Review Estrogen and cerebrovascular regulation in menopause Limor Raz ⇑ Department of Neurology, University of New Mexico, Albuquerque, NM, United States article info Article history: Available online xxxx Keywords: 17b-Estradiol Menopausal hormone therapy Cerebrovascular Cardiovascular Stroke Review abstract Estrogen (E2), classically viewed as a reproductive steroid hormone, has non-reproductive functions throughout the body including in the brain and vasculature. Studies report diminished neuroprotection with declining E2 levels, corresponding with higher incidence of cerebrovascular and neurological disease. However, the effects of menopausal hormone therapy (MHT) on the cerebral vasculature and brain function remain controversial. This review will focus on evidence of 17b-estradiol actions in the cerebral vasculature, with a particular emphasis on the vasoactive, anti-inflammatory, anti-oxidant, metabolic and molecular properties. Controversies surrounding MHT in relation to cerebrovascular disease and stroke risk will be discussed, particularly the emerging evidence from clinical trials support- ing the critical period hypothesis of estrogen protection. Ó 2014 Elsevier Ireland Ltd. All rights reserved. Contents 1. Introduction .......................................................................................................... 00 1.1. Mechanisms of E2 and cerebral vascular regulation ..................................................................... 00 1.1.1. Estrogen receptor-mediated mechanisms of cerebrovascular protection ............................................. 00 1.1.2. Signaling pathways of estrogen-induced vasoprotection .......................................................... 00 1.1.3. Indirect actions of estrogen on cerebrovasculature cells of the intravascular compartment.............................. 00 1.2. Inflammation and the blood brain barrier ............................................................................. 00 1.3. Mitochondrial bioenergetics ........................................................................................ 00 1.4. Effects of E2 on cerebral circulation.................................................................................. 00 2. Menopausal hormone therapy and cerebrovascular outcomes .................................................................. 00 2.1. Controversies surrounding menopausal hormone therapy and cerebrovascular risk ........................................... 00 2.2. Hormonal regulation of microvascular pathology ....................................................................... 00 3. Conclusions and future directions ......................................................................................... 00 Author contributions..................................................................................................... 00 Acknowledgements .................................................................................................. 00 References ........................................................................................................... 00 1. Introduction The brain is the most highly perfused organ in the body, consist- ing of specialized vascular beds that sustain metabolic needs of cellular structures (Cipolla et al., 2009). Autoregulatory mecha- nisms of the cerebral circulation assure efficient perfusion by responding to changes in perfusion pressure. Physiological pro- cesses regulating cerebral blood flow (CBF) include perivascular nerve constriction or arterial dilation (Drake and Iadecola, 2007), arterial diameter modifications by astrocyte end feet (Gordon et al., 2007) and endothelial release of hemodynamic and vasoac- tive elements (Faraci and Heistad, 1998). In addition, a distinct and selective microenvironment known as the blood brain barrier (BBB) separates the brain from the peripheral circulation. The BBB consists of cerebral microvascular endothelium, astrocytes, peri- cytes, neurons and extracellular matrix, which altogether forms the ‘‘neurovascular unit’’ (Hawkins and Davis, 2005). These distinct cerebrovascular structures are particularly susceptible to aging pathophysiology and neurological damage (Cipolla et al., 2009; Duckles and Krause, 2007). http://dx.doi.org/10.1016/j.mce.2014.01.015 0303-7207/Ó 2014 Elsevier Ireland Ltd. All rights reserved. ⇑ Address: University of New Mexico, Dominici Hall, MSC11 6035, Albuquerque, NM 87131, United States. Tel.: +1 505 272 0846; fax: +1 505 272 6692. E-mail address: [email protected]Molecular and Cellular Endocrinology xxx (2014) xxx–xxx Contents lists available at ScienceDirect Molecular and Cellular Endocrinology journal homepage: www.elsevier.com/locate/mce Please cite this article in press as: Raz, L. Estrogen and cerebrovascular regulation in menopause. Molecular and Cellular Endocrinology (2014), http:// dx.doi.org/10.1016/j.mce.2014.01.015
Transcript
Molecular and Cellular Endocrinology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Molecular and Cellular Endocrinology
journal homepage: www.elsevier .com/locate /mce
Review
Estrogen and cerebrovascular regulation in menopause
http://dx.doi.org/10.1016/j.mce.2014.01.0150303-7207/� 2014 Elsevier Ireland Ltd. All rights reserved.
⇑ Address: University of New Mexico, Dominici Hall, MSC11 6035, Albuquerque,NM 87131, United States. Tel.: +1 505 272 0846; fax: +1 505 272 6692.
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular regulation in menopause. Molecular and Cellular Endocrinology (2014),dx.doi.org/10.1016/j.mce.2014.01.015
Limor Raz ⇑Department of Neurology, University of New Mexico, Albuquerque, NM, United States
Estrogen (E2), classically viewed as a reproductive steroid hormone, has non-reproductive functionsthroughout the body including in the brain and vasculature. Studies report diminished neuroprotectionwith declining E2 levels, corresponding with higher incidence of cerebrovascular and neurologicaldisease. However, the effects of menopausal hormone therapy (MHT) on the cerebral vasculature andbrain function remain controversial. This review will focus on evidence of 17b-estradiol actions in thecerebral vasculature, with a particular emphasis on the vasoactive, anti-inflammatory, anti-oxidant,metabolic and molecular properties. Controversies surrounding MHT in relation to cerebrovasculardisease and stroke risk will be discussed, particularly the emerging evidence from clinical trials support-ing the critical period hypothesis of estrogen protection.
The brain is the most highly perfused organ in the body, consist-ing of specialized vascular beds that sustain metabolic needs ofcellular structures (Cipolla et al., 2009). Autoregulatory mecha-nisms of the cerebral circulation assure efficient perfusion byresponding to changes in perfusion pressure. Physiological pro-cesses regulating cerebral blood flow (CBF) include perivascular
nerve constriction or arterial dilation (Drake and Iadecola, 2007),arterial diameter modifications by astrocyte end feet (Gordonet al., 2007) and endothelial release of hemodynamic and vasoac-tive elements (Faraci and Heistad, 1998). In addition, a distinctand selective microenvironment known as the blood brain barrier(BBB) separates the brain from the peripheral circulation. The BBBconsists of cerebral microvascular endothelium, astrocytes, peri-cytes, neurons and extracellular matrix, which altogether formsthe ‘‘neurovascular unit’’ (Hawkins and Davis, 2005). These distinctcerebrovascular structures are particularly susceptible to agingpathophysiology and neurological damage (Cipolla et al., 2009;Duckles and Krause, 2007).
2 L. Raz / Molecular and Cellular Endocrinology xxx (2014) xxx–xxx
Estrogen (17b-estradiol or E2), classically known as a reproduc-tive steroid hormone, may function as a neurosteroid in the brain,given its neuroactive properties. In addition to regulation ofgonadotropin releasing hormone (GnRH) release from the hypo-thalamus (Kenealy et al., 2013), de novo synthesis of E2 from tes-tosterone via aromatase within the brain suggest broaderneurosignaling capacity for the hormone (Balthazart and Ball,2006; Hojo et al., 2004; Krause et al., 2011; McEwen et al., 2012;Saldanha et al., 2011). E2 synthesis may occur in the arterial wallof vascular smooth muscle cells directly, thus providing localizedE2 effects and neuroprotection. Aromatase knockout (AKO) ani-mals experience significantly greater ischemic damage as com-pared to controls, with E2 administration entirely reversing theknockout phenotype (McCullough et al., 2003). Although systemiccontributions of E2 to peripheral vascular physiology have alsobeen extensively studied (White, 2002; Babiker et al., 2002; Smileyand Khalil, 2009), the contribution of E2 in modifying cerebrovas-cular function in women throughout the aging process remains tobe elucidated.
As women age, a permanent loss in ovarian follicles results inthe sharp decline in circulating 17b-estradiol (E2) and estrone(E1) levels (Simpson and Davis, 2001; Trevoux et al., 1986) associ-ated, in part, with the onset of menopausal symptoms. Animalstudies and human clinical trials have shown that a state of E2depletion promotes secondary changes in metabolic parameters,thus resulting in a higher prevalence of strokes, which may leadto vascular dementia and Alzheimer’s disease. This review will fo-cus on two important aspects underlying E2 actions in the brainduring menopause. First, the molecular actions of E2 in the cere-bral vasculature will be discussed, with a particular emphasis onvascular tone regulation, inflammation and mitochondrial bioener-getics. Second, clinical evidence and controversies in the field willbe presented on the contributions of menopausal hormone treat-ment (MHT) to cerebrovascular risks and benefits. Examples fromischemic injury models and clinical stroke outcomes will be uti-lized to provide insight to E2 function in disease pathology.
1.1. Mechanisms of E2 and cerebral vascular regulation
The cerebral vasculature contributes to sexually dimorphic neu-rovascular diseases (Bushnell et al., 2006). Evidence supports neu-roprotective, anti-inflammatory, vasodilatory and metaboliceffects of E2 on cerebral blood vessels (Duckles and Krause,2007; Miller and Duckles, 2008). The loss of E2 is thought to pro-mote neurovascular disease in postmenopausal women and wor-sen stroke outcomes and has been associated with cognitivedecline (Henderson and Lobo, 2012). The following sections willhighlight the multifactorial and atheroprotective actions of E2,focusing on the positive and negative experimental data in the fol-lowing areas: (1) cerebrovascular control of myogenic tone, (2)anti-inflammatory actions and generation of cytokines and (3)mitochondrial bioenergetics and free radical production.
1.1.1. Estrogen receptor-mediated mechanisms of cerebrovascularprotection
Receptors for the sex steroids, including E2, are present in thecerebral vasculature, serving as a direct receptor-operated targetby which hormonal actions are mediated. These receptor signalsact through genomic and non-genomic processes to induce vasodi-lation, anti-inflammatory and anti-oxidant effects within cerebralblood vessels (Duckles and Krause, 2011). Both estrogen receptor(ER) a and ERb isoforms are localized within endothelial cells, withERa-specific localization in vascular smooth muscle cells (VSMCs)of cerebral arteries (Dan et al., 2003; Stirone et al., 2003). Molecu-lar techniques demonstrate ERa upregulation in cerebrovasculararterial walls of ovariectomized rats with E2 treatment, with
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
concomitant upreglation of endothelial nitric oxide synthase(eNOS) and nitric oxide (NO)-dependent vasodilation (Stironeet al., 2003). In contrast to ERa, ERb mRNA expression increasesin vascular endothelial and smooth muscle cells after vascularinjury (Lindner et al., 1998). The ERb isoform has been found inthe nuclei of cerebral vascular endothelial cells (Tu and Jufri,2013) and in VSMC (Hodges et al., 2000). Moreover, ERb activationhas also been implicated to reduce blood brain barrier (BBB) break-down and vasogenic edema following cerebral ischemia in studiesof an ERb-selective agonist (Shin et al., 2013). Yet evidence fromERb knockout studies suggests that this receptor subtype is unnec-essary to preserve E2-mediated cerebrovascular protection (Karaset al., 1999). The discovery of a novel membrane-bound G-pro-tein-coupled estrogen receptor-1 (GPER-1/GPR30) and its regula-tion by E2 provide additional support for the vasoprotectiveactions of E2 through rapid signaling. Studies using the GPER-1agonist, G-1, show a reduction in endothelin-1 (ET-1)-mediatedvasoconstriction, corresponding to a decrease in vascular tone(Brunsing and Prossnitz, 2011; Meyer et al., 2012; Revankaret al., 2005).
Recent evidence demonstrates the loss for tissue-specific E2sensitivity with aging. For example, anti-inflammatory effects ofE2 were lost following a 10 day period of hypoestrogenicity butnot immediately following ovariectomy in animal models of stroke(Suzuki et al., 2007). Recent studies have confirmed these observa-tions in middle-aged female rats and showed that a prolonged per-iod of E2 deprivation leads to neuronal cell death and a specific lossof hippocampus CA1 ERa levels, while the uterotropic effects ofERa were preserved following global cerebral ischemia (Zhanget al., 2009). Moreover, it was demonstrated that the degradationof ERa via ubiquitination in neurons occurs as a result of a pro-longed period of hypoestrogenicity in aged rats (Zhang et al.,2011), a mechanism which may help explain the critical periodhypothesis. These studies demonstrate factors affecting complexi-ties and efficacy of MHT on cerebral vascular parameters in post-menopausal women (see Section 2).
1.1.2. Signaling pathways of estrogen-induced vasoprotectionSimilar to the peripheral circulation, E2 regulates cerebral vas-
cular tone by modulating endothelial and smooth muscle cell func-tions, resulting in changes in vascular tone. As in peripheralendothelial cells, E2 facilitates the production of endothelium-de-rived release of vasoactive substances such as nitric oxide (NO),cyclooxygenase (COX) derivatives of arachidonic acid, the contrac-tile agent ET-1 and endothelium-derived hyperpolarizing factor(EDHF) Duckles and Krause, 2007.
E2 decreases myogenic tone of the cerebrovasculature,indirectly, through NO-dependent vasorelaxation. NO has beenextensively studied for its remarkable ability to signal the sur-rounding smooth muscle to relax by elevating cyclic guanosinemonophosphate (cGMP) activity, thus increasing arterial bloodflow (Pelligrino and Galea, 2001; Snyder and Bredt, 1992). Studiesof female cerebral arteries demonstrate stronger NO-dependentvasodilation as compared to age-matched males or ovariectomizedfemales (Geary et al., 2000). Additional evidence suggests thatexogenous E2 supplementation in ovariectomized femalespromotes NO upregulation and cerebral arterial dilation, corre-sponding to a notable reduction in vascular tone (Geary et al.,1998; Weiner et al., 1994). One mechanism responsible forE2-induced NO-dependent vasodilation promotes an elevation ineNOS expression and activation. Prolonged E2 treatment increaseseNOS production and enzymatic activity through an ER-dependentmechanism, as shown by using the selective ERa and ERb inhibitorcompound, ICI182,780 (McNeill et al., 2002). These effects arepartially due to the activation of extracellular, pro-survival,
tion in menopause. Molecular and Cellular Endocrinology (2014), http://
L. Raz / Molecular and Cellular Endocrinology xxx (2014) xxx–xxx 3
phosphatidylinositol-3-kinase-serine-threonine protein kinase(PI3K-Akt) signaling (Stirone et al., 2005).
In addition, EDHF has emerged as an endothelium-dependentvasodilatory factor, regulating CBF through VSMC hyperpolariza-tion (Taylor and Weston, 1988). In vivo studies argue that EDHFis an important regulator of CBF during a pathophysiological state(Bryan et al., 2005), yet the effects of E2 on EDHF in the cerebralcirculation remain controversial. Additional vasoactive factorssuch as prostacyclin (PGI2) and thromboxane A2 (TXB A2) also playan important role in vascular tone maintenance. The balance ofPGI2 to TXB A2 ratio has been associated with regulation of ade-quate CBF (Ospina et al., 2003). Prostaglandins are synthesizedby COX enzymes, thereby inducing changes in blood vessel diam-eter. Interestingly, chronic E2 administration to ovariectomizedfemale rats has been shown to increase prostaglandin productionin cerebral blood vessels through COX-1 and prostacyclin synthaseupregulation (Ospina et al., 2002).
E2 also exerts direct effects on VSMC, including suppressing in-flux of extracellular Ca2+, resulting dilation of cerebral arteriesthrough an ER-independent mechanism (Salom et al., 2001).
In spite of the strong evidence presented above in support ofthe vasodilatory, anti-inflammatory and anti-oxidant role of E2within the cerebral blood vessels, the literature remains inconclu-sive. For instance, E2 only partially restored myogenic tone incerebral arteries from female rodents during focal ischemia(Goodrow et al., 2005). Additional claims for E2-mediated vasore-laxation are contrasted by in vitro studies of EDHF suppression byE2 (Golding et al., 2002), while other in vivo data reports no effect(Xu et al., 2002). Reports documenting the negative effects ofMHT on vascular remodeling show an exacerbation of intimalhyperplasia as a result of common carotid arterial injury, consis-tent with matrix metalloproteinases (MMP)-2 and (MMP)-9upregulation (Mountain et al., 2013). Discrepancies in theseobserved results may be attributed to differences in animal mod-el type, experimental ischemia procedure performed and the ageof animal.
1.1.3. Indirect actions of estrogen on cerebrovasculature cells of theintravascular compartment1.1.3.1. Platelets. E2 also influences myogenic tone through theregulation of blood borne elements such as platelets, lipids andcytokines found within the intravascular compartment. Vasoac-tive factors stored and secreted from platelet dense granulesinclude serotonin (5-hydroxytryptamine, 5-HT), TXB A2 andPGI2, triggering vasoconstriction and vasodilation of cerebralblood vessels and effecting platelet activation (Park and Pepine,2010). The release of 5-HT from activated platelets initiatesserotonergic receptor binding on platelets, vascular endotheliumand VSMC, inducing platelet pro-coagulation associated withincreased cardiovascular risk (Luscher and Vanhoutte, 1986;Yubero-Lahoz et al., 2013). Furthermore, the ability of17b-estradiol to modulate serotonergic receptors has beenshown (McQueen et al., 1997) and is further supported by theassociations between peripheral and cerebral 5-HT levels(Nakatani et al., 2008).
1.1.3.2. Leukocytes and inflammation. Leukocytes are the first line ofdefense against infection, counteracting pathogen invasion byadhering to the endothelial surface during the inflammatory cas-cade. E2 can promote leukocyte de-adhesion in endothelial cellswithin the cerebral microvasculature, thus exerting anti-inflam-matory effects (Pelligrino and Galea, 2001). Evidence shows thatprolonged E2 depletion increases leukocyte adhesion in cerebralpial venules, while E2 replacement reversed these effects (Santizoand Pelligrino, 1999). Reperfusion studies in a model of forebrainischemia further strengthen these observations by showing an
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
E2-mediated reduction in the number of adherent leukocytes inpial venules of the cerebrovasculature (Santizo et al., 2000). Takenas a whole, these findings provide a partial mechanistic explana-tion for the anti-inflammatory cerebrovascular effects of E2.
1.2. Inflammation and the blood brain barrier
Active immune cells release pro-inflammatory mediatorsinducing local and systemic factors which alter vascular tone.Interestingly, sexual dimorphism underlies neuroimmunity and ismediated, in part, by E2 (Chakrabarti et al., 2008). Although thecontributions of neuroinflammation to cardiovascular risk are stillbeing explored in postmenopausal women, the anti-inflammatoryactions of E2 in the cerebral circulation have been extensivelystudied (Ritzel et al., 2013; Straub, 2007).
The anti-inflammatory effects of E2 in the cerebral circulationinclude the suppression of pro-inflammatory cytokines, the reduc-tion in free radical production and a decrease in blood brain per-meability and edema (Zhang et al., 2009; Galea et al., 2002;Sunday et al., 2007). E2 has been shown to decrease cerebralinflammation by the attenuation of endothelial pro-inflammatorymarkers such as interleukin-1 (IL-1) expression, nuclear factor kap-pa-B (NF-kB) activation and leukocyte adhesion (Santizo et al.,2000; Galea et al., 2002; Ospina et al., 2004). In vivo experimentsof lipopolysacharride (LPS)-induced inflammatory responses incerebral blood vessels find greatly diminished inducible NO syn-thase (iNOS) and cyclooxygenase-2 (COX-2) levels during estrous,a period characterized by high serum E2 concentrations (Sundayet al., 2006). Similar reports of chronic E2 treatment have con-firmed the suppression of IL-1, a potent inducer of pro-inflamma-tory prostaglandins in female rat cerebral arteries by the COX-2pathway (Ospina et al., 2004). E2 protection was shown by theattenuation of intercellular adhesion molecule type-1 (I-CAM1)cells and reduction in the transcription factor NF-kB activation incultured brain endothelial cells (Galea et al., 2002). Another wayby which E2 protects cerebral arteries and prevents pro-inflamma-tory vascular damage is by diminishing free radical production. Po-tent oxidative stress markers (i.e. superoxide and peroxynitrite)promote endothelial activation and the recruitment of leukocyteadhesion molecules, thus resulting cell death (Closa and Folch-Puy, 2004). Further discussion on E2 regulation of free radical pro-duction in the cerebral vascular mitochondria is found inSection 1.3.
Cerebrovascular inflammation promotes a secondary wave ofdamage, characterized by leukocyte infiltration, BBB leakinessand vasogenic edema. E2-mediated vasoprotective and anti-inflammatory actions involve the inhibition of leukocyte recruit-ment under basal (Santizo and Pelligrino, 1999) and ischemicstates (Santizo et al., 2000) in intact and ovariectomized femalerats. Under a pathological state, the cerebral endothelium formingthe BBB will become compromised (Balthazart and Ball, 2006;Rosenberg, 2012). E2 has been shown to restore BBB propertiesby decreasing permeability through a reduction in matrix-metallo-proteinase (MMP) expression and by limiting vasogenic edemafollowing cerebral injury (Cipolla et al., 2009).
Although the abovementioned evidence in support of the anti-inflammatory role of E2 is compelling, E2 has also been shown toexert pro-inflammatory actions. Physiological levels of 17b-estra-diol increase endothelial BBB permeability by altering intercellularjunctions and leukocyte recruitment (Groten et al., 2005), conse-quently increasing edema in reproductive senescent females ascompared to controls (Bake and Sohrabji, 2004). Furthermore, instudies of vasoinflammatory disease states such as diabetes andstroke, E2 may initiate pro-inflammatory cascades (Mukherjeeet al., 2005; Xu et al., 2004). These studies show a loss in E2 protec-tion and exacerbation of pro-inflammatory factors, including an
tion in menopause. Molecular and Cellular Endocrinology (2014), http://
4 L. Raz / Molecular and Cellular Endocrinology xxx (2014) xxx–xxx
upregulation of the receptor for advanced glycation end products(RAGE) in diabetics and a post-ischemic increase in leukocyteadhesion response and microglia activation (Marriott et al.,2002). Cumulatively, these data support the critical periodhypothesis of E2 protection, whereby a ‘‘window of opportunity’’for E2 protection exists in animals and humans (Gibbs, 2000;Maki, 2006; Sherwin, 2009). The hypothesis claims that E2 mustbe administered soon after ovarian E2 depletion to be neuro- andvaso-protective and exert positive effects on brain circuitry(Rocca et al., 2011).
1.3. Mitochondrial bioenergetics
The prevalence of mitochondria in cerebral endothelial cells isthree to five times higher as compared with those present in othervascular beds (Grammas et al., 2011). Mitochondria contain the‘‘power house’’ of the cell, generating energy in the form of ATPin a process known as oxidative phosphorylation. Reactive oxygenspecies (ROS) are the byproducts of oxidative phosphorylation.These potent free radicals are elevated with aging, inducing cellu-lar and tissue damage by releasing factors such as superoxide,hydrogen peroxide and peroxynitrite into the mitochondrial ma-trix, resulting BBB leakiness (Enciu et al., 2013; Madamanchiet al., 2005) and leading to atherosclerotic progression (Yu et al.,2012).
E2 antioxidant and vasoprotective actions are partly mediatedby subcellular regulation of the cerebrovascular mitochondria(Rettberg et al., 2013). For instance, E2 has been shown to elevateantioxidant defense enzymes by promoting manganese superoxidedismutase (Mn-SOD) protein expression in cerebral arteries, thusattenuating superoxide (Razmara et al., 2007) and hydrogen perox-ide (Stirone et al., 2005) production. E2 can also reduce intercellu-lar pro-apoptotic signaling cascades responsible for vascular celldeath (Yang et al., 2005). Cytochrome c, a key pro-apoptotic factorand component of oxidative phosphorylation in the mitochondria(Razmara et al., 2008) and mitochondrial subunit complexes Iand IV, supporting the electron transport chain during oxidativephosphorylation, are known to be regulated by E2 (Stirone et al.,2005). Recent evidence by Kemper and colleagues suggests thatE2 depletion in ovary compromised mice suppresses mitochondrialbiogenesis in cerebral endothelial cells. The mechanism underlyingthe loss in E2 vasoprotection involves diminished expression of thetranscription factor, peroxisome proliferator-activated receptor ccoactivator 1 (PGC-1), commonly referred to as the ‘‘master regula-tor’’ of mitochondrial function (Kemper et al., 2013).
Mechanisms for E2-mediated increase in mitochondrial effi-ciency and antioxidant properties depend on ERa and ERb sub-types. E2 upregulates Mn-SOD activation, catalyzing superoxidein an ERa-dependent mechanism (Razmara et al., 2007, 2008).Findings reveal co-localization of ERb in the mitochondria usingDiarylpropionitrile (DPN), a selective ERb-specific agonist (Irwinet al., 2012).
1.4. Effects of E2 on cerebral circulation
The brain vasculature is uniquely different from other vascularbeds throughout the body in its autoregulatory ability to maintainconstant blood flow to meet high metabolic demands. In the brain,both large and small cerebral arterioles contribute to vascularresistance in a mechanism designed to preserve steady CBF,involving interplay of myogenic tone and metabolic mechanisms(Paulson et al., 1990; van Beek et al., 2008).
Studies of E2 effects on the cerebral circulation remain contro-versial. Numerous reports have investigated vasoactive arterialresponse following ovary removal or upon E2 supplementation(Krejza et al., 2001; Nevo et al., 2007). Fifty percent of total
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
cerebrovascular resistance consists of pial arteries, which partiallyregulate CBF and protect downstream microvascular structuresfrom increased hydrostatic pressure and injury (McHedlishvili,1980). Other cerebrovascular components include penetratingparenchymal arterioles (PA), which are long, unbranched vesselsconnecting the pial circulation to the microcirculation Cipollaet al., 2009 and exhibiting greater basal tone and low concentra-tions of 5-HT receptors (Cipolla et al., 2004). Administration ofsupraphysiological concentrations of E2 improved perfusion tothe hippocampus following ischemic injury as compared to intactand ovariectomized female rats (He et al., 2002). E2-mediated in-crease in CBF was also documented during resting state in healthy,elderly and postmenopausal women with underlying cerebrovas-cular pathology (Maki and Resnick, 2001). Clinical studies demon-strate that hemodynamic changes inversely correlate to menstrualcycle patterns, as associations between E2 concentrations andcerebrovascular resistance were found in women of late follicularphase (Brackley et al., 1999; Krejza et al., 2004). Thus, E2 appearsto augment blood flow (Diomedi et al., 2001).
Contrary to these results, in rodent models of ischemic injury,chronic, physiological concentrations of E2 protected the cortexagainst Middle Cerebral Artery Occlusion (MCAO)-induced damagewithout effecting CBF (Alkayed et al., 2000; Dubal and Wise, 2001;Rusa et al., 1999). These findings were further strengthened by evi-dence from young, ovariectomized rats where basal CBF wasunchanged following 6 weeks of E2 deprivation (Holschneiderand Scremin, 1998). Discrepancies in findings highlight the impor-tance of E2 concentration administered, the timing of initiationand type of ischemic insult in experimental animal models.
E2 also promotes the formation of new blood vessels and im-proves functional recovery following ischemic insult in a processknown as angiogenesis. 17b-estradiol increases vascular densityby 22% in the peri-infarcted region 10 days following ischemia(Alqahtani et al., 2007). Other reports support the regenerativeeffects of E2 by demonstrating an increase in vascularity, improvedperfusion and motor function after MCAO (Ardelt et al., 2012).Speculative reports suggest that the mechanism for E2-mediatedendothelial regeneration may involve the upregulation of circulat-ing endothelial progenitor cells (Strehlow et al., 2003).
2. Menopausal hormone therapy and cerebrovascular outcomes
The effects of MHT on cerebrovascular and neurological healthin humans have been extensively studied, suggesting that E2administration is protective (Brann et al., 2007; Harman et al.,2011). The correlation between systemic vascular disease and cere-bral blood vessel health has also been well characterized (Cotteret al., 2003; Slark et al., 2012).
One major limitation of clinical trials conducted thus far is thatsome are representative of an elderly menopausal women popula-tion with cardiovascular risk factors, years past the onset ofmenopause. As a result, MHT usage may not be effective in reduc-ing the risk of stroke or VTE among women with establishedvascular disease or against stroke prevention. Several randomizedand placebo-controlled trials investigated stroke outcomes inwomen with pre-established vascular disease. The Heart and Estro-gen/progestin Replacement Study (HERS) and Women’s Estrogenfor Stroke Trial (WEST), preceded the Women’s Health Initiative(WHI) studies and measured stroke risk as a secondary outcome.HERS participants were postmenopausal women with pre-existingcoronary heart disease taking MHT or placebo (Hulley et al., 1998).The WEST enrolled postmenopausal women with a history ofischemic stroke or transient ischemic attacks (Viscoli et al.,2001). In both trials, the incidence of stroke for women given E2or placebo was HR: 2.89 (95% CI: 1.50–5.58) and RR: 2.9 (95% CI:0.9–9.0), respectively.
tion in menopause. Molecular and Cellular Endocrinology (2014), http://
L. Raz / Molecular and Cellular Endocrinology xxx (2014) xxx–xxx 5
The WHI findings are in contrast to prior studies which haveconsistently reported non-significant increases in stroke risk withMHT (Viscoli et al., 2001; Shumaker et al., 2004; Simon et al.,2001). These studies, however, provided the turning point in clin-ical care of menopausal women. Due to the high incidence of breastcancer and elevated stroke risk in the E2+Progestin trial, WHIinvestigators concluded that the overall health risks exceed thebenefits, leading to the early termination of the study (Rossouwet al., 2002). The WHI were randomized, multi-centered, placebo-controlled trials measuring stroke as a secondary outcome. Thewomen had an elevated cardiovascular risk profile reflecting thegeneral population, with a mean age of 63. Participants were givenconjugated estrogens with or without medroxy-progesterone ace-tate. Evidence from the WHI trials indicates that oral conjugatedequine estrogen (oCEE) alone, or with a progestin, increases awoman’s risk of stroke by approximately one third, equivalent tonine additional cases of stroke per 10,000 person-years of hormoneuse (Rossouw et al., 2002; Anderson et al., 2004; Hendrix et al.,2006; Wassertheil-Smoller et al., 2003).
The risk of venous thromboembolism (VTE), defined as the for-mation of clots in blood vessels, has also been well-studied in men-opausal women. Meta-analyses for the risk of venousthromboembolism (VTE) with oral and transdermal hormoneadministration documented similar findings to those of strokestudies specified above. Although the risk from VTE was elevatedin users of both preparations, it did not reach statistical signifi-cance (Canonico et al., 2008). Consequently, a risk for VTE wasfound upon comparison of former and non-users of oral MHT prep-aration in the WHI studies. The observed increase in clotting factorproduction may be attributed to oral estradiol hepatic actions(Heiss et al., 2008). Nevertheless, studies of cognition in meno-pausal women report improved cognitive abilities as measuredby verbal memory and abstract reasoning (Wharton et al., 2009).Taken altogether, these findings reiterate the importance of thecritical window of E2 in humans and confirm that timing of hor-mone administration must be considered to preserve the favorablephysiological effects of E2.
Approximately a decade following the WHI studies, whichdefined the standard of care for menopausal women, MHT admin-istration remains controversial. The WHI study design has beenquestioned by many experts in the field (Clark, 2006; Naftolinet al., 2004). Reanalysis of the WHI data from the E2 plus progestinarm of the trial suggests that reduced risk for coronary heartdisease in perimenopausal women may appear only after five to6 years of treatment (Clark, 2006; Taylor and Manson, 2011). Theseclaims were further confirmed by additional reanalysis of the WHIE2-Alone Trial, supporting the beneficial effects of prolonged E2exposure. Hysterectomized, postmenopausal women taking oralE2 for a duration of 6 years were not at an increased ordecreased risk of coronary heart disease and strokes, whileyounger women showed a significant reduction in these outcomes(LaCroix et al., 2011). Findings suggest that a longer duration ofMHT may be needed to induce neuro- and cardio-protectivephysiological mechanisms, thus producing improved vascularoutcomes (Harman et al., 2011).
Of particular interest, support for the timing hypothesis of E2protection was observed in the Danish Osteoporosis PreventionStudy of perimenopausal women. Following 10 years of random-ized treatment, women receiving MHT early after menopausehad a significantly reduced risk of mortality, heart failure, or myo-cardial infarction, without an increase in the risk of stroke, VTE andcancer (Schierbeck et al., 2012).
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
Current data demonstrates that the route of E2 administrationand steroid metabolism may be responsible for the observed car-diovascular outcomes. Reports from a case-control study of theWomen’s International Study of Long-Duration Estrogen afterMenopause (WISDOM) compared oCEE or estradiol and transder-mal therapy, finding a dose–response relationship, whereby areduction in the risk of stroke only occurs with low-dose transder-mal but not oral administration (650 lg/day) (Renoux et al., 2010;Speroff, 2010). Additional support for transdermal MHT wasobserved in menopausal Japanese women, emphasizing the com-plexities of E2 effects on blood vessel physiological function andatherosclerotic progression (Sumino and Murakami, 2013). Anexplanation for these findings may involve differences in hormonemetabolism and bioavailability. The transdermal route of hormoneadministration avoids first-pass hepatic metabolism, thereby cre-ating stable E2 serum levels and less pronounced effects on hepaticprotein synthesis, thus blocking pro-inflammatory and pro-throm-botic coagulation cascades. Conversely, oral treatment inducessupraphysiological E2 concentrations in the liver with hyper-coag-ulant effects, leading to an increase in hepatic sex hormone bindingglobulin (SHBG) production and elevated estrone (E1) levels(Goodman, 2012).
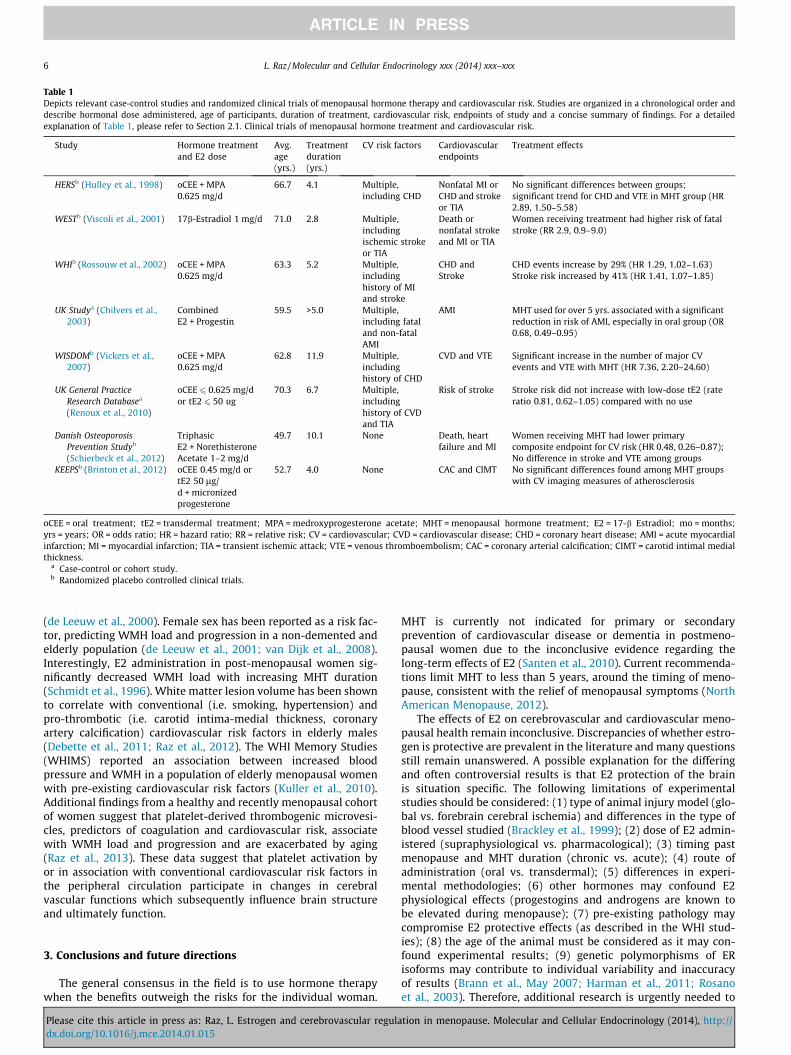
Table 1 provides a summary of MHT treatment effects on car-diovascular outcomes presented in chronologically ordered clinicaltrials. It is noteworthy that MHT timing, duration, dose and thehealth status of women predict cardiovascular risk. Several keyconcepts are illustrated in Table 1: (1) the ‘‘critical period hypoth-esis’’ of E2 protection is consistent with studies of relatively young,recently menopausal women, when MHT is administered closer tothe timing of menopause. Studies depicting healthy cohorts ofwomen at the onset of studies offer additional support for thehypothesis. The window of opportunity for E2 protection isthought to exist during pre- and peri-menopausal years, as E2has been shown to exert detrimental effects on previously dam-aged vasculature, an effect exacerbated by aging (Chen et al.,2006); (2) the composition of estradiol preparation consists of aconglomeration of hormones. Estrone (E1) sulfate is the primaryhormone of conjugated equine estrogen (CEE) formulations. Sinceestradiol (E2) binds to the ER with greater affinity than E1, clinicaloutcomes may be altered in accordance to the biochemical natureof the MHT preparation (Bhavnani et al., 1998; Punnonen andLukola, 1982); (3) lower, transdermal doses of E2 have been asso-ciated with greater cardiovascular benefit, negating E2 effects onhepatic metabolism and reducing pro-coagulant factors (Speroff,2010) and (4) short MHT duration of less than 5 years may beinsufficient to induce significant physiological change. LongerMHT administration of 10 or more years may exert positive effectson cardiovascular and neurovascular biology (Harman et al., 2011).
2.2. Hormonal regulation of microvascular pathology
Aging is accompanied by histopathological and structural cere-brovascular changes, including inconsistencies in CBF, hypoperfu-sion and decreased vascular density (Moody et al., 1990). Thesecerebrovascular abnormalities may lead to the development oftortuous arterioles and increased vascular resistance, promotingwhite matter susceptibility to injury (Moody et al., 1991). Lesionsof white matter can be detected on magnetic resonance imaging(MRI), and are typically seen in the aging population. These whitematter hyperintensities (WMH) predict stroke risk and cognitivedecline in large population studies (Brown and Thore, 2011).Serving as indicators for small vessel pathology, WMH have beenassociated with gliosis, endothelial dysfunction and BBBpermeability (Wardlaw et al., 2013).
WMH physiology and histopathology remain poorly under-stood, with only a few studies addressing sex-specific differences
tion in menopause. Molecular and Cellular Endocrinology (2014), http://
Table 1Depicts relevant case-control studies and randomized clinical trials of menopausal hormone therapy and cardiovascular risk. Studies are organized in a chronological order anddescribe hormonal dose administered, age of participants, duration of treatment, cardiovascular risk, endpoints of study and a concise summary of findings. For a detailedexplanation of Table 1, please refer to Section 2.1. Clinical trials of menopausal hormone treatment and cardiovascular risk.
Study Hormone treatmentand E2 dose
Avg.age(yrs.)
Treatmentduration(yrs.)
CV risk factors Cardiovascularendpoints
Treatment effects
HERSb (Hulley et al., 1998) oCEE + MPA0.625 mg/d
66.7 4.1 Multiple,including CHD
Nonfatal MI orCHD and strokeor TIA
No significant differences between groups;significant trend for CHD and VTE in MHT group (HR2.89, 1.50–5.58)
WESTb (Viscoli et al., 2001) 17b-Estradiol 1 mg/d 71.0 2.8 Multiple,includingischemic strokeor TIA
Death ornonfatal strokeand MI or TIA
Women receiving treatment had higher risk of fatalstroke (RR 2.9, 0.9–9.0)
WHIb (Rossouw et al., 2002) oCEE + MPA0.625 mg/d
63.3 5.2 Multiple,includinghistory of MIand stroke
CHD andStroke
CHD events increase by 29% (HR 1.29, 1.02–1.63)Stroke risk increased by 41% (HR 1.41, 1.07–1.85)
a Case-control or cohort study.b Randomized placebo controlled clinical trials.
6 L. Raz / Molecular and Cellular Endocrinology xxx (2014) xxx–xxx
(de Leeuw et al., 2000). Female sex has been reported as a risk fac-tor, predicting WMH load and progression in a non-demented andelderly population (de Leeuw et al., 2001; van Dijk et al., 2008).Interestingly, E2 administration in post-menopausal women sig-nificantly decreased WMH load with increasing MHT duration(Schmidt et al., 1996). White matter lesion volume has been shownto correlate with conventional (i.e. smoking, hypertension) andpro-thrombotic (i.e. carotid intima-medial thickness, coronaryartery calcification) cardiovascular risk factors in elderly males(Debette et al., 2011; Raz et al., 2012). The WHI Memory Studies(WHIMS) reported an association between increased bloodpressure and WMH in a population of elderly menopausal womenwith pre-existing cardiovascular risk factors (Kuller et al., 2010).Additional findings from a healthy and recently menopausal cohortof women suggest that platelet-derived thrombogenic microvesi-cles, predictors of coagulation and cardiovascular risk, associatewith WMH load and progression and are exacerbated by aging(Raz et al., 2013). These data suggest that platelet activation byor in association with conventional cardiovascular risk factors inthe peripheral circulation participate in changes in cerebralvascular functions which subsequently influence brain structureand ultimately function.
3. Conclusions and future directions
The general consensus in the field is to use hormone therapywhen the benefits outweigh the risks for the individual woman.
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
MHT is currently not indicated for primary or secondaryprevention of cardiovascular disease or dementia in postmeno-pausal women due to the inconclusive evidence regarding thelong-term effects of E2 (Santen et al., 2010). Current recommenda-tions limit MHT to less than 5 years, around the timing of meno-pause, consistent with the relief of menopausal symptoms (NorthAmerican Menopause, 2012).
The effects of E2 on cerebrovascular and cardiovascular meno-pausal health remain inconclusive. Discrepancies of whether estro-gen is protective are prevalent in the literature and many questionsstill remain unanswered. A possible explanation for the differingand often controversial results is that E2 protection of the brainis situation specific. The following limitations of experimentalstudies should be considered: (1) type of animal injury model (glo-bal vs. forebrain cerebral ischemia) and differences in the type ofblood vessel studied (Brackley et al., 1999); (2) dose of E2 admin-istered (supraphysiological vs. pharmacological); (3) timing pastmenopause and MHT duration (chronic vs. acute); (4) route ofadministration (oral vs. transdermal); (5) differences in experi-mental methodologies; (6) other hormones may confound E2physiological effects (progestogins and androgens are known tobe elevated during menopause); (7) pre-existing pathology maycompromise E2 protective effects (as described in the WHI stud-ies); (8) the age of the animal must be considered as it may con-found experimental results; (9) genetic polymorphisms of ERisoforms may contribute to individual variability and inaccuracyof results (Brann et al., May 2007; Harman et al., 2011; Rosanoet al., 2003). Therefore, additional research is urgently needed to
tion in menopause. Molecular and Cellular Endocrinology (2014), http://
L. Raz / Molecular and Cellular Endocrinology xxx (2014) xxx–xxx 7
investigate the optimal dose, timing, duration and route ofadministration for MHT in light of individual genetic differences.Cumulative evidence in the field suggests that E2 may exertpositive or negative effects on vascular health, depending on thespecific circumstance.
Author contributions
Dr. Raz: collection of literature, conceptualization, interpreta-tion of the data, drafting the manuscript.
Acknowledgements
The author would like to thank Dr. Virginia Miller, Dr. VictorHenderson and Dr. Gary Rosenberg for their contributions andguidance during the revisions process.
References
Alkayed, N.J., Murphy, S.J., Traystman, R.J., Hurn, P.D., Miller, V.M., 2000.Neuroprotective effects of female gonadal steroids in reproductivelysenescent female rats. Stroke; J. Cereb. Circ. 31 (1), 161–168.
Alqahtani, S., Coffin, C.S., Burak, K., Chen, F., MacGregor, J., Beck, P., 2007. Hepaticportal venous gas: a report of two cases and a review of the epidemiology,pathogenesis, diagnosis and approach to management. Can. J. gastroenterol = J.canadien de gastroenterologie. 21 (5), 309–313.
Anderson, G.L., Limacher, M., Assaf, A.R., et al., 2004. Effects of conjugated equineestrogen in postmenopausal women with hysterectomy: the Women’s HealthInitiative randomized controlled trial. JAMA, J. Am. Med. Assoc. 291 (14), 1701–1712.
Ardelt, A.A., Carpenter, R.S., Lobo, M.R., et al., 2012. Estradiol modulates post-ischemic cerebral vascular remodeling and improves long-term functionaloutcome in a rat model of stroke. Brain Res. 1461, 76–86.
Babiker, F.A., De Windt, L.J., van Eickels, M., Grohe, C., Meyer, R., Doevendans, P.A.,2002. Estrogenic hormone action in the heart: regulatory network and function.Cardiovasc. Res. 53 (3), 709–719.
Bake, S., Sohrabji, F., 2004. 17beta-estradiol differentially regulates blood–brainbarrier permeability in young and aging female rats. Endocrinology 145 (12),5471–5475.
Balthazart, J., Ball, G.F., 2006. Is brain estradiol a hormone or a neurotransmitter?Trends Neurosci. 29 (5), 241–249.
Bhavnani, B.R., Cecutti, A., Gerulath, A., 1998. Pharmacokinetics andpharmacodynamics of a novel estrogen delta8-estrone in postmenopausalwomen and men. J. Steroid. Biochem. Mol. Biol. 67 (2), 119–131.
Brackley, K.J., Ramsay, M.M., Broughton Pipkin, F., Rubin, PC., 1999. The effect of themenstrual cycle on human cerebral blood flow: studies using Dopplerultrasound. Ultrasound Obs. Gynecol.: Off. J. Int. Soc. Ultrasound Obstet.Gynecol. 14 (1), 52–57.
Brann, D.W., Dhandapani, K., Wakade, C., Mahesh, V.B., Khan, M.M., 2007.Neurotrophic and neuroprotective actions of estrogen: basic mechanisms andclinical implications. Steroids 72 (5), 381–405.
Brinton, E., Nanjee, N., Hopkins, P., et al., 2012. Oral estrogen vs. transdermalestrogen effects on lipids and lipoproteins in recently menopausal women: thekronos early estrogen prevention study (KEEPS). North American MenopauseSociety Meeting, p. 88.
Brunsing, R.L., Prossnitz, E.R., 2011. Induction of interleukin-10 in the T helper type17 effector population by the G protein coupled estrogen receptor (GPER)agonist G-1. Immunology 134 (1), 93–106.
Bryan Jr., R.M., You, J., Golding, E.M., Marrelli, S.P., 2005. Endothelium-derivedhyperpolarizing factor: a cousin to nitric oxide and prostacyclin. Anesthesiology102 (6), 1261–1277.
Bushnell, C.D., Hurn, P., Colton, C., et al., 2006. Advancing the study of stroke inwomen: summary and recommendations for future research from an NINDS-Sponsored Multidisciplinary Working Group. Stroke; J. Cereb. Circ. 37 (9), 2387–2399.
Canonico, M., Plu-Bureau, G., Lowe, G.D., Scarabin, P.Y., 2008. Hormone replacementtherapy and risk of venous thromboembolism in postmenopausal women:systematic review and meta-analysis. BMJ 336 (7655), 1227–1231.
Chakrabarti, S., Lekontseva, O., Davidge, S.T., 2008. Estrogen is a modulator ofvascular inflammation. IUBMB Life 60 (6), 376–382.
Chen, S., Nilsen, J., Brinton, R.D., 2006. Dose and temporal pattern of estrogenexposure determines neuroprotective outcome in hippocampal neurons:therapeutic implications. Endocrinology 147 (11), 5303–5313.
Chilvers, C.E., Knibb, R.C., Armstrong, S.J., et al., 2003. Post menopausal hormonereplacement therapy and risk of acute myocardial infarction-a case controlstudy of women in the East UK. Eur Heart J. 24 (24), 2197–2205.
Cipolla, M.J., Li, R., Vitullo, L., 2004. Perivascular innervation of penetrating brainparenchymal arterioles. J. Cardiovasc. Pharmacol. 44 (1), 1–8.
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
Cipolla, M.J., Godfrey, J.A., Wiegman, M.J., 2009. The effect of ovariectomy andestrogen on penetrating brain arterioles and blood–brain barrier permeability.Microcirculation 16 (8), 685–693.
Clark, J.H., 2006. A critique of Women’s Health Initiative Studies (2002–2006). Nucl.Receptor Signaling 4, e023.
Closa, D., Folch-Puy, E., 2004. Oxygen free radicals and the systemic inflammatoryresponse. IUBMB Life 56 (4), 185–191.
Cotter, G., Cannon, C.P., McCabe, C.H., et al., 2003. Prior peripheral arterial diseaseand cerebrovascular disease are independent predictors of adverse outcome inpatients with acute coronary syndromes: are we doing enough? Results fromthe orbofiban in patients with unstable coronary syndromes-thrombolysis inmyocardial infarction (OPUS-TIMI) 16 study. Am. Heart J. 145 (4), 622–627.
Dan, P., Cheung, J.C., Scriven, D.R., Moore, E.D., 2003. Epitope-dependent localizationof estrogen receptor-alpha, but not -beta, in en face arterial endothelium. Am. J.Physiol. Heart Circ. Physiol. 284 (4), H1295–H1306.
de Leeuw, F.E., de Groot, J.C., Bots, M.L., et al., 2000. Carotid atherosclerosis andcerebral white matter lesions in a population based magnetic resonanceimaging study. J. Neurol. 247 (4), 291–296.
de Leeuw, F.E., de Groot, J.C., Achten, E., et al., 2001. Prevalence of cerebral whitematter lesions in elderly people: a population based magnetic resonanceimaging study. The Rotterdam Scan Study. J. Neurol., Neurosurg. psychiatr. 70(1), 9–14.
Debette, S., Seshadri, S., Beiser, A., et al., 2011. Midlife vascular risk factor exposureaccelerates structural brain aging and cognitive decline. Neurology 77 (5), 461–468.
Diomedi, M., Cupini, L.M., Rizzato, B., Ferrante, F., Giacomini, P., Silvestrini, M., 2001.Influence of physiologic oscillation of estrogens on cerebral hemodynamics. J.Neurol. Sci. 185 (1), 49–53.
Drake, C.T., Iadecola, C., 2007. The role of neuronal signaling in controlling cerebralblood flow. Brain Lang. 102 (2), 141–152.
Dubal, D.B., Wise, P.M., 2001. Neuroprotective effects of estradiol in middle-agedfemale rats. Endocrinology 142 (1), 43–48.
Duckles, S.P., Krause, D.N., 2011. Mechanisms of cerebrovascular protection:oestrogen, inflammation and mitochondria. Acta Physiol. 203 (1), 149–154.
Enciu, A.M., Gherghiceanu, M., Popescu, B.O., 2013. Triggers and effectors ofoxidative stress at blood–brain barrier level: relevance for brain ageing andneurodegeneration. Oxidative Med. Cell longevity 2013, 297512.
Faraci, F.M., Heistad, D.D., 1998. Regulation of the cerebral circulation: role ofendothelium and potassium channels. Physiol. Rev. 78 (1), 53–97.
Galea, E., Santizo, R., Feinstein, D.L., et al., 2002. Estrogen inhibits NF kappa B-dependent inflammation in brain endothelium without interfering with I kappaB degradation. NeuroReport 13 (11), 1469–1472.
Geary, G.G., Krause, D.N., Duckles, S.P., 1998. Estrogen reduces myogenic tonethrough a nitric oxide-dependent mechanism in rat cerebral arteries. Am. J.Physiol. 275 (1 Pt 2), H292–H300.
Geary, G.G., Krause, D.N., Duckles, S.P., 2000. Estrogen reduces mouse cerebralartery tone through endothelial NOS- and cyclooxygenase-dependentmechanisms. Am. J. Physiol. Heart Circ. Physiol. 279 (2), H511–H519.
Gibbs, R.B., 2000. Long-term treatment with estrogen and progesterone enhancesacquisition of a spatial memory task by ovariectomized aged rats. Neurobiol.Aging 21 (1), 107–116.
Golding, E.M., Ferens, D.M., Marrelli, S.P., 2002. Altered calcium dynamics do notaccount for attenuation of endothelium-derived hyperpolarizing factor-mediated dilations in the female middle cerebral artery. Stroke; J. Cereb. Circ.33 (12), 2972–2977.
Goodman, M.P., 2012. Are all estrogens created equal? A review of oral vs.transdermal therapy. J. Women’s Health 21 (2), 161–169.
Goodrow, G.J., Vitullo, L., Cipolla, M.J., 2005. Effect of estrogen therapy on cerebralarteries during stroke in female rats. Menopause 12 (1), 99–109.
Gordon, G.R., Mulligan, S.J., MacVicar, B.A., 2007. Astrocyte control of thecerebrovasculature. Glia 55 (12), 1214–1221.
Harman, S.M., Vittinghoff, E., Brinton, E.A., et al., 2011. Timing and duration ofmenopausal hormone treatment may affect cardiovascular outcomes. Am. J.Med. 124 (3), 199–205.
Hawkins, B.T., Davis, T.P., 2005. The blood–brain barrier/neurovascular unit inhealth and disease. Pharmacol. Rev. 57 (2), 173–185.
He, Z., He, Y.J., Day, A.L., Simpkins, J.W., 2002. Proestrus levels of estradiol duringtransient global cerebral ischemia improves the histological outcome of thehippocampal CA1 region: perfusion-dependent and-independent mechanisms.J. Neurol. Sci. 193 (2), 79–87.
Heiss, G., Wallace, R., Anderson, G.L., et al., 2008. Health risks and benefits 3 yearsafter stopping randomized treatment with estrogen and progestin. JAMA, J. Am.Med. Assoc. 299 (9), 1036–1045.
Henderson, V.W., Lobo, R.A., 2012. Hormone therapy and the risk of stroke:perspectives 10 years after the Women’s Health Initiative trials. Climacteric: J.Int. Menopause Soc. 15 (3), 229–234.
Hendrix, S.L., Wassertheil-Smoller, S., Johnson, K.C., et al., 2006. Effects ofconjugated equine estrogen on stroke in the Women’s Health Initiative.Circulation 113 (20), 2425–2434.
tion in menopause. Molecular and Cellular Endocrinology (2014), http://
8 L. Raz / Molecular and Cellular Endocrinology xxx (2014) xxx–xxx
Hodges, Y.K., Tung, L., Yan, X.D., Graham, J.D., Horwitz, K.B., Horwitz, L.D., 2000.Estrogen receptors alpha and beta: prevalence of estrogen receptor beta mRNAin human vascular smooth muscle and transcriptional effects. Circulation 101(15), 1792–1798.
Hojo, Y., Hattori, T.A., Enami, T., et al., 2004. Adult male rat hippocampussynthesizes estradiol from pregnenolone by cytochromes P45017alpha andP450 aromatase localized in neurons. Proc. Nat. Acad. Sci. USA 101 (3), 865–870.
Holschneider, D.P., Scremin, O.U., 1998. Effects of ovariectomy on cerebral flow ofrats. Neuroendocrinology 67 (4), 260–268.
Hulley, S., Grady, D., Bush, T., et al., 1998. Randomized trial of estrogen plusprogestin for secondary prevention of coronary heart disease inpostmenopausal women. Heart Estrogen/Progestin Replacement Study (HERS)Research Group. JAMA: The J. Am. Med. Assoc. 280 (7), 605–613.
Irwin, R.W., Yao, J., To, J., Hamilton, R.T., Cadenas, E., Brinton, R.D., 2012. Selectiveoestrogen receptor modulators differentially potentiate brain mitochondrialfunction. J. Neuroendocrinol. 24 (1), 236–248.
Karas, R.H., Hodgin, J.B., Kwoun, M., et al., 1999. Estrogen inhibits the vascular injuryresponse in estrogen receptor beta-deficient female mice. Proc. Nat. Acad. Sci.USA 96 (26), 15133–15136.
Kemper, M.F., Zhao, Y., Duckles, S.P., Krause, D.N., 2013. Endogenous ovarianhormones affect mitochondrial efficiency in cerebral endothelium via distinctregulation of PGC-1 isoforms. J. Cerebr. Blood Flow Metab: Off. J. Int. Soc. Cereb.Blood Flow Metab. 33 (1), 122–128.
Kenealy, B.P., Kapoor, A., Guerriero, K.A., et al., 2013. Neuroestradiol in thehypothalamus contributes to the regulation of gonadotropin releasinghormone release. J. Neurosci.: Off. J. Soc. Neurosci. 33 (49), 19051–19059.
Krause, D.N., Duckles, S.P., Gonzales, R.J., 2011. Local oestrogenic/androgenicbalance in the cerebral vasculature. Acta Physiol. 203 (1), 181–186.
Krejza, J., Mariak, Z., Huba, M., Wolczynski, S., Lewko, J., 2001. Effect of endogenousestrogen on blood flow through carotid arteries. Stroke; J. Cereb. Circ. 32 (1),30–36.
Krejza, J., Mariak, Z., Nowacka, A., Melhem, E.R., Babikian, V.L., 2004. Influence of 17-beta-estradiol on cerebrovascular impedance during menstrual cycle in women.J. Neurol. Sci. 221 (1–2), 61–67.
Kuller, L.H., Margolis, K.L., Gaussoin, S.A., et al., 2010. Relationship of hypertension,blood pressure, and blood pressure control with white matter abnormalities inthe Women’s Health Initiative Memory Study (WHIMS)-MRI trial. J. Clin.Hypertens. 12 (3), 203–212.
LaCroix, A.Z., Chlebowski, R.T., Manson, J.E., et al., 2011. Health outcomes afterstopping conjugated equine estrogens among postmenopausal women withprior hysterectomy: a randomized controlled trial. JAMA, J. Am. Med. Assoc. 305(13), 1305–1314.
Lindner, V., Kim, S.K., Karas, R.H., Kuiper, G.G., Gustafsson, J.A., Mendelsohn, M.E.,1998. Increased expression of estrogen receptor-beta mRNA in male bloodvessels after vascular injury. Circ. Res. 83 (2), 224–229.
Luscher, T.F., Vanhoutte, P.M., 1986. Endothelium-dependent responses to plateletsand serotonin in spontaneously hypertensive rats. Hypertension 8 (6 Pt 2), II55-60.
Madamanchi, N.R., Vendrov, A., Runge, M.S., 2005. Oxidative stress and vasculardisease. Arterioscler. Thromb. Vasc. Biol. 25 (1), 29–38.
Maki, P.M., 2006. Hormone therapy and cognitive function: is there a critical periodfor benefit? Neuroscience 138 (3), 1027–1030.
Maki, P.M., Resnick, S.M., 2001. Effects of estrogen on patterns of brain activity atrest and during cognitive activity: a review of neuroimaging studies.NeuroImage 14 (4), 789–801.
Marriott, L.K., Hauss-Wegrzyniak, B., Benton, R.S., Vraniak, P.D., Wenk, G.L., 2002.Long-term estrogen therapy worsens the behavioral and neuropathologicalconsequences of chronic brain inflammation. Behav. Neurosci. 116 (5), 902–911.
McCullough, L.D., Blizzard, K., Simpson, E.R., Oz, O.K., Hurn, P.D., 2003. Aromatasecytochrome P450 and extragonadal estrogen play a role in ischemicneuroprotection. J Neurosci.: Off. J. Soc. Neurosci. 23 (25), 8701–8705.
McEwen, B.S., Akama, K.T., Spencer-Segal, J.L., Milner, T.A., Waters, E.M., 2012.Estrogen effects on the brain: actions beyond the hypothalamus via novelmechanisms. Behav. Neurosci. 126 (1), 4–16.
Moody, D.M., Bell, M.A., Challa, V.R., 1990. Features of the cerebral vascular patternthat predict vulnerability to perfusion or oxygenation deficiency: an anatomicstudy. AJNR Am. J. Neuroradiol. 11 (3), 431–439.
Moody, D.M., Santamore, W.P., Bell, M.A., 1991. Does tortuosity in cerebralarterioles impair down-autoregulation in hypertensives and elderly
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
normotensives? A hypothesis and computer model. Clin. Neurosurg. 37, 372–387.
Mountain, D.J., Freeman, M.B., Kirkpatrick, S.S., et al., 2013. Effect of hormonereplacement therapy in matrix metalloproteinase expression and intimalhyperplasia development after vascular injury. Ann. Vasc. Surg. 27 (3), 337–345.
Mukherjee, T.K., Reynolds, P.R., Hoidal, J.R., 2005. Differential effect of estrogenreceptor alpha and beta agonists on the receptor for advanced glycation endproduct expression in human microvascular endothelial cells. Biochim. Biophys.Acta 1745 (3), 300–309.
Naftolin, F., Taylor, H.S., Karas, R., et al., 2004. The Women’s Health Initiative couldnot have detected cardioprotective effects of starting hormone therapy duringthe menopausal transition. Fertil. Steril. 81 (6), 1498–1501.
Nakatani, Y., Sato-Suzuki, I., Tsujino, N., et al., 2008. Augmented brain 5-HT crossesthe blood–brain barrier through the 5-HT transporter in rat. Eur. J. Neurosci. 27(9), 2466–2472.
Nevo, O., Soustiel, J.F., Thaler, I., 2007. Cerebral blood flow is increased duringcontrolled ovarian stimulation. Am. J. Physiol. Heart Circ. Physiol. 293 (6),H3265–H3269.
North American Menopause, S., 2012. The 2012 hormone therapy positionstatement of: the north american menopause society. Menopause 19 (3),257–271.
Ospina, J.A., Duckles, S.P., Krause, D.N., 2003. 17beta-estradiol decreases vasculartone in cerebral arteries by shifting COX-dependent vasoconstriction tovasodilation. Am. J. Physiol. Heart Circ. Physiol. 285 (1), H241–H250.
Ospina, J.A., Brevig, H.N., Krause, D.N., Duckles, S.P., 2004. Estrogen suppresses IL-1beta-mediated induction of COX-2 pathway in rat cerebral blood vessels. Am.J. Physiol. Heart Circ. Physiol. 286 (5), H2010–H2019.
Park, K.E., Pepine, C.J., 2010. Pathophysiologic mechanisms linking impairedcardiovascular health and neurologic dysfunction: the year in review. Clevel.Clin. J. Med. 77 (Suppl 3), S40–S45.
Pelligrino, D.A., Galea, E., 2001. Estrogen and cerebrovascular physiology andpathophysiology. Jpn. J. Pharmacol. 86 (2), 137–158.
Punnonen, R., Lukola, A., 1982. High-affinity binding of estrone, estradiol and estriolin human cervical myometrium and cervical and vaginal epithelium. J.Endocrinol. Invest. 5 (4), 203–207.
Raz, N., Yang, Y., Dahle, C.L., Land, S., 2012. Volume of white matter hyperintensitiesin healthy adults: contribution of age, vascular risk factors, and inflammation-related genetic variants. Biochim. Biophys. Acta 1822 (3), 361–369.
Raz, L., Jayachandran, M., Tosakulwong, N., et al., 2013. Thrombogenic microvesiclesand white matter hyperintensities in postmenopausal women. Neurology 80(10), 911–918.
Razmara, A., Duckles, S.P., Krause, D.N., Procaccio, V., 2007. Estrogen suppressesbrain mitochondrial oxidative stress in female and male rats. Brain Res. 1176,71–81.
Razmara, A., Sunday, L., Stirone, C., et al., 2008. Mitochondrial effects of estrogen aremediated by estrogen receptor alpha in brain endothelial cells. J. Pharmacol.Exp. Ther. 325 (3), 782–790.
Renoux, C., Dell’aniello, S., Garbe, E., Suissa, S., 2010. Transdermal and oral hormonereplacement therapy and the risk of stroke: a nested case-control study. BMJ340, c2519.
Rettberg, J.R., Yao, J., Brinton, R.D., 2013. Estrogen: a master regulator ofbioenergetic systems in the brain and body. Front. Neuroendocrinol. Aug 29.
Ritzel, R.M., Capozzi, L.A., McCullough, L.D., 2013. Sex, stroke, and inflammation: thepotential for estrogen-mediated immunoprotection in stroke. Horm. Behav. 63(2), 238–253.
Rocca, W.A., Grossardt, B.R., Shuster, L.T., 2011. Oophorectomy, menopause,estrogen treatment, and cognitive aging: clinical evidence for a window ofopportunity. Brain Res. 1379, 188–198.
Rosano, G.M., Vitale, C., Silvestri, A., Fini, M., 2003. Hormone replacement therapyand cardioprotection: the end of the tale? Ann. N. Y. Acad. Sci. 997, 351–357.
Rosenberg, G.A., 2012. Neurological diseases in relation to the blood–brain barrier. J.Cerebr. Blood Flow Metab: Off. J. Int. Soc. Cereb. Blood Flow Metab. 32 (7),1139–1151.
Rossouw, J.E., Anderson, G.L., Prentice, R.L., et al., 2002. Risks and benefits ofestrogen plus progestin in healthy postmenopausal women: principal resultsFrom the Women’s Health Initiative randomized controlled trial. JAMA, J. Am.Med. Assoc. 288 (3), 321–333.
Rusa, R., Alkayed, N.J., Crain, B.J., et al., 1999. 17beta-estradiol reduces stroke injuryin estrogen-deficient female animals. Stroke; J. Cereb. Circ. 30 (8), 1665–1670.
Saldanha, C.J., Remage-Healey, L., Schlinger, B.A., 2011. Synaptocrine signaling:steroid synthesis and action at the synapse. Endocr. Rev. 32 (4), 532–549.
Salom, J.B., Burguete, M.C., Perez-Asensio, F.J., Torregrosa, G., Alborch, E., 2001.Relaxant effects of 17-beta-estradiol in cerebral arteries through Ca(2+) entryinhibition. J. Cerebr. Blood Flow Metab: Off. J. Int. Soc. Cereb. Blood Flow Metab.21 (4), 422–429.
Santen, R.J., Allred, D.C., Ardoin, S.P., et al., 2010. Postmenopausal hormone therapy:an Endocrine Society scientific statement. J. Clin. Endocrinol. Metab.95 (7 Suppl 1), s1–s66.
tion in menopause. Molecular and Cellular Endocrinology (2014), http://
Shumaker, S.A., Legault, C., Kuller, L., et al., 2004. Conjugated equine estrogens andincidence of probable dementia and mild cognitive impairment inpostmenopausal women: Women’s Health Initiative Memory Study. JAMA, J.Am. Med. Assoc. 291 (24), 2947–2958.
Simon, J.A., Hsia, J., Cauley, J.A., et al., 2001. Postmenopausal hormone therapy andrisk of stroke: The Heart and Estrogen-progestin Replacement Study (HERS).Circulation 103 (5), 638–642.
Simpson, E.R., Davis, S.R., 2001. Minireview: aromatase and the regulation ofestrogen biosynthesis–some new perspectives. Endocrinology 142 (11), 4589–4594.
Straub, R.H., 2007. The complex role of estrogens in inflammation. Endocr. Rev. 28(5), 521–574.
Strehlow, K., Werner, N., Berweiler, J., et al., 2003. Estrogen increases bone marrow-derived endothelial progenitor cell production and diminishes neointimaformation. Circulation 107 (24), 3059–3065.
Sumino, H., Murakami, M., 2013. Investigation of atherosclerosis in postmenopausalwomen: alteration of atherosclerosis-associated factors and vascularatherosclerosis by oral and transdermal estrogen replacement. Rinsho byori.Jpn. J. Clin. Pathol. 61 (3), 256–262.
Sunday, L., Osuna, C., Krause, D.N., Duckles, S.P., 2007. Age alters cerebrovascularinflammation and effects of estrogen. Am. J. Physiol. Heart Circ. Physiol. 292 (5),H2333–H2340.
Suzuki, S., Brown, C.M., Dela Cruz, CD., Yang, E., Bridwell, DA., Wise, PM., 2007.Timing of estrogen therapy after ovariectomy dictates the efficacy of its
Please cite this article in press as: Raz, L. Estrogen and cerebrovascular reguladx.doi.org/10.1016/j.mce.2014.01.015
neuroprotective and antiinflammatory actions. Proc. Nat. Acad. Sci. USA 104(14), 6013–6018.
Taylor, H.S., Manson, J.E., 2011. Update in hormone therapy use in menopause. J.Clin. Endocrinol. Metab. 96 (2), 255–264.
Taylor, S.G., Weston, A.H., 1988. Endothelium-derived hyperpolarizing factor: a newendogenous inhibitor from the vascular endothelium. Trends Pharmacol. Sci. 9(8), 272–274.
Trevoux, R., De Brux, J., Castanier, M., Nahoul, K., Soule, J.P., Scholler, R., 1986.Endometrium and plasma hormone profile in the peri-menopause and post-menopause. Maturitas 8 (4), 309–326.
Tu, J., Jufri, N.F., 2013. Estrogen Signaling through estrogen receptor beta andG-protein-coupled estrogen receptor 1 in human cerebral vascularendothelial cells: implications for cerebral aneurysms. BioMed Res. Int.2013, 524324.
van Beek, A.H., Claassen, J.A., Rikkert, M.G., Jansen, R.W., 2008. Cerebralautoregulation: an overview of current concepts and methodology withspecial focus on the elderly. J. Cerebr. Blood Flow Metab: Off. J. Int. Soc.Cereb. Blood Flow Metab. 28 (6), 1071–1085.
van Dijk, E.J., Prins, N.D., Vrooman, H.A., Hofman, A., Koudstaal, P.J., Breteler, M.M.,2008. Progression of cerebral small vessel disease in relation to risk factors andcognitive consequences: Rotterdam Scan study. Stroke; J. Cereb. Circ. 39 (10),2712–2719.
Vickers, M.R., MacLeannan, A.H., Lawton, B., et al., 2007. Main morbidities recordedin women’s international study of long duration oestrogen after menopause(WISDOM): A randomized controlled trial of hormone replacement therapy inpostmenopausal women. BMJ. 335 (7613), 239.
Viscoli, C.M., Brass, L.M., Kernan, W.N., Sarrel, P.M., Suissa, S., Horwitz, R.I., 2001. Aclinical trial of estrogen-replacement therapy after ischemic stroke. New Engl. J.Med. 345 (17), 1243–1249.
Wardlaw, J.M., Smith, E.E., Biessels, G.J., et al., 2013. Neuroimaging standards forresearch into small vessel disease and its contribution to ageing andneurodegeneration. Lancet Neurol. 12 (8), 822–838.
Wassertheil-Smoller, S., Hendrix, S.L., Limacher, M., et al., 2003. Effect of estrogenplus progestin on stroke in postmenopausal women: the Women’s HealthInitiative: a randomized trial. JAMA, J. Am. Med. Assoc. 289 (20), 2673–2684.
Weiner, C.P., Lizasoain, I., Baylis, S.A., Knowles, R.G., Charles, I.G., Moncada, S., 1994.Induction of calcium-dependent nitric oxide synthases by sex hormones. Proc.Nat. Acad. Sci. USA 91 (11), 5212–5216.
Wharton, W., Dowling, M., Khosropour, C.M., Carlsson, C., Asthana, S., Gleason, C.E.,2009. Cognitive benefits of hormone therapy: cardiovascular factors andhealthy-user bias. Maturitas 64 (3), 182–187.
Yang, S.H., Liu, R., Perez, E.J., Wang, X., Simpkins, J.W., 2005. Estrogens as protectantsof the neurovascular unit against ischemic stroke. Curr. Drug Targets CNSNeurol. Disord. 4 (2), 169–177.
Yu, E., Mercer, J., Bennett, M., 2012. Mitochondria in vascular disease. Cardiovasc.Res. 95 (2), 173–182.
Yubero-Lahoz, S., Robledo, P., Farre, M., de laTorre, R., 2013. Platelet SERT as aperipheral biomarker of serotonergic neurotransmission in the central nervoussystem. Curr. Med. Chem. 20 (11), 1382–1396.
Zhang, Q.G., Raz, L., Wang, R., et al., 2009. Estrogen attenuates ischemic oxidativedamage via an estrogen receptor alpha-mediated inhibition of NADPH oxidaseactivation. J. Neurosci.: Off. J. Soc. Neurosci. 29 (44), 13823–13836.
Zhang, Q.G., Han, D., Wang, R.M., et al., 2011. C terminus of Hsc70-interactingprotein (CHIP)-mediated degradation of hippocampal estrogen receptor-alphaand the critical period hypothesis of estrogen neuroprotection. Proc. Nat. Acad.Sci. USA 108 (35), E617–E624.
tion in menopause. Molecular and Cellular Endocrinology (2014), http://