Evaluation of microbial diversity in sulfite-added and sulfite-free wine by culture-dependent and -independent methods Masayuki Takahashi, * Tami Ohta, Kazuo Masaki, Akihiro Mizuno, and Nami Goto-Yamamoto National Research Institute of Brewing (NRIB), 3-7-1 Kagamiyama, Higashi-hiroshima, Hiroshima 739-0046, Japan Received 8 May 2013; accepted 15 October 2013 Available online 14 November 2013 The difference in microbiota including non-lactic acid bacteria, non-acetic acid bacteria, and wild yeast during winemaking and in the end-products between sulfite-added and sulfite-free wine, was investigated using polymerase chain reaction-denaturing gradient gel electrophoresis (PCR-DGGE) and a culture-dependent method. There were dif- ferences between the microorganisms detected by PCR-DGGE and those detected by the culture-dependent method, probably because of the selectivity of culture medium and the characteristics of PCR-based method. In both the red wine and white wine, the microbial diversity of the sulfite-added wine was lower than that of the sulfite-free wine during fermentation. Tatumella terrea was detected from the fermenting must by PCR-DGGE and by the culture-dependent method, even though sulfite inhibited its growth to some extent. We confirmed that the addition of sulfite plays an important role in winemaking by inhibiting the growth of unexpected microorganisms, but on the other hand, it was revealed that some microorganisms can survive and grow in sulfite-added fermenting must. We also analyzed 15 samples of commercial wines by the PCR-DGGE method and detected various microorganisms. Among them, Sphingo- monas sp., Pseudozyma sp., Ochromonas sp. and Methylophilus sp. were found for the first time in wine as far as we know. We did not identify a specific microorganism that was detected only from wines without sulfite addition. Thus, the microbiota of end-products seemed to be influenced by other factors, such as filtration before bottling, the production equipment and the storage environment. Ó 2013, The Society for Biotechnology, Japan. All rights reserved. [Key words: Polymerase chain reaction-denaturing gradient gel electrophoresis; Wine; Microbiota; Sulfite; SO 2 ; Tatumella terrea] In the wine industry, the inhibition of growth of unfavorable or unexpected microorganisms is very important to control wine quality and safety. Numerous microorganisms are found on grape skin and on the winemaking environment, e.g., production equip- ment (1e3), and these microorganisms are transferred into the fermenting wine called must (4). However, the growth of almost all microorganisms is inhibited by low pH, the addition of sulfite, and yeast inoculation (5,6). Sulfite has been used as an antimicrobial agent and antioxidant for winemaking for many decades (7). Sulfite is added to wine must before and after vinification as SO 2 gas or K 2 S 2 O 5 , and sulfite inhibits wild yeasts and most spoilage bacteria, especially lactic acid bacteria (LAB). Several research groups have investigated the effects of the addition of sulfite on wild yeasts, LAB, and acetic acid bacteria (AAB) using both culture-dependent and -independent methods (5). However, the effect of sulfite on non- LAB and non-AAB microorganisms is not well known, even though the production of biodynamic or additive-free wine has increased in recent years. Thus, in the present study, we examined the microbiota including the non-LAB and non-AAB in sulfite-added and sulfite-free wine must during fermentation using polymerase chain reaction-denaturing gradient gel electrophoresis (PCR-DGGE) and plate culture method. MATERIALS AND METHODS Strains used We used the commercial strain Saccharomyces cerevisiae Zymaflore F15 (Laffort, Bordeaux, France) for red wine and Zymaflore VL1 (Laffort) for white wine fermentation. Propionibacterium acnes JCM 6425 T was obtained from the Japan Collection of Microorganism (JCM). Wine preparation For bench-scale red wine and white wine fermentation, we used 1.0 kg of Cabernet Sauvignon grapes and 800 ml of juice from Chardonnay grapes, respectively, which were grown in the experimental vineyard of our research institute in Southwestern Japan. For sulfite-added wine musts, 150 mg/l of potas- sium metabisulfite was added to the grape musts, and after the musts were left for 6 h at room temperature, yeast was added to the musts. For the sulfite-free wine musts, yeast was added to the must immediately after the berries were crushed for red wine and juiced for white wine. Red wine musts and white wine musts were fermented at 28 C and 15 C, respectively. The progress of fermentation was fol- lowed with the decrease in weight due to CO 2 release and in density. The ethanol concentration was calculated from decrease in weight and density. We sampled an aliquot of fermenting musts during fermentation every day. In addition, 15 bottles of commercial wine were purchased from the market. DNA extraction We carried out the DNA extraction from wine must following the method described by Renouf et al. (8). A 10 ml of fermenting must was centrifuged, and the pellet was physically disrupted by bead-beating with glass beads (<106 mm dia.; SigmaeAldrich, St. Louis, MO, USA) using a Multi-Beads Shocker (Yasui Kikai, Osaka, Japan). After bead-beating, DNA was extracted using nuclei lysis solution (Promega, Fitchburg, WI, USA) and protein precipitation solution (Promega), and purified with an OneStep PCR Inhibitor Removal Kit (Zymo Research, Orange, CA, USA). For the commercial wine, 100 ml of wine was filtered by a membrane (pore size, 0.20 mm; dia., 47 mm; mixed cellulose ester, Toyo Roshi, Tokyo) to collect microorganisms. DNA was extracted from the shredded membrane by the same method used for the fermenting must. DNA solution was stored at 30 C until use. * Corresponding author. Tel.: þ81 82 420 8227; fax: þ81 82 420 8228. E-mail address: [email protected](M. Takahashi). www.elsevier.com/locate/jbiosc Journal of Bioscience and Bioengineering VOL.117 No. 5, 569e575, 2014 1389-1723/$ e see front matter Ó 2013, The Society for Biotechnology, Japan. All rights reserved. http://dx.doi.org/10.1016/j.jbiosc.2013.10.012
Transcript
www.elsevier.com/locate/jbiosc
Journal of Bioscience and BioengineeringVOL. 117 No. 5, 569e575, 2014
Evaluation of microbial diversity in sulfite-added and sulfite-free wine byculture-dependent and -independent methods
National Research Institute of Brewing (NRIB), 3-7-1 Kagamiyama, Higashi-hiroshima, Hiroshima 739-0046, Japan
Received 8 May 2013; accepted 15 October 2013Available online 14 November 2013
* CorrespondE-mail add
1389-1723/$http://dx.doi
The difference in microbiota including non-lactic acid bacteria, non-acetic acid bacteria, and wild yeast duringwinemaking and in the end-products between sulfite-added and sulfite-free wine, was investigated using polymerasechain reaction-denaturing gradient gel electrophoresis (PCR-DGGE) and a culture-dependent method. There were dif-ferences between the microorganisms detected by PCR-DGGE and those detected by the culture-dependent method,probably because of the selectivity of culture medium and the characteristics of PCR-based method. In both the red wineand white wine, the microbial diversity of the sulfite-added wine was lower than that of the sulfite-free wine duringfermentation. Tatumella terrea was detected from the fermenting must by PCR-DGGE and by the culture-dependentmethod, even though sulfite inhibited its growth to some extent. We confirmed that the addition of sulfite plays animportant role in winemaking by inhibiting the growth of unexpected microorganisms, but on the other hand, it wasrevealed that some microorganisms can survive and grow in sulfite-added fermenting must. We also analyzed 15samples of commercial wines by the PCR-DGGE method and detected various microorganisms. Among them, Sphingo-monas sp., Pseudozyma sp., Ochromonas sp. andMethylophilus sp. were found for the first time in wine as far as we know.We did not identify a specific microorganism that was detected only from wines without sulfite addition. Thus, themicrobiota of end-products seemed to be influenced by other factors, such as filtration before bottling, the productionequipment and the storage environment.
� 2013, The Society for Biotechnology, Japan. All rights reserved.
In the wine industry, the inhibition of growth of unfavorable orunexpected microorganisms is very important to control winequality and safety. Numerous microorganisms are found on grapeskin and on the winemaking environment, e.g., production equip-ment (1e3), and these microorganisms are transferred into thefermenting wine called must (4). However, the growth of almost allmicroorganisms is inhibited by low pH, the addition of sulfite, andyeast inoculation (5,6). Sulfite has been used as an antimicrobialagent and antioxidant for winemaking for many decades (7). Sulfiteis added to wine must before and after vinification as SO2 gas orK2S2O5, and sulfite inhibits wild yeasts and most spoilage bacteria,especially lactic acid bacteria (LAB). Several research groups haveinvestigated the effects of the addition of sulfite onwild yeasts, LAB,and acetic acid bacteria (AAB) using both culture-dependent and-independent methods (5). However, the effect of sulfite on non-LAB and non-AAB microorganisms is not well known, even thoughthe production of biodynamic or additive-free wine has increasedin recent years. Thus, in the present study, we examined themicrobiota including the non-LAB and non-AAB in sulfite-addedand sulfite-free wine must during fermentation using polymerasechain reaction-denaturing gradient gel electrophoresis (PCR-DGGE)and plate culture method.
e see front matter � 2013, The Society for Biotechnology, Japan..org/10.1016/j.jbiosc.2013.10.012
MATERIALS AND METHODS
Strains used We used the commercial strain Saccharomyces cerevisiaeZymaflore F15 (Laffort, Bordeaux, France) for red wine and Zymaflore VL1 (Laffort)for white wine fermentation. Propionibacterium acnes JCM 6425T was obtained fromthe Japan Collection of Microorganism (JCM).
Wine preparation For bench-scale red wine and white wine fermentation,we used 1.0 kg of Cabernet Sauvignon grapes and 800 ml of juice from Chardonnaygrapes, respectively, whichwere grown in the experimental vineyard of our researchinstitute in Southwestern Japan. For sulfite-added wine musts, 150 mg/l of potas-sium metabisulfite was added to the grape musts, and after the musts were left for6 h at room temperature, yeast was added to the musts. For the sulfite-free winemusts, yeast was added to the must immediately after the berries were crushed forred wine and juiced for white wine. Red wine musts and white wine musts werefermented at 28 �C and 15 �C, respectively. The progress of fermentation was fol-lowed with the decrease in weight due to CO2 release and in density. The ethanolconcentration was calculated from decrease in weight and density. We sampled analiquot of fermenting musts during fermentation every day. In addition, 15 bottles ofcommercial wine were purchased from the market.
DNA extraction We carried out the DNA extraction from wine mustfollowing the method described by Renouf et al. (8). A 10 ml of fermenting must wascentrifuged, and the pellet was physically disrupted by bead-beating with glassbeads (<106 mm dia.; SigmaeAldrich, St. Louis, MO, USA) using a Multi-BeadsShocker (Yasui Kikai, Osaka, Japan). After bead-beating, DNA was extracted usingnuclei lysis solution (Promega, Fitchburg, WI, USA) and protein precipitationsolution (Promega), and purified with an OneStep PCR Inhibitor Removal Kit(Zymo Research, Orange, CA, USA). For the commercial wine, 100 ml of wine wasfiltered by a membrane (pore size, 0.20 mm; dia., 47 mm; mixed cellulose ester,Toyo Roshi, Tokyo) to collect microorganisms. DNA was extracted from theshredded membrane by the same method used for the fermenting must. DNAsolution was stored at �30 �C until use.
FIG. 1. PCR-DGGE profile of white wine. The number above each lane indicates the day of fermentation. (AeC) PCR-DGGE profile using GC-341F/531R (eukaryote and prokaryote) (A),GC-NL1/LS2 (eukaryote) (B), and GC-968F/1378R (prokaryote) (C) primer sets. SO2þ, sulfite addition; SO2�, sulfite-free; Sc, Saccharomyces cerevisiae; Vv, Vitis vinifera; Lm, Leuco-nostoc mesenteroides; L, Lactococcus sp.; Tt, Tatumella terrea; Ck, Candida kofuensis; C, Candida sp.; S, Streptococcus sp.; Me, Methylophilus sp.; A, Actinomyces sp.; X, non-sequenceable.
570 TAKAHASHI ET AL. J. BIOSCI. BIOENG.,
PCR amplification PCR was carried out using three sets of primers (1,9e12)(Table S1) in a total volume of 50 ml, containing 1� KOD-plus Neo buffer, 1.5 mMMgSO4, 0.2 mM of each dNTP, 1 U KOD-plus Neo (Toyobo, Osaka, Japan), 0.3 mMeach primer set [GC-341F/531R for universal (prokaryote and eukaryote), GC-NL1/LS2 for eukaryote, and GC-968F/1378R for prokaryote], and 100 ng genome DNA.The thermal cycler program was 96 �C for 3 min, 40 cycles of 96 �C for 15 s, 50 �Cfor 30 s, and 68 �C for 30 s. The size of a PCR amplicon was estimated using100 bp of DNA Ladder Marker (Takara Bio, Shiga, Japan) and 1.2% agarose gelelectrophoresis.
DGGE analysis of PCR amplicon The DGGE analysis was carried out usingthe DCode System (Bio-Rad Laboratories, Hercules, CA, USA). The denaturantgradient gel consisted of 8% acrylamide [acrylamide-HG:N,N0-methylenebis (acryl-amide) eHG ¼ 37.5:1 (Wako Pure Chemical Industries, Osaka, Japan)], 1� TAE andchemical denaturant. The 100% chemical denaturant consisted of 7 M urea (Bio-RadLaboratories) and 40% formamide (SigmaeAldrich). The concentration gradient of
chemical denaturant differed among the primer sets: the concentration gradientranged from 20% to 80% for GC-341F/531R, from 40% to 55% for GC-NL1/LS2, andfrom 40% to 70% for GC-968F/1378R. The electrophoresis was performed at 60 �C and100 V for 16 h. Following electrophoresis, the gel was stained for 30minwith GelRedNucleic Acid Gel Stain (Biotium, Hayward, CA, USA), and the DGGE bands werevisualized using a UV transilluminator (UVP, Upland, CA, USA).
DNA sequencing and data analysis In order to identify the microbial spe-cies, we excised DGGE bands from the gels using a sterilized Pasteur pipette. Theexcised gel plugs were transferred into sterilized PCR tubes and reamplified underthe conditions described above. The PCR amplicons were purified with NucleoSpinExtract II (MachereyeNagel, Düren, Germany) and directly sequenced by an ABIPRISM 310 Genetic Analyzer using a BigDye Terminator v1.1 Cycle Sequencing Kit(Applied Biosystems, Foster City, CA, USA). The sequences were aligned to search theclosest known species using BLAST in the GenBank database (http://blast.ncbi.nlm.nih.gov/) except uncultured/environmental sample sequences. The threshold
FIG. 2. PCR-DGGE profile of red wine. The number above each lane indicates the day of fermentation. (AeC) PCR-DGGE profile using GC-341F/531R (eukaryote and prokaryote) (A),GC-NL1/LS2 (eukaryote) (B), and GC-968F/1378R (prokaryote) (C) primer sets. SO2þ, sulfite addition; SO2�, sulfite-free; Sc, Saccharomyces cerevisiae; Vv, Vitis vinifera; C, Candida sp.;Me, Methylophilus sp.; P, Pediococcus sp.; Ca, Caulobacteraceae; Sz, Schizosaccharomyces sp.; Mi, Microbacterium sp.; Sp, Sphingomonas sp.; Au, Aureobasidium sp.; F, Fungal sp.; Sa,Saccharomyces sp.; B, Brevundimonas sp.; Pa, Propionibacterium acnes; U, unidentified bacterium.
VOL. 117, 2014 MICROBIOTA OF SULFITE-FREE WINE 571
similarity for BLAST identification of a partial sequence was 98%. For the identifi-cation of prokaryote, we analyzed the sequence of V6eV8 region of SSU-rRNA geneat first. When one species was not selected from the sequence of V6eV8 region, weanalyzed the sequence of V3 region. When both of the sequences showed thehighest similarity to one species, the DGGE fragment was identified to this species.For the analysis of eukaryote, we have identified to the genus level with same way,because the sequences obtained from DGGE analysis were too short for thespecies level. For discrimination of Metschnikowia and Candida, genus specificprimers for amplification of ITS region; 18 bases, 50-CCGTAGGTGAACCTGCGG-30; 20bases, 50-CTTAAGTTCAGCGGGTAGTC-30 were used for PCR amplification andsequencing. We identified the detected sequences of plant or organelle of plant asVitis vinifera and eukaryotic sequences derived from thick DGGE bands detectedafter the addition of wine yeast as S. cerevisiae. When some DGGE bands were tooclose from each other to excise separately, the PCR amplicons were cloned using a
Zero Blunt TOPO PCR Cloning Kit for Sequencing (Invitrogen, Carlsbad, CA, USA) andEscherichia coli strain DH5a. Then the insert sequence of each isolate was obtainedfollowing the Cloning Kit and Sequencing Kit instructions from the manufacturer.The accession numbers of sequences obtained through this study are fromAB769285 to AB769301 and from AB846588 to AB846643.
Culture conditions Potato dextrose agar (PDA; BD Difco, Detroit, MI, USA)medium containing 10 mg/ml chloramphenicol was used for the detection of fungi,the plate culture of which was performed at 30 �C for 24 h. Nutrient mediumcomposed of 0.5% peptone, 0.3% beef extract, 0.5% NaCl, 1.5% agar and 10 mg/mlcycloheximide was used for the detection of bacteria, the plate culture of which wasperformed at 37 �C for 48 h. For the detection of LAB, de Man, Rogosa and Sharpeagar (MRS agar; BD Difco) containing 10 mg/ml cycloheximide was used, and theplate culture was performed at 30 �C under anaerobic condition using AnaeroPack(Mitsubishi Gas Chemical, Tokyo) for 1 week. The GAM agar, modified Nissui
TABLE 1. Isolation of bacteria by nutrient medium culture from fermenting must of white wine.
Name (accession no.) Day 1 Day 2 Day 3 Day 4 Day 5 Day 7 Day 11 Day 15 Day 19
Cellulosimicrobium sp. (AB769288) þ þ þ þ þ e e e e
Bacillus megaterium (AB769287) þ þ þ þ þ þ e e e
Lactococcus sp. (AB769293) e þ þ þ þ þ e e e
Paenibacillus sp. (AB769286) þ þ þ þ þ þ e e e
a Calculated by a decrease in weight due to CO2 release and density.b Population of total microorganisms detected by nutrient medium.
572 TAKAHASHI ET AL. J. BIOSCI. BIOENG.,
(Nissui, Tokyo) was used for the detection of P. acnes, the plate culture of which wasperformed at 37 �C under anaerobic condition using AnaeroPack for 1 week. Thefermenting must was diluted 10 times with sterilized water, and then 100 ml of thediluted sample was inoculated onto each agar plate described above. The red winefermenting must was sampled and plated every day during fermentation, and thewhite wine fermenting must sampled and plated at day 1, 2, 3, 4, 5, 7, 11, 15, and 19.The number of colonies was counted, and the CFU per 1 ml was calculated.
Colony isolation and 16S rRNA gene sequence We have chosen the 10colonies per each plate (when colonies were less than 10, all of colonies wereanalyzed) to maximize the diversity based on the shapes and sizes of colonies.Isolated microbes were identified by a homology search of the partial sequence ofthe 16S rRNA gene using BLAST. The 16S rRNA gene of bacteria was amplified bycolony PCR with the 341F/1378R primer set (13) (Table S1) and sequenced using341F or 1378R primer as described above. For identifying eukaryote, the large-subunit rRNA gene of eukaryote was amplified by colony PCR with the NL1/NL4primer set described by O’Donnell (14), and sequenced using NL1 or NL4 primer(Table S1).
Statistical analysis The bands with the same mobility were consideredidentical fragments. DGGE bands were quantified by Image J software (http://rsb.info.nih.gov/ij/), and we analyzed them to determine the relationships among thesamples with R packages (http://www.R-project.org/) and Community EcologyPackage (http://CRAN.R-project.org/package¼vegan).
RESULTS
Analysis of microbiota during fermentation of red and whitewine by PCR-DGGE Judging from alcohol concentration, thefermentation profile of the sulfite-free musts was almost the sameas that of the sulfite-added musts in both the red wine and white
TABLE 2. Isolation of bacteria by nutrient medium
Name (accession no.) Day 1 Day 2 Day 3
Sulfite additionEthanol concentration (v/v%)a 0.0 3.2 6.8cfu/mLb 1.5 � 102 0 0Bacillus cereus (AB769298) þ e e
a Calculated by a decrease in weight due to CO2 release and density.b Population of total microorganisms detected by nutrient medium.
wine. The fermentation of red wine started quickly, but that ofwhite wine was delayed. The alcohol contents of the red and whitewine at the end of fermentation were 14.6 v/v% and 13.3 v/v%,respectively, with or without sulfite.
Various microorganisms were detected from the wine mustswith and without sulfite. Using the same primer sets, the DGGEprofiles show the change of microbiota during fermentation. In thewhite wine (Fig. 1), the use of universal primer set (Fig. 1A) andprokaryote primer set (Fig. 1C) showed that the addition of sulfiteinhibited the growth of Leuconostoc mesenteroides during fermen-tation, and a weak inhibition of the growth of T. terrea by sulfite atday 4e5 was observed. With the eukaryote primer set (Fig. 1B),Candida sp. was detected from day 4 to day 11 in the sulfite-freesamples, but no corresponding DGGE bands were detected in thesulfite-added samples.
In the red wine (Fig. 2) using the prokaryote primer set (Fig. 2C),an unidentified bacterium was detected from day 6 to day 9 in themust without sulfite. The closest relative to this unidentified bac-terium was Pseudomonas sp., but the nucleotide sequence homol-ogy was rather low (approximately 95%). A comparison of Shannonindex calculated from the DGGE profiles of the white wine and redwine revealed that the bacterial diversity of the red wine wasgreater than that of the white wine (Table S2). The use of universalprimer set (Fig. 2A) and eukaryote primer set (Fig. 2B) showed nosignificant difference between sulfite-added and sulfite-freesamples.
culture from fermenting must of red wine.
Day 4 Day 5 Day 6 Day 7 Day 8 Day 9
9.9 11.7 12.7 13.3 13.9 14.60 1.0 � 102 0 0 0 0e þ e e e e
e e e e e e
9.3 11.4 12.7 13.4 14.0 14.60 0 0 0 0 0e e e e e e
TABLE 3. The microorganisms detected from 15 commercial wines by PCR-DGGE.a
Species 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Acinetobacter sp. þ þ þ þ þ þ þ þ þ þ þ þ þ þ þMethylophilus sp. þ þ þ þ þ þ þ þ þ þ þ þ þ þ þSphigomonas sp. e e e e e e þ e e e e e þ þ e
Propionibacterium acnes þ þ þ þ þ þ þ þ þ þ þ þ þ þ þPropionibacterium acnesb þ þ þ þ þ e þ þ þ þ e þ þ e e
Pseudomonas sp. þ þ e þ þ þ þ þ þ þ þ þ þ þ þPediococcus sp. þ e þ e e e þ e e e e e e e e
Enterobacteriaceae þ þ e e þ þ þ þ þ þ þ þ þ þ þAcetobacteraceae þ e þ e e e e e e e e e e e e
Staphylococcus sp. e e þ e e e e e e e e e þ e e
Pseudozyma sp. þ þ e e e e e e e e e e e e e
Candida sp. e þ e e e þ e e þ e e e þ e e
Ochromonas sp. e þ e e þ þ þ e þ e þ þ þ e e
Aureobasidium sp. þ e e e e e e þ e e e e e þ e
Saccharomyces cerevisiae þ e þ þ þ e þ þ þ þ e þ e þ þa Underlined numbers indicate sulfite-free wines. Information of wines used in this study is described in Table S3.b Determined by PCR using a species-specific primer (15).
VOL. 117, 2014 MICROBIOTA OF SULFITE-FREE WINE 573
Principal response curve analysis To compare the effectsof sulfite on the transition of microbiota in red wine and whitewine, we analyzed the DGGE profiles in a principal responsecurve (PRC) analysis. The PRC results for the red wine showedthat the microbiota were influenced by sulfite during fermen-tation, the results for the white wine showed that the micro-biota were strongly influenced by sulfite during an earlier stage(Fig. S1).
Analysis of bacterial diversity using a culture-dependentmethod Since PCR can also amplify DNA derived from deadcells, the microorganisms detected by PCR-DGGE may or may notbe alive. To detect the live microorganisms in fermenting must, weanalyzed all of the fermenting must samples by plate culture(Tables 1 and 2). Using a nutrient medium with cycloheximide forwhite wine, the number of colonies increased until day 7, but nocolony was detected after day 11. The cfu/ml values of the sulfite-free samples were greater than those of the sulfite-addedsamples at an early stage of fermentation (day 4 and 5). The V3to V8 region of the 16S rRNA gene of several colonies selecteddescribed above were amplified by colony PCR, and sequenced.The microorganisms identified were T. terrea, Bacillus sp.,Lactococcus sp. and Paenibacillus sp. (Table 1), and almost all ofisolates identified were T. terrea. If we assume that the colonieswere selected randomly, the detection ratio of T. terrea was over90%.
With the use of the nutrient medium for the red wine, the cfu/ml values of the red wine were lower than that of the white wine.Using the same method for the white wine, we identified the col-onies with the sequence of the 16S rRNA gene (Table 2). The mi-croorganisms inhibited by sulfite were Microbacterium testaceum,Microbacterium phyllosphaerae, Curtobacterium flaccumfaciens,Curtobacterium sp., Bacillus pumilus, and B. megaterium. Micro-bacterium sp. were also detected by PCR-DGGE from both sulfite-added and -free samples, and the results of the plate culturesindicated that Microbacterium spp. died out after the addition ofsulfite and with an increase in the alcohol concentration (Table 2).Since the number of microorganisms detected by nutrient mediumwas very small, we inoculated red wine samples onto PDA agar andMRS agar. Hanseniaspora vineae, Penicillium sp., Aspergillus sp., andAcremonium sp. were detected using PDA agar from the wine mustbefore the addition of S. cerevisiae starter. However, it was impos-sible to elucidate the eukaryotic microbiota after the addition ofwine yeast by plate culture, because S. cerevisiaewas overwhelmingmajority. No microorganisms were detected from MRS agar (datanot shown). We did not carry out PDA andMRS agar plate culture ofwhite wine.
PCR-DGGE analysis of commercial wine We purchased 15bottles of commercial wine (including three sulfite-free wines)from the market (Table S3). We extracted DNA from samples of thewines and analyzed them by PCR-DGGE using the three primer sets(Fig. S2). Although 15 microorganisms including S. cerevisiae weredetected (Table 3), no characteristic microorganism for sulfite-free wine or fruit wine was identified. We quantified the DGGEbands using Image J and analyzed the DGGE profile by principalcomponent analysis (PCA) and cluster analysis using R package,but no influence of sulfite was extracted (data not shown).
Detection of P. acnes P. acnes was detected in all samples ofcommercial wine, as well as in red wine musts by PCR-DGGE. Usinga species-specific primer set (15), P. acnes was detected in 11 out of15 commercial wine samples. We examined whether P. acnes wasdetectable by the culture-dependent method at the same stage offermentation when this bacteria was detected by PCR-DGGE. Wemade a red wine must as described above with the addition of1 ml of P. acnes JCM 6425T bacterial suspension (>106 cfu/ml)into approx. 800 ml of crushed grapes just before the addition ofSO2. We sampled the wine must 3 h after the addition of yeastand every other day during fermentation, and we inoculated thesamples onto GAM agar, but no colony of P. acnes was detectedfrom any samples (data not shown).
DISCUSSION
In recent years, many consumers are concerned for the use offood additives. Thus, the production of additives-free foods such assulfite-free wine is increasing (16). It is known that there is a largevariety of microorganisms (including wild yeast, LAB and AAB) ongrapes, and the addition of sulfite to must inhibits the growth ofLAB (17,18) and wild yeast (19). Thus in the present study, weanalyzed the differences in microbial communities (including non-LAB and non-AAB) between sulfite-added and sulfite-free wineusing a culture-dependent and -independent method (PCR-DGGE).
The PCR-DGGE analysis revealed that the addition of sulfiteinhibited both L. mesenteroides, that grew in fermenting mustwithout sulfite, and Candida sp., that was detected at a middle stageof fermentation. Because certain LAB such as Lactobacillus hilgardii,Pediococcus parvulus and L. mesenteroideswere reported to producebiogenic amines such as cadaverine, putrescine, histamine andtyramine (20,21), it is important to inhibit their growth by addingsulfite. In addition, in the red wine we examined, an unidentifiedbacterium was detected in fermenting must without sulfite at thelater stage of fermentation.
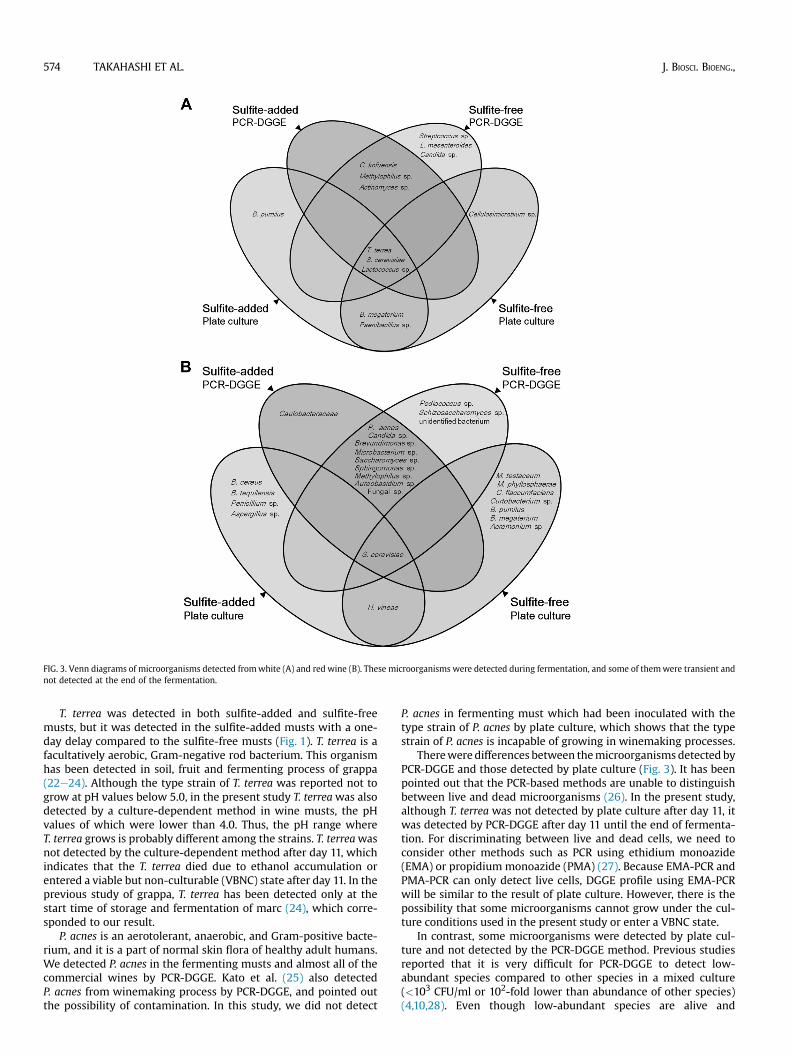
FIG. 3. Venn diagrams of microorganisms detected fromwhite (A) and red wine (B). These microorganisms were detected during fermentation, and some of themwere transient andnot detected at the end of the fermentation.
574 TAKAHASHI ET AL. J. BIOSCI. BIOENG.,
T. terrea was detected in both sulfite-added and sulfite-freemusts, but it was detected in the sulfite-added musts with a one-day delay compared to the sulfite-free musts (Fig. 1). T. terrea is afacultatively aerobic, Gram-negative rod bacterium. This organismhas been detected in soil, fruit and fermenting process of grappa(22e24). Although the type strain of T. terrea was reported not togrow at pH values below 5.0, in the present study T. terreawas alsodetected by a culture-dependent method in wine musts, the pHvalues of which were lower than 4.0. Thus, the pH range whereT. terrea grows is probably different among the strains. T. terreawasnot detected by the culture-dependent method after day 11, whichindicates that the T. terrea died due to ethanol accumulation orentered a viable but non-culturable (VBNC) state after day 11. In theprevious study of grappa, T. terrea has been detected only at thestart time of storage and fermentation of marc (24), which corre-sponded to our result.
P. acnes is an aerotolerant, anaerobic, and Gram-positive bacte-rium, and it is a part of normal skin flora of healthy adult humans.We detected P. acnes in the fermenting musts and almost all of thecommercial wines by PCR-DGGE. Kato et al. (25) also detectedP. acnes from winemaking process by PCR-DGGE, and pointed outthe possibility of contamination. In this study, we did not detect
P. acnes in fermenting must which had been inoculated with thetype strain of P. acnes by plate culture, which shows that the typestrain of P. acnes is incapable of growing in winemaking processes.
Therewere differences between themicroorganisms detected byPCR-DGGE and those detected by plate culture (Fig. 3). It has beenpointed out that the PCR-based methods are unable to distinguishbetween live and dead microorganisms (26). In the present study,although T. terrea was not detected by plate culture after day 11, itwas detected by PCR-DGGE after day 11 until the end of fermenta-tion. For discriminating between live and dead cells, we need toconsider other methods such as PCR using ethidium monoazide(EMA) or propidium monoazide (PMA) (27). Because EMA-PCR andPMA-PCR can only detect live cells, DGGE profile using EMA-PCRwill be similar to the result of plate culture. However, there is thepossibility that some microorganisms cannot grow under the cul-ture conditions used in the present study or enter a VBNC state.
In contrast, some microorganisms were detected by plate cul-ture and not detected by the PCR-DGGE method. Previous studiesreported that it is very difficult for PCR-DGGE to detect low-abundant species compared to other species in a mixed culture(<103 CFU/ml or 102-fold lower than abundance of other species)(4,10,28). Even though low-abundant species are alive and
VOL. 117, 2014 MICROBIOTA OF SULFITE-FREE WINE 575
culturable, they are not detectable in case of coexisting a largeamount of dead cells compared with them by PCR-DGGE, becausePCR-DGGE cannot discriminate between live and dead cells. Withthe use of plate culture that is suitable for their growth, low-abundant species were detectable in a mixed culture. These resultsindicated that the use of multiple methods is necessary to achieve acomprehensive analysis of microbiota.
According to our PRC analysis of the PCR-DGGE band patterns,the influence of sulfite on the microbiota of fermenting must ofwhite wine is strong at the early stage of fermentation, but weak onthe microbiota of fermenting must of red wine at the early stage offermentation. The fermentation speed of the red wine was fasterthan that of the white wine, and the ethanol concentration wasincreased in a short time. Thus, the influence of ethanol concen-tration on microbiota in the fermenting must of red wine was moresignificant than that in the fermenting must of white wine. In thisstudy, because of the delay in the fermentation process of whitewine, sulfite addition seemed to have a significantly greater effecton the microbiota in white wine rather than in red wine.
As both the PCR-DGGE and plate culturing showed, in both redand white wine, the microbial diversity of the sulfite-added wine(including non-LAB, non-AAB and wild yeast) in the fermentingmusts was lower than that of the fermenting musts of the sulfite-free wine. We confirmed that the addition of sulfite plays animportant role in winemaking by inhibiting the growth of unex-pected microorganisms. On the other hand, we revealed that manymicroorganisms were able to survive and grow in fermenting musteven when we added the usual amount of sulfite.
We detected a large variety of microorganisms by PCR-DGGE inthe commercial wines. Although several microorganisms weredetected fromwine and grape skin in previous studies (3,4,8,25,29),Sphingomonas sp., Pseudozyma sp., Ochromonas sp. and Methyl-ophilus sp. were found for the first time in wine as far as we know.Acinetobacter sp.,Methylophilus sp. and P. acneswere detected in allof the samples examined in this study. These microorganisms havea wide distribution in environment (e.g., water, plant, soil). There-fore, the contamination of these microorganisms during wine-making and bottling may occur easily. S. cerevisiae, which can beremoved by the filtration procedure, was not detected in severalwines. In this study, there was almost no difference between themicrobiota of fruit wine made from peach or strawberry and that ofgrape wine. Although it is not clear whether the microorganismswe detected in the wine were alive or not, it is clear that thesemicroorganisms had existed at a certain stage of winemaking. Inthis study, we found no significant difference between the micro-organisms detected in the sulfite-added wine and sulfite-free wine.This result indicates that the microbial diversity in wine is influ-enced not only by sulfite, but also by filtration before bottling,production equipment, and the storage environment.
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.jbiosc.2013.10.012.
References
1. Bae, S., Fleet, G. H., and Heard, G. M.: Lactic acid bacteria associated with winegrapes from several Australian vineyards, J. Appl. Microbiol., 100, 712e727(2006).
2. Costantini, A., Vaudano, E., Del Prete, V., Danei, M., and Garcia-Moruno, E.:Biogenic amine production by contaminating bacteria found in starter prepa-rations used in winemaking, J. Agric. Food Chem., 57, 10664e10669 (2009).
3. Barata, A., Malfeito-Ferreira, M., and Loureiro, V.: The microbial ecology ofwine grape berries, Int. J. Food Microbiol., 153, 243e259 (2012).
4. Nisiotou, A. A., Spiropoulos, A. E., and Nychas, G. J.: Yeast communitystructures and dynamics in healthy and Botrytis-affected grape must fermen-tations, Appl. Environ. Microbiol., 73, 6705e6713 (2007).
5. Andorra, I., Landi, S., Mas, A., Guillamon, J. M., and Esteve-Zarzoso, B.: Effectof oenological practices on microbial populations using culture-independenttechniques, Food Microbiol., 25, 849e856 (2008).
6. Bartowsky, E. J.: Bacterial spoilage of wine and approaches to minimize it, Lett.Appl. Microbiol., 48, 149e156 (2009).
7. Ough, C. S. and Crowell, E. A.: Use of sulfur dioxide in winemaking, J. Food Sci.,52, 386e388 (1987).
8. Renouf, V., Claisse, O., and Lonvaud-Funel, A.: Inventory and monitoring ofwine microbial consortia, Appl. Microbiol. Biotechnol., 75, 149e164 (2007).
9. Muyzer, G., de Waal, E. C., and Uitterlinden, A. G.: Profiling of complex mi-crobial populations by denaturing gradient gel electrophoresis analysis of po-lymerase chain reaction-amplified genes coding for 16S rRNA, Appl. Environ.Microbiol., 59, 695e700 (1993).
10. Cocolin, L., Bisson, L. F., and Mills, D. A.: Direct profiling of the yeast dynamicsin wine fermentations, FEMS Microbiol. Lett., 189, 81e87 (2000).
11. Nubel, U., Engelen, B., Felske, A., Snaidr, J., Wieshuber, A., Amann, R. I.,Ludwig, W., and Backhaus, H.: Sequence heterogeneities of genes encoding16S rRNAs in Paenibacillus polymyxa detected by temperature gradient gelelectrophoresis, J. Bacteriol., 178, 5636e5643 (1996).
12. Heuer, H., Krsek, M., Baker, P., Smalla, K., and Wellington, E. M.: Analysis ofactinomycete communities by specific amplification of genes encoding 16SrRNA and gel-electrophoretic separation in denaturing gradients, Appl. Envi-ron. Microbiol., 63, 3233e3241 (1997).
13. Andrighetto, C., Lombardi, A., Ferrati, M., Guidi, A., Corrain, C., andArcangeli, G.: Lactic acid bacteria biodiversity in Italian marinated seafoodsalad and their interactions on the growth of Listeria monocytogenes, FoodControl, 20, 462e468 (2009).
14. O’Donnell, K.: Fusarium and its near relatives, p. 225e233, in: Reynolds, D. andTaylor, J. (Eds.), The fungal holomorph: mitotic, meiotic and pleomorphicspeciation in fungal systematics. CAB International, Wallingford, UnitedKingdom (1993).
15. Nakamura, M., Kametani, I., Higaki, S., and Yamagishi, T.: Identification ofPropionibacterium acnes by polymerase chain reaction for amplification of 16Sribosomal RNA and lipase genes, Anaerobe, 9, 5e10 (2003).
16. Kurihara, S., Shimoura, S., Maruyama, A., Nakamura, T., and Brennan, M. A.:International difference in consumers’ food safety concern: comparisonbetweenJapan, the United States, China, and Ireland, HortResearch, 64, 67e75 (2010).
17. Carr, J. G., Davies, P. A., and Sparks, A. H.: The toxicity of sulphur dioxidetowards certain lactic acid bacteria from fermented apple juice, J. Appl.Microbiol., 40, 201e212 (1976).
18. Lafon-Lafourcade, S., Carre, E., and Ribereau-Gayon, P.: Occurrence of lacticacid bacteria during the different stages of vinification and conservation ofwines, Appl. Environ. Microbiol., 46, 874e880 (1983).
19. Romano, P. and Suzzi, G.: Sulphur dioxide and wine microorganisms, pp.373e393, in: Fleet, G. H. (Ed.), Wine microbiology and biotechnology. HarwoodAcademic Publishers, Chur, Switzerland (1993).
20. Landete, J. M., Ferrer, S., and Pardo, I.: Which lactic acid bacteria areresponsible for histamine production in wine? J. Appl. Microbiol., 99, 580e586(2005).
21. Pircher, A., Bauer, F., and Paulsen, P.: Formation of cadaverine, histamine,putrescine and tyramine by bacteria isolated from meat, fermented sausagesand cheeses, Eur. Food Res. Technol., 226, 225e231 (2006).
22. Kageyama, B., Nakae, M., Yagi, S., and Sonoyama, T.: Pantoea punctata sp.nov., Pantoea citrea sp. nov., and Pantoea terrea sp. nov. isolated from fruit andsoil samples, Int. J. Syst. Evol. Microbiol., 42, 203e210 (1992).
23. Brady, C. L., Venter, S. N., Cleenwerck, I., Vandemeulebroecke, K., De Vos, P.,and Coutinho, T. A.: Transfer of Pantoea citrea, Pantoea punctata and Pantoeaterrea to the genus Tatumella emend. as Tatumella citrea comb. nov., Tatumellapunctata comb. nov. and Tatumella terrea comb. nov. and description of Tatu-mella morbirosei sp. nov., Int. J. Syst. Evol. Microbiol., 60, 484e494 (2010).
24. Maragkoudakis, P. A., Nardi, T., Bovo, B., D’Andrea, M., Howell, K. S.,Giacomini, A., and Corich, V.: Biodiversity, dynamics and ecology of bacterialcommunity during grape marc storage for the production of grappa, Int. J. FoodMicrobiol., 162, 143e151 (2013).
25. Kato, S., Ishihara, T., Hemmi, H., Kobayashi, H., and Yoshimura, T.: Alter-ations in D-amino acid concentrations and microbial community structuresduring the fermentation of red and white wines, J. Biosci. Bioeng., 111,104e108 (2010).
26. Cocolin, L., Rantsiou, K., Iacumin, L., Zironi, R., and Comi, G.: Moleculardetection and identification of Brettanomyces/Dekkera bruxellensis and Bretta-nomyces/Dekkera anomalus in spoiled wines, Appl. Environ. Microbiol., 70,1347e1355 (2004).
27. Nocker, A., Cheung, C. Y., and Camper, A. K.: Comparison of propidiummonoazide with ethidium monoazide for differentiation of live vs. dead bac-teria by selective removal of DNA from dead cells, J. Microbiol. Methods, 67,310e320 (2006).
28. Prakitchaiwattana, C. J., Fleet, G. H., and Heard, G. M.: Application andevaluation of denaturing gradient gel electrophoresis to analyse the yeastecology of wine grapes, FEMS Yeast Res., 4, 865e877 (2004).
29. Ruiz, P., Sesena, S., Izquierdo, P. M., and Palop, M. L.: Bacterial biodiversityand dynamics during malolactic fermentation of Tempranillo wines as deter-mined by a culture-independent method (PCR-DGGE), Appl. Microbiol. Bio-technol., 86, 1555e1562 (2010).