ICES mar. Sei. Symp., 197: 159-171. 1993 Estimation of primary production by the simulated in situ method Steven E. Lohrenz Lohrenz, S. E. 1993. Estimation of primary production by the simulated in situ method. - ICES mar. Sei. Symp. ,197: 159-171. The simulation of in situ conditions for primary production incubations may seem a clear objective. However, the inherent assumptions of the method, that subsurface in situ conditions are fully understood and can be duplicated in a surface incubator, are inevitably invalid. It is not surprising then that differences exist between in situ and simulated in situ (SIS) incubation measurements. The objective of this paper is to evaluate the SIS incubation method and discuss its utility. I consider evidence for potential factors causing errors in SIS estimates of actual in situ primary production. These factors may be grouped as (1) irradiance effects, (2) temperature effects, (3) sampling handling effects including, for example, sample collection, processing, and incubation duration, and (4) methods of data analysis by which a final production number is calculated. I argue that proper control of these factors can minimize, although not necessarily eliminate, differences between SIS incubation measurements and actual in situ primary production. Evidence is cited to show that residual differences are comparable to the limits of precision of in vitro primary production estimates. It is concluded that the SIS method, because of its capabilities for extended spatial coverage and logistical control over incubation conditions, remains a valuable tool for the assessment of primary production and microplankton rate processes. Steven E. Lohrenz: Center for Marine Science, University of Southern Mississippi, Stennis Space Center, MS 39529, USA. “The precise determination of in situ daily production beneath a unit of sea surface is extremely difficult and has probably never been achieved.” J. D. H. Strickland, 1965 Introduction While most oceanographers today would agree with the first part of Strickland’s statement, the issue of whether we have yet achieved precise determination of in situ daily production is more controversial. This paper criti- cally examines one approach, the simulated in situ method. In early applications of in vitro techniques to measure primary production in aquatic environments (e.g., Gaarder and Gran, 1927), the use of in situ incubation methods was preferred. However, conven- tional in situ incubation methods were time-consuming and, hence, expensive for oceanographic research con- ducted with large ships. An alternative method was the “imitated” or “simulated" in situ technique for measuring primary production. The simplest version of this method involved collecting surface samples and suspending incubation bottles in a water bath on the deck of a ship (Riley, 1939). Subsequently, the approach was modified to include collection of samples from several depths and imitation of light conditions with neutral density (Steemann Nielsen, 1958; Rytheref a/., 1966) and color filters (Jitts, 1963; Laws et al., 1990). The simulated in situ (SIS) method offers several practical advantages over conventional in situ (IS) incu- bation methods. The lack of recovery and deployment of an IS array makes ship operations less complicated. SIS incubations do not require remaining on or returning to a given location, thus allowing for increased spatial resolution. In addition, SIS incubations provide greater flexibility and more logistical control over incubation conditions (e.g., increased temporal resolution). A dis- advantage of the SIS method is that in situ conditions can only be approximated. Despite this, the SIS method is widely used. For example, at least 75% of productivity data currently archived with the US National Oceano- graphic Data Center were obtained using SIS methods (W. Balch, pers. comm.). Several studies have compared SIS and IS estimates of
Transcript
ICES mar. Sei. Symp., 197: 159-171. 1993
Estimation of primary production by the simulated in situ method
Steven E. Lohrenz
Lohrenz, S. E. 1993. Estimation of primary production by the simulated in situ method. - ICES mar. Sei. Symp. ,197: 159-171.
The simulation of in situ conditions for primary production incubations may seem a clear objective. However, the inherent assumptions of the method, that subsurface in situ conditions are fully understood and can be duplicated in a surface incubator, are inevitably invalid. It is not surprising then that differences exist between in situ and simulated in situ (SIS) incubation measurements. The objective of this paper is to evaluate the SIS incubation method and discuss its utility. I consider evidence for potential factors causing errors in SIS estimates of actual in situ primary production. These factors may be grouped as (1) irradiance effects, (2) temperature effects, (3) sampling handling effects including, for example, sample collection, processing, and incubation duration, and (4) methods of data analysis by which a final production number is calculated. I argue that proper control of these factors can minimize, although not necessarily eliminate, differences between SIS incubation measurements and actual in situ primary production. Evidence is cited to show that residual differences are comparable to the limits of precision of in vitro primary production estimates. It is concluded that the SIS method, because of its capabilities for extended spatial coverage and logistical control over incubation conditions, remains a valuable tool for the assessment of primary production and microplankton rate processes.
Steven E. Lohrenz: Center fo r Marine Science, University o f Southern Mississippi, Stennis Space Center, M S 39529, U SA.
“The precise determination of in situ daily production beneath a unit of sea surface is extremely difficult and has probably never been achieved.”
J. D. H. Strickland, 1965
Introduction
While most oceanographers today would agree with the first part of Strickland’s statement, the issue of whether we have yet achieved precise determination of in situ daily production is more controversial. This paper critically examines one approach, the simulated in situ method. In early applications of in vitro techniques to measure primary production in aquatic environments (e.g., Gaarder and Gran, 1927), the use of in situ incubation methods was preferred. However, conventional in situ incubation methods were time-consuming and, hence, expensive for oceanographic research conducted with large ships. An alternative method was the “imitated” or “simulated" in situ technique for measuring primary production. The simplest version of this method involved collecting surface samples and
suspending incubation bottles in a water bath on the deck of a ship (Riley, 1939). Subsequently, the approach was modified to include collection of samples from several depths and imitation of light conditions with neutral density (Steemann Nielsen, 1958; R ytheref a / . , 1966) and color filters (Jitts, 1963; Laws et al., 1990).
The simulated in situ (SIS) method offers several practical advantages over conventional in situ (IS) incubation methods. The lack of recovery and deployment of an IS array makes ship operations less complicated. SIS incubations do not require remaining on or returning to a given location, thus allowing for increased spatial resolution. In addition, SIS incubations provide greater flexibility and more logistical control over incubation conditions (e.g., increased temporal resolution). A disadvantage of the SIS method is that in situ conditions can only be approximated. Despite this, the SIS method is widely used. For example, at least 75% of productivity data currently archived with the US National Oceanographic Data Center were obtained using SIS methods (W. Balch, pers. comm.).
Several studies have compared SIS and IS estimates of
160 S. E. Lohrenz IC E S m ar . Sei . S y m p . , 197 ( 1993)
primary production (Table 1). The majority of SIS primary production estimates at discrete depths fall within 100% (factor of 2) of IS values. Agreement is better between SIS and IS estimates of water column- integrated primary production. Generally, errors in SIS measurements of in situ primary production can be attributed to the following: (1) irradiance effects, (2) temperature effects, (3) sample handling (sample collection/processing, incubation duration), and (4) methods of data analysis and computation. The objective of this paper is critically to examine evidence that these factors contribute to errors in SIS (and IS) estimates of primary production. Based on this analysis, recommendations for SIS incubation procedures were made. Finally, the precision and accuracy of SIS incubation estimates of in situ primary production were considered. It was concluded that the SIS method, because of its ability to provide extended spatial coverage and flexibility in sample treatment and incubation conditions, remains an important tool for the measurement of primary production and microplankton rate processes.
Evaluation of factors contributing to variations in IS and SIS estimates of primary production
Irradiance effects
Variations in total irradiance
Control of irradiance in SIS incubations is routinely accomplished using filters (neutral density or spectral) to adjust irradiance (solar or artificial) to a desired level. Determination of variations in irradiance with depth can be accomplished by direct measurement (e.g. Jerlov, 1976; J it tseJa /. , 1976). Alternatively, irradiance may be derived indirectly using estimates of vertical attenuation of surface irradiance through the water column. For the latter approach, a common method (e.g. Parsons e/«/.,1984) has been to determine an average irradiance attenuation coefficient ( K \ m ^ 1) for the euphotic zone. The coefficient, K ', can be estimated from subsurface irradiance data or from empirical relationships based on Secchi depth (Isdo and Gilbert, 1974; cf. Preisendorfer, 1986; Megard and Berman, 1989) or other optical properties. Errors in estimating subsurface irradiance may result from the assumption of a uniform K' with depth. Depth-dependent variations in K(z) occur because attenuation of irradiance by sea water varies with wavelength and is affected by composition of particulate and dissolved substances (e.g., Tailing, 1957; Jerlov, 1976; Kirk, 1983; Siegel and Dickey, 1987). Largest depth-related variations in K(z) generally occur near the surface (Fig. 1). Errors in calculating I(z) from a uniform
K (z) ( m 1)0.0 0.1 0.2 0 .3 0 . 4 0 .5
20
40
60
a
0 TCF
20
25
30
Figure 1. Vertical profiles of attenuation coefficients (K(z)) in the northern Gulf of Mexico during October 1990 estimated by direct measurement (solid line) using a Li-Cor LI-192SA underwater quantum sensor with LI-190SA surface reference sensor (both 2j i sensors). The average value of K(z) expected from a Secchi depth determination is shown for comparison (dashed line), (a) Offshore station, 28°31.15'N, 89°2 1 .9 1 'W .
(b) Coastal station, 28°46.71'N, 89°30.55'W.
K' occur in both surface and deep waters for oligotro- phic conditions and primarily in deeper water for more turbid conditions (Fig. 2). More precise approximation of K(z) can be accomplished by modeling it as a linear sum of absorption and scattering properties associated with pure water, particulate matter, and dissolved materials (e.g.. Smith and Baker, 1978; Austin and Petzold, 1984; Prieur and Sathyendranath, 1981; Sath- yendranath and Platt, 1988; Lohrenz era/., 1992).
Temporal variations in irradiance contribute to variations in estimates of production over a wide range of scales (e.g., Marra, 1980; Falkowski, 1984). For many types of variations, simulating in situ irradiance conditions in an incubator is essentially impossible. An example is wave focusing and reflection (Jerlov, 1976; Walsh and Legendre, 1983; Queguiner and Legendre, 1986). In addition, both IS and SIS incubations have been criticized because their “static” nature fails to account for photosynthetic responses to fluctuating irradiance associated with vertical motions (e.g., Harris
IC E S m ar . Sei. S y m p . . 197(1 993) Primary production by the simulated in situ method 161
Table 1. Absolute differences between SIS and IS measured primary production expressed as a percentage of the IS value. All measurements were determined by the l4C method unless otherwise indicated. Means are given, along with standard deviations (s.d.) and ranges, to provide an indication of dispersion (n = number of observations).
Grande et al. ( 1989e) 0 2 method 43(18) 30-56 2I4C 39 (34) 2-70 3 - - -
Gieskes et al. (1990°) 32 (42) 4-250 39 - - -Chiswell et al. (1990) 38 (27) 2-100 31 26 (26) 2-62 4Winn et al. (1992) 53 (34) 1-180 39 20 (9) 10-28 5Lohrenz et al. (1992)
12 h incubations 52 (43) 9-170 28 27(14) 17-42 31 h incubations 30 (5) 24-33 3 - - -
Barber, unpublished 20 (27) 2-67 5 - - -
“Statistics based on digitization of plotted data contained in reference.bBecause of the large number of closely spaced data, digitization of data for discrete depths was not attempted. ^'Negative rates were omitted.
I r r a d i a n c e
( m o l q u a n t a m h )4 5 - 1 0 0
% E r r o r
-50 0 50 100
20
40
60
so
20
25
30
Figure 2. Offshore (A) and coastal (B) vertical profiles of irradiance based on measured (solid line) and depth-invariant (dashed line) estimates of K(z) as in Figure 1. Also shown are the errors in estimated irradiance resulting from the assumption of a uniform K(z) with depth for the offshore (C) and coastal (D) stations. Locations as in Figure 1.
and Lott, 1973; Harris and Piccinin, 1977; Harris, 1978, 1980, 1984; Marra, 1978, 1980). Under some conditions a significant negative bias may exist for static incubation estimates because of increased photoinhibition with increasing exposure time (see under “Incubation duration and non-linearity in rates”).
Alternatively, photoinhibition may occur naturally as, for example, during diel stratification events (Vincent et al., 1984). Internal wave activity has also been considered as a source of variation in in situ irradiance (Haury etal., 1983; Fahnenstiel et al., 1988). Some of the problems of irradiance fluctuations can be addressed through models which incorporate information about photosynthesis-irradiance (P-I) properties and susceptibility to photoinhibition with descriptions of optical and physical regimes (e.g., Gallegos and Platt, 1985). Data for constructing P-I relationships have generally been obtained by incubating samples in artificial light gradients (e.g., Fee, 1973a, b; Jitts etal., 1976; Harrison et al., 1985). Recent work has considered time dependence of P-I parameters (Cullen and Lewis, 1988; Pahl- Wostl, 1990). It is also possible to derive "composite” P - I parameters from pooled water column measurements of available irradiance and primary production deter-
162 S. E. Lohrenz I C E S m ar . Sei. S y m p . . 197 ( 1993)
mined by IS or SIS techniques (e .g ., Menzel and Ryther, 1961; Herman and Platt, 1986; Bower et al., 1987; Lohrenz et al., 1990, 1992). A potential limitation of composite P-I curves is that the derived P-I parameters are not representative of any one phytoplankton population or photoadaptive state (cf. Lewis et al., 1984; Cullen, 1990). Even so, the utility of the composite P-I approach has been demonstrated in coastal (Yoder and Bishop. 1985) and oceanic (Lohrenz et al., 1992) waters.
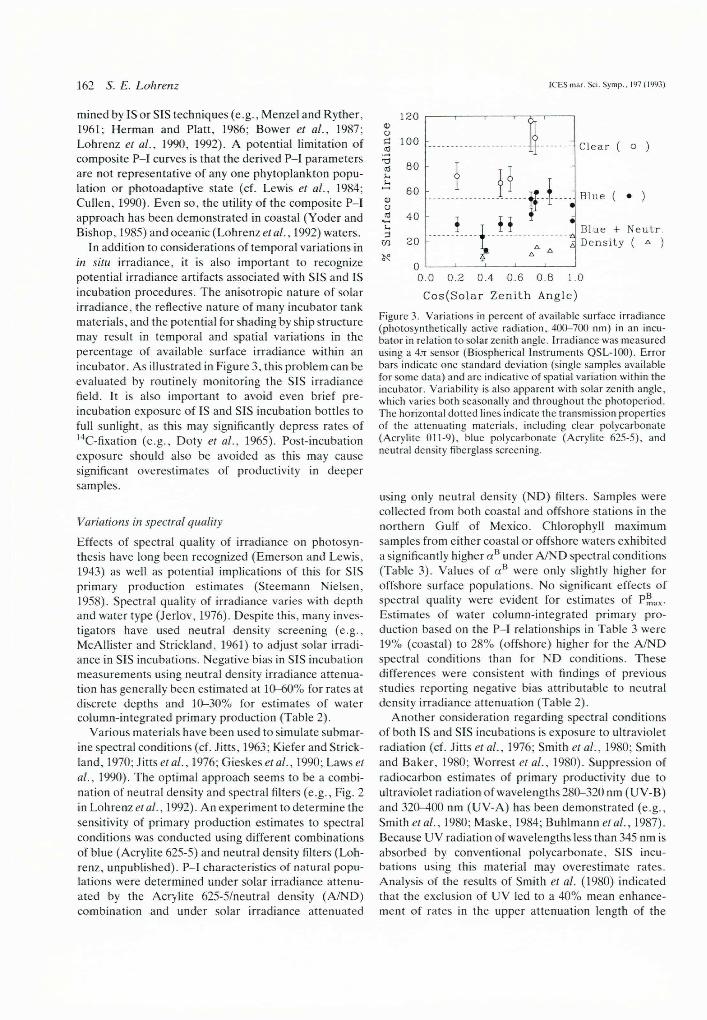
In addition to considerations of temporal variations in in situ irradiance, it is also important to recognize potential irradiance artifacts associated with SIS and IS incubation procedures. The anisotropic nature of solar irradiance, the reflective nature of many incubator tank materials, and the potential for shading by ship structure may result in temporal and spatial variations in the percentage of available surface irradiance within an incubator. As illustrated in Figure 3, this problem can be evaluated by routinely monitoring the SIS irradiance field. It is also important to avoid even brief preincubation exposure of IS and SIS incubation bottles to full sunlight, as this may significantly depress rates of 14C-fixation (e.g., Doty et al., 1965). Post-incubation exposure should also be avoided as this may cause significant overestimates of productivity in deeper samples.
Variations in spectral quality
Effects of spectral quality of irradiance on photosynthesis have long been recognized (Emerson and Lewis, 1943) as well as potential implications of this for SIS primary production estimates (Steemann Nielsen, 1958). Spectral quality of irradiance varies with depth and water type (Jerlov, 1976). Despite this, many investigators have used neutral density screening (e.g., McAllister and Strickland, 1961) to adjust solar irradiance in SIS incubations. Negative bias in SIS incubation measurements using neutral density irradiance attenuation has generally been estimated at 10-60% for rates at discrete depths and 10-30% for estimates of water column-integrated primary production (Table 2).
Various materials have been used to simulate submarine spectral conditions (cf. Jitts, 1963; Kiefer and Strickland, 1970; Jitts etal., 1976; Gieskes etal., 1990; Laws et al., 1990). The optimal approach seems to be a combination of neutral density and spectral filters (e.g., Fig. 2 in Lohrenz et al., 1992). An experiment to determine the sensitivity of primary production estimates to spectral conditions was conducted using different combinations of blue (Acrylite 625-5) and neutral density filters (Lohrenz, unpublished). P-I characteristics of natural populations were determined under solar irradiance attenuated by the Acrylite 625-5/neutral density (A/ND) combination and under solar irradiance attenuated
120
00
80
60 Blue (
4 0Blue + N e u t r .
20
00 .0 0 .2 0 .4 0 .6 0 .8 1.0
Cos(Solar Zen i th Angle)
Figure 3. Variations in percent of available surface irradiance (photosynthetically active radiation, 400-700 nm) in an incubator in relation to solar zenith angle. Irradiance was measured using a 4jt sensor (Biospherical Instruments QSL-100). Error bars indicate one standard deviation (single samples available for some data) and are indicative of spatial variation within the incubator. Variability is also apparent with solar zenith angle, which varies both seasonally and throughout the photoperiod. The horizontal dotted lines indicate the transmission properties of the attenuating materials, including clear polycarbonate (Acrylite 011-9), blue polycarbonate (Acrylite 625-5), and neutral density fiberglass screening.
using only neutral density (ND) filters. Samples were collected from both coastal and offshore stations in the northern Gulf of Mexico. Chlorophyll maximum samples from either coastal or offshore waters exhibited a significantly higher a B under A/ND spectral conditions (Table 3). Values of a B were only slightly higher for offshore surface populations. No significant effects of spectral quality were evident for estimates of PELx- Estimates of water column-integrated primary production based on the P-I relationships in Table 3 were 19% (coastal) to 28% (offshore) higher for the A/ND spectral conditions than for ND conditions. These differences were consistent with findings of previous studies reporting negative bias attributable to neutral density irradiance attenuation (Table 2).
Another consideration regarding spectral conditions of both IS and SIS incubations is exposure to ultraviolet radiation (cf. Jitts e ta/., 1976; Smith et al., 1980; Smith and Baker. 1980; Worrest et al., 1980). Suppression of radiocarbon estimates of primary productivity due to ultraviolet radiation of wavelengths 280-320 nm (UV-B) and 320-400 nm (UV-A) has been demonstrated (e.g., Smith et al., 1980; Maske, 1984; Buhlmann etal., 1987). Because UV radiation of wavelengths less than 345 nm is absorbed by conventional polycarbonate, SIS incubations using this material may overestimate rates. Analysis of the results of Smith et al. (1980) indicated that the exclusion of UV led to a 40% mean enhancement of rates in the upper attenuation length of the
IC E S m ar . Sei. S y m p . , 197(1993) Primary production by the simulated in situ method 163
Table 2. Estimated negative bias attributable to inappropriate spectral conditions in SIS primary production measurements using neutral density light attenuation (%).
Reference
Underestimation relative to in situ:
Discrete depths Water column
Jitts (1963) 0-60 _
Kiefer and Strickland (1970) 66 -
Shimura and Ichimura (1973) 10-30 _
Morel (1978) 45 -
Harrison et al. (1985) 40-60 10-15Lewis et al. (1985a) 38 -
Lewis et al. (1985b) - 30Herman and Platt (1986) 10-30 20Prezelin et al. (1989) - 6-13Laws et al. (1990) 50 -
Prezelin and Glover (1991) - 8-14Schofield el al. ( 1991 ) 30-85 -
This paper 0-54 19-28
water column in the oligotrophic Pacific Ocean. Enhancement was negligible at greater depths. More precise estimates of effects of U V exposure can be made using models that employ UV dose-response relationships (e.g., Smith and Baker, 1980; Smith et a l., 1980; Cullen and Lesser, 1991; Smith et a l ., 1992) in the context of the physical regime.
Temperature effects
Simulation of in situ temperatures is, in principle, less complicated than irradiance because in situ temperatures can be easily measured and incubation temperatures can be controlled precisely. An exception would be in ecosystems having significant temporal and spatial variations (e.g., diel variation in stratification and surface warming). Correction for errors in incubation temperatures may be possible through the evaluation of
photosynthesis-temperature responses. Tailing (1957) expressed the view that usually only light-saturated rates of photosynthesis were temperature-dependent (however, cf. Tilzer et a l., 1986). Li (1985) examined natural phytoplankton assemblages over a wide latitudinal range and found that temperature responses of light- saturated photosynthetic rates were characterized by an optimum range and declined at either suboptimal or supraoptimal temperatures. Based on these results, small differences (i.e., 2-3°C) between ambient and incubation temperatures would have a minimal effect on rates provided that ambient temperatures were near the optimum. However, away from the optimum, rates would be expected to be much more sensitive to temperature changes above or below ambient. A correspondence between optimum and ambient temperatures has been observed only at higher temperatures (>16-18°C) (Aruga, 1965; Li, 1985). Given that surface tempera-
Table 3. A comparison of photosynthesis-irradiancc parameters obtained under different spectral regimes in the northern Gulf of Mexico, October 1990 (n = number of samples, standard errors in parentheses).
Units: P%;,x: gC gChl 1 h a B and ß n: gC gChl 1 (mol quanta m 2)
164 S. E. Lohrenz IC E S m ar . Sei. S y m p . , 1 97(1993)
tures in many productive areas in the ocean are lower than this, it would seem that temperature control should be a critical consideration in SIS incubations.
A common approach to regulation of temperature for SIS methods is to circulate surface sea water through decktop incubators. Inappropriate incubation temperatures may result if there is a large vertical gradient from surface to depth (e.g., Lohrenz et al., 1992) or large horizontal gradients encountered during the incubations. In addition, temperature of surface water supplied by ships’ seawater systems may be poorly regulated (e.g., Marra et al., 1988). A more precise method is to control incubator temperatures using thermostatic baths (e.g., Lohrenz et al., 1992).
Sample handling/incubation duration
Sample handling
There is extensive literature discussing various aspects of sample handling for in vitro (both IS and SIS) primary production measurements. Specific issues include, for example, toxicity effects (Carpenter and Lively, 1980; Fitzwater et al., 1982; Colijn et al., 1983; although cf. Marra and Heinemann, 1984), bottle size or type (Gieskesefa/., 1979; Sakamoto eta l. , 1984), selection of sample depths (Doty etal., 1965; Richardson, 1991), l4C solutions (Bresta eta l., 1987), filtration/sample fixation procedures (Schindler et al., 1972; Lean and Burnison, 1979; Gachter et al., 1984; Goldman and Dennett,1985), and radioassay techniques (Doty et al., 1965; Richardson, 1991). Variations in any procedural step can introduce variability into measurements. For example, average coefficients of variation attributed to processing of identical subsamples by different laboratories were shown to be on the order of 20% (Doty etal., 1965; Richardson, 1991).
Another issue of some concern is the extent to which the physiology of phytoplankton populations may be altered during sample collection prior to incubations (e.g., Taylor et al., 1982). One approach to avoid these problems is to perform all aspects of sampling and incubation in situ. Results from such in situ approaches have been compared to simultaneous SIS incubations that used conventional sample collection methods (Fig. 4). These results support the view that conventional sampling operations do not introduce serious bias, given proper treatment of samples as discussed in the references cited above.
Incubation duration and non-linearity in rates
The choice of incubation duration represents a compromise between (1) incubating long enough to obtain a measurable response integrated over appropriate time scales, and (2) incubating short enough to minimize
effects of confinement. Incubations of up to 24 h are routinely used for estimating daily production, with the understanding that such measurement periods integrate over diel periodicity in rates and that dark respiratory losses are reflected by a decrease in 14C-particulate carbon (Eppley et al., 1973; Eppley and Sharp, 1975). However, long incubations increase the likelihood that population characteristics may be altered by confinement in bottles (e.g., Venrick et al., 1977; Goldman et al., 1981). For example, summation of a series of short incubations has been shown to produce an estimate higher than obtained with a single long-term incubation (Rodhe, 1958; Vollenweider and Nauwerck, 1961; Goldman and Dennett, 1984). An alternative is to conduct short incubations and linearly extrapolate rates over the photoperiod. However, this approach gives rise to other uncertainties.
P r o d u c t i o n ( m gC m h )
0 .0 0 .2 0 .4 0 .6 0 .8 1.00
20
40• • o
60 • in s i t u o d e c k
• o80
12 16840
20
25
Figure 4. Comparison of simulated in situ (SIS) measurements with measurements of primary production in which sample collection, innoculation with 14C, and incubation were all performed in situ. Samples for SIS measurements were collected using Niskin bottles on a Rosette. (A) SIS measurements in relation to in situ measurements determined using an autonomous sampler incubation device as described in Taylor and Doherty (1990) and Lohrenz et al. (1992). (B) SIS measurements (methods in Huntsman and Barber. 1977) compared with in situ measurements executed by scuba divers (R. Barber, unpublished).
IC E S m ar . Sei. S y m p . . 197 (1993) Primary production by the simulated in situ method 165
Compartmentalization of cellular carbon pools may cause short incubations with l4C-inorganic carbon to reflect something between gross and net photosynthetic rates (cf. Hobson et a i , 1976; Dring and Jewson, 1982). Another problem is that extrapolation of short-term rates is complicated by diel periodicity (Doty and Oguri, 1957; Shimada, 1958; Ryther et al., 1961; Lorenzen. 1963; Fee, 1975; Harding et al., 1982a, b). Photosynthetic maxima have been reported to occur at various times of day (Harding et al., 1982a). Consequently, there is no a priori best time of day for making short-term incubation measurements. Finally, there is no way to estimate night respiratory losses directly from short 14C incubations (e.g., Eppley and Sharp, 1975).
Despite the problems associated with both long- and short-term incubations, daily primary production estimated from photoperiod extrapolation of a single midday short-term incubation can agree well (within 50% on
25
20
15
10
5
00 1 2 3
0 1 2 3
4 00
0 20 40 60
S h o r t T e r m P r o d u c t i o n— 3 — 1
(mgC m h )
Figure 5. Relationship between rates estimated with single mid-day short-term (2-6 h) versus 24 h simulated in situ incubations in various locations and times of the year. Short-term rates were expressed on an hourly basis and 24 h rates were expressed on a daily basis. Methods of incubation as described in Lohrenz et al. (1988, 1990). The dotted lines indicate the predicted relationship if hourly rates were extrapolated over the photoperiod. (A) Western Mediterranean, May 1986, 13 h photoperiod. The average difference between hourly extrapolated and 24 h incubation estimates expressed as a percentage of the 24 h value was 42% (range 0.3-98, n=20); (B) western Mediterranean, November 1987,6 h photoperiod. The average difference was 20% (0.4-57, n=30), (C) northwest Atlantic, October 1988, 8 h photoperiod. The average difference was 25% (0.7-180, n=51); (D) Gulf of Mexico, April 1990, 12 h photoperiod. The average difference was 45% (9-110. n=8).
average) with 24 h incubation estimates (Fig. 5). The results in Figure 5 illustrate how a combination of incubation strategies can be used to determine sensitivity of production estimates to incubation duration effects.
Data analysis and computation o f integral primary production
Use of different computational approaches can contribute to variation in both IS and SIS measurements (e.g., Richardson, 1991). The simplest computational method is to integrate discrete estimates over the depth of the euphotic zone. However, simple integration is sensitive to the number and position of samples in a vertically heterogeneous water column. Furthermore, results may be strongly influenced by measurement errors in portions of the water column where production is high. As an alternative to simple integration, various numerical approaches have been used (e.g., Table 4 and cf. Balch et al., 1992). Comparisons between P-I modeled and IS incubation estimates of primary production reveal differences similar in magnitude to those observed between SIS and IS incubation estimates (cf. Table 4 and Table 1). However, the predictive capabilities of P-I methods allow for increased temporal and spatial resolution. “Composite” P-I parameters may be derived from water-column measurements of available irradiance and primary production determined by IS or SIS techniques (see under “Variations in total irradiance”). This approach reduces the possibility of bias due to a single measurement and allows for adjustment for errors in simulating in situ irradiance (e.g., Lohrenz el al., 1992).
Conclusions
R ecom m endations for SIS incubations
(1) Irradiance (incubator and in situ) should be measured precisely and temporal variations considered (even for IS incubations, irradiance conditions may be altered from those experienced by unconfined populations). Determination of P-I relationships in the context of the physical regime may help in determining sensitivity of rates to variations in irradiance and allow for corrections to errors in simulating in situ irradiance. Simulation of spectral quality is recommended, particularly for accuracy in determination of rates at depth.
(2) Temperatures should be precisely controlled, particularly where differences between ambient and optimal temperatures can be expected. Simulation of in situ temperatures should be less difficult than irradiance, except where temperature structure is spatially and/
166 S. E. Lohrenz IC E S m ar . Sei. S y m p . . 197 ( 1993)
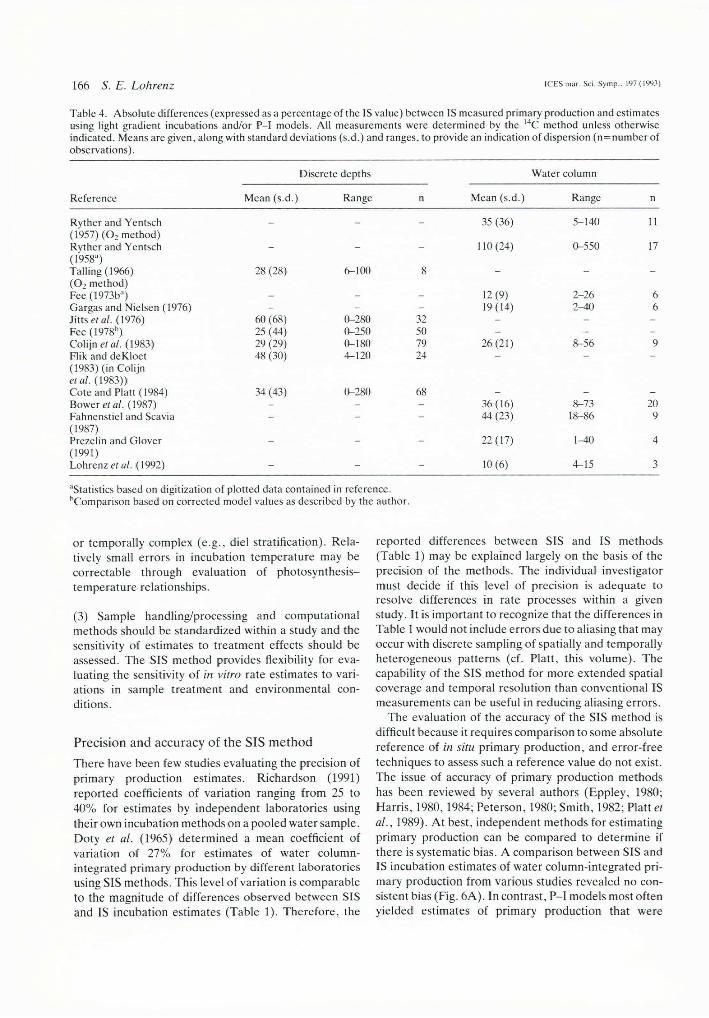
Tabic 4. Absolute differences (expressed as a percentage of the IS value) between IS measured primary production and estimates using light gradient incubations and/or P-l models. All measurements were determined by the l4C method unless otherwise indicated. Means are given, along with standard deviations (s.d.) and ranges, to provide an indication of dispersion (n = number of observations).
(O? method) F e e (1973bü) _ _ _ 12(9) 2-26 6Gargas and Nielsen (1976) - - - 19(14) 2-40 6Jitts el al. (1976) 60 (68) 0-280 32 - - -Fee (1978b) 25 (44) 0-250 50 - - -Colijn etal. (1983) 29 (29) 0-180 79 26(21) 8-56 9Flik and deKloet 48 (30) 4-120 24 - - -(1983) (in Colijn etal. (1983))Cote and Piatt (1984) 34 (43) 0-280 68Bower et al. (1987) - - - 36(16) 8-73 20Fahnenstiel and Scavia - - - 44 (23) 18-86 9(1987)Prezelin and Glover _ _ 22(17) 1—40 4(1991)Lohrenz etal. (1992) - - - 10(6) 4-15 3
“Statistics based on digitization of plotted data contained in reference. C om parison based on corrected model values as described by the author.
or temporally complex (e.g., diel stratification). Relatively small errors in incubation temperature may be correctable through evaluation of photosynthesis- temperature relationships.
(3) Sample handling/processing and computational methods should be standardized within a study and the sensitivity of estimates to treatment effects should be assessed. The SIS method provides flexibility for evaluating the sensitivity of in vitro rate estimates to variations in sample treatment and environmental conditions.
Precision and accuracy o f the SIS method
There have been few studies evaluating the precision of primary production estimates. Richardson (1991) reported coefficients of variation ranging from 25 to 40% for estimates by independent laboratories using their own incubation methods on a pooled water sample. Doty et al. (1965) determined a mean coefficient of variation of 27% for estimates of water column- integrated primary production by different laboratories using SIS methods. This level of variation is comparable to the magnitude of differences observed between SIS and IS incubation estimates (Table 1). Therefore, the
reported differences between SIS and IS methods (Table 1) may be explained largely on the basis of the precision of the methods. The individual investigator must decide if this level of precision is adequate to resolve differences in rate processes within a given study. It is important to recognize that the differences in Table 1 would not include errors due to aliasing that may occur with discrete sampling of spatially and temporally heterogeneous patterns (cf. Platt, this volume). The capability of the SIS method for more extended spatial coverage and temporal resolution than conventional IS measurements can be useful in reducing aliasing errors.
The evaluation of the accuracy of the SIS method is difficult because it requires comparison to some absolute reference of in situ primary production, and error-free techniques to assess such a reference value do not exist. The issue of accuracy of primary production methods has been reviewed by several authors (Eppley, 1980; Harris, 1980, 1984; Peterson, 1980; Smith, 1982; Platter al., 1989). At best, independent methods for estimating primary production can be compared to determine if there is systematic bias. A comparison between SIS and IS incubation estimates of water column-integrated primary production from various studies revealed no consistent bias (Fig. 6A). In contrast, P-I models most often yielded estimates of primary production that were
IC E S m ar . Sei. S y m p . . 197 (1993) Primary production by the simulated in situ method 167
100
80
60
40
20
VII-
0 10 20 30 40 50o
10080
60
40
20
V I I I- 2 0- 4 0
- 6 0
- 8 0
- 1 0 0
I I I
IV
0 10 20 30 40 50 60 70 80
O b s e r v a t i o n
Figure 6. Differences between IS and (A) SIS or (B) P-l modeled estimates of water column-integrated daily primary production for selected studies. Differences are expressed as a percentage of the IS value. Roman numerals correspond to data from the following citations: (A) I, Jitts (1963); 11, Head (1976); III, Brown (1982) ( 0 2 method); IV, Brown (1982) ( l4C method); V, Chiswell etal. (1990); VI, Winn etal. (1992); VII, Lohrenz et al. (1992). (B) I, Ryther and Yentsch (1957); II, Ryther and Yentsch (1958); III, Fee (1973b); IV, Gargas and Nielsen (1976); V. Colijn etal. (1983); VI, Bower et al. (1987); VII, Fahnenstiel and Scavia (1987); VIII, Prezelin and Glover (1991); IX. Lohrenz et al. (1992).
higher than those determined by IS incubations (Fig. 6B). Explanations include the fact that IS (and SIS) incubations were generally longer than those used to determine P-I parameters and, therefore, more susceptible to sample confinement effects.
This leads to the question of the accuracy of in vitro
(both SIS and IS) methods. Evidence from a variety of studies suggests that in vitro estimates, including both 14C and oxygen methods, have not vastly underestimated in situ primary production. This view has been supported by studies comparing in vitro measurements with observed changes in algal cultures (Eppley and Sloan, 1965; Ryther and Menzel, 1965; Peterson, 1978; Lohrenz and Taylor, 1987), ecosystem enclosures (McAllister et a l.. 1961; Antia et a l ., 1963; Davies and
Williams, 1984), and unconfined waters (Ryther et a l.,
1971; Bower e ta l . , 1987; Fahnenstiel and Carrick, 1988; Williams and Purdie, 1991; Daneri, in press). While there do not appear to be large discrepancies, there is a trend of slight negative bias (ca. 30-60%) in the in vitro
measurements compared with determinations in unconfined waters (Bower et a l., 1987; Fahnenstiel and Carrick, 1988; Williams and Purdie, 1991). Conclusions about the significance of a negative bias of 30-60% must remain tentative, since this is comparable to the precision of the in vitro measurements. Further investigation of the precision and accuracy of both in vitro and in situ methods is needed.
SIS measurements o f microplankton rate processes
The question of the relationship of photosynthetic production to carbon cycling in natural populations remains elusive. Attempts to use in vitro rate estimates in constructing carbon budgets for natural populations have been complicated by the fact that net in situ changes may be small relative to production or loss rates (e.g., Jassby and Goldman, 1974; Reynolds et a l ., 1985). A clearer understanding of the role of the non-algal microbial community in carbon cycling is required to resolve these issues. To address these issues, some research requires the use of SIS methods. Consequently, as long as in vitro
techniques are used for studying microplankton processes, the SIS technique will continue to be an important tool.
Acknowledgements
I gratefully acknowledge the comments of G. L. Fahnenstiel and G. A. Knauer on earlier versions of this paper. R. Barber and W. Balch graciously provided data. This research was supported by the office of Naval Research (N00014-88-K-0155), the National Science Foundation (OCE-8801089) through a subcontract with the Bermuda Biological Station for Research, Inc., and by the Coastal Ocean Program Office of the National Oceanic and Atmospheric Administration (NA90AA- D-SG688, Project No. R/LR-25 to Mississippi-Alabama Sea Grant). The Naval Research Laboratory at Stennis Space Center, Mississippi provided facilities support.
References
Antia, N. J., McAllister, C. D., Parsons, T. R., Stephens, K., and Strickland, J. D. H. 1963. Further measurements of primary production using a largc-volumc plastic sphere. Limnol. Occanogr.. 8: 166-183.
Aruga, Y. 1965. Ecological studies of photosynthesis and
168 S. E. Lohrenz IC E S m ar . Sei. S y m p . . 197 (1993)
matter production of phytoplankton. 1. Seasonal changes in photosynthesis of natural phytoplankton. Bot. Mag. Tokyo, 78: 280-288.
Austin, R. W., and Pctzold, T. J. 1984. Spectral dependence of the diffuse attenuation of light in ocean waters. Proc. Soc. Photo-Opt. Instrum. Eng., 489, Ocean Optics VII, 168- 178.
Balch. W., Evans, R., Brown, J., Feldman, G .. McClain, C., and Esaias, W. 1992. The remote sensing of ocean primary productivity: use of a new data compilation to test satellite algorithms. J. Geophys. Res., 97: 2279-2293.
Berge, G. 1958. The primary production in the Norwegian Sea inJunc 1954, measured by an adapted C-14tcchnique. Rapp. P.-v. Réun. Cons. perm. Int. Explor. Mer, 144: 38-46.
Boalch, G. T., Harbour, D .S ., and Butler, E. I. 1978. Seasonal phytoplankton production in the western English Channel. J. mar. biol. Ass. UK, 58: 943-953.
Bower. P. M.. Kelly, C. A., Fee, E. J., Shearer, J. A., DeClercq, D. R., and Schindler, D. W. 1987. Simultaneous measurement of primary production by whole-lake and bottle radiocarbon additions. Limnol. Oceanogr., 32: 299- 312.
Bresta, A. M., Ursin, C., and Jensen, L. M. 1987. Intercomparison of 14C-labellcd bicarbonate solutions prepared by different institutes for measurement of primary productivity in natural waters. J. Plankt. Res., 9(2): 317-325.
Brown, P. C. 1982. Phytoplankton production measured in situ and under simulated in situ conditions in the southern Ben- guela upwelling region. Fish. Bull. S. Afr., 16: 31-37.
Buhlmann, B., Bossard, P., and Uehlinger, U. 1987. The influence of longwave ultraviolet (u.v.-A) on the photosyn- thctic activity ( C-assimilation) of phytoplankton. J. Plankt. Res., 9: 935-943.
Cadée, G. C., and Hegeman, J. 1979. Phytoplankton primary production, chlorophyll and composition in an inlet of the western Wadden Sea (Marsdicp). Neth. J. Sea Res., 13: 224— 241.
Carpenter, E. J., and Lively, J. S. 1980. Review of estimates of algal growth using l4C tracer techniques. In Primary productivity in the sea, pp. 161-178. Ed. by P. G. Falkowski. Plenum Press, New York and London.
Chiswell, S., Firing, E ., Karl, D., Lukas, R., and Winn. C. 1990. Hawaii Ocean Timc-series Data Report 1, 1988-1990. SOEST Technical Report #1.
Colijn, F. W., Gieskes, W. W. C., and Zevenboom, W. 1983. The measurement of primary production: problems and recommendations. Hydrobiol. Bull., 17: 29-51.
Cote, B., and Platt, T. 1984. Utility of the light saturation curve as an operational model for quantifying effects of environmental conditions on phytoplankton photosynthesis. Mar. Ecol. Progr. Ser., 18: 57-66.
Cullen, J. C. 1990. On models of growth and photosynthesis in phytoplankton. Deep-Sea Res., 37: 667-683.
Cullen, J. J . , and Lesser, M. P. 1991. Inhibition of photosynthesis by ultraviolet radiation as a function of dose and dosage rate: results for a marine diatom. Mar. Biol.. I l l : 183-190.
Cullen, J. J., and Lewis, M. R. 1988. The kinetics of algal photoadaptation in the context of vertical mixing. J. Plankt. Res., 10: 1039-1063.
Daneri, G. 1992. Comparison between in vitro and in situ estimates of primary production within two tracked water bodies. Archiv, für Hydrobiologie.
Davies, J. M., and Williams, P. J. leB. 1984. Verification of l4C and 0 2 derived primary organic production measurements using an enclosed ecosystem. J. Plankt. Res., 6: 457-474.
Doty, M. S., Jitts, H. R., Koblentz-Mishke,O. J. ,andSaijo , Y.
Doty. M. S.. and Oguri, M. 1957. Evidence for a photosynthc- tic daily periodicity. Limnol. Oceanogr., 2: 37-40.
Dring, J. J., and Jewson, D . H . 1982. What does l4C uptake by phytoplankton really measure? A theoretical modeling approach. Proc. R. Soc. Lond., 214: 351-368.
Emerson, R., and Lewis, C. M. 1943. The dependence of the quantum yield of Chlorella photosynthesis on wave length of light. Am. J. Bot., 30: 165-178.
Eppley, R. W. 1980. Estimating phytoplankton growth rate in the central oligotrophic oceans. In Primary productivity in the sea, pp. 231-242. Ed. by P. G. Falkowski. Plenum Press, New York and London.
Eppley, R. W., Renger, E. H., Venrick, E. L., and Mullin, M. M. 1973. A study of plankton dynamics and nutrient cycling in the central gyre of the North Pacific Occan. Limnol. Oceanogr., 18: 534-551.
Eppley, R. W., and Sharp. J. H. 1975. Photosynthetic measurements in the central North Pacific: the dark loss of carbon in 24 h incubations. Limnol. Oceanogr., 20: 981-987.
Eppley, R. W., and Sloan, P. R. 1965. Carbon balance experiments with marine phytoplankton. J. Fish. Res. B dC an ., 22: 1083-1097.
Fahnenstiel, G. L., and Carrick, H. J. 1988. Primary production in lakes Huron and Michigan: in vitro and in situ comparisons. J. Plankt. Res., 10: 1273-1283.
Fahnenstiel, G. L., and Scavia, D. 1987. Dynamics of Lake Michigan phytoplankton, primary production and growth. Can. J. Fish, aquat. Sei., 44: 499-508.
Fahnenstiel, G. L., Scavia, D., Lang, G. A., Saylor, J. H., Miller, G. S., and Schwab, D. J. 1988. Impact of inertial period internal waves on fixed-depth primary production estimates. J. Plankt. Res., 10: 77-87.
Falkowski, P. G. 1984. Physiological responses of phytoplankton to natural light regimes. J. Plankt. Res., 6: 295- 307.
Fee, E. J. 1973a. A numerical model for determining integral primary production and its application to Lake Michigan. J. Fish. Res. Bd Can., 30: 1447-1468.
Fcc, E. J. 1973b. Modelling primary production in water bodies: a numerical approach that allows vertical inhomoge- ncities. J. Fish. Res. Bd Can., 30: 1469-1473.
Fee, E. J. 1975. The importance of diurnal variation of photosynthesis vs. light curves to estimates of integral primary production. Vcrh. Int. Verein. Limnol., 19: 39-46.
Fee, E. J. 1978. A procedure for improving estimates of in situ primary production at low irradiances with an incubator technique. Vcrh. Int. Verein. Limnol., 20: 59-67.
Fitzwater, S. E., Knauer, G. A., and Martin, J. H. 1982. Metal contamination and its effect on primary production measurements. Limnol. Oceanogr., 27: 544-551.
Gaarder, T ., and Gran, H. H. 1927. Investigation of the production of plankton in the Oslo fjord. Rapp. P.-v. Réun., Cons. perm. int. Explor. Mer, 42: 1-8.
Gachter, R., Mares, A., and Tilzer, M. M. 1984. Determination of phytoplankton production by the radiocarbon method: a comparison between the acidification and bubbling method (ABM) and the filtration technique. J. Plankt. Res., 6(2): 359-364.
Gallegos, C. L.. and Platt, T. 1985. Vertical advection of phytoplankton and productivity estimates: a dimensional analysis. Mar. Ecol. Prog. Ser., 26: 125-134.
Gallegos, C. L., and Schiebe, F. R. 1986. A device for the continuous measurement of algal photosynthctic response to light. Wat. Res., 20: 509-516."
Gargas, E., and Nielsen, C. S. 1976. An incubator method for
IC E S m ar . Sei. S y m p . . 197 (1993) Primary production by the simulated in situ method 169
estimating the actual daily plankton algae primary production. Water Res., 10: 853-860.
Gieskes, W. W. C., Kraay, G. W., and Baars, M. A. 1979. Current l4C methods for estimating primary production: gross underestimates in natural waters. Neth. J. Sea Res., 13: 58-78.
Gieskes, W. W. C., Kraay, G. W., Nontji, A ., Setiapermana, D., and Sutomo 1990. Monsoonal differences in primary production in the eastern Banda Sea (Indonesia). Neth. J. Sea Res., 25: 473-483.
Goldman, J. C., and Dennett, M. R. 1984. Effect of photoinhibition during bottle incubations on the measurement of seasonal primary production in a shallow coastal water. Mar. Ecol. Prog. Ser., 15: 169-180.
Goldman, J. C., and Dennett, M. R. 1985. Susceptibility of some marine phytoplankton species to cell breakage during filtration and post-filtration rinsing. J. exp. mar. Biol. Ecol., 86: 47-58.
Goldman, J. C., Taylor, C. D., and Glibert, P. M. 1981. Nonlinear time-course uptake of carbon and ammonium by marine phytoplankton. Mar. Ecol. Prog. Ser., 6: 137-148.
Grande, K. D., Williams, P. J. leB., Marra, J., Purdie, D. A., Heinemann, K., Eppley, R. W., and Bender, M. L. 1989. Primary production in the North Pacific gyre: a comparison of rates determined by the l4C, 0 2 concentration and lsO methods. Deep-Sea Res., 36: 1621-1634.
Harding, L. W., Jr., Prezelin, B. B., Sweeney, B. M., and Cox, J. L. 1982a. Diel oscillations of the photosynthesis-irradiance (P-I) relationship in natural assemblages of phytoplankton. Mar. Biol., 67: 167-178.
Harding, L. W., Jr., Prezelin, B. B., Sweeney, B. M., and Cox, J. L. 1982b. Primary production as influenced by diel periodicity of phytoplankton photosynthesis. Mar. Biol., 67: 179— 186.
Harris, G. P. 1978. Photosynthesis, productivity and growth: the physiological ecology of phytoplankton. Arch. Hydro- biol. Beih. Ergebn, Limnol., 10: 1-171.
Harris, G. P. 1980. The measurement of photosynthesis in natural populations of phytoplankton. In The physiological ecology of phytoplankton, pp. 129-187. Ed. by I. Morris, University of California Press, Berkeley.
Harris, G. P. 1984. Phytoplankton productivity and growth measurements: past, present and future. J. Plankt. Res., 6: 219-237.
Harris, G. P., and Lott, J. N. A. 1973. Light intensity and photosynthetic rates in phytoplankton. J. Fish. Res. Bd Can., 30: 1771-1778.
Harris, G. P., and Piccinin, B. B. 1977. Photosynthesis by natural phytoplankton populations. Arch. Hydrobiol., 80: 405-457.
Harrison, W. G ., Platt, T., and Lewis, M. R. 1985. The utility of light-saturation models for estimating marine primary productivity in the field: a comparison with conventional “simulated" in situ methods. Can. J. Fish, aquat. Sei., 4: 864- 872.
Haury, L. R., Wiebe, P. H., Orr, M. H., and Briscoc, M. G. 1983. Tidally generated high-frequency internal wave packets and their effects on plankton in Massachusetts Bay. J. mar. Res., 41: 65-112.
Head, A. 1976. Primary production in an estuarinc environment: a comparison of in situ and simulated in situ C-14 techniques. Estuarine Coastal Mar. Sei., 4: 575-578.
Herman, A. W., and Platt, T. 1986. Primary production profiles in the ocean, estimation from a chlorophyll/light model. Occanol. Acta, 9: 31^10.
Hesslein, R. H., Broccker, W. S., Quay, P. D., and Schlindler, D. W. 1980. Whole-lake radiocarbon experiment in an
oligotrophic lake at the Experimental Lakes Area. Can. J. Fish, aquat. Sei., 37: 454-463.
Hobson, L. A.. Morris, W. J., and Pirquet, K. T. 1976. Theoretical and experimental analysis of the l4C technique and its use in studies of primary production. J. Fish. Res. Bd Can., 33: 1715-1721.
Huntsman, S. A., and Barber, R. T. 1977. Primary production off northwest Africa: the relationship to wind and nutrient conditions. Deep-Sea Res., 24: 25-34.
Idso, S. B., and Gilbert, R. G. 1974. On the universality of the Poole and Atkins Secchi disk-light extinction equation. J. Appl. Ecol., 11: 399-401.
Jassby, A. D., and Goldman, C. R. 1974. Loss rates from a lake phytoplankton community. Limnol. Oceanogr., 19: 618— 627.
Jerlov, N. G. 1976. Marine optics. Elsevier Scientific, New York.
Jitts, H. R. 1963. The simulated in situ measurement of oceanic primary production. Aust. J. Mar. Freshw. Res., 14: 139— 147.
Jitts, H. R., Morel, A., and Saijo, Y. 1976. The relation of oceanic primary production to available photosynthetic irradiance. Aust. J. Mar. Freshw. Res., 27: 441-454.
Kiefer, D. A ., and Strickland, J. H. 1970. A comparative study of photosynthesis in seawater samples incubated under two types of light attenuator. Limnol. Oceanogr., 15: 408-412.
Kirk, J. T. O. 1983. Light and photosynthesis in aquatic ecosystems. Cambridge University Press, Cambridge.
Laws, E. A., DiTullio, G. R., Carder, K. L., Betzer, P. R., and Hawes, S. 1990. Primary production in the deep blue sea. Deep-Sea Res., 37: 715-730.
Lean, D. R. S., and Burnison, B. K. 1979. An evaluation of errors in the l4C method of primary production measurement. Limnol. Oceanogr., 24(5): 917-928.
Lewis, M. R., Cullen, J. J . , and Platt, T. 1984. Relationships between vertical mixing and photoadaptation of phytoplankton: similarity criteria. Mar. Ecol. Prog. Ser., 15: 141— 149.
Lewis, M. R., Warnock, R. E ., Irwin, B., and Platt, T. 1985a. Measuring photosynthetic action spectra of natural phytoplankton populations. J. Phycol., 21: 310-315.
Lewis, M. R., Warnock. R. E., and Platt, T. 1985b. Absorption and photosynthetic action spectra for natural phytoplankton populations: implications for production in the open ocean. Limnol. Oceanogr., 30: 794-806.
Li, W. K. W. 1985. Photosynthetic response to temperature of marine phytoplankton along a latitudinal gradient (16°N to 74°N). Deep-Sea Res., 32: 1381-1391.
Lohrenz, S. E., Dagg, M. J., and Whitledge, T. E. 1990. Enhanced primary production at the plume/oceanic interface of the Mississippi River. Cont. Shelf Res., 10: 639-664.
Lohrenz, S. E., and Taylor, C. D. 1987. Primary production of protein: I Comparison of net cellular carbon and protein synthesis with l4C-derived rate estimates in steady state cultures of marine phytoplankton. Mar. Ecol. Prog. Ser., 356: 277-292.
Lohrenz, S. E ., Wiesenburg, D. A ., DePalma, I. P., Johnson, K. S., and Gustafson, D. E., Jr. 1988. Interrelationships among primary production, chlorophyll, and environmental conditions in frontal regions of the western Mediterranean Sea. Deep-Sea Res., 35: 793-810.
Lohrenz, S. E ., Wiesenburg, D. A., Rein, C. R., Arnone, R. A., Taylor, C. D., Knauer, G. A ., and Knap, A. H. 1992. A comparison of in situ and simulated in situ methods for estimating oceanic primary production. J. Plankt. Res., 14: 201- 221 .
Lorenzen, C. J. 1963. Diurnal variation in photosynthetic
170 S. E. Lohrenz IC E S m ar . Sei. S y m p . , 197 ( 1993)
activity of natural phytoplankton populations. Limnol. Oceanogr., 8: 56-62.
Marra. J. 1978. Phytoplankton photosynthctic response to vertical movement in a mixed layer. Mar. Biol., 46: 203-208.
Marra, J. 1980. Vertical mixing and primary production. In Primary productivity in the sea, pp. 121-137. Ed. by P. G. Falkowski. Plenum Press, New York.
Marra, J.. Haas, L. W., and Heinemann, K. R. 1988. Time course of C assimilation and microbial food web. J. exp. mar. Biol. Ecol., 115: 263-280.
Marra, J., and Heinemann, K. 1984. A comparison between noncontaminating and conventional incubation procedures in primary production measurements. Limnol. Oceanogr.. 29: 389-392.
Maske, H. 1984. Daylight ultraviolet radiation and the photoinhibition of phytoplankton carbon uptake. J. Plankton Res., 6: 351-357.
McAllister, C. D., and Strickland, J. D. H. 1961. Light attenuators for use in phytoplankton photosynthesis studies. Limnol. Oceanogr., 6: 226-228.
McAllister, C. D., Parsons, T. R., Stephens, K.. and Strickland, J. D. H. 1961. Measurements of primary production in coastal sea water using a large-volumc plastic sphere. Limnol. Oceanogr., 6: 237-258.
Megard, R. O.. and Berman, T. 1989. Effects of algae on the Secchi transparency of the southeastern Mediterranean Sea. Limnol. Oceanogr., 34: 1640-1655.
Menzel, D. W., and Ryther, J. H., 1961. Annual variations in primary production of the Sargasso Sea off Bermuda. Deep- Sea Res., 7: 282-288.
Morel, A. 1978. Available, usable and stored radiant energy in relation to marine photosynthesis. Deep-Sea Res., 25: 673- 688 .
Morris, 1., and Glover, H. E. 1974. Questions on the mechanism of temperature adaptation in marine phytoplankton. Mar. Biol., 24: 147-154.
Myers, J. 1971. Enhancement studies in photosynthesis. Ann. Rev. Plant Physiol., 22: 289-312.
Neale, P. J., and Richerson, P. J. 1987. Photoinhibition and the diurnal variation of phytoplankton photosynthesis - I. Development of a photosynthesis-irradiance model from studies of in situ responses. J. Plankt. Res., 9: 167-193.
Neori, A ., and Holm-Hanscn, O. 1982. Effect of temperature on rate of photosynthesis in Antarctic phytoplankton. Polar Biol., 1: 33-38.
Pahl-Wostl, C. 1990. DY PHO RA - a dynamic model for the rate of photosynthesis of algae. J. Plankt. Res., 12: 1207- 1 2 2 1 .
Parsons, T. R., Takahashi, M., and Hargrave, B. 1984. Biological oceanographic processes. Pergamon Press, New York.
Peterson, B. J. 1978. Radiocarbon uptake: its relation to net particulate carbon production. Limnol. Oceanogr., 23: 179— 184.
Peterson, B. J. 1980. Aquatic primary productivity and the l4C 0 2 method: a history of the productivity problem. Annu. Rev. Ecol. Syst., 11: 369-385.
Peterson, D. H., Perry, M. J., Bencala, K. E., and Talbot, M. C. 1987. Phytoplankton productivity in relation to light intensity, a simple equation. Estuarine Coast. Shelf Sei., 24: 813-832.
Platt, T., Gallegos, C. L., and Harrison, W. G. 1980. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. mar. Res., 38: 687-701.
Platt, T ., Harrison, W. G ., Lewis, M. R., Li, W. K. W., Sathyendranath, S., Smith, R. E., and Vezina, A. F. 1989.
Biological production of the oceans: the case for a consensus. Mar. Ecol. Progr. Ser., 52: 77-88.
Platt, T., and Sathyendranath, S. 1988. Oceanic primary production: estimation by remote sensing at local and regional scales. Science, 241: 1613-1620.
Preisendorfcr, R. W. 1986. Secchi disk science: visual optics of natural waters. Limnol. Oceanogr., 31: 909-926.
Prezelin, B. B., and Glover, H. E. 1991. Variability in time/ space estimates of phytoplankton, biomass and productivity in the Sargasso Sea. J. Plankt. Res., 13: 45-67.
Prezelin, B. B., Glover, H. E ., Ver Hoven, B., Steinberg, D., Matlick, H. A., Schofield, O., Nelson, N., Wyman, M., and Campbell, L. 1989. Blue-green light effects on light-limited rates of photosynthesis: relationship to pigmentation and productivity estimates for Synechococcus populations from the Sargasson Sea. Mar. Ecol. Prog. Ser., 54: 121-136.
Prieur, L., and Sathyendranath, S. 1981. An optical classification of coastal and oceanic waters based on the specific spectral absorption curves of phytoplankton pigments, dissolved organic matter, and other particulate materials. Limnol. Oceanogr., 26: 671-689.
Queguiner, B., and Legendre, L. 1986. Phytoplankton photosynthctic adaptation to high frequency light fluctuations simulating those induced by sea surface waves. Mar. Biol., 90: 483-491.
Reynolds, C. S., Harris, G. P., and Gouldney, D. N. 1985. Comparison of carbon-specific growth rates and rates of cellular increase of phytoplankton in large limnetic enclosures. J. Plankt. Res., 7: 791-820.
Richardson, K. 1991. Comparison of 14C primary production determinations made by different laboratories. Mar. Ecol. Prog. Ser., 72: 189-201.
Riley. G. A. 1939. Plankton studies. II. The western North Atlantic, May-June 1939. J. mar. Res., 2: 145-162.
Rodhe, W. 1958. The primary production in lakes: some results and restrictions of the l4C method. Rapp. P.-v. Réun. Cons, perm. int. Explor. Mer, 144: 122-128.
Ryther. J. H. 1956. Photosynthesis in the ocean as a function of light intensity. Limnol. Oceanogr., 1: 61-70.
Ryther, J. H . Hall, J. R.. Pease, A. K., Bakun, A., and Jones, M. M. 1986. Primary organic production in relation to the chemistry and hydrography of the western Indian Ocean. Limnol. Oceanogr., 11: 371-380.
Ryther. J. H ., and Menzel, D. W. 1965. Comparison of the l4C technique with direct measurement of carbon fixation. Limnol. Oceanogr., 10: 490-492.
Ryther, J. H ., Menzel, D. W., Hulburt, E. M., Lorenzen, C. J., and Corwin, N. 1971. The production and utilization of organic matter in the Peru coastal current. Inv. Pesq . ,35:43- 50.
Ryther, J. H., Menzel, D. W., and Vaccaro, R. F. 1961. Diurnal variations in some chemical and biological properties of the Sargasso Sea. Limnol. Oceanogr., 6: 149-153.
Ryther, J. H ., and Vaccaro, R. F. 1954. A comparison of O? and l4C methods of measuring marine photosynthesis. J. Cons. perm. int. Explor. Mer, 20: 25-37.
Ryther, J. H., and Yentsch, C. S. 1957. The estimation of phytoplankton production in the ocean from chlorophyll and light data. Limnol. Oceanogr., 2: 281-286.
Ryther, J. H., and Yentsch, C. S. 1958. Primary production of continental shelf waters off New York. Limnol. Oceanogr., 3: 327-335.
Sakamoto, M., Tilzer, M. M., Gachter, R ., Rai, H ., Collos, Y., Tschumi, P., Berner, P., Zbaren, D ., Zbaren, J., Doku- lil. M., Bossard, P., Uehlinger, U ., and Nusch, E. A. 1984. Joint field experiments for comparisons of measuring
I C E S m ar . Sci. S y m p . . 1 9 7 ( I T O ) Primary production by the simulated in situ method 171
methods of photosynthetic production. J. Plankt. Res., 6: 365-383.
Sathyendranath, S., and Platt, T. 1988. The spcctral irradiance field at the surface and in the interior of the ocean: a model for applications in oceanography and remote sensing. J. Geophys. Res., 93: 9270-9280.
Schindler, D. W., Schmidt, R. V., and Reid, R. A. 1972. Acidification and bubbling as an alternative to filtration in determining phytoplankton production by the l4C method. Can. J. Fish. Res. Bd Can., 29: 1627-1631.
Schofield, O ., Prezelin, B. B., Smith, R. C., Stegmann, P. M., Nelson, N. B., Lewis, M. R., and Baker, K. S. 1991. Variability in spectral and nonspectral measurements of photosynthetic light utilization efficiencies. Mar. Ecol. Prog. Ser., 78: 253-271.
Shimada, B. M. 1958. Diurnal fluctuation of photosynthctic rate and chlorophyll “a" content of phytoplankton from eastern Pacific waters. Limnol. Oceanogr., 3: 336-339.
Shimura. S., and Ichimura, S. 1973. Selective transmission of light in the ocean waters and its relation to phytoplankton photosynthesis. J. Oceanogr. Soc. Japan, 29: 257-266.
Siegel, D. A., and Dickey, T. D. 1987. Observations of the vertical structure of the diffuse attenuation coefficient spectrum. Deep-Sea Res., 34: 547-563.
Slawyk, G ., Maclsaac, J. J., and Dugdale, R. C. 1976. Inorganic nitrogen uptake by marine phytoplankton under in situ and simulated in situ incubation conditions: results from the northwest African upwelling region. Limnol. Occanogr., 21 : 149-152.
Smith, J. C., and Platt, T. 1985. Temperature responses of ribulose bisphosphate carboxylase and photosynthetic capacity in the arctic and tropical phytoplankton. Mar. Ecol., 25: 31-37.
Smith, R. C., and Baker, K. S. 1978. Optical classification of natural waters. Limnol. Oceanogr., 23: 260-267.
Smith, R., C., and Baker, K. S. 1980. Biologically effective dose transmitted by culture bottles in l4C productivity experiments. Limnol. Oceanogr., 25: 364-366.
Smith. R. C., and Baker, K. S. 1981. Optical properties of clearest natural waters (200-800 nm). Appl. Opt., 20: 177— 184.
Smith, R. C., Baker, K. S., Holm-Hansen, O., and Olson, R. 1980. Photoinhibition of photosynthesis and middle ultraviolet radiation in natural waters. Photochem. Photobiol.,31: 585-592.
Smith, R. C., Prezelin, B. B., Baker, K. S.. Bidigare, R. R., Boucher, N. P., Coley, T., Karentz, D., MacIntyre, S., Matlick, H. A ., Mcnzics, D., Ondrusck, M., Wan, Z., and Waters, K. J. 1992. Ozone depletion: ultraviolet radiation and phytoplankton biology in Antarctic waters. Science, 255: 952-959.
Smith, R. E. 1982. The estimation of phytoplankton production and excretion by carbon-14. Mar. Biol. Lett.,3 :3 2 5 - 334.
Steele, J. H. 1962. Environmental control of photosynthesis in the sea. Limnol. Oceanogr., 7: 137-150.
Stccmann Nielsen, E. 1952. The use of radio-activc carbon (C 14) for measuring organic production in the sea. J. Cons, perm. int. Explor. Mer, 18: 117-140.
Stccmann Nielsen, E. 1958. Experimental methods for measuring organic production in the sea. Rapp. P.-v. Rcun. Cons. perm. int. Explor. Mer, 144: 38-46.
Strickland, J. D. H. 1965. Phytoplankton and marine primary production. Ann. Rev. Microbiol., 19: 127-162.
Tailing, J. F. 1957. Photosynthctic characteristics of some freshwater plankton diatoms in relation to underwater radiation. New Phytol., 56: 29-50.
Tailing, J. F. 1966. Photosynthetic behaviour in stratified and unstratified lake populations of a planktonie diatom. J. Ecol., 54: 99-127.
Taylor, C. D., and Doherty, K. W. 1990. Submersible incubation device (SID), autonomous instrumentation for the in situ measurement of primary production and other microbial rate processes. Deep-Sea Res., 37: 343-358.
Taylor, C. D., Molongoski, J. J., and Lohrenz, S. E. 1982. Instrumentation for the measurement of phytoplankton primary production. Limnol. Oceanogr., 28: 781-787.
Tilzer, M. M., Elbrachter, M., Gieskes, W. W., and Beese, B. 1986. Light-temperature interactions in the control of photosynthesis of Antarctic phytoplankton. Polar Biol., 5: 105- i l l .
Vcnrick, E. L., Beers, J. R., and Heinbokel, J. F. 1977. Possible consequences of containing microplankton for physiological rate measurements. J. exp. mar. Biol. Ecol., 26: 55-76.
Vincent, W. F., Neale, P. J., and Richardson, P. J. 1984. Photoinhibition: algal responses to bright light during diel stratification and mixing in a tropical alpine lake. J. Phycol., 2 0 : 201 - 2 1 1 .
Vollcnweidcr, R. A ., and Nauwcrck, A. 1961. Some observations on the l4C method for measuring primary production. Verh. Int. Verein. Limnol., 14: 134-139.
Walsh, P.. and Legendre, L. 1983. Photosynthesis of natural phytoplankton under high frequency light fluctuations simulating those induced by sea surface waves. Limnol. Oceanogr., 28: 688-697.
Williams, P. J. leB., and Purdie, D. A. 1991. In vitro and in situ derived rates of gross oxygen production, net community production and respiration of oxygen in the oligotrophic subtropical gyre of the North Pacific Ocean. Deep-Sea Res., 38: 891-910.'
Winn, C., Chiswell, S., Firing, E ., Karl, D., and Lukas, R. 1992. Hawaii Ocean Time-series Data Report 2, 1990. SOEST-92-l.
Worrest, R. C., Booker, D. L., and Van Dyke, H. 1980. Results of a primary productivity study as affected by the type of glass in the culture bottle. Limnol. Oceanogr., 25: 360-364.
Yoder, J. A., and Bishop, S. S. 1985. Effects of mixing-induced irradiance fluctuations on phytosynthesis of natural assemblages of coastal phytoplankton. Mar. Biol., 90: 87-93.