Ethanol Oral Self-Administration and Rewarding Brain Stimulation
G E O R G E T. B A I N z A N D C O N A N K O R N E T S K Y
Laboratory of Behavioral Pharmacology, Boston University School of Medicine 80 East Concord Street, Boston, MA 02118
R e c e i v e d 30 Oc tobe r 1987; A c c e p t e d 9 June 1989
BAIN, G. T. AND C. KORNETSKY. Ethanol oral self-administration and rewarding brain stimulation. ALCOHOL 6(6) 499-503, 1989.--The effects of orally self-administered ethanol (ETOH) on responding for rewarding brain stimulation were studied. Bipolar electrodes were implanted in either the lateral hypothalamic region of the medial forebrain bundle (MFB-LH) or the ventral tegmental area (VTA) of 6 male F-344 rats. After surgery subjects were trained in a continuous reinforcement procedure (CRF) for constant current rewarding brain stimulation. On alternate days subjects were allowed to drink an ethanol and sucrose solution (12% and 5%, respectively) for 30 min and subsequently tested on the brain stimulation procedure. All subjects showed facilitation of responding (increase in rate) after ingesting low to moderate doses of ETOH (0.4-1.7 g/kg). Depression of responding (decrease in rate) or return to baseline levels (control solution rate) was observed only in those subjects which ingested 2 g/kg or greater during the drinking period. These results indicate that low to moderate doses of self-administered ethanol will increase responding for rewarding brain stimulation. Further, the results suggest that this facilitation of responding is, at least in part, a function of the method of administration and/or the contingent nature of the ethanol delivery (self-administration).
MANY substances which are readily abused by humans have also been found to facilitate responding for rewarding electrical stim- ulation to the brain (5, 6, 10, 11, 15). Although several groups have studied the effects of ethanol (ETOH) on brain stimulation reward in the rat the results have been notably mixed. This variability may be due to differences in the methods of presenta- tion and electrical characteristics of the brain stimulation, the area of the brain stimulated or the route of administration and dose range of the ETOH. Large increases in rate of responding (13), lowered thresholds (12) and increased time spent responding for rewarding brain stimulation (4) have been reported for low doses of intraperitoneally (IP) administered ETOH. Other investigators have reported more diverse effects ranging from attenuation of rate or no effect depending on stimulation site with ETOH doses which produced a marked state of intoxication (21) to increases in both response rate and peak force at low doses and decreased rate but increased duration of response at higher doses (26). Increased response rates with no effect on threshold for rewarding stimula- tion at low doses as well as increased threshold and no effect on response rate at high doses have also been reported (3). High doses of ETOH which have produced near comatose states in rats have been found to inhibit initiation of brain stimulation but not to block self-stimulation once initiated by the experimenter (18). Experi- ments conducted in our laboratory have yielded only decreases in response rate at high doses of ETOH and no effect at any dose tested (0.025-1.6 g/kg, IP) on threshold for rewarding brain stimulation (25).
With the exception of the study by DeWitte and Bada (4), in which one experimental group of rats drank only ETOH during the test days, all other experiments cited above, including our own, involved intraperitoneally (IP) administered ETOH at concentra- tions which may be irritating to the rats (26) or, as reported by other investigators, are unsuitable for administration by this route (14,27).
In a recent experiment by Schaefer and Michael (19) an alternative to the IP administration was examined. They reported when ETOH was administered intragastrically (IG) via an animal feeding needle, response rates significantly decreased at relatively high doses (3.0 and 4.5 g/kg) while threshold was not affected at any dose tested. In this study IP administration also yielded decreases in response rates from rewarding brain stimulation with little concurrent alteration in threshold levels.
Because of the potentially aversive aspects of IP administration of ETOH as employed in the majority of the previously reported studies and the possible confounding of results, as well as the potentially stressful nature of IG administration (via a feeding needle), we decided to examine the effects of orally self-admin- istered ETOH. Rate of response for brain stimulation was em- ployed since this type of measure or a variant thereof appeared to be most frequently cited as affected by ETOH.
~Supported by NIAAA Grant AA 05950 and NIDA Research Scientist Award DA 00099 to C.K. 2Requests for reprints should be addressed to George T. Bain, Ph.D., Boston University School of Medicine, 80 East Concord Street, L-602, Boston,
MA 02118.
499
500 BAIN AND KORNETSKY
VA) (0.013 cm in diameter and insulated except at the tip) were stereotaxically implanted in the brain of male F-344 rats (approx- imately 300 g at surgery) (Charles River Laboratories). The electrodes were aimed at either the lateral hypothalamic region of the medial forebrain bundle (MFB-LH) ( N = 3 ) or the ventral tegmental area (VTA) (N = 3). Surgical anesthesia was produced by systemic administration of xylazine and ketamine (13 mg/kg and 87 mg/kg of body weight, respectively). MFB-LH coordinates were 4.0 mm posterior to bregma, 1.4 mm lateral from the midline suture and 8.0 mm ventral to the dorsal surface of the skull. VTA coordinates were 2.2 mm anterior to lambda, 1.4 mm lateral from the midline suture and 8.0 mm ventral to the dorsal surface of the skull. Coordinates were determined with the plane passing through bregma and lambda level with the horizontal. The electrodes were placed through small burr holes in the skull and attached perma- nently to the surface with an acrylic platform. After surgery animals received 60,000 units of penicillin (Dual Pen ® ) and were given at least one week for postoperative recovery before behav- ioral testing was begun. Animals were maintained on a 12-hour light/dark cycle, housed individually in stainless steel cages and had ad lib access to food (during the testing period access to water but not to food was restricted according to the experimental design described below).
Animals were trained to respond on a lever manipulandum under a continuous reinforcement (CRF schedule) for rewarding brain stimulation. Training and testing were conducted in standard operant chambers (Grason Stadler, Concord, MA). A constant current stimulator (Sunrise Systems, Pembroke, MA) was used to deliver biphasic symmetrical pulses. Each stimulus consisted of a 500-msec train with a pulse width of 0.2 msec and a delay of 0.2 msec between the positive and negative pulses at a frequency of 160 Hz. When responding for electrical stimulation was estab- lished each subject was then tested at a series of current intensities to determine a complete rate-current intensity function (power function). Each subject was then tested for the balance of the study at a current intensity which resulted in responding at 75% of its maximal rate.
After stable responding for electrical stimulation had been established oral self-administration was begun. In order to induce the rat to readily drink substantial volumes of ETOH a modifica- tion of a technique described by Hubbell et al. (9) was used. With this procedure rats were reported to readily drink up to 2.5 g/kg of ETOH (12% ethanol-5% sucrose v/w solution) in presence of tap water after 22 hours of water deprivation. Food was continuously available during this period.
In our procedure the subjects were deprived of water for 22 hours then tested in the operant chamber for 5 minutes to determine baseline rate of responding for electrical stimulation. Subjects were then returned to their home cages and allowed access to either an ethanol solution (12% ethanol, 5% sucrose v/w) or a control solution for either a maximum of 30 minutes or until a predetermined volume of the solution was consumed. The subjects were then returned to the operant chamber and tested for 35 minutes. The 35-minute test session consisted of 4 signaled (chamber illuminated) 5-minute periods during which each re- sponse on the lever resulted in the delivery of a rewarding brain stimulation, a CRF schedule, interspersed with 3 unsignaled periods (chamber darkened) when no stimulation was delivered. The test session was followed approximately 45 minutes later by a 30 minute period during which each rat had free access to tap water. Ethanol solutions consisted of 12% by volume of 100% ethyl alcohol and 5% by weight of sucrose dissolved in distilled water. Control solutions were either tap water or 5% by weight of sucrose dissolved in distilled water. Under this schedule of fluid deprivation the subjects maintained body weights within 90 to
95% of their starting value throughout the study. The subjects were divided into two experimental groups.
Group 1 ( N = 3 ) was composed of two rats with electrodes implanted in the VTA and one with electrodes implanted in the MFB-LH. This group was tested after drinking either a control solution of tap water or the 12% ETOH-5% sucrose solution.
In Group 2 (N = 3) two subjects had electrodes implanted in the MFB-LH and one had electrodes implanted in VTA. This group was tested after drinking either a control solution of 5 ~ sucrose in distilled water or the 12% ETOH-5% sucrose solution. Animals in both Group 1 and 2 were tested 5 times each week (Monday Friday) for a 6 to 8-week period with ethanol available during the 30-min dosing period on 2 or 3 days and vehicle on the alter- nate days.
Due to variation in the volumes ,~f ETOH-sucrose solutions consumed from one test session to another volumes or doses of similar magnitude were grouped together. For example, if prior to 3 different testing sessions a subject drank 1.90, 2.0 and 2.1 g/kg. respectively, the rates of responding were averaged and the dose then labeled as 2.0 g/kg. This method of data reduction allowed for a reasonable comparison of dose-effect curves between the subjects.
After completion of the behavioral testing the animals were killed with an overdose of Chloropent ® and perfused intracardially with saline. The brains were then removed from the skull, fixed, embedded and sectioned at 40 it. Sections were stained with cresyt violet and Luxol fast blue and examined under a light microscope to determine the site of electrode placement.
RESULTS
All subjects in Group 1 and 2 showed facilitation of response rate for rewarding brain stimulation at low to moderate doses of ETOH (0.4-1.7 g/kg). Figure 1 shows time-effect curves for two doses of ETOH for each subject. Dose-effect curves for the second 5-minute test period during which the most consistent and greatest facilitation of rate was observed in all subjects of both groups are shown in Fig. 2. Doses of 2 g/kg or greater resulted in reduction of response rate from control levels in 3 subjects Itwo from Group l and one from Group 2). The two subjects from Group 2 and one from Group l that did not show a reduction in rate could not be induced to drink greater doses than those shown. These individuals appeared to self-regulate their intake to levels of ETOH below which either their ability to respond was not affected or the rewarding effects of the brain stimulation was not attenuated.
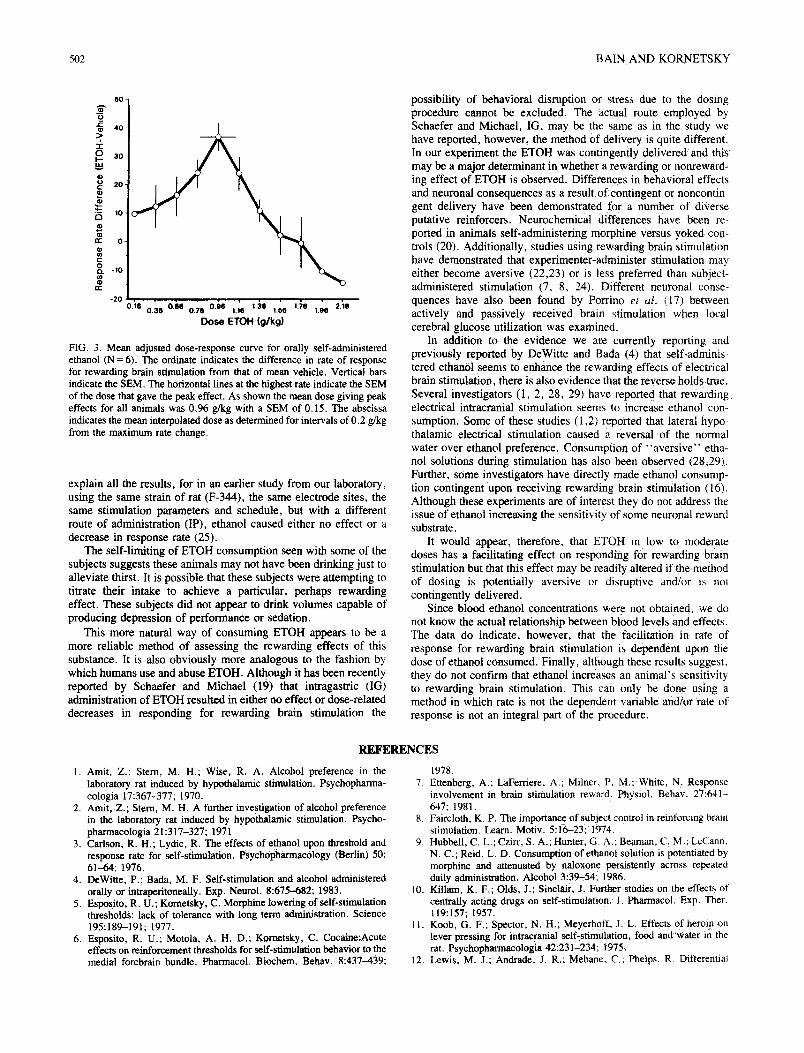
Response rates obtained after consumption of the two control solutions differed to such an extent (H20 x = 210 responses/5 min vs. sucrose x - 2 8 0 responses/5 min) relative to the increases seen after ethanol consumption that these two groups could not be combined for group statistical analysis using only rate of response. However. the data for all animals were combined using as an effect the difference from mean vehicle rate for individual am- mals. respectively. Figure 3 shows the adjusted dose-response curve resulting from equating animals at the point of maximum effect. This gives a mean dose of 0.96 g/kg with a SEM of 0.t5.
A repeated measure ANOVA based on the difference in rates of response between vehicle and each adjusted dose of ethanol (Fig. 3) during Period 2 yielded an F ratio of 13.3 (p<0.001). Test of differences between individual means was carried out using the Newman-Keuls multiple comparison procedure. This analysis indicated that 0.76. 0.96 and 1.16 g/kg of ethanol significantly increased rate of response as compared to vehicle (p<0.01). Also. rate of response after 0.96 g/kg was significantly different than the rate of response after 1.36 and 1.56 g/kg qo<0.01 ). Excluded from analysis was rate of response after doses of 1.76 g/kg and
ETHANOL AND BRAIN STIMULATION 501
< ¢C
300
#50 260 "
. o , ' J~ . ° °
200 ~
150 ,T
• ~'.&Ig.~.
TIME PERIOD
$O0 '
280
200
150
380 -
800 -
250 -
200
1233 O COXmOL
• 2dLi/J~
TIME PERIOD
O CONTROL
• ,..,¢~ #234 • ~,s ,/K#,
35O
#190
300 / ~ LZ. "1,.....
250 ~ 0 CONTR?~ T
20o i i 3 i TIME PERIOD
400
36O
I£ 300
260
850
$ 0 0 -
#261 .)'-..~,
e o % d ' ""
• ldC~.xl
TIME PERIOD
#262
r ~ - - t \ .0i
e""°" ""m ,, L . . . t . . . ~ . . . ~ " 200: o + ...... %
• ;d,~.K.9
TIME PERIOD TIME PERIOD
FIG. l. Individual time-effect curves for two doses of ETOH and vehicle for each animal. Data points indicate response rates, lever presses, for each 5-min test period for control solutions and two doses of ethanol. Group 1 subjects (H20 control) are shown in left panel and Group 2 subjects (sucrose control) are shown in the right panel.
above because of failure of 2 or more of the subjects to ingest that amount of ethanol.
Histological analysis of subjects in Group 1 revealed that the electrode tips were located slightly superior to the MFB within the caudal aspect of the LH in subject No. 50 and slightly superior and within the rostral aspect of the VTA in subjects No. 233 and No. 234, respectively. In Group 2 electrode tips were found to be in the rostral aspect of the VTA in subject No. 190 and in both No. 261 and No. 262 to be slightly dorsal to the MFB and within the caudal aspect of the LH. There were no significant differences in results as a function of electrode placement.
DISCUSSION
The findings of the current study indicate that low to moderate doses of orally self-administered ETOH result in consistent facil- itation of responding (increases in rate) for rewarding electrical stimulation to the brain. These results would suggest that the decreases in self-stimulation behavior observed with similar doses intraperitoneally administered (3, 4, 26) may in part be due to the potentially irritating nature of the ETOH concentrations used (14,
, • i • i • , • i . , • r o 0 . 4 o ,8 1 .2 1 .8 2 2 .4
9/kg ETOIt
35o
300
280
. / e ' * ~ / *
e
#261 Period 2
*:4o.'a ,.~ , . ~ 2.4 g/kg ETOH
200
o J * /
O
#234 Period 2
,< rr
350
300
250
200
#262 Period 2
O / \ • •
\ ° ~ o
0.'4 o.'B I .~ 1 . e ; 2.4 ~ 0.', 0.'. ~.'~ I . ~ ; 2., glkg ETOH glkg ETOH
FIG. 2. Individual dose effect curves for the second 5-min test period (Period 2) for each animal. Data points indicate response rates, lever presses, for the second 5-min test period. Group 1 subjects (H20 control) are shown in left panel and Group 2 subjects (sucrose control) are shown in the right panel.
21, 25, 26, 27). Those reports describing increased self-stimula- tion behavior employing the intraperitoneal (IP) route for the most part involved relatively low doses/concentrations (4, 12, 13) which were probably not irritating. It would then appear that the IP route of ETOH administration is aversive when concentrations are moderate to high and this may then result in "art ifactually"- induced decreases in self-stimulation behavior. Such decreases, therefore, resulting from pain at or near the site of injection and not due to a centrally mediated interaction of ETOH with reward- ing brain stimulation.
Decreases in rates of responding either below vehicle rates or below maximum rates achieved, which were observed as the ETOH dose was increased, may not reflect a decrease in the rewarding effect of the brain stimulation, but rather sedating effects. It has been reported that animals in "near comatose" states resulting from high doses of ETOH have been, after a period of experimenter-delivered stimulation, readily roused from this state and would then crawl to the lever and respond "not dissimilar from the undrugged state" (18).
It could be argued that the increase in response rate seen in the current study may be the result of a simple stimulation action of the ethanol. Although this is a viable hypothesis, it does not
502 BAIN AND KORNETSKY
60- A (D O z 4 0 -
30"
2oi g
10
ee" O"
g t.~ -10. g
-20 o.ie
0 .3S O,7S I. |e 1.5e 1,96
Dose ETOH (g/kgl
FIG. 3. Mean adjusted dose-response curve for orally self-administered ethanol (N = 6). The ordinate indicates the difference in rate of response for rewarding brain stimulation from that of mean vehicle. Vertical bars indicate the SEM. The horizontal lines at the highest rate indicate the SEM of the dose that gave the peak effect. As shown the mean dose giving peak effects for all animals was 0.96 g/kg with a SEM of 0.15. The abscissa indicates the mean interpolated dose as determined for intervals of 0.2 g/kg from the maximum rate change.
explain all the results, for in an earlier study from our laboratory, using the same strain of rat (F-344), the same electrode sites, the same stimulation parameters and schedule, but with a different route of administration (IP), ethanol caused either no effect or a decrease in response rate (25).
The self-limiting of ETOH consumption seen with some of the subjects suggests these animals may not have been drinking just to alleviate thirst. It is possible that these subjects were attempting to titrate their intake to achieve a particular, perhaps rewarding effect. These subjects did not appear to drink volumes capable of producing depression of performance or sedation.
This more natural way of consuming ETOH appears to be a more reliable method of assessing the rewarding effects of this substance. It is also obviously more analogous to the fashion by which humans use and abuse ETOH. Although it has been recently reported by Schaefer and Michael (19) that intragastric (IG) administration of ETOH resulted in either no effect or dose-related decreases in responding for rewarding brain stimulation the
possibility of behavioral disruption or stress due to the dosing procedure cannot be excluded. The actual route employed by Schaefer and Michael, IG, may be the same as in the study we have reported, however, the method of delivery is quite different. In our experiment the ETOH was contingently delivered and this may be a major determinant in whether a rewarding or nonreward- ing effect of ETOH is observed. Differences in behavioral effects and neuronal consequences as a result of contingent or noncontin- gent delivery have been demonstrated for a number of diverse putative reinforcers. Neurochemical differences have been re- ported in animals self-administering morphine versus yoked con- trois (20). Additionally, studies using rewarding brain stimulation have demonstrated that experimenter-administer stimulation may either become aversive (22,23) or is less preferred than subject- administered stimulation (7, 8, 24). Different neuronal conse- quences have also been found by Porrino et at. (17) between actively and passively received brain Stimulation when local cerebral glucose utilization was examined.
In addition to the evidence we are currently reporting and previously reported by DeWitte and Bada (4) that self-adminis- tered ethanol seems to enhance the rewarding effects of electrical brain stimulation, there is also evidence that the reverse holds true. Several investigators (1, 2, 28, 29) have reported that rewarding electrical intracranial stimulation seems to increase ethanol con- sumption. Some of these studies (1,2) reported that lateral hypo- thalamic electrical stimulation caused a reversal of the normal water over ethanol preference. Consumption of "avers ive" etha- nol solutions during stimulation has also been observed (28,29)i. Further, some investigators have directly made ethanol consump- tion contingent upon receiving rewarding brain stimulation (16). Although these experiments are of interest they do not address the issue of ethanol increasing the sensitivity of some neuronal reward substrate.
It would appear, therefore, that ETOH in low to moderate doses has a facilitating effect on responding for rewarding brain stimulation but that this effect may be readily altered if the method of dosing is potentially aversive or disruptive and/or is not contingently delivered.
Since blood ethanol concentrations were not obtained, we do not know the actual relationship between blood levels and effects. The data do indicate, however, that the facilitation in rate Of response for rewarding brain stimulation is dependent upon the dose of ethanol consumed. Finally, although these results Suggest, they do not confirm that ethanol increa~s an animal's sensitivity to rewarding brain stimulation. This can only be done using a method in which rate is not the dependent variable and/or rate of response is not an integral part of the procedure.
REFERENCES
1. Amit, Z.; Stem, M. H.; Wise, R. A. Alcohol preference in the laboratory rat induced by hypothalamic stimulation. Psychopharma- cologia 17:367-377; 1970.
2. Amit, Z.; Stem, M. H. A further investigation of alcohol preference in the laboratory rat induced by hypothalamic stimulation. Psycho- pharmaeotogia 21:317-327; 1971.
3. Carlson, R. H.; Lydic, R. The effects of ethanol upon threshold and response rate for self-stimulation. Psychopharmacology (Berlin) 50: 61-64; 1976.
4. DeWitte, P.; Bada, M. F. Self-stimulation and alcohol administered orally or intraperitoneally. Exp. NeuroL 8:675-682; 1983.
5. Esposito, R. U.; Kometsky, C. Morphine lowering of self-stimulation thresholds: lack of tolerance with long term administration. Science 195:189-191; 1977.
6. Esposito, R. U.; Motola, A. H. D.; Kometsky, C. Cocaine:Acute effects on reinforcement thresholds for self-stimulation behavior to the medial forebrain bundle. Pharmacol. Biochem, Behav. 8:437-439;
1978. 7. Ettenberg, A.; LaFerriere, A.; MiMer, P. M.; White, N. Response
involvement in brain stimulation reward. Physiol. Behav. 27:641-- 647; 1981.
8. Faircloth, K. P. The importance of subject control in reinforcing brain stimulation. Learn. Motiv. 5:16-23; t974~
9. Hubbell, C. L.; Czirr, S. A.; Hunter, G. AI; Beaman, C. M.; LeCann, N. C.; Reid, L. D. Consumption of ethanol Solution is potentiated by morphine and attenuated by naloxone persistently across repeated daily administration. Alcohol 3:39.54; 1986.
10. Killam, K. F.; Olds, L; Sinclair, J. Further studies on the effects of centrally acting drugs on self-stimulation, J. Pharmacol: Exp. Ther. 119:157; 1957.
11. Koob, G. F.; Spector, N. H.; MeyerhofL J. L. Effects of heroin oil lever pressing for inlxacranial self-stimUlation, food and water in the rat. Psyehopbmrmacologia 42:231--234; i975.
12. Lewis, M. J.; Andrade, J. R.; Mebane. C.: Phelps. R Differential
E T H A N O L A N D B R A I N S T I M U L A T I O N 503
effects of ethanol and opiates on BSR threshold. Soc. Neurosci. Abstr. 10:960; 1984.
13. Lorens, S. A.; Sainati, S. M. Naloxone blocks the excitatory effect of ethanol and chlordiazepoxide on lateral hypothalamic self-stimulation behavior. Life Sci. 23:1359-1364;1978.
14. Majchrowicz, E.; Bercaw, B. L.; Cole, W. M.; Gregory, D. H. Nicotinamide adenine dinucleotide and the metabolism of ethanol and acetaldehyde. Q. J. Stud. Alcohol 28:213-224; 1967.
15. Marcus, R.; Kometsky, C. Negative and positive intracranial rein- forcement thresholds: effects of morphine. Psychopharmacologia 38:1-13; 1974.
16. Martin, G. E.; Myers, R. D. Ethanol ingestion in the rat induced by rewarding brain stimulation. Physiol. Behav. 8:1151-1160; 1972.
17. Porrino, L. J.; Esposito, R. U.; Seeger, T. F.; Crane, A. M.; Pert, A.; Sokoloff, L. Metabolic mapping of the brain during rewarding self-stimulation. Science 224:306-309; 1984.
18. Routtenberg, A. Drugs of abuse and the endogenous reinforcement system: the resistance of intracranial self-stimulation behavior to the inebriating effects of ethanol. Ann. NY Acad. Sci. 362:60-66; 1981.
19. Schaefer, G. J.; Michael, R. P. Ethanol and current thresholds for brain self-stimulation in the lateral hypothalamus of the rat. Alcohol 4:209-213; 1987.
20. Smith, J. E.; Co, C.; Freeman, M. E.; Lane, J. D. Brain neurotrans- mitter turnover correlated with morphine-seeking behavior of rats.
Pharmacol. Biochem. Behav. 16:509-519; 1982. 21. St. Laurent, J. Brain centers of reinforcement and the effects of
alcohol. In: Kissen, B. ; Begleiter, H., eds. The biology of alcoholism, volume 2: Physiology and behavior. New York: Plenum Press; 1972: 85-106.
22. Steiner. S. S.; Beer, B.; Shaffer, M. M. Escape from self-produced rates of brain stimulation. Science 163:90-91 ; 1969.
23. Steiner, S. S.; Bodnar, R. J.; Ackermann, R. F.; Ellman, S. J. Escape from rewarding brain stimulation of dorsal brainstem and hypothala- mus. Physiol. Behav. 11:589-591; 1973.
24. Tsang, W. K.; Stutz, R. M. Subject control as a determinant of the reinforcing properties of intracranial stimulation. Physiol. Behav. 32:795-802; 1984.
25. Unterwald, E. M.; Clark, J. A.; Bain, G.; Kometsky, C. The effects of ethanol on rewarding intracranial stimulation: rate and threshold measures. Soc. Neurosci. Abstr. 10:572; 1984.
26. Vrtunski, P.; Murray, R.; Wolin, L. R. The effect of alcohol on intracranially reinforced response. Q. J. Stud Alcohol 34:718-725; 1973.
27. Walgren, H.; Barry, H. Actions of alcohol. Amsterdam: Elsevier Publishing Co.; 1970.
28. Wayner, M. J.; Greenberg, I. Effects of hypothalamic stimulation, acclimation and periodic withdrawal on ethanol consumption. Phys- iol. Behav. 9:737-740; 1972.