Individual electrofusion of a pair of sunflower protoplasts with a given cell diameter can be performed by different combinations of physical parameters (pulse duration and field strength). The determination of ethane was used to indicate the extent of membrane lipid degradation as a result of different fusion parameters. The degree of membraneous damage measured by lipid peroxidation is negatively correlated with the regeneration ability of protoplasts, which was demonstrated by determining the protoplast division rate after electrofusion. Thus, it was possible to develop a tool for monitoring the protoplast regeneration potential at an early stage.
Lipid peroxidation of unsaturated fatty acids is a highly detrimental process to cell membrane structures and is linked to effects such as membrane damage, increased ion permeability, loss of fluidity, inactivation of membrane enzymes and receptors, and loss in membrane stability (Girotti, 1990).
Ethane production has often been described as a tool for measuring lipid peroxidation, and it serves as a sensitive indicator for membrane deterioration (Galliard, 1970; Konze and Elstner, 1978; Youngman et al., 1980; Schnabl et al., 1983 a, b; Schnabl and Youngman, 1985). Ethane formation is only a minor reaction pathway in lipid peroxidation, but the major advantage is that it can be used as a non-invasive in vivo measure of membrane lipid degradation. The release of ethane from plant tissues or protoplasts can be stimulated by chemical treatment with ecotoxicological substances such as pentachlorophenol or HgCh (Schnabl et al., 1984; Schnabl and Youngman, 1987), by aging, infection, wounding (Konze and Elstner, 1978) or electric pulses (Biedinger et al., 1990). It
can be delayed by immobilization in an alginate matrix (Schnabl et al., 1983 a) or by adding radical scavengers such as ascorbate, SOD and tocopherol (Biedinger et al., in press).
Electrofusion is a well experienced and often described method to generate somatic hybrids (Zachrisson and Bornman, 1986; Zimmermann and Scheurich, 1981; Zimmermann, 1986; Zimmermann et al., 1988). The formation of somatic hybrids is followed by their regeneration to intact plants with different and new genetic information. Although electro fusion offers advantages over more traditional chemical fusogens, we recently showed that the conditions of electromanipulations represent a significant stress (Biedinger et al., 1990). Electric pulses of high intensity are able to induce the lipid peroxidation process by the generation of free lipid radicals (Biedinger et al., in press). In the presence of oxygen, free lipid radicals are oxygenated to lipid peroxide radicals, which initiate a chain reaction. They are then degraded to a range of end products, including ethane.
With respect to subsequent regeneration electrofusion parameters have to be optimized to achieve a high yield of fusion products and to attain protoplast viability and the
418 URSULA BIEDINGER and HEIDE SCHNABl
regeneration potential. Recently, Mehrle et al. (1990) demonstrated a strong correlation between the protoplast diameter, field strength and pulse duration. This indicates that the fusion event of a pair of individual protoplasts is achieved by various combinations of field strength and pulse duration with a given cell diameter. It is of great interest to identify the electrical parameter combinations for successful fusion events with respect to their effects on protoplast membrane integrity and regeneration capacity. Ethane degeneration proved to be a sensitive indicator for membrane lipid degradation and a useful tool for estimating and characterizing the ability of fused protoplasts for regeneration at a very early state. Therefore, in this report, we developed a method to characterize the viability of protoplasts treated with different pulse combinations by means of the division rate.
Materials and Methods
Plant material and isolation of protoplasts
Mesophyll cell protoplasts were isolated from the first expanded leaves of 14-day-old sunflower plants (cv. Primasol from the Institut fiir Pflanzenziichtung, Universitat Giessen, FRG) grown on MS-medium under sterile conditions. Plant growth conditions and the isolation method have been previously described by Schmitz and Schnabl (1989).
Protoplast fusion
In order to determine the fusion event of a pair of two similiar protoplasts, the protoplast suspension was highly diluted and ad~ justed to contain about 4 x 103 protoplastsl mL. The protoplast diameter was determined microscopically using a calibrated micrometer. Fusion experiments were carried out in a fusion chamber consisting of a pair of V 2A-steel electrodes that were fixed on a microscope slide with a distance of 1 mm to each other and covered by a slide.
Applying an alternating field of 100 VI cm and 2 MHz, the protoplasts were aligned to form singles, pairs or short chains of protoplasts between the two electrodes, due to mutual dielectrophoresis. For each individual experiment a pair of protoplasts with equal diameter was observed before applying a single direct current pulse. On the basis of a given pulse length (1 to 99.9!-,sec), the field strength was varied until the parameter combination of a definite ~ulse length and the required field strength resulted in protoplast fusIOn.
The protoplast fusion was performed for the ethane determination and for regeneration purposes in a bulk fusion chamber filled with 400!-,L of a protoplast suspension. The density of the protoplast suspension was adjusted to about 106 protoplasts per mL. Electrical fields were applied by a Zimmerman Cell Fusion Power Supply.
Non-pulse treated (non-fused) protoplasts were investigated as a control.
Determination of ethane
The incubation of pulse-treated and unpulsed protoplasts and the determination of ethane was described previously (Schnabl et al., 1984; Biedinger et al., 1990). After fusion, aliquots of protoplasts were transferred to Fernbach flasks fitted with serum rubber stoppers and incubated at 25°C for 16 h in the dark. Then 1 mL headspace samples were withdrawn from flasks using disposable sy-
ringes and analyzed for ethane by gas chromatography. A Hewlett Packard model with an alumina column (length 305 mm, diameter 3.2 mm) was used with N2 as carrier gas.
Viability test
The effects of different parameter combinations of pulse length and field strength were tested by FDA (fluorescein diacetate) staining during e1ectrofusion on the viability of protoplasts. The final FDA concentration was 0.025 %.
Protoplast culture
After electrofusion, intact protoplasts were resuspended in VKM-medium, a nutrient medium by Binding and Nehls (1978) and embedded in agarose (final concentration 0.6 %) according to the technique of Shillito et al. (1983). Droplets were formed with an Eppendorf pipette and allowed to fall onto petri dishes. To prevent the protoplasts from drying out, droplets were overlayed by liquid VKM-medium.
The addition of 2 mg/L NAA and 0.5 mg/L BAP to the V-KMmedium stimulated the cell division activity.
All experiments were performed at least 8 -10 times.
Results
Influence of the fusion parameter on electrofusion of protoplasts
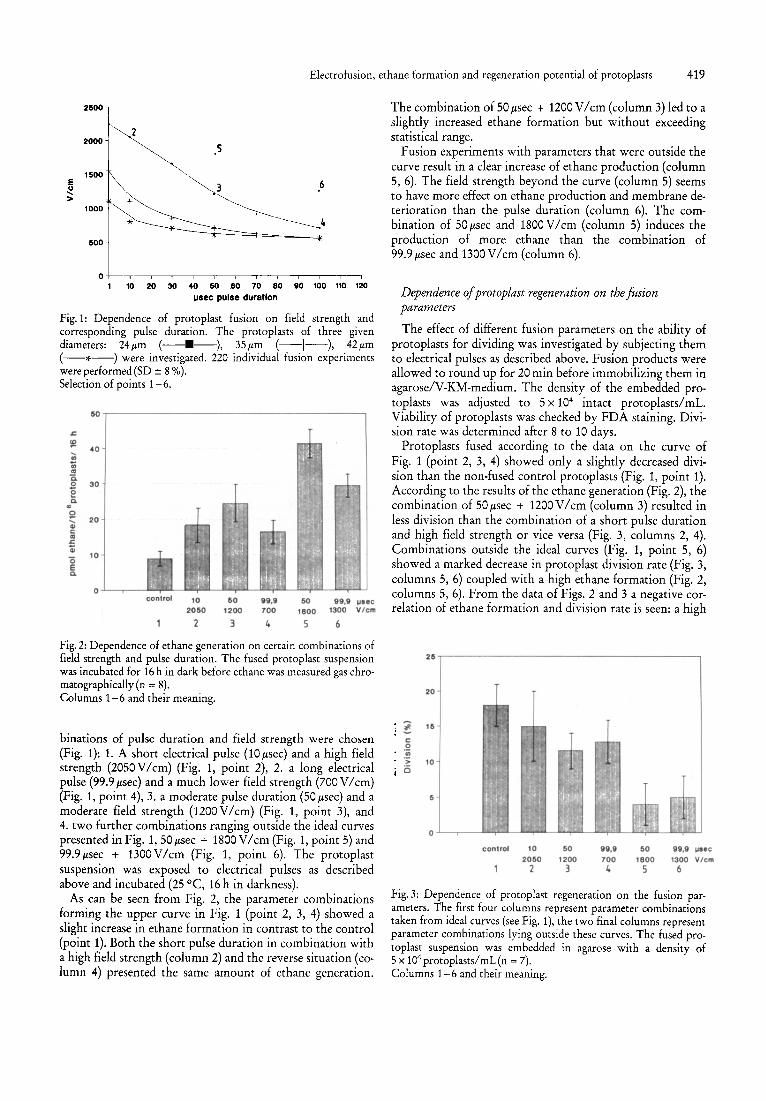
We investigated different fusion parameter combinations for sunflower mesophyll cell protoplasts. The individual fusion of pairs of protoplasts with a diameter of 24, 35 and 42ltm (Fig. 1) was used to show a correlation between the diameter of protoplasts, the required field strength and respectively, the pulse length. The electrical membrane breakdown (= fusion event) was achieved with a pair of protoplasts with a given cell diameter by different combinations of pulse duration and field strength. The shorter the pulse duration, the higher was the field strength necessary for a successful protoplast fusion. Smaller protoplasts needed a higher field strength for fusion than larger ones. The fusion parameter for protoplasts of a 24ltm diameter showed a linear correlation between field strength and pulse length in the range of 1-70 Itsec. A lower field strength was coupled with a longer pulse duration and vice versa for achieving protoplast fusion. Using a field strength of 800 V / cm, the fusion event becomes more or less independent of the pulse duration (minimum pulse duration 100 Itsec). For protoplasts of 35 and 42ltm diameter, there is a sharp decrease of required field strength between 1 and 10 Itsec pulse duration, while it decreases more gradually between 10 and 100 Itsec.
According to Mehrle et al. (1990) the field strength is the limiting factor for fusion events.
Dependence of ethane generation on different fusion parameter combinations
In order to determine the membrane degradation by lipid peroxidation, we measured ethane production as the result of protoplast fusion. Since the size of the sunflower protoplasts in the suspension was about 241tm, subsequent com-
Electrofusion, ethane formation and regeneration potential of protoplasts 419
2600
1 2000
1500 E u .... >
1000
600
2
5
6
+
*
ro ro ~ ~ 50 60 ro 50 50 m 00 m IIsec pulse duration
Fig. 1: Dependence of protoplast fusion on field strength and corresponding pulse duration. The protoplasts of three given diameters: 24 p,m (--.--), 35 p,m (--1--), 42 p,m (--*--) were investigated. 220 individual fusion experiments were performed(SD ± 8 %). Selection of points 1-6.
5O ~------------------------------------,
&;
!!! 40 , !! ., CD c. £ 30 ~ Co .. ~ 20 .... ., c .. &;
~ 10 (;
E Co
eontrot 10 60 1111,11 50 99,11 ~ .. e 2050 1200 700 1800 t300 V tern
2 3 4 5 6
Fig. 2: Dependence of ethane generation on cenain combinations of field strength and pulse duration. The fused protoplast suspension was incubated for 16 h in dark before ethane was measured gas chromatographically(n = 8). Columns 1-6 and their meaning.
binations of pulse duration and field strength were chosen (Fig. 1): 1. A short electrical pulse (10/lsec) and a high field strength (2050 V / cm) (Fig. 1, point 2), 2. a long electrical pulse (99.9/lsec) and a much lower field strength (700 V / cm) (Fig. 1, point 4), 3. a moderate pulse duration (50/lsec) and a moderate field strength (1200V/cm) (Fig. 1, point 3), and 4. two further combinations ranging outside the ideal curves presented in Fig. 1,50 /lsec + 1800 V / cm (Fig. 1, point 5) and 99.9 /lsec + 1300 V / cm (Fig. 1, point 6). The protoplast suspension was exposed to electrical pulses as described above and incubated (25 °C, 16 h in darkness).
As can be seen from Fig. 2, the parameter combinations forming the upper curve in Fig. 1 (point 2, 3, 4) showed a slight increase in ethane formation in contrast to the control (point 1). Both the short pulse duration in combination with a high field strength (column 2) and the reverse situation (column 4) presented the same amount of ethane generation.
The combination of 50 /lsec + 1200 V / cm (column 3) led to a slightly increased ethane formation but without exceeding statistical range.
Fusion experiments with parameters that were outside the curve result in a clear increase of ethane production (column 5, 6) . The field strength beyond the curve (column 5) seems to have more effect on ethane production and membrane deterioration than the pulse duration (column 6). The combination of 50/lsec and 1800V/ cm (column 5) induces the production of more ethane than the combination of 99.9/lSec and 1300V/ cm (column 6).
Dependence of protoplast regeneration on the fusion parameters
The effect of different fusion parameters on the ability of protoplasts for dividing was investigated by subjecting them to electrical pulses as described above. Fusion products were allowed to round up for 20 min before immobilizing them in agarose/V-KM-medium. The density of the embedded protop lasts was adjusted to 5 x 104 intact protoplasts/mL. Viability of protoplasts was checked by FDA staining. Division rate was determined after 8 to 10 days.
Protoplasts fused according to the data on the curve of Fig. 1 (point 2, 3, 4) showed only a slightly decreased division than the non-fused control protoplasts (Fig. 1, point 1). According to the results of the ethane generation (Fig. 2), the combination of 50/lsec + 1200V/cm (column 3) resulted in less division than the combination of a short pulse duration and high field strength or vice versa (Fig. 3, columns 2, 4). Combinations outside the ideal curves (Fig. 1, point 5, 6) showed a marked decrease in protoplast division rate (Fig. 3, columns 5, 6) coupled with a high ethane formation (Fig. 2, columns 5, 6). From the data of Figs. 2 and 3 a negative correlation of ethane formation and division rate is seen: a high
211
20
~ 16
c: 0
OJ -; 10 0
6
eontrot 10 50 1111,11 50 99,9 ~ •• e 20110 1200 700 1800 1300 V tern
2 3 4 5 6
Fig. 3: Dependence of protoplast regeneration on the fusion parameters. The first four columns represent parameter combinations taken from ideal curves (see Fig. 1), the two final columns represent parameter combinations lying outside these curves. The fused protoplast suspension was embedded in agarose with a density of 5x 104 protoplasts/mL(n = 7). Columns 1-6 and their meaning.
420 URSULA BIEDINGER and HEIDE SCHNABL
ethane production is reflected by a low division ability of fused protoplasts.
Discussion
By varying the field strength, pulse duration and protoplast size during electrofusion, we succeeded in confirming the results of Mehrle et ai. (1990) using tobacco protoplasts. Our results demonstrated the strong correlation of these parameter by investigating sunflower mesophyll cell protoplasts.
The interdependence of field strength and pulse duration shows that for a given protoplast size, the same fusion event occurs by either applying field pulses of high intensity and short duration, or vice versa. Our main interest was to study the effect of different electrical treatments on the membrane stability (ethane production as an indicator) and its relationship with the protoplast regeneration ability (division rate as an indicator).
Our data on ethane generation indicate differences between the various parameter combinations. The application of a field pulse of high intensity and short duration, and vice versa, resulted in an equal amount of ethane generation (Fig. 2, columns 2, 4), whereas the moderate combination of 50 psec pulse duration combined with a field strength of 1200 V / cm (Fig. 2, column 3) led to a slightly increased ethane formation. Parameter deviations from the ideal curves indicate a markedly higher stress, which resulted in a strongly increased ethane production (Fig. 2, columns 5, 6).
Previously, we demonstrated that electric pulses used for electro manipulation of protoplasts lead to the generation of free lipid radicals (Biedinger et aI., in press). This resulted in the formation of ethane by inducing the lipid peroxidation process. Lipid peroxidation is linked with membrane degradation, inhibition of enzyme activity, increase in membrane permeability and loss of membrane integrity (Chance et aI., 1979; Girotti, 1990). We proposed that these detrimental effects reduce the ability of protoplasts to regenerate. Comparing Fig. 2 and Fig. 3 the generation of ethane is negatively correlated with the division rate of protoplasts. An increase in ethane formation is accompanied by a decreased protoplast division.
We therefore conclude that a successful fusion of protoplasts of a given size can be achieved by different combinations of field strength and pulse duration. The effect, however, of these different parameter variations indicates differences in protoplast membrane deterioration and ability of regeneration. Thus, it is particularly important to choose the optimal physical factors in order to achieve optimal protoplast fusion and a high capacity for regeneration.
The study reported here shows that the determination of ethane as an indicator of lipid peroxidation induced by fusion conditions can be used as a highly sensitive tool to estimate the regeneration potential of fused protoplasts at a very early stage, before time-consuming morphological changes can be observed.
Acknowledgements
The authors gratefully acknowledge the financial support of the Bundesministerium fiir Forschung and Technologie (BMFT/ DLR) to H.S. (0IQV87672).
References
BIEDINGER, U., R.J. YOUNGMAN, and H. SCHNABL: Differential effects of electrofusion and electropermeabilization parameters on the membrane integrity of plant protoplasts. Planta 180,598-602 (1990).
BIEDINGER, U ., CHR. BICKERT, R. J. YOUNGMAN, and H. SCHNABL: The formation of free lipid radicals during the electromanipulation of protoplasts (Vicia /aba). Botanica acta, in press.
BINDING, H. and R. NEHLS: Regeneration of isolated protoplasts of Vicia faba L. Z. Pflanzenphysio!. 88, 327 - 332 (1978).
CHANCE, B., H. SIES, and A. BOVERIS: Hydroperoxide metabolism in mammalian organs. Physio!. Rev. 53, 527 -605 (1979).
GALLIARD, T.: The enzymic breakdown of lipids in potato tuber by phospholipid- and galactolipid acyl hydrolase activities and by lipoxygenase. Phytochemistry 9,1725-1734 (1970).
GIROTTI, A. W.: Photobiology school: Photodynamic lipid peroxidation in biological systems. Photochemistry and Photobiology Vo!' 51, No.4, 497-509 (1990).
KONZE, J. R. and E. F. ELSTNER: Ethane and ethylene formation by mitochondria as an indication of aerobic lipid degradation in response to wounding of plant tissue. Biochim. Biophys. Acta 528, 213 -221 (1978).
MEHRLE, W., B. NATON, and R. HAMPP: Determination of physical membrane properties of plant cell protoplasts via the electrofusion technique: prediction of optimal fusion yields and protoplast viability. Plant Cell Reports 8, 687 -691 (1990).
SCHMITZ, P. and H. SCHNABL: Regeneration and evacuolation of protoplasts from mesophyll, hypocotyl and petioles from Helianthus annuus L. J. Plant Physio!. Vo!' 135, 223 - 227 (1989).
SCHNABL, H ., R. J. YOUNGMAN, and U. ZIMMERMANN: Maintenance of plant cell membrane integrity and function by the immobilisation of protoplasts in alginate matrices. Planta 158, 392-397 (1983 a).
SCHNABL, H ., CH. ELBERT, and R. J. YOUNGMAN: Release of ethane from immobilized plant cell protoplasts in response to chemical treatment. Physio!. Plant. 59, 46-49 (1983 b).
SCHNABL, H ., J. KUNNE, K. FLORSCHUTZ, and R. J. YOUNGMAN: Differential effects of ecotoxicological substances on lipid peroxidation of plant cell protoplasts. Plant Cell Environ. 7, 499-505 (1984).
SCHNABL, H . and R. J. YOUNGMAN: Immobilisation of plant cell protoplasts inhibits enzymic lipid peroxidation. Plant Sci. 40, 65-69 (1985).
- - A characterization of the effects of ecotoxicological substances on plant cell metabolic reactions. Angew. Bot. 61, 493 - 504 (1987).
SHILUTO, R . D., J. PASZKOWSKI, and 1. POTRYKUS: Agarose plating and bead type culture technique enable and stimulate development of protoplast derived colonies in a number of plant species. Plant Cell Reports 2, 244-247 (1983).
YOUNGMAN, R. J., K. E. PALLETT, and A. D. DODGE: Active oxygen species in herbicide action. In: Chemical and biochemical aspects of superoxide and superoxide dismutase, pp. 402-411, BANNISTER, J. V. and H. A. O. HILL (eds.). Elsevier/North-Holland, New York (1980).
ZACHRISSON, A. and C. H . BORNMAN: Electromanipulation of plant protoplasts. Physio!. Plant. 67, 507 -516 (1986).
ZIMMERMANN, U. and P. SCHEURICH: High frequency fusion of plant protoplasts by electric fields. Planta 151, 26-32 (1981).
ZIMMERMANN, U., R. SCHNETTLER, and K. HANNIG: Biotechnology in space: Potentials and perspectives. In: Biotechnology, Vol. 6 b, pp. 637-672, REHM, H.-J. and G. REED (eds.). VCH Verlagsgesellschaft, Weinheim (1988).