ELSEVIER Biochimica et Biophysica Acta 1310 (1996) 317-324
Biochi~ic~a et Biophysica A~ta
Opposite regulation of IGF-I and IGF-I receptor mRNA and concomitant changes of GH receptor and IGF-II/M6P receptor mRNA in human
IM-9 lymphoblasts 1
Y. Yang b, A. Hoeflich a,2, O. Butenandt a, W. Kiess a,. a Children's Hospital, University ofM~nchen, Lindwurmstrasse 4, D-80337 Munich, Germany
b Children's Hospital, Shanghai Medical University, Feng Lin Road, ShanghaL People's Republic of China
Received 14 June 1995; revised 7 November 1995; accepted 24 November 1995
Abstract
Human IM-9 lymphoblasts synthesize IGF-I and express IGF-I receptors, IGF-II/M6P receptors and GH receptors. We have studied the regulation of mRNA expression of IGF-I, IGF-I receptors, IGF-II/M6P receptors and GH receptors in IM-9 cells upon serum-withdrawal and re-addition of serum. IM-9 cells were cultured in RPMI-1640 medium with or without serum for various periods of time. RNA was prepared using guanidinium thiocyanate and CsC1. Antisense riboprobes for human IGF-I, IGF-I receptor, IGF-II/M6P receptor, GH receptor and for comparison for human beta-actin were synthesized and labeled with 32p. Protected fragments of 379 bases and of 420 and 350 bases with the IGF-I receptor and with the IGF-I probe respectively and protected fragments of 670 bases and of 51 and 121 bases with the GH receptor and with the beta-actin probe were detected. For the human IGF-II/M6P receptor probe protected fragments of 260 bases were visualized in RNA samples. The amount of mRNA present in each lane (10/zg total RNA) was determined by computed densitometry. The amount of IGF-I mRNA expressed by IM-9 cells decreased rapidly (within two hours) and dramatically (more than 120%) after the withdrawal of serum and increased significantly (220%) after the re-addition of serum. This increase of IGF-I mRNA preceded the increase: in cell number that was seen after 48 h of medium change. Conversely, the expression of IGF-I receptor mRNA and beta-actin mRNA increased by more than 250% after the withdrawal of serum within 2 and 8 h respectively, while GH receptor mRNA fell within 2-4 h. The expression of IGF-II/M6P receptor mRNA continued to increase throughout the duration of the cell culture experiment. We conclude that IGF-I and IGF-I receptor mRNAs are regulated in an opposite direction in serum-deprived IM-9 lymphoblasts. In addition, GH receptor mRNA expression parallels IGF-I mRNA expression.
The growth hormone-insulin-l ike growth factor axis plays a key role in the hormonal regulation of growth and cell proliferation [1-3]. GH binds to a high affinity cell surface receptor that signals its biologic function through a phosphorylation cascade involving a family of tyrosine
kinases ( JAK kinases). GH receptors are mainly expressed in human liver but are also found in many other tissues [4]. The insulin-like growth factors ( IGF-I and IGF-II) are polypeptides which share homology with insulin in respect to amino acid sequence and protein structure. The IGFs exert insulin-like metabolic effects in vitro and in vivo. The IGFs are potent mitogens for cells in culture. IGF-I mediates most of the anabolic effects of pituitary growth hormone, whereas IGF-I I is thought to play an important role as a fetal growth factor. Recent experimental evidence also suggests that IGF-II might be an important regulator of the lysosomal degradation pathway [1,2]. The IGFs bind to at least two classes of high affinity receptors that are present on many cells and tissues. The IGF-I receptor is a heterotetramer that closely resembles the insulin receptor and mediates most if not all of the IGF-induced biologic
318 Y. Yang et al. / Biochimica et Biophysica Acta 1310 (1996) 317-324
effects. The IGF-I receptor is a tyrosine kinase that signals its biologic response through a cascade of phosphorylation events that include phosphorylation of insulin-responsive substrate-1 (IRS-I), phosphatidyl-inositol-3-kinase (PI3- kinase) and mitogen-activated protein kinases (MAP ki- nases) [5,6]. The IGF-II/M6P receptor is a bifunctional binding protein that binds both IGF-II and mannose-6- phosphate bearing lysosmal enzymes at distinct binding sites. The two classes of ligands that bind to the IGF- I I /M6P receptor reciprocally inhibit binding of the other class of ligand [7-11 ].
Little is known as to the regulation of GH receptor and IGF receptor expression in vivo and in vitro. While insulin and developmental factors seem to be the major regulators of IGF-II/M6P receptor expression [9], regulation of IGF-I receptor expression is poorly understood. GH and IGF-I itself have been implicated in the regulation of IGF-I receptor protein expression in cells in culture [12-14]. It has also been suggested that IGF-I and its receptor are regulated in an opposite way [15-18]. There are no studies that report upon the concomitant regulation of IGF-I and the IGF receptors either in vivo or in vitro.
We have used the IM-9 lymphoblast cell line to exam- ine the regulation of IGF-I and of IGF-I receptor gene expression during proliferative phases of human cells [3]. For comparison we have also measured the presence of beta-actin RNA, growth hormone receptor RNA and IGF- I I /M6P receptor RNA in human IM-9 lymphoblasts dur- ing serum-deprivation and after serum re-addition to cell cultures. The results obtained suggest that the GH receptor may be involved in regulating IGF-I production by IM-9 cells. In addition, our data give evidence as to a putative compensatory up-regulatory mechanism involving the IGF-I receptor after serum withdrawal and decrease of endogenous IGF-I expression.
2. Materials and methods
2.1. Peptides and cDNA probes
The following probes were used in solution hybridisa- tion/RNase protection assays and were a kind gift from Dr. C.T. Roberts, Bethesda: a 442 bases hlGF-I probe [19], and a 379 bases (394 bases full-length probe) hlGF-I receptor probe. The hlGF-II/M6P receptor probe was 260 nucleotides in length with 250 bases of the bases comple- mentary to the IGF-II/M6P receptor mRNA. The human GH receptor DNA probe (670 bases) was obtained from Dr. Guo, Shanghai, People's Republic of China [3,20]. Recombinant human growth hormone (GH) was kindly donated by NovoNordisk, Mainz, Germany. IGF-I was kindly donated by Dr. Steinkamp, Pharmacia, Kabi Peptide Hormones, Erlangen, Germany. Interleukin-1 beta (IL-1 beta) was purchased from Sigma Chemicals, St. Louis, USA.
2.2. Cell culture
IM-9 cells were obtained from ATTC, Rockville, USA, and were grown in RPMI-1640 medium (Seromed, Berlin, Germany) containing 10% FCS at 37°C in 5% CO2. Cells were subcultured every 2-3 days. For experiments cells were washed 3 times with serum-free RPMI-1640 medium and then were kept serum-free for 2, 4, 8, 12, 24, 48, and 72 h as indicated. In a subset of experiments cells were cultured in serum-free RPMI-1640 medium with or with- out hGH (100 ng/ml), or IGF-I (50 ng/ml) or IL-1 beta (5 U/ml). Where indicated cells were re-exposed to serum-containing medium (10% FCS) after they had been cultured in serum-free medium for 24 h for the indicated time periods.
2.3. Cell proliferation
Viable cells were identified and counted after the indi- cated times in culture after Trypan blue staining (Sigma, Heidelberg, Germany) using a hemocytometer chamber. Cell numbers are expressed as cells per ml of culture medium.
2.4. RNA extraction
Cells were harvested in 4 M guanidinium thiocyanate (Fluka, Switzerland) with 25 mM sodium acetate and 0.835% beta-mercaptoethanoi. The suspension was ho- mogenized with a polytron (Kinematika, Germany) for 15 s. The RNA was pelleted by ultra-centrifugation in 5.7 M CsC1 solution with 25 mM sodium acetate in polyallomer tubes (Beckman, Munich, Germany) in a Beckman Ti-50 rotor for 18-23 h at 24°C. The RNA pellet was dissolved in DEPC-treated water and precipitated in ethanol overnight at -20°C. The amount of RNA was calculated by mea- surement of optical density at 260 nm. The ratio of ab- sorbance at 260 nm and 280 nm was routinely greater than 1.7. The quality of the RNA was further examined by electrophoresis in 0.8% TBE-agarose, where the bands of the 28S and of the 18S rRNA showed a typical ratio and clear shape [21,22].
A pGEM-3 plasmid containing a 442 bases PstI-RsaI fragment of the human IGF-I cDNA which included an 18 bases GC tail, 89 bases of 5'UTR derived from exon 1, and 329 bases encoding the pre, B, C, A, D and part of the E domain was linearized with HindlII and the antisense riboprobe was transcribed using T7 RNA polymerase. Identity of the IGF-I cDNA probe was confirmed by partial sequencing of the probe (courtesy of Dr. J. Haenze, Molecular Biology Laboratory, Children's Hospital, Giessen, Germany). An antisense IGF-I receptor RNA probe was transcribed from a 379 bases SalI-EcoRI hu-
Y. Yang et al. / Biochimica et Biophysica A cta 1310 (1996) 317-324 319
man IGF-I receptor eDNA fragment. An antisense ribo- probe containing 250 bases complementary to the IGF- II /M6P receptor eDNA and 10 bases of PGEM-2 se- quence was generated with T7 RNA polymerase. A 670 bases EcoRI-HindIII eDNA fragment derived from the human growth hormone receptor gene, exon 10, was cloned in the PTZ 19u vector. This construct was linearized with EcoRI, and the antisense RNA was generated using T7 RNA polymerase. A 50 bases SalI-AvaI human beta-actin eDNA fragment was also transcribed. Riboprobes were synthesized and labeled u'sing the Riboprobe Gemini II Core system (Promega, Heidelberg, Germany, Cat. P1270) and 32p (Amersham, Braunschweig, Germany). Solution hybridization was performed with antisense RNA probes (approx. 200 000 cpm 32 P-UTP labeled) at 45°C overnight in a buffer containing 75% formamide, Tris 20 mM (pH 7.6), EDTA 1 mM (pH 8.0), NaCI 0.4 M, 0.1% SDS. After hybridization samples were digested with RNase (RNase A, and RNase TI, Boehringer Mannheim, Mannheim, Germany), the protected fragments were extracted with phenol-choloroform, precipitated with ethanol, and elec- trophoresed on an 8% polyacrylamide-8 M urea denaturing gel [20]. Protected radiolabeled fragments were identified by exposing the gels to Kodak X-AR films with DuPont enhancer screens at - 70°(; for 1-11 days [21,22].
2.6. Quantitation and statistical analysis
Autoradiograms of polyacrylamide gels were scanned using the Biolmage system (TF-3X, Biolmage system, USA). Density of the protected bands was measured and relative optical densities (arbitrary units) of the respective bands from at least three independent experiments (inde- pendent cell cultures) were compared using unpaired Stu- dent's t-test. Equal loading of the gels was assured by parallel analysis of RNA preparations by electrophoresis in 0.8% TBE-agarose and methylene blue staining of the gels [21,22]. In addition, the pattern of/3-actin RNA expression was analyzed in the same gels as were the RNA species of interest, fl-actin RNA expression showed a consistent and reproducible pattern also indicating equal loading of the gels.
3. Results
3. I. Cell proliferation
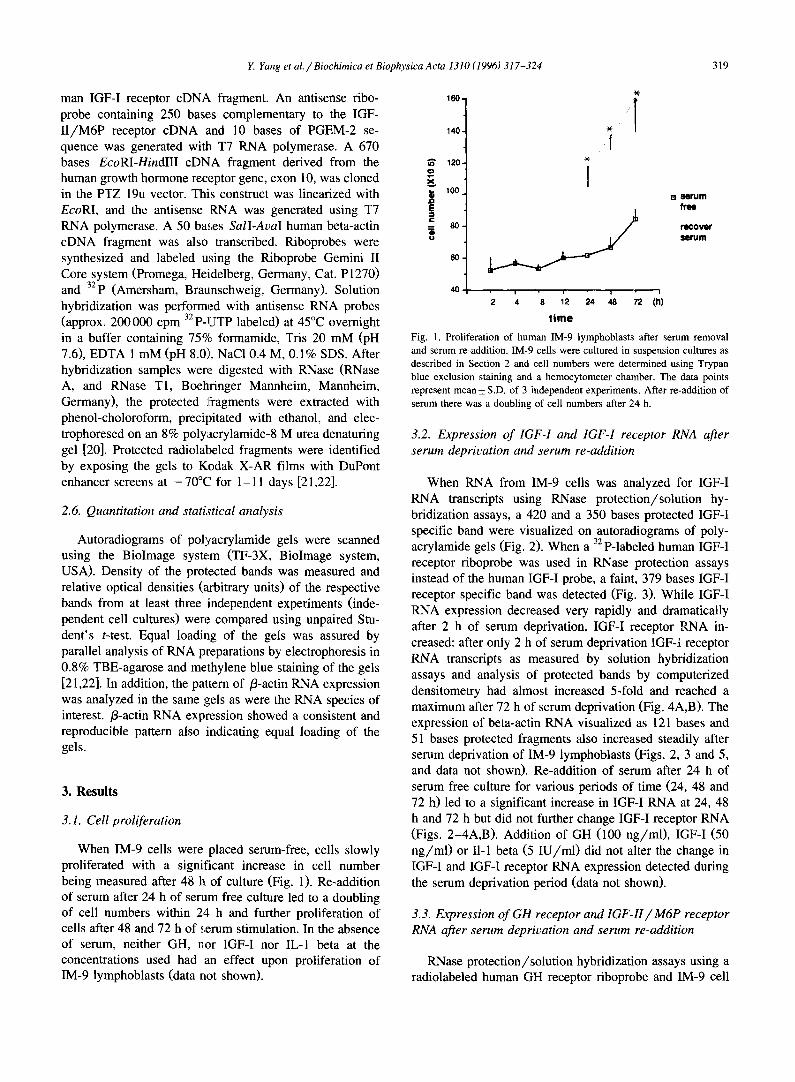
When IM-9 cells were placed serum-free, cells slowly proliferated with a significant increase in cell number being measured after 48 h of culture (Fig. 1). Re-addition of serum after 24 h of serum free culture led to a doubling of cell numbers within 24 h and further proliferation of cells after 48 and 72 h of s, erum stimulation. In the absence of serum, neither GH, nor IGF-I nor IL-I beta at the concentrations used had an effect upon proliferation of IM-9 lymphoblasts (data not shown).
160-
140
~' 120 - o
x ~oo
J~ E
serum free
r e o o v ~ r
serum
40 i ! i
2 4 8 12 24 48 72 (h)
t i m e
Fig. l. Proliferation of human IM-9 lymphoblasts after serum removal and serum re-addition. IM-9 cells were cultured in suspension cultures as described in Section 2 and cell numbers were determined using Trypan blue exclusion staining and a hemocytometer chamber. The data points represent mean + S.D. of 3 independent experiments. After re-addition of serum there was a doubling of cell numbers after 24 h.
3.2. Expression of IGF-I and IGF-I receptor RNA after serum deprivation and serum re-addition
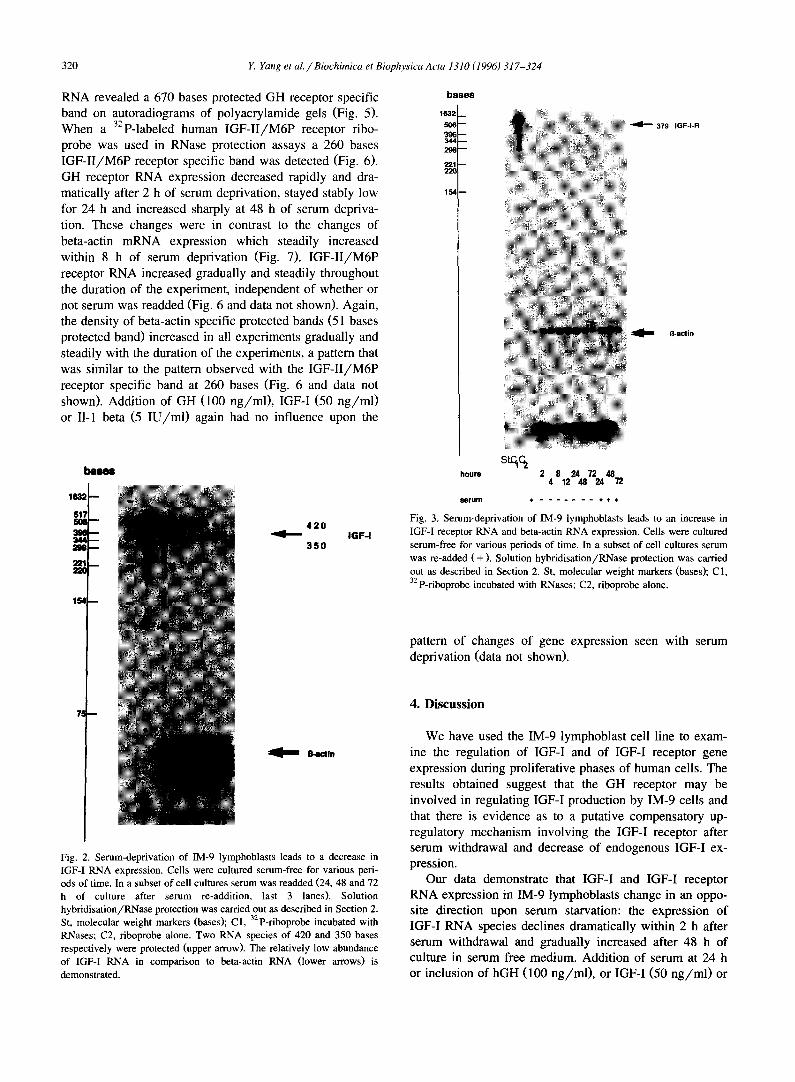
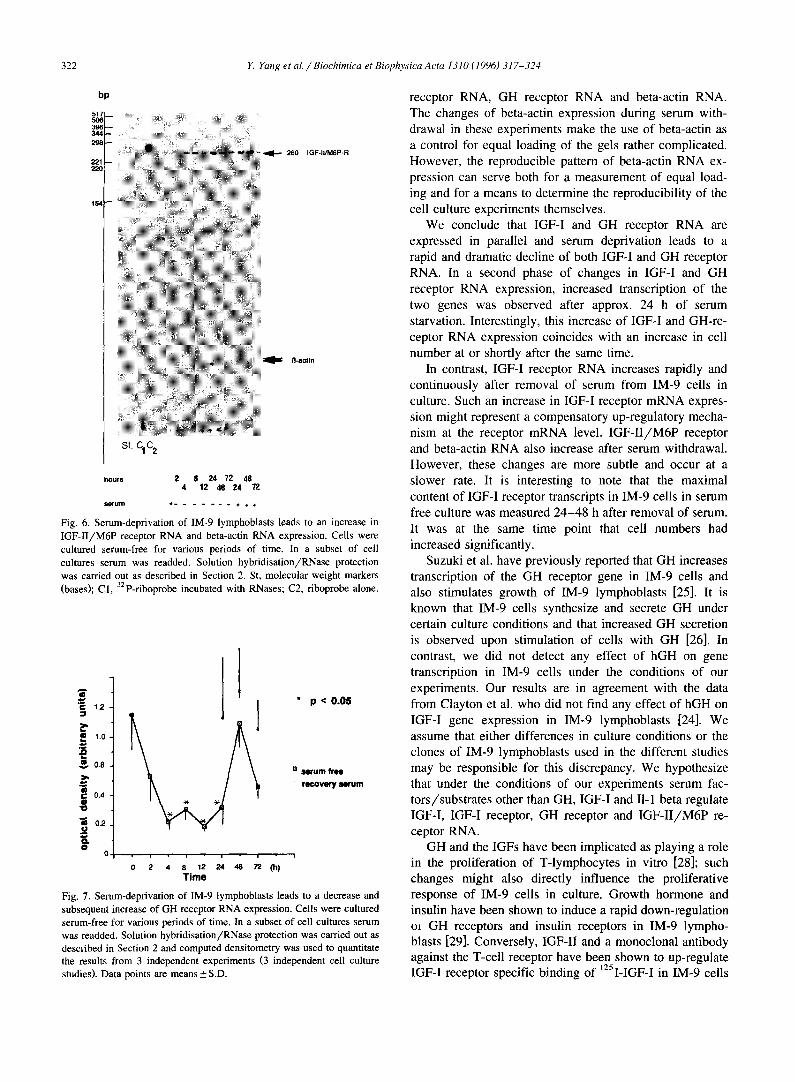
When RNA from IM-9 cells was analyzed for IGF-I RNA transcripts using RNase protection/solution hy- bridization assays, a 420 and a 350 bases protected IGF-I specific band were visualized on autoradiograms of poly- acrylamide gels (Fig. 2). When a 32 P-labeled human IGF-I receptor riboprobe was used in RNase protection assays instead of the human IGF-I probe, a faint, 379 bases IGF-I receptor specific band was detected (Fig. 3). While IGF-I RNA expression decreased very rapidly and dramatically after 2 h of serum deprivation, IGF-I receptor RNA in- creased: after only 2 h of serum deprivation IGF-I receptor RNA transcripts as measured by solution hybridization assays and analysis of protected bands by computerized densitometry had almost increased 5-fold and reached a maximum after 72 h of serum deprivation (Fig. 4A,B). The expression of beta-actin RNA visualized as 121 bases and 51 bases protected fragments also increased steadily after serum deprivation of IM-9 lymphoblasts (Figs. 2, 3 and 5, and data not shown). Re-addition of serum after 24 h of serum free culture for various periods of time (24, 48 and 72 h) led to a significant increase in IGF-I RNA at 24, 48 h and 72 h but did not further change IGF-I receptor RNA (Figs. 2-4A,B). Addition of GH (100 ng/ml), IGF-I (50 ng/ml) or II-1 beta (5 IU/ml) did not alter the change in IGF-I and IGF-! receptor RNA expression detected during the serum deprivation period (data not shown).
3.3. Expression of GH receptor and IGF-H / M6P receptor RNA after serum deprivation and serum re-addition
RNase protection/solution hybridization assays using a radiolabeled human GH receptor riboprobe and IM-9 cell
320 Y. Yang et al. / Biochimica et Biophysica Acta 1310 (1996) 317-324
RNA revealed a 670 bases protected GH receptor specific band on autoradiograms of polyacrylamide gels (Fig. 5). When a 32p-labeled human IGF-II/M6P receptor ribo- probe was used in RNase protection assays a 260 bases IGF-II/M6P receptor specific band was detected (Fig. 6). GH receptor RNA expression decreased rapidly and dra- matically after 2 h of serum deprivation, stayed stably low for 24 h and increased sharply at 48 h of serum depriva- tion. These changes were in contrast to the changes of beta-actin mRNA expression which steadily increased within 8 h of serum deprivation (Fig. 7). IGF-II/M6P receptor RNA increased gradually and steadily throughout the duration of the experiment, independent of whether or not serum was readded (Fig. 6 and data not shown). Again, the density of beta-actin specific protected bands (51 bases protected band) increased in all experiments gradually and steadily with the duration of the experiments, a pattern that was similar to the pattern observed with the IGF-II/M6P receptor specific band at 260 bases (Fig. 6 and data not shown). Addition of GH (100 ng/ml), IGF-I (50 ng/ml) or I1-1 beta (5 IU/ml) again had no influence upon the
b l l m
4 2 0 IGF-I
3 5 0
B-~tin
Fig. 2. Serum-deprivation of IM-9 lymphoblasts leads to a decrease in IGF-I RNA expression. Cells were cultured serum-free for various peri- ods of time. In a subset of cell cultures serum was readded (24, 48 and 72 h of culture after serum re-addition, last 3 lanes). Solution hybridisation/RNase protection was carried out as described in Section 2. St, molecular weight markers (bases); C1, 32p-riboprobe incubated with RNases; C2, riboprobe alone. Two RNA species of 420 and 350 bases respectively were protected (upper arrow). The relatively low abundance of IGF-I RNA in comparison to beta-actin RNA (lower arrows) is demonstrated.
bases
1 6 3 2 _
S 0 6 i - -
N
1 5 4 D
hours
379 I G F - I - R
= R-actin
stqc
serum .* . . . . . . . * *
Fig. 3. Serum-deprivation of IM-9 lymphoblasts leads to an increase in IGF-I receptor RNA and beta-actin RNA expression. Cells were cultured serum-free for various periods of time. In a subset of cell cultures serum was re-added (+ ) . Solution hybridisation/RNase protection was carried out as described in Section 2. St, molecular weight markers (bases); C1, 32p-riboprobe incubated with RNases; C2, riboprobe alone.
pattern of changes of gene expression seen with serum deprivation (data not shown).
4. Discussion
We have used the IM-9 lymphoblast cell line to exam- ine the regulation of IGF-I and of IGF-I receptor gene expression during proliferative phases of human cells. The results obtained suggest that the GH receptor may be involved in regulating IGF-I production by IM-9 cells and that there is evidence as to a putative compensatory up- regulatory mechanism involving the IGF-I receptor after serum withdrawal and decrease of endogenous IGF-I ex- pression.
Our data demonstrate that IGF-I and IGF-I receptor RNA expression in IM-9 lymphoblasts change in an oppo- site direction upon serum starvation: the expression of IGF-I RNA species declines dramatically within 2 h after serum withdrawal and gradually increased after 48 h of culture in serum free medium. Addition of serum at 24 h or inclusion of hGH (100 ng/ml), or IGF-I (50 ng/ml) or
Y. Yang et al. / Biochimica et Biophysica Acta 1310 (1996) 317-324 321
IL-1 beta (5 IU/ml) during the serum free conditions did not change IGF-I RNA expression in IM-9 lymphoblasts further. Since the concentrations of single stimulants used in these experiments is in accordance to biologically effec- tive and/or physiological doses, it is most likely that IM-9 cells do not respond to these agents under the conditions of our experiments. Two protected fragments of IGF-I RNA were detected in IM-9 cell RNA in our study. Similar observations were made in the human ovary [20] and also in IM-9 lymphoblasts [23,24]: it is thought that multiple RNA species are caused by the use of different promoters of the IGF-I gene. Indeed, multiple transcripts of the IGF-I gene have been found to be expressed in IM-9 lympho- blasts with exon 2 being the major leader exon [24]. In contrast to the IGF-I RNA species, IGF-I receptor RNA increased rapidly after withdrawal of serum and increased in a continuous fashion (Fig;. 4). Such an increase in IGF-I receptor mRNA expression might indicate a putative com-
1,0 •
c :1
0,8. m
0,6.
m ¢: Q "o • ~ 0,4. u
o
0,2
0,2.
J~ 0,1
m c (D "0
0
0 0,0
/
i ! i i ! 0 2 4 8 12 24 48 72 (h)
Time
w i !
0 2 4 8 12 24 48 72 (h) Tinw
* p < 0 . 0 5
r= Imrum
free
recover serum
* p < 0 . 0 S
B I N l r u m free
, , r e c o v e r
8 ( i t u m
Fig. 4. Serum deprivation of IM-9 lymphoblasts leads to a decrease in IGF-I RNA (panel A) and an increase in IGF-I receptor RNA (panel B) expression. Cells were cultured serum-free for various periods of time. In a subset of cell cultures serum was readded ( + ) . Solution hybridisation/RNase protection was carded out as described in Section 2. Autoradiographs were analyzed by computed densitometry as described in Section 2. The data points represent mean + S.D. of three (panel A) or four (panel B) independent cultures and the representative RNAse protec- tion assays.
517 506
3N
344
29e
154
) 8 8 e 8
670 GH-R
sqc2
hours 2 8 24 72 48 4 12 48 24 "/2
Fig. 5. Serum-deprivation of IM-9 lymphoblasts leads first to a rapid decrease and in a second phase to an increase in GH receptor RNA expression. Cells were cultured serum-free for various periods of time. In a subset of cell cultures serum was readded ( + ) . Solution hybridisation/RNase protection was carried out as described in Section 2. St, molecular weight markers (bases); C1, 32p-riboprobe incubated with RNases; C2, riboprobe alone. The lower part of the panel shows the expression of beta-actin mRNA of the same experiment as analyzed in a parallel gel.
pensatory up-regulatory mechanism at the receptor mRNA level under condi t ions where IGF-I l igand expression/concentrations are low.
Conversely, changes in GH receptor mRNA initially paralleled the changes of IGF-I RNA that were seen upon the removal of serum from the cell cultures. Indeed, it has been shown that in EBV-transformed B-lymphoblasts from healthy subjects, GH regulates IGF-I production presum- ably through interaction with its own receptor [27]. In addition, the content of IGF-II/M6P receptor RNA and beta-actin RNA of IM-9 cells increased throughout the duration of the experiment in a continuous fashion. These changes were again observed independently of whether or not the cells were kept serum free or serum free with the addition of either IGF-I, hGH or II-1 beta. Also, the re-addition of serum after 24 h under serum free conditions did not significantly change the expression of IGF-II/M6P
322 Y. Yang et al. /Biochimica et Biophysica Acta 1310 (1996) 317-324
bp
2 9 ~
1 5 4 ~
St. C1C 2
2 6 0 I G F - I I / M G P - R
~-actin
hours 2 8 24 72 48 4 12 48 Z4 72
serum ¢ . . . . . . . + . ÷
Fig. 6. Serum-deprivation of IM-9 lymphoblasts leads to an increase in IGF-II/M6P receptor RNA and beta-actin RNA expression. Cells were cultured serum-free for various periods of time. In a subset of cell cultures serum was readded. Solution hybridisation/RNase protection was carried out as described in Section 2. St, molecular weight markers (bases); C1, 32p-riboprobe incubated with RNases; C2, riboprobe alone.
~-~ 1.2-
, 1 . 0 -
£ v m 0.8
m t~ 0.4
o.2
0 0 • i • i • i • i
0 2 4 8 12 24 48 72 (h) T i m e
* p < 0.05
'~ s e r u m f ree
• r e c o v e r y s e r u m
Fig. 7. Serum-deprivation of IM-9 lymphoblasts leads to a decrease and subsequent increase of GH receptor RNA expression. Cells were cultured serum-free for various periods of time. In a subset of cell cultures serum was readded. Solution hybridisation/RNase protection was carried out as described in Section 2 and computed densitometry was used to quantitate the results from 3 independent experiments (3 independent cell culture studies). Data points are means + S.D.
receptor RNA, GH receptor RNA and beta-actin RNA. The changes of beta-actin expression during serum with- drawal in these experiments make the use of beta-actin as a control for equal loading of the gels rather complicated. However, the reproducible pattern of beta-actin RNA ex- pression can serve both for a measurement of equal load- ing and for a means to determine the reproducibility of the cell culture experiments themselves.
We conclude that IGF-I and GH receptor RNA are expressed in parallel and serum deprivation leads to a rapid and dramatic decline of both IGF-I and GH receptor RNA. In a second phase of changes in IGF-I and GH receptor RNA expression, increased transcription of the two genes was observed after approx. 24 h of serum starvation. Interestingly, this increase of IGF-I and GH-re- ceptor RNA expression coincides with an increase in cell number at or shortly after the same time.
In contrast, IGF-I receptor RNA increases rapidly and continuously after removal of serum from IM-9 cells in culture. Such an increase in IGF-I receptor mRNA expres- sion might represent a compensatory up-regulatory mecha- nism at the receptor mRNA level. IGF-I I /M6P receptor and beta-actin RNA also increase after serum withdrawal. However, these changes are more subtle and occur at a slower rate. It is interesting to note that the maximal content of IGF-I receptor transcripts in IM-9 cells in serum free culture was measured 24-48 h after removal of serum. It was at the same time point that cell numbers had increased significantly.
Suzuki et al. have previously reported that GH increases transcription of the GH receptor gene in IM-9 cells and also stimulates growth of IM-9 lymphoblasts [25]. It is known that IM-9 cells synthesize and secrete GH under certain culture conditions and that increased GH secretion is observed upon stimulation of cells with GH [26]. In contrast, we did not detect any effect of hGH on gene transcription in IM-9 cells under the conditions of our experiments. Our results are in agreement with the data from Clayton et al. who did not find any effect of hGH on IGF-I gene expression in IM-9 lymphoblasts [24]. We assume that either differences in culture conditions or the clones of IM-9 lymphoblasts used in the different studies may be responsible for this discrepancy. We hypothesize that under the conditions of our experiments serum fac- tors/substrates other than GH, IGF-I and II-1 beta regulate IGF-I, IGF-I receptor, GH receptor and IGF-I I /M6P re- ceptor RNA.
GH and the IGFs have been implicated as playing a role in the proliferation of T-lymphocytes in vitro [28]; such changes might also directly influence the proliferative response of IM-9 cells in culture. Growth hormone and insulin have been shown to induce a rapid down-regulation ot GH receptors and insulin receptors in IM-9 lympho- blasts [29]. Conversely, IGF-II and a monoclonal antibody against the T-cell receptor have been shown to up-regulate IGF-I receptor specific binding of 125I-IGF-I in IM-9 cells
Y. Yang et al. / Biochimica et Biophysica Acta 1310 (1996) 317-324 323
or IGF-I receptor RNA content in peripheral T-lympho- cytes [14,17]. Other hormones such as human chorionic gonadotropin have been shown to induce IGF-I receptor gene expression [30-32] or down-regulate IGF-I receptor RNA [33,34]. Developmental regulation of the IGF-I re- ceptor and the IGF-II/M6P receptor gene has been de- scribed [9,15,35].
Both inhibitory and stirnulatory effects of sera upon IGF gene transcription have been described [36]. A bidirec- tional effect of serum removal upon IGF-I RNA expres- sion was measured in our study. It is conceivable that removal of serum from the IM-9 cells leads to a rapid decline in IGF-I RNA expression through the withdrawal of a stimulatory factor. A number of hormones including growth hormone have been described to stimulate IGF-I gene transcription in other cells [1,27,37,38].
It has been reported that both IGF-II and IGF-II/M6P receptor levels increase during skeletal muscle differentia- tion. In addition, IGF-II/M6P receptor gene expression is increased during compensatory growth of hepatic, renal and thyroid tissue and during endochondrial bone develop- ment [7,9,11,15,39,40]. Since the IGF-II/M6P receptor does not serve as a signal transduction pathway for the IGFs [41] but rather ser~'es to direct the transport of lysosomal enzymes to the; lysosome [7,10], changes in IGF-II/M6P receptor gene expression during cellular pro- liferation might not be directly related to growth and proliferation but rather be important for lysosomal enzyme function.
In summary, we have observed that the genes encoding IGF-I, the IGF-I receptor, the IGF-II/M6P receptor and the GH receptor are differentially regulated in IM-9 lym- phoblasts. The mechanisms by which expression of these genes is being regulated are not known but are thought to involve a number of different transcription factors such as the WT1 Wilms tumor gene product [42].
Acknowledgements
This work was supported by Deutsche Forschungsge- meinschaft, Bonn, German~y (DFG Ki 365/1) (W.K.) and Deutscher Akademischer Austauschdienst, Bonn, Germany (Y.Y.). The authors wish to express their gratitude to Drs. C.R. Roberts and D. LeRoith, Bethesda, USA, and J. Guo, Shanghai, China, for their kind gift of cDNA probes.
References
[1] Rechler, M.M. and Nissley, S.P. (1990) in: Peptide Growth Factors and Their Receptors. I. Handbook of Pharmacology (Sporn, M.B. and Roberts, A.B., eds.), Springer, Heidelberg, pp. 263-367.
[2] Rotwein, P. (1991) Growth Factors 5, 3-18. [3] Yang, Y., Hoeflich, A., Kessler, U., Barenton, B., Blum, W.F.,
Schwarz, H.P. and Kiess, W. (1993) Regul. Pept. 48, 41-53.
[4] Delehaye-Zervas, M.C., Mertani, H., Martini, J.F., Nihoul-Fekete, C., Morel, G. and Postel-Vinay, M.C. (1994) J. Clin. Endocrinol. Metabol. 78, 1473-1480.
[5] Nissley, S.P. and Lopaczynski, W. (1991) Growth Factors 5, 29-43. [6] Oh, Y., Miiller, H.L., Neely, E.K., Lamson, G. and Rosenfeld, R.G.
(1993) Growth Regul. 3, 113-123. [7] Nissley, S.P., Kiess, W. and Sklar, M.M. (1991) in: Insulin-like
Growth Factors: Molecular and Cellular Aspects (D. LeRoith, ed.), CRC Press, Boca Raton, pp. 111-150.
[8] Kiess, W., Thomas, C.L., Greenstein, L.A., Lee, L., Sklar, M.M., Rechler, M.M., Sahagian, G.G. and Nissley, S.P. (1989) J. Biol. Chem. 264, 4710-4714.
[9] Kiess, W., Hoeflich, A., Yang, Y., Kessler, U., Flyvbjerg, A. and Barenton, B. (1994) in: Current Directions in Insulin-like Growth Factor Research (D. LeRoith and M.K. Raizada, eds.), Plenum Press, New York.
[10] Kessler, U., Aumeier, S., Funk, B. and Kiess, W. (1992) Mol. Cell. Endocrinol. 90, 147-153.
[11] Kornfeld, S. (1992)Ann. Rev. Biochem. 61,307-330. [12] Adashi, E.Y., Resnick, C.E., Vera, A. and Hernadez, E.R. (1991)
Endocrinology 128, 3130-3137. [13] Yamasaki, H., Prager, D., Gebremedhin, A. and Melmed, S. (1991)
E.J., Goldfine, I.D. and Rosenthal, S.M. (1992) J. Cell. Biochem. 48, 81-85.
[18] Ohlsson, C., Nilsson, A., Isaksson, O., Bentham, J. and Lindahl, A. (1992) J. Endocrinol. 135, 115-123.
[19] Sussenbach, J.S., Sittenbergh, P.H. and Holthuizen, P. (1992) Growth Regul. 2, 1-9.
[20] Hernandez, E.R., Hurwitz, A., Vera, A., Pellicer, A., Adashi, E.Y., LeRoith, D. and Roberts, C.T. Jr. (1992) J. Clin. Endocrinol. Metabol. 74, 419-425.
[21] Hoeflich, A., Yang, Y., Kessler, U., Heinz-Erian, P., Kolb, H. and Kiess, W. (1994) Mol. Cell. Endocrinol. 101, 141-150.
[22] Raile, K., Hoeflich, A., Kessler, U., Yang, Y., Pfuender, M., Blum, W.F., Kolb, H., Schwarz, H.P. and Kiess, W. (1994) J. Cell. Physiol. 159, 531-541.
[23] Clayton, P.E., Hellmann, P., Day, K.H. and Day, R.N. (1992). Differential expression of IGF-I mRNAs in human IM-9 lympho- cytes. 74 th Annual Meeting, Endocrine Society, San Antonio, TX, abstract 685.
[24] Clayton, P.E., Day, R.N., Silva, C.M., Hellmann, P., Day K.H. and Thorner, M.O. (1994) J. Mol. Endocrinol. 13, 127-136.
[25] Suzuki, K., Suzuki, S., Saito, Y., Ikebuchi, H. and Tereao, T. (1990) J. Biol. Chem. 265, 11320-11327.
[26] Hattori, N., Shimatsu, A., Sugita, M., Kumagai, S. and Imura, H. (1990) Biochem. Biophys. Res. Commun. 168, 396-401.
[27] Merimee, T.J., Grant, M.B., Broder, C.M. and Cavalli-Sforza, L.L. (1989) J. Clin. Endocrinol. Metabol. 69, 97-984.
[28] Kooijman, R., Willems, M., Rijkers, G.T., Brinkman, A., Van Buul-Offers, S.C., Heijnen, C.J. and Zegers, B.J.M. (1992) J. Neu- roimmunol. 38, 95-104.
[29] Torossian, K., Freedman, D. and Fantus, I.G. (1988) J. Biol. Chem. 263, 9353-9359.
[30] Lin, T., Blaisdell, J. and Haskell, J.F. (1988) Endocrinology 123, 134-139.
[31] Mulroney, S.E., Haramati, A., Werner, H., Bondy, C., Roberts, C.T. and LeRoith, D. (1992) Endocrinology 130, 249-256.
[32] Nagpal, M.L., Wang, D., Calkins, J.H., Chang, W. and Lin, T. (1991) Endocrinology 129, 2820-2826.
324 Y. Yang et al. / Biochimica et Biophysica Acta 1310 (1996) 317-324





![Biochi~ic~a - core.ac.uk · planes and the O~Xi axe of the two molecules are first superimposed. Following ... and Perun et al. [19] for erythromycin, another macrolide](https://static.documents.pub/doc/80x56/5b407f6f7f8b9af46b8d831c/biochiica-coreacuk-planes-and-the-oxi-axe-of-the-two-molecules-are-first.jpg)



![Biochi~ic~a et Biophysica A~ta - COnnecting REpositoriesc a and o has been purified from A. vinelandii membranes [12]. This complex was found to be able to oxidize ascor- bate-N, N,](https://static.documents.pub/doc/80x56/60cf08ca3bd7762e217d18c5/biochiica-et-biophysica-ata-connecting-repositories-c-a-and-o-has-been-purified.jpg)



![Ulster University Biochi… · Web viewThe [A14K] analogue (1 µM) produced a significant increase in cAMP concentration in BRIN-BD11 cells and down-regulation of the protein kinase](https://static.documents.pub/doc/80x56/5f269236bbfd65169e078685/ulster-university-biochi-web-view-the-a14k-analogue-1-m-produced-a-significant.jpg)








