig. 3A). In A. laxa, the fraction obtained at a retentionime (RT) of 14.7 min showed fungicidal activity (Fig. 3). None ofhe fractions belonging to A. variabilis showed any fungicidal activ-ty, and it can be surmised that the absence of specific peak (withetention time of 14.7 min) can be associated with lack of fungicidalctivity in A. variabilis (Fig. 3C).
.4. Elucidation of chemical structure of the bioactive fraction bypectroscopic methods
.4.1. Fourier transform infrared spectroscopy (FT-IR)The chemical nature of the HPLC purified bioactive fraction
RT 14.7) from A. laxa was characterized using Fourier trans-orm infrared spectroscopy (FT-IR) analyses. The FT-IR spectrumf the bioactive fraction (RT 14.7) of A. laxa showed absorbance at443 cm−1 and 3221 cm−1 corresponding to NH group of amidesNHC O) (Fig. 4) and absorbance at 1736 cm−1 and 1680 cm−1 dueo carbonyl group of amide and ester, respectively. The absorbanceeak at 2963 cm−1 signifies the presence of C–CH3 bonding or longlkyl chains.
.4.2. 1H and 13C NMR analysesThe 1H NMR spectrum showed broad multiplet at � 7.99 due to
econdary amide group, four aromatic protons at � 7.18–6.98, aro-atic methyl protons as singlet at � 2.34, seven methine protons
etween � 4.90–4.21 as doublet and triplets, four methylene pro-ons as singlet at � 4.10, N-CH3 protons at � 3.23 along with methylrotons of isopropyl group at � 0.91 (Supplementary Fig. S3A). The3C NMR showed presence of fifty carbon atoms. It also showedne carbonyl carbon of carboxyl at � 209 and nine carbonyl car-ons of amide and ester between � 170–168, two aromatic tertiaryarbons at � 135 and 133, four aromatic methine carbons between
129–128, one OCH3 carbon at � 55.00 and three N-CH3 between 30.4–29. It also showed 11 methine carbons, six methylene and2 methyl carbons (Supplementary Fig. S3B). The 1H, 13C NMR datand HMBC of the isolated antifungal compound were tabulated asupplementary Table S1.
Supplementary material related to this article found, in thenline version, at http://dx.doi.org/10.1016/j.procbio.2013.04.002.
.4.3. Mass spectrometryThe structural similarity of our fungicidal compound with
ajusculamide C was further confirmed by GC–MS spectrometry.
562.94 C29H48O7N4
771.27 C41H65O9N5
The mass spectrum of the fungicidal compound using GC–MS spec-trometry showed six fragment ion peaks at m/z 221, 355, 395, 429,563 and 771 (Fig. 5). The tentative chemical formulas of the com-pound with respect to mass fragments are given in Table 2.
4. Discussion
Cyanobacteria, among the earth’s oldest organisms, haveevolved sophisticated biosynthetic pathways to produce a tremen-dous diversity of bioactive metabolites. The chemicals produced bythese organisms are known to provide them a competitive advan-tage and permit proliferation in specific environments [28]. In thecurrent study, the physiological conditions such as growth period
and environmental/nutritional conditions (light–dark, phospho-rous, pH) were optimized for enhancing the highest productionof fungicidal metabolite in A. laxa which was then purified and
haracterized. The fungicidal activity study under different timeeriods (14, 21, 28 and 42 d) and light dark conditions (CL; CD;:D-16:8; L:D-8:16) in A. laxa revealed highest activity in the con-inuous light (CL) incubation and during the stationary phase (28 d).uch observations are supported by Volk [29], who showed that anntimicrobial compound was accumulated during the stationaryhase in Nostoc insulare. Chitosanase/antifungal activity in our pre-ious investigation [23] was also found to be higher in the 28 d oldultures of A. fertilissima.
Fungicidal activity measured under different light–dark condi-ions was found t be highest in CL and LD (16:8), which revealedhe significance of length of light phase. High light intensity favorsoxin production in cyanobacteria as revealed in several investiga-ions [30–32], while contrasting reports are also available [33–36].he high levels of exogenous organic carbon sources (fructose andlucose) during a long photosynthetic phase can lead to more avail-bility and C and N skeletons directly or through breakdown ofhotosynthetically derived glycogen, leading to increased produc-ion of fungicidal compound. However, our previous investigationevealed highest chitosanase/fungicidal activity in A. fertilissimauring long dark period (L:D-8:16) [23]. Hence, in-depth analyses ofhe differential expression patterns of this enzyme may shed moreight on this aspect of regulation.
Phosphorous being an essential element is known to stimulateroductivity and growth in cyanobacteria [37,38] and its con-entration influences toxin production in cyanobacteria [39]. Theungicidal activity measured at different environmental/nutritionalonditions under the optimized CL condition and growth periodf 28 d suggested that the fungicidal activity is associated withigh P levels. Earlier studies showed an increase in biocidal activ-
ty with the high P levels in the growth medium [27,40]. However,he effects of phosphorus on toxin production are highly variablend sometimes contradictory [41–45]. In our previous study, chi-osanase/antifungal activity was highest at low P concentration in A.ertilissima [25]. Thus, the synthesis of biocidal compound/enzymen cyanobacteria is differentially regulated and may also vary fromtrain to strain; this requires more focused efforts on a large col-ection of strains.
The measurement of fungicidal activity at different pH valuesnder the optimized P level, CL and at growth period of 28 d showed
significant increase at high pH (9.0). However, fungicidal activ-ty was drastically reduced at pH values below 7.5 and above 9.0.oxin production was found to be highest in Calothrix and Microcys-is sp. at pH values between 8.4–8.8 [39,40]. However, a negative
Fig. 6. Structure of majusculamide C from A. laxa
istry 48 (2013) 768–774
correlation was observed in pH and toxin production in O. laete-virens [27]. In our previous study, chitosanase/fungicidal activitywas also observed to be higher at pH 9.0. A pH of 9.0 may thereforebe stimulatory for the activity of antifungal compound/enzymes inAnabaena.
The compound from both the Anabaena strains in control andboth treatments (T6 and T9) was partially purified using DCM asa solvent. In A. laxa, the highest fungicidal activity was observedin T9 treatment as compared to the control and T6 treatment.This revealed that the production of extracellular diffusible anti-fungal compounds in this strain was stimulated by increase in Plevel and pH. Earlier studies have shown that methanolic extractsof cyanobacterial extracellular filtrates exhibited potent fungicidalactivity [46]. Hexane extracts of Pseudomonas aeruginosa Sha8were found highly active against a number of phytopathogenicfungi viz. Aspergillus niger, Helminthosprium sp. and F. oxysporum[47,48].
Thin layer chromatography (TLC) of the crude extracts of A. laxarevealed highest fungicidal activity in T9 treatment with Rf value of0.13. However in A. variabilis, no corresponding spot was detected,indicating the lack of active fungicidal metabolite in A. variabilis.The extracted compound from this treatment also appeared densecompared to that of control and T6 treatments, indicating theenhanced level of secretion of fungicidal compound in A. laxa athigh pH (9.0) and P concentration. The peptide nature of the fungi-cidal metabolite in A. laxa was revealed using ninhydrin spraying,which showed a spot at the same Rf value as observed with iodinevapors.
In order to validate the TLC data, different fractions of controland treatments (T6 and T9) in A. laxa and A. variabilis were collectedusing reverse phase HPLC and tested for their fungicidal activity. InA. laxa, the specific peak obtained at the retention time of 14.7 minwas identified as a bioactive fraction in terms of fungicidal activity,which was absent in A. variabilis.
FT-IR spectrum of the HPLC purified bioactive fraction (RT 14.7)from A. laxa showed the presence of secondary and tertiary nitro-gen and carbonyl group of ester and amide. This type of the spectrais the characteristic of the peptide nature of the fungicidal com-pound and was found nearly similar to a novel antifungal cyclicpeptide compound, majusculamide C from Lyngbya majuscula [49].
1 13
Further, H and C NMR spectrum and structure of the bioactivecompound was compared with this previously reported majus-culamide C which revealed similar number and type of carbonsleading to its structural similarity with majusculamide C. Thus, the
revealed by 1H and 13C NMR spectroscopy.
ochem
siHtcea
aamfiApdn
dstTtffra
A
M(Mu
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
V. Gupta et al. / Process Bi
tructural conclusions drawn from 1H and 13C NMR suggested thedentified antifungal compound as majusculamide C like (Fig. 6).ence, on the basis of IR spectra, 1H and 13C NMR and mass spec-
ral data, the most probable structure assigned to the antifungalompound from A. laxa is highly similar to majusculamide C. How-ver, further crystallographic analyses of this isolated compoundre required for further validation.
Earlier studies identified the derivatives of spermine and piper-zine exhibiting antimicrobial activities (antibacterial, antifungalnd antialgal) from three Oscillatoria species using GC/MS spectro-etric analyses [50–53]. To the best of our knowledge, this is the
rst time reported antifungal compound from the cyanobacterium. laxa. The solubility of the isolated majusculamide C like com-ound from cyanobacterium A. laxa. in solvents such as methanol,ichloromethane and acetone is dissimilar, but its water insolubleature is in consonance with the earlier data on majusculamide C.
In conclusion, the optimization of different physiological con-itions revealed that the fungicidal activity in A. laxa is enhancedignificantly in 28 d (stationary phase) cultures grown under con-inuous light, in BG 11 medium with 344 �M P and high pH (9.0).he fungicidal compound purified from the culture grown underhese optimized physiological conditions and characterized wasound to be highly similar to majusculamide C, previously identifiedrom other cyanobacterium – L. majuscula. This study is a first timeeport which reveals the promise of cyanobacteria as a biocontrolgent, besides it use as a nitrogen fixer.
cknowledgements
This study was supported by AMAAS Network project onicroorganisms, granted by Indian Council of Agricultural Research
ICAR), New Delhi. We thank the authorities of the Division oficrobiology, IARI, New Delhi, for providing necessary facilities for
ndertaking this study.
eferences
[1] Liu YH, Chung YC, Xiong Y. Purification and characterization of a dimethoate-degrading enzyme of Aspergillus niger ZHY256, isolated from sewage. ApplEnviron Microbiol 2001;67:3746–9.
[2] Nain L, Rana A, Joshi M, Shrikrishna JD, Kumar D, Shivay YS, et al. Evaluationof synergistic effects of bacterial and cyanobacterial strains as biofertilizers forwheat. Plant Soil 2010;331:217–30.
[3] Kulik MM. The potential for using cyanobacteria (blue green algae) and algae inthe biological control of plant pathogenic bacteria and fungi. Eur J Plant Pathol1995;101:585–99.
[4] Prasanna R, Lata N, Tripathi R, Gupta V, Middha S, Joshi M, Ancha R, et al. Evalu-ation of fungicidal activity of extracellular filtrates of cyanobacteria – possiblerole of hydrolytic enzymes. J Basic Microbiol 2008;48:186–94.
[5] Manjunath M, Prasanna R, Lata Dureja P, Singh R, Kumar A, et al. Biocontrolpotential of cyanobacterial metabolites against damping off disease caused byPythium aphanidermatum in solanaceous vegetables. Arch Phytopathol PlantProtect 2010;43:666–77.
[6] Dukare AS, Prasanna R, Dubey SC, Chaudhary V, Nain L, Singh R, et al. Evaluatingnovel microbe amended composts as biocontrol agents in tomato. Crop Protect2011;30:436–42.
[7] Mundt S, Kreitlow S, Nowotny A, Effmert U. Biochemical and pharmacolog-ical investigations of selected cyanobacteria. Int J Hygiene Environ Health2001;203:327–34.
[8] Simmons TL, Andrianasolo E, McPhail K, Flatt P, Gerwick WH. Marine naturalproducts as anticancer drugs. Mol Cancer Ther 2005;4:333–42.
[9] Tan LT. Bioactive natural products from marine cyanobacteria for drug discov-ery. Phytochemistry 2007;68:954–79.
10] Hagmann L, Juttner F. Fischerellin A, a novel photosystem-II-inhibiting alle-lochemical of the cyanobacterium Fischerella muscicola with antifungal andherbicidal activity. Tetrahedron Lett 1996;37:6539–42.
11] Moon SS, Chen JL, Moore RE, Patterson GML. Calophycin, a fungicidal cyclicdecapeptide from the terrestrial blue green algae Calothrix fusca. J Org Chem
1992;57:1097–103.
12] Moore RE. Cyclic peptides and depsipeptides from cyanobacteria: a review. JInd Microbiol 1996;16:134–43.
13] Helms GL, Moore RE, Patterson GML. Scytonemin A, a novel calcium antagonistfrom a blue-green alga. J Org Chem 1988;53:1298–307.
[
istry 48 (2013) 768–774 773
14] Pergament I, Carmeli S. Schizotrin A; a novel antimicrobial cyclic peptide froma cyanobacterium. Tetrahedron Lett 1994;35:8473–6.
15] Singh IP, Milligan KE, Gerwick WH. Tanikolide, a toxic and antifungallactone from the marine cyanobacterium Lyngbya majuscula. J Nat Prod1999;62:1333–5.
16] Carmeli S, Moore RE, Patterson GML. Tolytoxin and new scytophycins fromthree species of Scytonema. J Nat Prod 1990;53:1533–42.
17] Frankmolle WP, Larsen LK, Caplan FR, Patterson GML, Knubel G, Moore RE.Antifungal cyclic peptides from the terrestrial blue green algae Anabaena laxa.J Ant 1992;45:1451–7.
18] Yoon YS, Lee CG. Partial purification and characterization of a novel antifungalcompound against Aspergillus spp. from Synechocystis sp. PCC 6803. BiotechnolBioprocess Eng 2009;14:383–90.
19] Natarajan C, Prasanna R, Gupta V, Dureja P, Nain L. Dissecting the fungicidalactivity of Calothrix elenkinii using chemical analyses and microscopy. ApplBiochem Microbiol 2011;48:51–7.
20] Jaiswal P, Singh PK, Prasanna R. Cyanobacterial bioactive molecules – anoverview of their toxic properties. Can J Microbiol 2008;54:701–17.
21] Prasanna R, Kumar R, Sood A, Prasanna BM, Singh PK. Morphological, physio-chemical and molecular characterization of Anabaena strains. Microbiol Res2006;161:187–92.
22] Prasanna R, Gupta V, Natarajan C, Chaudhary V. Bioprospecting for genesinvolved in the production of chitosanases and microcystin-like compoundsin Anabaena strains. World J Microbiol Biotechnol 2010;26:717–24.
23] Gupta V, Prasanna R, Natrajan C, Srivastava AK, Sharma J. Identification, char-acterization and regulation of a novel antifungal chitosanase (cho) in Anabaenasp. Appl Environ Microbiol 2010;76:2769–77.
24] Gupta V, Natarajan C, Kumar K, Prasanna R. Identification and characteriza-tion of endoglucanases for fungicidal activity in Anabaena laxa. J Appl Phycol2011;23:73–81.
25] Gupta V, Prasanna R, Srivastava AK, Sharma J. Purification and characterizationof a novel antifungal endo-type chitosanase from Anabaena fertilissima. AnnMicrobiol 2011;62:1089–98.
26] Stanier RY, Kunisawa R, Mandal M, Cohen-Bazire G. Purification and prop-erties of unicellular blue green algae (Order: Chroococcales). Bacteriol Rev1971;35:171–305.
27] Ray S, Bagchi SN. Nutrients and pH regulate algicide accumulation in culturesof cyanobacterium Oscillatoria laetevirens. New Phytol 2001;149:455–60.
28] Repka S, Meyerhofer M, Von Brockel K, Sivonen K. Associations of cyanobacte-rial toxin nodularin with environmental factors and zooplanktons in the Balticsea. Microbiol Ecol 2004;47:350–8.
29] Volk RB. Studies on culture age versus exometabolite production in batch cul-tures of the cyanobacterium Nostoc insulare. J Appl Phycol 2007;19:491–5.
30] Tonk L, Visser PM, Christiansen G, Dittmann E, Snelder EFM, Wiedner C,et al. The microcystin composition of the cyanobacterium Planktothrix agard-hii changes toward a more toxic variant with increasing light intensity. ApplEnviron Microbiol 2005;71:5177–81.
31] Lehtimaki J, Moisander P, Sivonen K, Kononen K. Growth nitrogen fixation andnodularin production by two Baltic Sea cyanobacteria. Appl Environ Microbiol1997;63:1647–56.
32] Wiedner C, Visser P, Fastner J, Metcalf JS, Codd GA, Mur LR. Effects of light onthe microcystin content of Microcystis strain PCC 7806. Appl Environ Microbiol2003;69:1475–81.
33] Lehtimaki J, Sivonen K, Luukkainen R, Niemela SI. The effects of incubationtime, temperature, light, salinity, and phosphorus on growth and hepatotoxinproduction by Nodularia strains. Arch Hydrobiol 1994;130:269–82.
34] Rapala J, Sivonen K, Luukkainen R, Niemela SI. Anatoxin-a concentration inAnabaena and Aphanizomenon under different environmental conditions andcomparison of growth by toxic and non-toxic Anabaena strains – a laboratorystudy. J Appl Phycol 1993;5:581–91.
35] Sivonen K. Effects of light, temperature, nitrate, orthophosphate, and bacteriaon growth of and hepatotoxin production by Oscillatoria agardhii strains. ApplEnviron Microbiol 1990;56:2658–66.
36] Hobson P, Fallowfield HJ. Effect of irradiance, temperature and salinityon growth and toxin production by Nodularia spumigena. Hydrobiologia2003;493:7–15.
37] Elmetri I, Bell PRF. Effects of phosphorus on the growth and nitrogen fixa-tion rates of Lyngbya majuscula: implication for management in Moreton BayQueensland. Mar Ecol Prog Ser 2004;281:27–35.
38] Albert S, O’Neil JM, Udy JW, Ahern KS, O’Sullivan CM, Dennison WC. Bloomsof the cyanobacterium Lyngbya majuscula in coastal Queensland, Australia:disparate sites, common factors. Mar Poll Bull 2005;51:428–37.
39] Pyo D, Jin J. Production and degradation of cyanobacterial toxin in water reser-voir, lake soyang. Bull Korean Chem Soc 2007;28:800–4.
40] Radhakrishnan B, Prasanna R, Jaiswal P, Nayak S, Dureja P. Modulation ofbiocidal activity of Calothrix sp. and Anabaena sp. by environmental factors.Biologia (Bratisl) 2009;64:881–9.
41] Watanabe MF, Oishi S. Effects of environmental factors on toxicity of acyanobacterium (Microcystis aeruginosa) under culture conditions. Appl Envi-ron Microbiol 1985;49:1342–4.
42] Kotak BG, Lam AKY, Prepas EE, Keneflick SL, Hrudey AE. Variability of the
hepatotoxin, microcystin-LR, in hypereutrophic drinking water lakes. J Phycol1995;31:248–63.
43] Oh HM, Lee SJ, Kim HS, Yoon BD. Microcystin production by Microcys-tis aeruginosa in a phosphorus-limited chemostat. Appl Environ Microbiol2000;66:176–9.
[et al. Highly active anti-Pneumocystis carinii compounds in a library of
74 V. Gupta et al. / Process Bi
44] Kameyama K, Sugiura N, Isoda H, Maekawa T. Effect of nitrate and phosphateconcentration on production of microcystins by Microcystis viridis NIES 102.Aquat Ecosys Health Manag 2002;5:443–9.
45] Vezie G, Rapala J, Vaitomaa J, Seitsonen J, Sivonen K. Effect of nitrogen and phos-phorus on growth of toxic and nontoxic Microcystis strains and on intracellularmicrocystin concentrations. Microb Ecol 2002;43:443–5.
46] Drobac-Cik AV, Dulic TI, Stojanovic DB, Svircev ZB. The importance ofextremophile cyanobacteria in the production of biologically active compound.Proc Natl Sci Matica Srpska Novi Sad 2007;112:57–66.
antagonistic activities of metabolite produced by Pseudomonas aeruginosa Sha8.J Appl Sci Res 2009;5:392–3.
48] Grover M, Nain L, Singh SB, Saxena AK. Molecular approaches for characteriza-tion of antifungal trait of a potent agent Bacillus subtilis RP24. Curr Microbiol2010;60:99–106.
[
istry 48 (2013) 768–774
49] Moore RE, Mynderse JS. A novel antifungal compound majusculamideC from blue-green algae Lyngbya majuscula. US patent 4342751;1982.
50] Dash GK, Suresh P, Sahu SK, Kar OM, Ganapaty S, Panda SB. Evaluation of Evolvu-lus alsinoides Linn. for anthelmintic and antimicrobial activities. J Nat Remedies2002;2:182–5.