COMPARISON TO BACILLUS THURINGIENSIS ON THE SURVIVAL AND
FEEDING PREFERENCES OF OSTRINIA NUBILALIS
by
Kelsey E. Fisher
A thesis submitted to the Faculty of the University of Delaware in partial fulfillment of the requirements for the degree of Master of Science in Entomology
INFORMATION TO ALL USERSThe quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscriptand there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States Code
Published by ProQuest LLC (2016). Copyright of the Dissertation is held by the Author.
ProQuest Number: 10014932
EVALUATION OF NATURAL PLANT CHEMICAL DEFENSES IN
COMPARISON TO BACILLUS THURINGIENSIS ON THE SURVIVAL AND
FEEDING PREFERENCES OF OSTRINIA NUBILALIS
by
Kelsey E. Fisher
Approved: ____________________________________________________________ Charles E. Mason, Ph.D. Professor in charge of thesis on behalf of the Advisory Committee Approved: ____________________________________________________________ Jacob L. Bowman, Ph.D. Chair of the Department of Entomology and Wildlife Ecology Approved: ____________________________________________________________ Mark W. Rieger, Ph.D. Dean of the College of Agriculture and Natural Resources Approved: ____________________________________________________________ Ann L. Ardis, Ph.D. Interim Vice Provost for Graduate and Professional Education
ACKNOWLEDGMENTS
I would like to thank my advisor, Dr. Charles Mason, for his on-going support
and encouragement through the duration of my degree. I truly enjoyed the many
conversations that jumpstarted a majority of the ideas for this thesis. I would like to
acknowledge Dr. Lindsey Flexner, for his constant interest and enthusiasm. Without him,
this project would not have been possible. I would also like to thank Dr. Judith Hough-
Goldstein for her knowledge and comments that improved this thesis. Furthermore, I
would like to acknowledge Dr. John McDonald and Zaiqi Pan for their abundant
knowledge and aid with my statistical analyses.
I would like to thank a few people for technical and logistical support. First, Keith
Bidne at Iowa State University, for maintaining a colony of Z-race European corn borers
and sending weekly shipments of eggs and 3rd instars. Additionally, I would like to thank
Bill Bartz, Rodney Dempsey, and the University of Delaware greenhouse staff for
making it possible for me to conduct experiments in the greenhouse and providing the
necessary materials. I would like to thank Scott Hopkins, Bill Cissel, and the University
of Delaware farm staff for helping set up my field experiments and teaching me about
irrigation. Next, I would like to thank Holly Walker, David Ingber, Maddie Chura,
Jessica Faucher, and Michael Palmer for help executing my many experiments.
Last, but certainly not least, I would like to thank all of my family and friends for
their understanding and encouragement.
TABLE OF CONTENTS LIST OF TABLES ............................................................................................................. vi LIST OF FIGURES .......................................................................................................... vii ABSTRACT ....................................................................................................................... ix Chapter
1 LITERATURE REVIEW ............................................................................1 Feeding Behavior Overview ........................................................................1 Agriculture Pest Behavior ............................................................................5 The European Corn Borer ............................................................................8
2 SURVIVORSHIP OF THE EUROPEAN CORN BORER ON A RANGE OF HOST PLANTS VARYING IN DEFENSIVE CHEMISTRY ...........15
Introduction ................................................................................................15 Materials and Methods ...............................................................................17
Insects and Plants ...........................................................................17 Growth Chamber ............................................................................20
Neonate Infestation ............................................................20 Third Instar Infested ...........................................................20
Neonate Infestation ............................................................26 Third Instar Infestation ......................................................27
Field ...............................................................................................27
Discussion ..................................................................................................29 3 EUROPEAN CORN BORER PLANT CONSUMPTION RATES AND
PREFERENCE AMONG A RANGE OF AGRICULTURAL HOST CROP SPECIES ........................................................................................43
Introduction ................................................................................................43 Materials and Methods ...............................................................................45
Insects and Plants ...........................................................................45 Consumption Rates and Preference ...............................................47 Statistical Analysis .........................................................................49
REFERENCES ..................................................................................................................58 Appendix A TEST FOR INDUCED DEFENSES IN CUCUMBER .............................66
LIST OF TABLES
Table 1 Average weight of mid-development larvae or pupae under all experimental designs. Letters indicate significantly different weights analyzed with two-way ANOVAs (P < 0.05)............................................36
LIST OF FIGURES Figure 1 Survival curve for neonate-infested growth ghamber feeding trials.
Survival curves based on the longevity of larvae fed various host plants determined by degree-days accumulated until death prior to pupation. Lines for hosts followed by the same letter did not differ in survival time (Tukey-Kramer mean separation test (P < 0.05)........................................37
Figure 2 Growth chamber survival through the first stadium of treatment.(A) Neonates or (B) third instars fed one of various host plants (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, T = tomato) through the duration of development. Bars represent the average percent of the individuals that survived to the subsequent instar based on degree-day accumulation (106 degree-days = 2nd instar; 132 degree-days = 4th instar). Bars within each graph with different letters are significantly different (Tukey-Kramer test, P < 0.05). Error bars represent SEM.........38
Figure 3 Survival curve for third instar-infested growth chamber feeding trials. Survival curves based on the longevity of larvae fed various host plants determined by degree-days accumulated until death prior to pupation. Lines for hosts followed by the same letter did not differ in survival time (Tukey-Kramer mean separation test (P < 0.05)........................................39
Figure 4 Greenhouse recovery. (A) Neonate infested plants (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) were dissected after 355 degree-days for mid-development recovery or (B) after 790 degree-days for pupal recovery. (C) Plants
infested with 3rd instars were dissected after 567 degree-days for pupal recovery. Bars represent the number of individuals recovered at each dissection with SEM. Different letters within one graph are significantly different (P < 0.05).....................................................................................40
Figure 5 Field recovery. (A) Neonate infested plant groupings (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) were dissected after 355 degree-days for a mid-development recovery or (B) after 790 degree-days for pupal recovery. (C) Plant groupings infested with 3rd instars were dissected after 567 degree-days for pupal recovery. Overall, recovery was very low; none of the experiments yielded significant results......................................................41
Figure 6 Pupal weight comparison from third instar infested plants in the
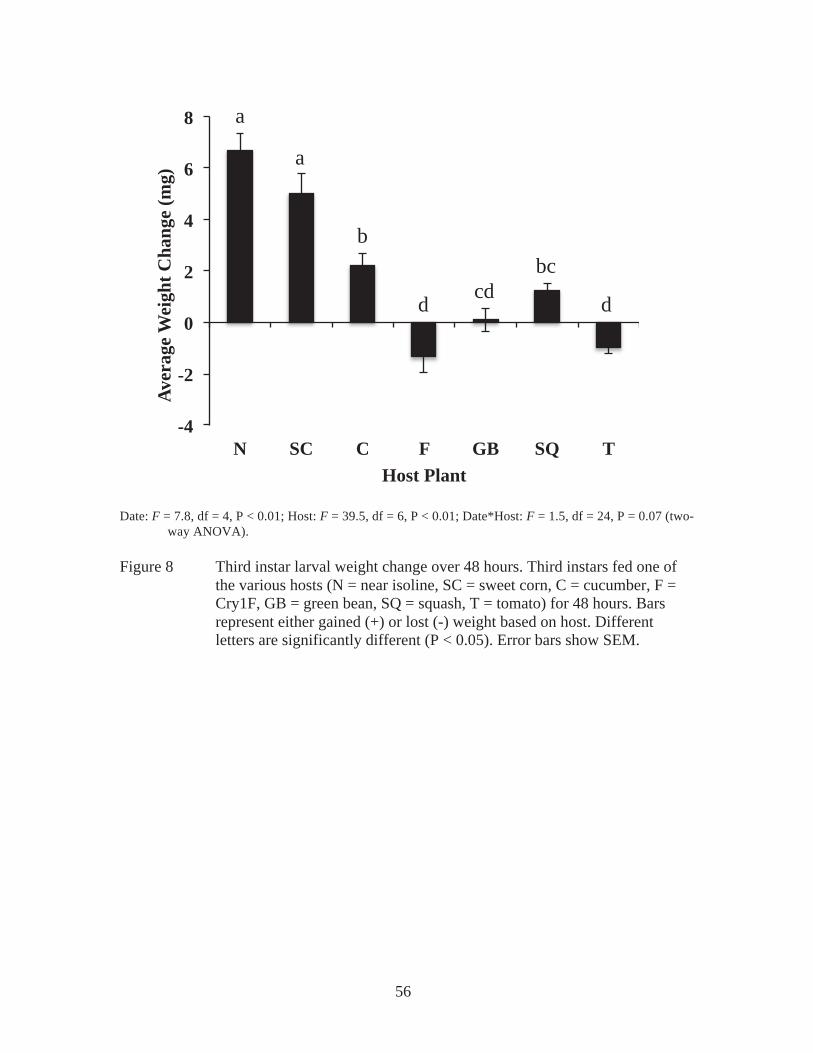
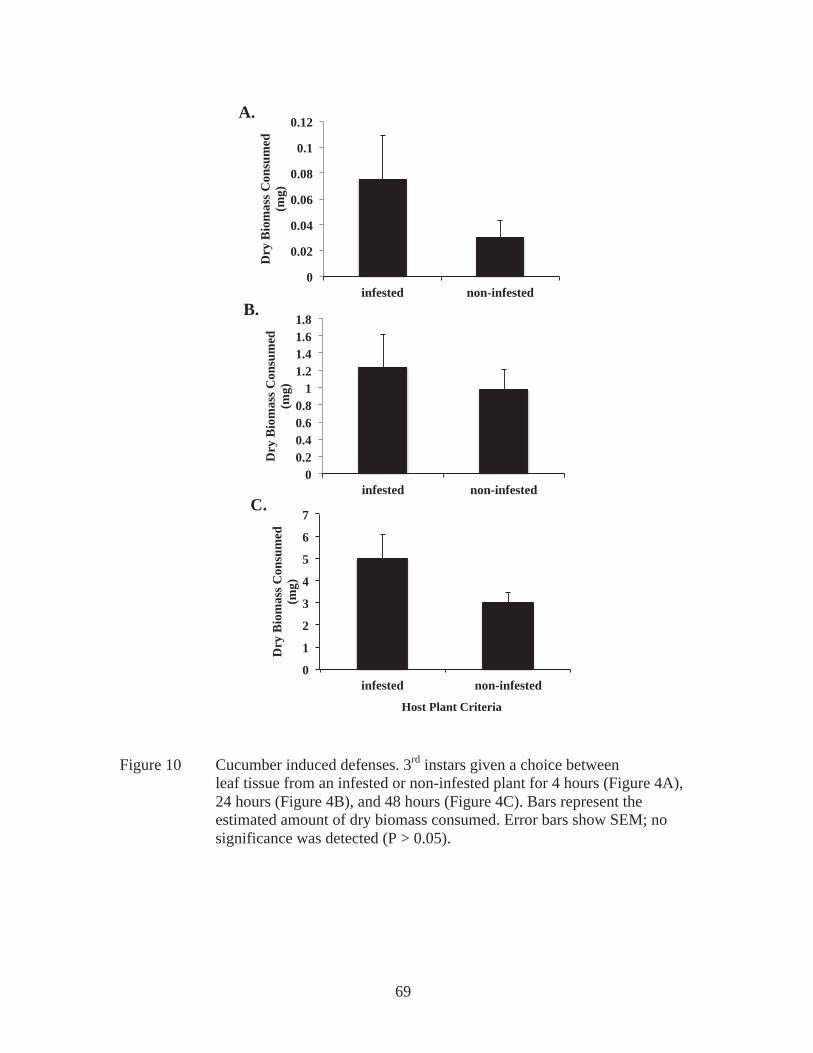
field. Average weight of pupae recovered from 3rd instar infested field experiments from non-Bt corn (sweet corn and near isoline) (n = 7) and tomato (n = 8). Significant difference represented by different letters (P < 0.01). Error bars represent SEM................................................................42 Figure 7 Dry biomass consumed. Third instars fed one of the various hosts (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) for 48 hours. Bars represent the estimated amount of dry biomass consumed. Different letters are significantly different (P < 0.05). Error bars show SEM................................................55 Figure 8 Third instar larval weight change over 48 hours. Third instars fed one of the various hosts (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) for 48 hours. Bars represent either gained (+) or lost (-) weight based on host. Different letters are significantly different (P < 0.05). Error bars show SEM..........56 Figure 9 Preference determined by dry biomass consumed. Third instars given a choice between 2 host plants (near isoline (N) and either sweet corn (SC), Cry1F (F), tomato (T), green bean (GB), or cucumber (C), which are presented in figure 3A, or Cry1F (F) and tomato (T), presented in figure 3B) for 48 hours. Bars represent the estimated dry biomass consumed. Significant differences, within one group are distinguished with an asterisk (*). Error bars show SEM..........................................................................57 Figure 10 Cucumber induced defenses. 3rd instars given a choice between leaf tissue from an infested or non-infested plant for 4 hours (Figure 4A), 24 hours (Figure 4B), and 48 hours (Figure 4C). Bars represent the estimated amount of dry biomass consumed. Error bars show SEM; no significance was detected (P > 0.05)..............................................................................69
ABSTRACT
The European corn borer (ECB), Ostrinia nubialis (Hübner), is an herbivorous
pest of over 200 documented hosts, especially corn (Zea mays L.), in the United States
and Canada, and had been labeled a “generalist herbivore.” The feeding behavior of
insects, including ECB, depends on perception of multiple physical and chemical features
presented by the plant and the insect’s response to these cues. Recently, in the United
States there has been little evidence of ECB presence outside of cornfields. Generalist
insect species have the ability to evolve over time into specialists when the focal plant is
common, predictable and abundant. All of these criteria are met by corn in the United
States.
This study investigated the growth and survivorship of ECB on a range of hosts
that vary in defensive chemistries and their preferences among these hosts (sweet corn,
Cry1F Bt corn (maize), non-Bt near isoline maize, cucumber, squash, tomato, and green
bean). Experiments were conducted in the growth chamber, greenhouse, and field to
determine survival under different conditions. Survival was significantly higher on non-
Bt corn hosts than any other host provided. Choice feeding assays were conducted to
determine preference based on biomass consumption. Results generally supported the
expected outcome with greater consumption of non-Bt corn hosts. From the results on
survival and preference, it can be concluded that non-Bt corn is the most suitable host
plant for ECB, thus strongly supporting ECB corn specialization.
1
Chapter 1
LITERATURE REVIEW
Feeding Behavior Overview
There are complex ecological and evolutionary relationships between herbivorous
insects and the plants they consume. Many factors affect the feeding behaviors in a plant-
insect relationship. Plants defend vegetative tissues against herbivorous predators by
mechanical and chemical mechanisms (Ehrlich and Raven 1964; Mooney et al. 2009).
Insects must overcome these defenses in order to grow and reproduce. In turn, damage
from herbivores affects photosynthesis and growth of plants, causing impacts on plant
reproduction (Futuyma and Gould 1979; Niesenbaum 1996).
Most herbivorous insects have host plant ranges. These are plants the insects are
capable of consuming to obtain adequate nutrition for growth and reproduction.
Additionally, these plants often provide a protective habitat when the herbivorous insect
is able to overcome the plant defenses (Ehrlich and Raven 1964). Typically, when
referring to a host plant, it is either a plant family or genus, not usually a single species
(Forister et al. 2015). A topic that has long been debated pertaining to herbivorous
insects’ host range is the generalist versus specialist argument including monophagy,
oligophagy, and polyphagy. Monophagous insects have one or few closely related
acceptable host plants. Many of these scenarios occur within Lepidoptera, Hemiptera, and
Coleoptera (Schoonhoven et al. 2005). An example of monophagy is the Monarch
Butterfly larvae’s ability to only feed on milkweed species (Van Zandt and Agrawal
2004). Oligophagous insects can feed on species limited to one family or genus, and
polyphagous insects can feed on many plants represented across several families.
2
Polyphagous insects will accept many host plants and rarely exercise choice
(Schoonhoven et al. 2005). An argument comes in when claims are made about generalist
and specialist herbivores. Generalists, typically oligophagous or polyphagous, can feed
on a wide array of plant types and are able to metabolize a multitude of plant defenses
(Schoonhoven et al. 2005). Specialist insect herbivores have the ability to tolerate
specific plant defenses and manipulate hosts to their benefit to reduce predation and
parasitism (Jaenike 1990). It has been documented that less than 10% of herbivores feed
on more than three different plant families (Bernays and Graham 1988). Based on these
definitions, specialists are much more common than generalists. According to a meta-
analysis by Ali and Agrawal (2012), the identification of generalist or specialist is still
unclear because of experimental limitations. Often researchers place insects into self-
determined categories based on their own definitions of generalist and specialist. There is
often more complexity in the plant-insect world, and the answer may not be as simple as
two categories.
Host plants can provide an assortment of nutrients and defenses in a number of
ways. Due to this variation, the host species may affect larval success by influencing rates
of growth and development (Singer et al. 2002). Nitrogen is a major component of
proteins and amino acids, which are sources for arthropod growth (Chen et al. 2009).
Higher plant nitrogen content correlates with greater growth, higher conversion of
ingested food into energy, and shorter developmental time (Slansky and Feeny 1977;
Mattson 1980; Woods 1999; Chen et al. 2008, 2009). Greater growth is adaptive because
it has been shown that larger insects perform better (Kingsolver and Huey 2008). Higher
conversion of ingested food into energy means that the insect would be able to consume
3
smaller quantities of food in order to obtain optimal energy, leaving more time to be
allocated to other activities and less time exposed to predators and parasitoids. In a study
on tobacco hornworm larvae, it was shown that when a diet is less nutrient rich, the
larvae feed longer in order to maintain their intake of nutrients (Timmins et al. 1988).
Furthermore, a shorter development time would cause earlier pupation, allowing less
opportunity for predation or parasitism (Benrey and Denno 1997) Additionally, a shorter
developmental time can increase fitness by increasing the number of generations per
year.
In addition to host plants having a direct impact on larval fitness based on
nutrition, there is an impact due to plant defenses. Plants have two types of biochemistry
that occur within their tissues. The basic chemical processes required for day-to-day
functions are classified as primary plant metabolism. This chemistry is involved in the
formation and breakdown of a limited set of chemicals to create proteins, carbohydrates,
lipids, and nucleic acids (Buchanan et al. 2000). Secondary chemistry is involved in plant
reproduction, insect attractants, and insect deterrents (Ehrlich and Raven 1964). These,
along with physical barriers like trichomes and leaf toughness, play a key role in
manipulating insect behavior (Wink 1988). Secondary chemistry develops as derivatives
from primary biosynthesis. Plant chemical attractants and deterrents can be present in
glandular trichomes, and in epicuticular waxes as volatile compounds, or stored in the
plant cells. Attractants play a small roll in host selection for herbivorous insects, as
continual feeding is highly influenced by deterrent chemistries (Jermy 1976). Some
deterrents are always present in the plant tissues, while others can be induced by an insect
behavior such as herbivory or oviposition (Schoonhoven et al. 2005).
4
An abundance of studies have been conducted to determine host plant preference.
Many analyze host plant range (Kogan and Goeden 1970), preference in the presence of
additive toxins (Davis and Coleman 1997; Gore et al. 2005), or pest preference for other
crops or weeds when the major host is not present (Losey et al. 2001; Tate et al. 2006).
Others have been conducted for specific analyses like allelochemical or nitrogen
abundance’s affect on feeding behavior (Glendinning and Slansky 1994; Elasyed 2011).
It has also been shown that herbivorous insects can detect host plant quality and feed or
oviposit preferentially on plants containing more nitrogen (White 1984; Fox et al. 1990;
Prudic et al. 2005; Chen et al. 2008).
There is a basic overview of insect feeding behavior that is widely accepted for
most chewing, herbivorous insects. First, the insect is hungry, which causes search
behavior as a random “walk” until the insect detects a plant cue. The term “walk” is used
for simplicity purposes. Most insects can use multiple modes of mobility including
jumping and flying, which are actions by insects with wings or strong legs like
grasshoppers or beetles. Other insects are less mobile, such as larval Lepidoptera. To
search for plant cues, less mobile insects use walking, crawling, or ballooning: a form of
movement where Lepidoptera larvae produce silk and hang from the plant resulting in
dropping below or floating away with a gust of wind (Zalucki et al. 2002; Goldstein et al.
2010). For Lepidoptera larvae, some of the searching behavior is shortened because these
individuals are placed on an acceptable host plant by adult oviposition. Nevertheless,
sometimes the larvae reject their natal plant in search of a new one. Insect mobility
provides potential for insects to switch host plant species one or more times (Singer et al.
2002). Once the insect detects a plant cue the “walk” becomes directional toward the
5
stimulus. Eventually there is contact between the plant and the insect, initiating selection
behavior. These behaviors include physical and chemical evaluation of the plant that
could not occur from a distance. Some examples are scratching, drumming with tarsi,
palpating, ovipositor dragging, and test biting. Test bites are often smaller than a regular
bite. The plant material may be kept longer in the pre-oral cavity than during regular food
intake due to gathering sensory information. Finally, continual feeding or discontinuation
of feeding and leaving the plant confirms host plant acceptance or rejection. (Elsayed
2011).
It is possible for generalist insect species to evolve over time into specialists
(Forister et al. 2015). Host plant specialization typically occurs where plants are
common, predictable, and abundant, making mating and oviposition site location easier
(Jaenike 1990). Host specialization allows the insect to gain ability to tolerate plant
defenses, manipulate hosts to their benefit, and evolve ways to reduce predation and
parasitism (Ali and Agrawal 2012). This also typically occurs when one host plant is
superior in many ways to other host plants in terms of nutrient availability, abundance,
and predation risk (Ehrlich and Murphy 1988).
Agriculture Pest Behavior
The agriculture industry produces the majority of food sources for humans.
According to the USDA National Agricultural Statistics Service for crop production, in
2014, United States farmers produced 149 billion dollars worth of crops. One of the most
abundant crops was corn (Zea mays L.), producing 84 million acres worth 63.9 billion
dollars. Stable monocultures of crop plants are grown in the United States because it is a
6
profitable platform for farmers and easy to make control decisions. A monoculture
consists of a single plant species over a wide area of land. However, monocultures can be
difficult to manage due to herbivorous agricultural pests exploiting the crop resources.
Monocultures are easy for herbivores to locate because of the amplification of plant cues
released. Pests are attracted to monocultures for reasons that can be explained by the
resource concentration hypothesis and the enemy free hypothesis. The resource
concentration hypothesis refers to the abundance of food sources available for an insect
and its future offspring. All of the plants in the area are healthy host plants (Grez and
Gonzalez 1995). The enemy free hypothesis states that due to the lowered plant diversity
the insect diversity is decreased as well. Typically, there are fewer natural enemies
present in agriculture fields to feed on or parasitize pest species (Letourneau 1987).
In addition to the enemy free and resource concentration hypotheses, plant
breeding plays a large role in herbivorous insects attraction to crops grown in
monoculture. Over time, plant breeders have selected for the highest yielding, largest, and
most palatable varieties. In order to do this, plants’ energy is directed toward growth
rather than defense (Rosenthal and Dirzo 1997). While selecting for large, high yielding
of cucurbitacin, a bitter deterrent that is specifically detected and avoided by ovipositing
ECB (Tallamy et al. 1997). Cucurbitacin content of squash is 0.02mg/g of fresh weight in
the leaves, while cucumber contains trace amounts (Metcalf et al. 1982). Cucurbit plants
were tested for larval feeding to see if results would coincide with results from ECB
oviposition studies where cucurbitacin was avoided (Tallamy et al. 1997).
19
Favorable host plants were classified as those with minimal or lacking known
defensive characteristics. These included non-Bt near isoline corn, sweet corn, and green
bean. Seedling corn contains high concentrations of DIMBOA (2,4-Dihydroxy-7-
methoxy-1,4-benzoxazin-3-one), a compound that prevents feeding and is toxic to ECB
neonates until corn reaches vegetative stage 5 (V5) (Guthrie et al. 1986). Vegetative stage
6 (V6) corn was utilized for experiments to avoid effects of DIMBOA. Near isoline corn
is genetically identical to Cry1F, except without the Bt toxin, and it is used commonly as
refuge corn. Sweet corn was tested to determine if it has greater suitability for ECB
compared to near isoline. Green bean does not provide many toxins that directly affect
ECB (Webb et al. 1987). ECB adults have been associated with green bean because it
offers a proper environment for rest during the day and mating at night, and adults are
known to oviposit on green bean (Mason et al. 1996). In addition, ECB even in low
densities can have a large economic impact on the green bean industry causing entire
fields to be rejected if one larva is found in the harvested beans.
Plants used in growth chamber and greenhouse experiments were grown in the
greenhouse maintained at 20-30°C, 40-60% humidity, and 16:8 L:D supplemented during
short day lengths. Plants for field experiments were seeded into the ground, except for
tomato, which consisted of two-week old seedlings. All plants were grown to
approximately 1000 accumulated degree-days to correspond with corn at V6. At this
point in development, plants were used to initiate experiments or their leaves were
excised to feed ECB.
20
Growth Chamber
Neonate Infestation
Neonate ECB hatched from egg masses in a plastic bag and were moved via their
silk using a fine brush into individual 37 mL Solo® plastic cups (Lake Forest, IL, USA).
Each cup contained approximately five cm2 of leaf tissue of one of the host plants and a
dry cotton wick to collect excess moisture. Both leaf material and cotton wicks were
changed daily. Seventy-five replicates were grouped into three blocks of 25 by date. Cups
were maintained in the growth chamber at the same growth chamber conditions described
previously. Each day, survival was recorded. At mid-development (355 degree days since
hatch), larvae were weighed. Individuals were monitored for degree-day accumulation to
pupation and pupal weight was recorded. Degree-day accumulation at death was recorded
for those that did not survive to pupation.
Third Instar Infestation
Third instar larvae were moved from the gregarious, ECB diet container into
individual 37 mL Solo® plastic cups. Each cup contained approximately five cm2 of leaf
tissue of one of the host plants and a dry cotton wick to collect excess moisture. Both leaf
material and cotton wicks were changed daily. Seventy-five replicates were grouped into
three blocks of 25 by date. Cups were maintained in the growth chamber at the same
conditions described previously. Each day, survival was recorded. Individuals were
monitored for degree-day accumulation to pupation and pupal weight was recorded.
Degree-day accumulation to death was recorded for individuals that did not survive to
pupation.
21
Greenhouse
Neonate Infestation
To infest whole plants with neonates, two blackhead stage egg masses were
placed in 1.5 mL microcentrifuge tubes (Fisher Scientific®, Waltham, WA, USA) and
attached to the plants with a paper twist tie (Staples®, Framingham, MA, USA).
Additional sweet corn plants were infested with neonates to serve as larval developmental
monitors after infestation. Plants were covered with a cage created from thin mesh fabric
(JoAnn Fabric®, Hudson, OH, USA) and either 84 cm or 107 cm galvanized steel wire
round tomato cages (Lowes®, Mooresville, NC, USA) based on host plant size to prevent
larval movement away from the plant. Using a split design, infested plants were either
dissected at mid-development (355 degree-days of development) or after estimated
pupation (790 degree-days of development). Ten plants were dissected over 5 dates for
mid-development larvae and eight plants were dissected over 4 dates for pupae on each
host type. The number of individuals collected per plant at dissection and their weights
were recorded. If larvae were recovered from plants at 790 degree-days, they were kept
with stalk/stem tissue in the growth chamber until pupation.
Third Instar Infestation
For 3rd instar experiments, five individuals were placed on different leaves of the
plant to prevent crowding effects. Additional sweet corn plants were infested with 3rd
instars to be dissected prior to experimental dissections to observe ECB development.
Subsequent to infestation, plants were covered with a cage created from thin mesh fabric
and either 84 cm or 107 cm galvanized steel wire round tomato cages based on host plant
size to prevent larval movement. Eight plants were dissected over 4 dates for pupae (567
22
degree-days of development after infestation) on each host type. The number of
individuals collected per plant at dissection and their weights were recorded. If larvae
were recovered, they were kept with stalk/stem tissue in the growth chamber until
pupation.
Field
The design for field experiments corresponded with the greenhouse experiments.
Field plots were created from small three-row clusters of plants of the same species with
three plants in each row (nine plants total). Neighboring plants were spaced 30 cm apart.
Plants were seeded into the ground, except for tomato, which consisted of two-week old
seedlings. If seeds failed to germinate in the field, back-up seedlings were transplanted
into empty spaces. If those failed, plants were transplanted with similar degree-day
accumulation from weekly greenhouse plantings. Groupings were arranged in a complete
random block design for the seven host plants with 45cm of space between groupings.
There were a total of twelve groupings planted for each host plant and each experimental
design. Plantings occurred in mid-May and late-May to stagger infestations and
dissections into more manageable numbers. Black plastic (Rain-Flo Irrigation, East Earl,
PA, USA) was laid between groupings to prevent weeds. Weeds were manually removed
within row middles and with Roundup® (Monsanto Company®, St. Louis, MO, USA)
outside of research plots.
Neonate Infestation
In neonate experiments, the center plant of a grouping was infested with a 1.5 mL
microcentrifuge tube containing two blackhead stage egg masses. Tubes were attached to
23
the plants with a paper twist tie. The surrounding eight plants were left uninfested to
receive only individuals moving off of the infested plant. Additional sweet corn
groupings were planted and infested with neonates to serve as developmental monitors.
Using a split plot design, neonate infested plants were either dissected at mid-
development (4th instar; 355 degree-days) or at estimated pupation (790 degree-days).
Both mid-development and pupal dissections consisted of 12 groupings split evenly over
the two plant dates for each host plant type. All nine plants in a grouping were inspected;
the number of individuals collected per grouping and their weights were recorded. In
addition, injury on plants without individuals recovered was documented. If larvae were
recovered at the later dissection, they were kept on ECB diet in the growth chamber until
pupation.
Third Instar Infestation
For 3rd instar experiments, the center plant of a grouping was infested with five 3rd
instars placed on separate leaves to minimize density dependent effects. The surrounding
eight plants were left uninfested to receive only individuals moving off the infested plant.
There were two replicates of six plant clusters per host. Additional sweet corn groupings
were planted and infested with 3rd instars as developmental monitors. All nine plants in a
grouping were inspected at estimated pupation (567 degree-days after infestation). The
number of individuals collected per grouping and their weights were recorded. In
addition, injury on plants without individuals recovered was documented. If larvae were
recovered, they were kept with ECB diet in the growth chamber until pupation.
24
Statistical Analysis
The mid-development and pupal weights in all experiments were analyzed using
two-way ANOVAs followed by Tukey-Kramer’s mean separation test in the program R
2.14 (R Development Core Team) to determine an interaction between the two
independent variables on the dependent variable. The nominal and independent variables
were date of experiment and host plant. The measurement/dependent variables were mid-
development or pupal weight. Growth chamber survival data was analyzed using the
Kaplan-Meier Method for survival analysis to compare the longevity of insects reared on
different hosts until pupation followed by the Wilcoxon test to adjust for multiple
comparisons and Tukey-Kramer’s mean separation test in the program SAS 9.3 (SAS
Institute Inc., Cary, NC). Overall length of survival was measured in degree-days
accumulated until death. Individuals that pupated were censored data points and removed
from the analysis, since very few survived to pupation. Percent survival after the one
stadium of treatment (neonate to 2nd instar; 3rd to 4th instar) based on degree-day
accumulation (2nd = 106 degree-days; 4th = 132 degree-days) was analyzed using a one-
way ANOVA in the program R based on survival percentages of the 3 replications. The
number of individuals recovered per plant (greenhouse) or plant grouping (field) was
analyzed with two-way ANOVAs followed by Tukey-Kramer’s mean separation test in
the program R. The nominal variables were date of experiment and host plant. The
measurement variable was the number of individuals recovered. For all analyses, P-
values of 0.05 or less were considered statistically significant.
25
Results
Growth Chamber
Within the growth chamber statistical analyses, it should be noted that there were
significant interactions regarding experiment replication (Table 1). There were occasions
when one date was significantly different from the other two when measuring weight or
survival The survival analyses conducted did not take this effect into consideration.
Neonate Infestation
Only neonates fed near isoline or sweet corn survived to the mid-development
(355 degree-days) and pupal weight observed points. Individuals were significantly
heavier at mid-development when fed near isoline than those fed sweet corn (Table 1).
There were no significant differences in pupal weight when larvae were fed near isoline
or sweet corn (Table 1).
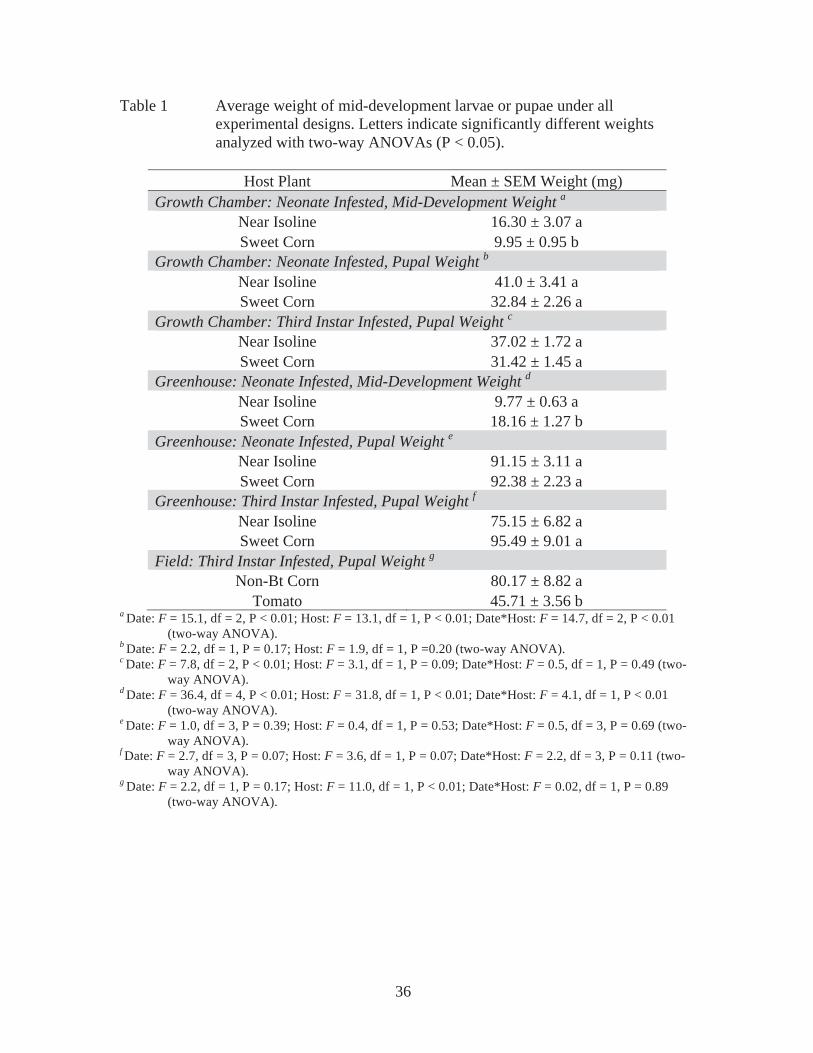
Survival curves show that larvae fed sweet corn or near isoline survived
approximately the same length of time, and significantly longer than those fed the other
four host plants (Figure 1). In addition, these were the only hosts that allowed individuals
to reach pupation. Larvae fed Cry1F, cucumber, or tomato survived the same length of
time, but shorter than the other host plants. Larvae fed green bean survived for an
intermediate period of time significantly different from larvae fed the other host plants
(Figure 1). Percentage of survival through the first stadium of treatment was consistent
with the overall survival (Figure 2A). Here, survival to 2nd instar is presented based on
degree-day accumulation. The largest percentage of individuals survived 106 degree-days
when fed sweet corn, near isoline, or green bean (Figure 2A). Individuals fed cucumber,
tomato, and Cry1F had the lowest survival (Figure 2A).
26
Third Instar Infestation
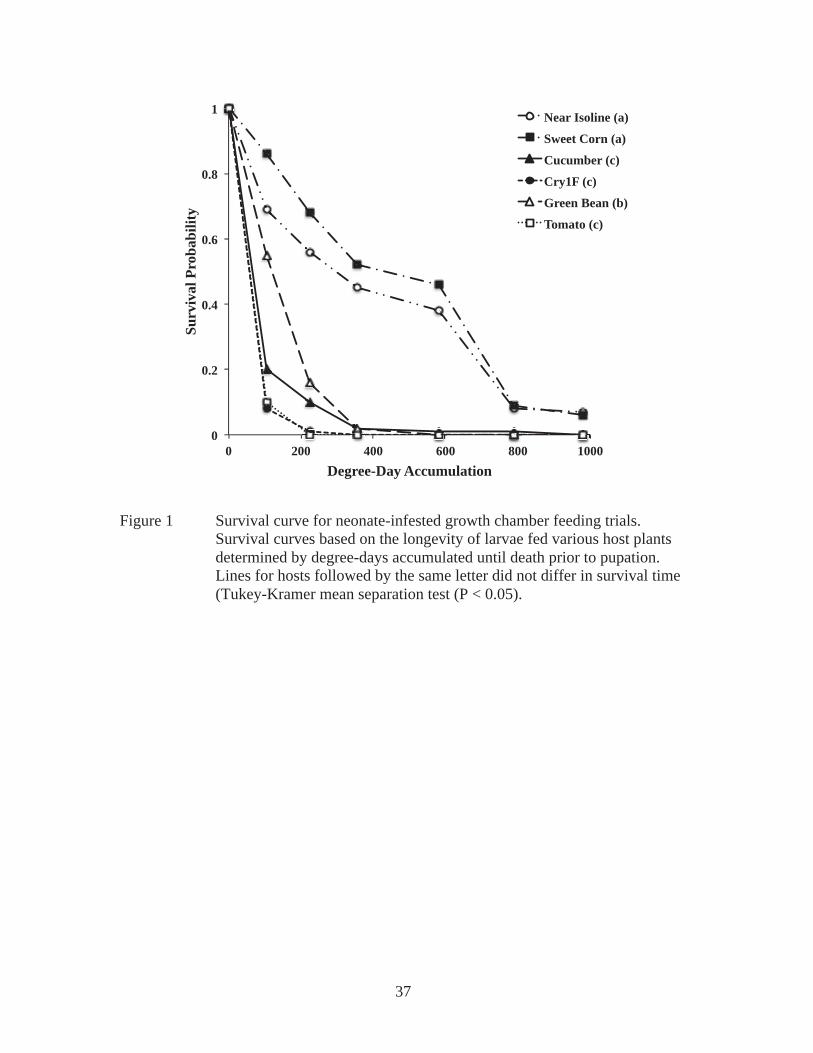
Third instars only survived to pupation on near isoline and sweet corn. Pupae
were not significantly heavier when fed near isoline than those fed sweet corn (Table 1).
Survival curves show third instars fed sweet corn or near isoline survived
statistically the same amount of time, and significantly longer than larvae fed any other
host plant tested (Figure 3). Larvae fed cucumber survived an intermediate length of
time, while larvae fed Cry1F, green bean, or tomato survived the shortest (Figure 3).
Percentage of survival through the first stadium of treatment was consistent with the
overall survival (Figure 2B). The largest percentages of individuals to survive 132
degree-days were fed sweet corn or near isoline, cucumber had intermediate survival, and
individuals fed Cry1F, green bean, or tomato had the lowest survival.
Greenhouse
Neonate Infestation
In the neonate experiments, only enough individuals were recovered from near
isoline or sweet corn to collect larval mid-development (355 degree-days) and pupal,
weights. Mid-development individuals were significantly heavier when fed sweet corn
than those fed near isoline (Table 1). There was no significant difference in pupal weights
when fed sweet corn or near isoline (Table 1).
At mid-development dissection, corn plants were in late vegetative stages and all
other plants were beginning to flower. There were significantly more individuals found
on sweet corn and near isoline than any other host tested (Figure 4A). There was an
average of 18 individuals recovered from sweet corn or near isoline and fewer than 1
27
individual per squash, cucumber, tomato, green bean, or Cry1F plant. At pupal dissection,
corn plants were in early reproductive stages and all other plants were flowering. There
were significantly more individuals recovered from sweet corn than from near isoline, but
still more individuals from near isoline than cucumber, Cry1F, green bean, tomato, or
squash (Figure 4B). No individuals were recovered from Cry1F or tomato. For both
dissections, there was rarely injury observed on tomato or Cry1F, minimal injury on
green bean, cucumber, and squash, and heavy injury on sweet corn and near isoline.
Third Instar Infestation
In the 3rd instar experiments, only enough individuals were recovered from near
isoline or sweet corn to collect pupal weights. At pupal dissection, corn plants were in
late vegetative stages and all other plants were beginning to flower. There was no
significant difference in pupal weights when fed sweet corn or near isoline (Table 1). The
highest pupal recovery was from non-Bt corn hosts, which were significantly higher than
all other hosts (Figure 4C). Of five individuals placed on each plant, on average two to
three were recovered from near isoline and sweet corn, and none from cucumber, Cry1F,
green bean, and tomato. There was heavy damage on near isoline and sweet corn; there
was minimal damage observed on tomato, green bean, cucumber, squash, and Cry1F.
Field
All of the plants grown in the field were noticeably larger than those grown in the
greenhouse. There were no pots to restrict growth and the plants were open to natural
sunlight. At plant dissection plants were much more developed than in greenhouse
experiments. Corn was typically in late vegetative stages for mid-development dissection
28
and early reproductive stages for pupal dissection. All of the other plants were fruiting at
both dissections. Additionally, plants were open to other herbivorous pests, while ECB
larvae were exposed to predators (especially ladybird beetles) and parasitoids.
Very few individuals were recovered from field experiments on any host plant.
There were no differences detected in recovery for any of the field experiments, neonate
or third instar infested plants (Figure 5A: P > 0.23; Figure 5B: P > 0.25; Figure 5C: P >
0.07). Of the two egg masses (~ 40 neonates) released in the 9-plant clusters, on average
0.25 or fewer were recovered per plant at mid-development (Figure 5A), with even fewer
at pupation (Figure 5B). Of the five third instars placed in the clusters of nine plants,
fewer than one per plant were recovered (Figure 5C).
However, ECB injury was observed on all plants. Minimal injury was found on
Cry1F, squash, and cucumber. Moderate injury was observed on sweet corn, near isoline,
green bean, and tomato. More injury was observed on green bean and tomato than
expected based on growth chamber and greenhouse results. Many empty galleries were
observed in sweet corn, near isoline, and green bean.
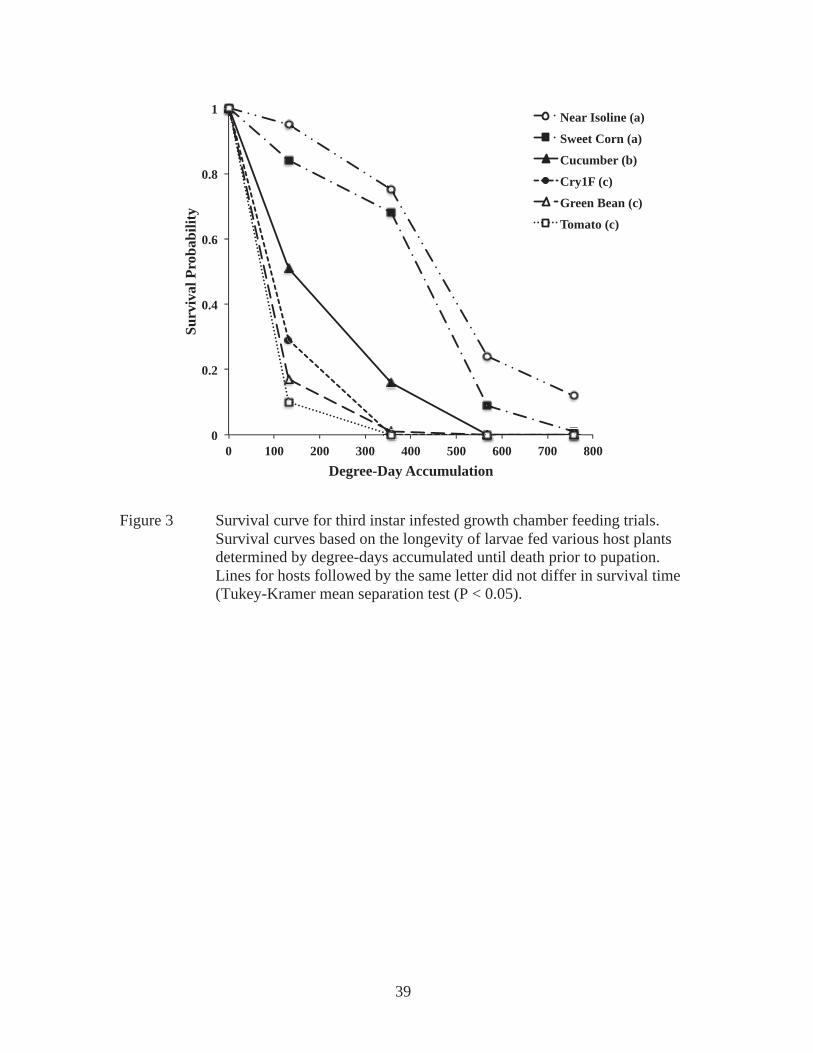
There were only enough individuals recovered from the plants that were infested
at third instar and dissected at pupation to analyze weight. In this case, to increase the
sample size, individuals from sweet corn and near isoline were pooled into one category
(“non-Bt corn”) since no significant differences were found in the growth chamber or
greenhouse trials. The average pupal weight recovered from non-Bt corn was
significantly higher than pupae recovered from tomato (Figure 6).
29
Discussion
Growth chamber trials yielded minimal survival to pupation. When the larvae
were fed plant tissue in the growth chamber, it was only leaf tissue without stalks or
stems. Since ECB are stem-boring insects, the larvae may have been stressed by being
limited to leaf tissue, resulting in a high amount of death. However, there were still
meaningful trends present in the growth chamber survival data (Figure 1 and 2). In both
experiments (neonate or 3rd instar infestation), larvae survived significantly longer on
non-Bt corn hosts (near isoline and sweet corn) than any of the other hosts, suggesting
that non-Bt corn is the most acceptable host among those tested. In turn, larvae survived
the shortest period (approximately 2-6 days) on Cry1F and tomato, suggesting that these
are very poor hosts for larval growth and development. Green bean was an intermediate
host for neonates and a poor host for 3rd instars. Cucumber was a poor host for neonates
and an intermediate host for 3rd instar larvae. Because of this, it is difficult to classify
either cucumber or green bean as a favorable or unfavorable host species. Consequently,
it is appropriate to classify them as intermediate in host suitability.
In the greenhouse, the greatest survival was on non-Bt corn hosts, with little to no
survival on any other plant species in all experiments (neonate and 3rd instar). Sweet corn
and near isoline were expected to provide optimal survival because of relatively reduced
host defenses (Wink 1988; Rosenthal and Dirzo 1997). When neonates remained and
survived on the plants until pupation sweet corn was able to sustain more individuals,
supporting the idea that sweet corn is a more suitable host than near isoline. Few
individuals were recovered and minimal plant injury was observed from ECB on squash,
green bean, and cucumber, but the differences in growth, development, and survival were
30
not enough to be statistically significant. The small ECB recovery numbers could be
because these are not sufficient hosts for complete ECB development, but also because
these plants offer less plant tissue and stem diameter than the corn hosts, providing
insufficient habitat and food resources for ECB development (Ehrlich and Raven 1964;
Losey et al. 2002). The average dry weight of the corn plants provided at infestation was
22.3 mg and 4.5 mg for non-corn hosts. It has been found that when the stem diameters of
non-corn hosts are too small, ECB will abandon the plant and search for a new host
(Losey et al. 2002). There was minimal injury and no individuals were recovered from
Cry1F. Rarely was any injury observed on tomato plants in the greenhouse; only one
individual survived to pupation from a 3rd instar infestation. These results suggest that
Cry1F and tomato are unsuitable host plants for ECB used in this experiment.
In the field, there were no significant trends observed in any experiment.
Recovery was so low, with most plants recovering zero individuals, that it is difficult to
draw any conclusions. Most unfailingly, one individual on average was recovered from
sweet corn groupings, but this did not occur frequently enough to cause a significant
difference. Strangely, there were a relatively large number of individuals recovered as
larvae and pupae from tomato plants. This does not coincide with the experiments
conducted in the incubator or greenhouse. There is potential that these individuals moved
from other host plants in the field. If a host plant is unsuitable, ECB are capable of
abandoning the host and surviving three weeks in search of a more suitable host (Losey et
al. 2002). There were never individuals recovered from Cry1F, cucumber, or squash in
the field.
31
There are a few external reasons that could have caused the low survival on all
host species in the field. While in the field, I noticed an abundance of generalist
predators, specifically ladybird beetles. It is likely that generalist predators consumed
many of the eggs and larvae since ladybird beetles provide biological control as general
predators (Dixon 2000). In addition, many of the larvae that we recovered and reared in
the laboratory were parasitized, which was another source of mortality in the study. Even
though degree-day accumulation was closely followed and monitor plants were
frequently checked, occasionally empty galleries were observed that appeared to have
housed pupae that had already eclosed as adults. The majority of pupae recovered were
female leading to the belief a portion of the males may have escaped. Female ECB eclose
a few days after males (Mason et al. 1996). If I were to conduct this study again in the
future, I would plan to dissect the host plants a few days earlier and rear the recovered
larvae on diet until pupation occurred instead of hoping to recover pupae.
Weight data was collected from all of the experiments in the growth chamber,
greenhouse, and field. In most experiments, only individuals from non-Bt corn hosts
survived to the point of weight data collection. It was not expected that there would be a
difference in weight or degree- day accumulation between individuals fed sweet corn or
near isoline since they are both non-Bt corn hosts providing relatively the same nutrition
and plant defenses. In some parts of the study, there was a significant difference between
the two, but which larvae were was heavier varied between the two host plants and was
not consistent. Sometimes sweet corn had insects heavier and other times near isoline
produced insects with greater weights. This variation was due to some other factor, not
the host itself.
32
Pupae recovered from tomato plants in the field were significantly lighter than
those recovered from non-Bt corn hosts (Figure 6). Larger pupae yield larger adults,
which have been shown to have greater fitness and are more successful (Kingsolver and
Huey 2008). This suggests that when ECB larvae successfully develop on tomato, there is
a reduction in their potential fitness. In the growth chamber and greenhouse experiments,
individuals fed tomato did not survive to pupation. The tomatine present in the green leaf
tissue of tomato plants may be initially acting as an antifeedant when larvae first
encounter it (Isman 2002). If there are no other host options, larvae will either starve to
death or eventually feed on tomato. However, those that do feed end up significantly
smaller, leading to reduced fitness when they become adults.
Components of host plant quality, such as nitrogen and defensive metabolite
concentrations, have been studied in depth. Host plant quality is a strong determining
factor in female fecundity. Individuals that were fed a better quality host plant have
greater reproductive potential and are, therefore, more fit (Awmack and Leather 2002).
When neonates remained on the plants in the greenhouse until pupation, sweet corn was
able to sustain more individuals, possibly because sweet corn contains greater nutritional
value than near isoline. Digestible protein was suspected as one nutritional difference,
however, we were unable to determine a difference when near isoline (n = 6; = 13.25%;
SE = 0.45) and sweet corn (n = 6; = 13.55%; SE = 1.28) tissue samples were sent to
AgriAnalysis in Leola, PA for a basic forage test (P = 0.83).
It is likely that the presence of plant deterrents and toxins caused the large
significant difference in length of survival in the growth chamber and recovery from the
greenhouse between the non-Bt corn hosts and all other plants. First, it is known that
33
Cry1F will kill ECB larvae upon feeding (Gill et al. 1992). Additionally, it has repeatedly
been shown that alkaloids are deterrents or toxins (Wink 1988; Thorsteinson 1960;
Bentley et al. 1984). There are some insect species that can detoxify alkaloids, but these
insects are plant specialists that utilize alkaloids to their defensive benefit, like the
tobacco hornworm (Boppre 1989). However, there is no evidence of ECB utilizing
alkaloids, which helps to explain the lack of survival on tomato plants. It has been shown
that cucurbitacin is an oviposition deterrent specifically for ECB (Tallamy et al. 1997).
This compound may be the cause for reduced larval survival on cucurbits. Reduced
survival on green bean may be explained by plant chemistry, as well. Some legumes
produce a high diversity of secondary metabolites used for defense against herbivores
(Wink 2013).
It is important to remember that Z-race ECB from a laboratory colony were
utilized for all of the experiments in this study. The Z pheromone race tends to feed and
oviposit mostly on corn because their first generation life cycle is more synchronized
with the availability of corn in V5-R1, while E pheromone race ECB have been known to
feed on alternative host plants because they eclose prior to corn availability (Clacagno et
al. 2007). If the same experiments were conducted with E-race ECB instead of Z-race
ECB, the results may show differing survival and host utilization.
All of the data presented here from the growth chamber and greenhouse
experiments support the idea that there is a tendency toward host plant specialization on
corn, or ECB was never as general of a pest as once documented. The criteria for host
specialization include: (1) the hosts provided should always be accepted or rejected, (2)
the hosts in an expanded diet would be of low suitability, and (3) the most suitable host is
34
easy to find (Jaenike 1990; Charnov and Stephens 1988; Futuyma 1983; Levins and
MacArthur 1969; Rausher 1983). Supporting the first and second criteria, less suitable
hosts are only acceptable occasionally when a more suitable host is not available, and
never consistently. In the growth chamber and greenhouse the vast majority of survival
was on non-Bt corn hosts with little to no survival to pupation on any other host. Further
supporting this, Cry1F and tomato were consistently rejected in the growth chamber and
greenhouse. Not only were there no survivors, but also there was no feeding damage. The
third criterion above is easily met by the agricultural corn system. Bt Corn is one of the
most abundant crops grown in the United States. Every Bt field requires a refuge of non-
Bt corn, the most suitable host for ECB, making it easy for ECB to find. Corn also is
much more abundant than any other potential host plant in the Corn Belt (the abundant
soybeans are not a preferred ECB host), where historically the worst ECB infestations
have occurred (Brindley and Dicke 1963). In addition, since corn is grown in
monoculture, both the resource concentration and reduced enemies may play a role in
host specialization. The resource concentration hypothesis refers to the abundance of
food sources an insect will have for itself and its future offspring. All of the plants in the
area are healthy host plants (Grez and Gonzalez 1995). The enemy free hypothesis states
that due to the lowered plant diversity the insect diversity is decreased as well. There are
fewer natural enemies present in agriculture fields to feed on or parasitize pest species
(Letourneau 1987). Since ECB is a stem borer, they have the capability of hiding from
potential enemies in corn’s strong stalks. Finally, as stated previously, corn provides
minimal defenses because plants have been selected for yield, size, and palatability.
35
Plants’ energy is directed more toward growth rather than defense (Rosenthal and Dirzo
1997).
In summation, sweet corn and near isoline are the only consistent favorable host
plants for Z-race ECB, supporting the idea of host plant specialization. As expected,
Cry1F was consistently a poor host, yielding no survival or feeding damage in any
experiment. Green bean, cucumber, and tomato are unsuitable hosts that occasionally
yield feeding damage and survival. They are not consistently adequate hosts, but there is
potential that ECB can survive on these plant species. However, when they survive on
tomato, they are less fit in comparison to larvae fed non-Bt corn. It appears very unlikely
that there will be an ECB infestation in any of these crops because survival was very rare.
Based on the history of many non-corn hosts in the early literature (Hodgson 1928; Hüber
et al. 1928), it appears that Z-race ECB has evolved a host plant specialization for corn.
Continual monitoring for Bt resistance, participation in resistance management strategies,
and high abundances of general predators and parasitoids (as was seen in the field
experiments) will continue to reduce ECB populations. Most ECB die because of Bt corn,
and those that do survive, have a high likelihood of predation or parasitism (field
observations). If it continues in this trend, Z-race ECB eventually may become rare in the
United States, or it may evolve back toward selecting other crops or weeds as hosts.
36
Table 1 Average weight of mid-development larvae or pupae under all experimental designs. Letters indicate significantly different weights analyzed with two-way ANOVAs (P < 0.05).
Host Plant Mean ± SEM Weight (mg) Growth Chamber: Neonate Infested, Mid-Development Weight a
Near Isoline 16.30 ± 3.07 a Sweet Corn 9.95 ± 0.95 b
Growth Chamber: Neonate Infested, Pupal Weight b
Near Isoline 41.0 ± 3.41 a Sweet Corn 32.84 ± 2.26 a
Growth Chamber: Third Instar Infested, Pupal Weight c
Near Isoline 37.02 ± 1.72 a Sweet Corn 31.42 ± 1.45 a
Greenhouse: Neonate Infested, Mid-Development Weight d
Near Isoline 9.77 ± 0.63 a Sweet Corn 18.16 ± 1.27 b
Greenhouse: Neonate Infested, Pupal Weight e
Near Isoline 91.15 ± 3.11 a Sweet Corn 92.38 ± 2.23 a
Greenhouse: Third Instar Infested, Pupal Weight f
Near Isoline 75.15 ± 6.82 a Sweet Corn 95.49 ± 9.01 a
Field: Third Instar Infested, Pupal Weight g
Non-Bt Corn 80.17 ± 8.82 a Tomato 45.71 ± 3.56 b
a Date: F = 15.1, df = 2, P < 0.01; Host: F = 13.1, df = 1, P < 0.01; Date*Host: F = 14.7, df = 2, P < 0.01 (two-way ANOVA). b Date: F = 2.2, df = 1, P = 0.17; Host: F = 1.9, df = 1, P =0.20 (two-way ANOVA). c Date: F = 7.8, df = 2, P < 0.01; Host: F = 3.1, df = 1, P = 0.09; Date*Host: F = 0.5, df = 1, P = 0.49 (two- way ANOVA). d Date: F = 36.4, df = 4, P < 0.01; Host: F = 31.8, df = 1, P < 0.01; Date*Host: F = 4.1, df = 1, P < 0.01 (two-way ANOVA). e Date: F = 1.0, df = 3, P = 0.39; Host: F = 0.4, df = 1, P = 0.53; Date*Host: F = 0.5, df = 3, P = 0.69 (two- way ANOVA). f Date: F = 2.7, df = 3, P = 0.07; Host: F = 3.6, df = 1, P = 0.07; Date*Host: F = 2.2, df = 3, P = 0.11 (two- way ANOVA). g Date: F = 2.2, df = 1, P = 0.17; Host: F = 11.0, df = 1, P < 0.01; Date*Host: F = 0.02, df = 1, P = 0.89 (two-way ANOVA).
37
Figure 1 Survival curve for neonate-infested growth chamber feeding trials. Survival curves based on the longevity of larvae fed various host plants determined by degree-days accumulated until death prior to pupation. Lines for hosts followed by the same letter did not differ in survival time (Tukey-Kramer mean separation test (P < 0.05).
0
0.2
0.4
0.6
0.8
1
0 200 400 600 800 1000
Surv
ival
Pro
babi
lity
Degree-Day Accumulation
Near Isoline (a)
Sweet Corn (a)
Cucumber (c)
Cry1F (c)
Green Bean (b)
Tomato (c)
38
Figure 2 Growth chamber survival through the first stadium of treatment. (A)
Neonates or (B) third instars fed one of various host plants (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, T = tomato) through the duration of development. Bars represent the average percent of the individuals that survived to the subsequent instar based on degree-day accumulation (106 degree-days = 2nd instar; 132 degree-days = 4th instar). Bars within each graph with different letters are significantly different (Tukey-Kramer test, P < 0.05). Error bars represent SEM.
0
20
40
60
80
100
N SC C F GB T
Aver
age
% S
urvi
val
0
20
40
60
80
100
N SC C F GB T
Aver
age
% S
urvi
val
Host Plant
A.
B.
ab a
bc c
abc
bc
a ab
bc
c
c c
39
Figure 3 Survival curve for third instar infested growth chamber feeding trials. Survival curves based on the longevity of larvae fed various host plants determined by degree-days accumulated until death prior to pupation. Lines for hosts followed by the same letter did not differ in survival time (Tukey-Kramer mean separation test (P < 0.05).
0
0.2
0.4
0.6
0.8
1
0 100 200 300 400 500 600 700 800
Surv
ival
Pro
babi
lity
Degree-Day Accumulation
Near Isoline (a)
Sweet Corn (a)
Cucumber (b)
Cry1F (c)
Green Bean (c)
Tomato (c)
40
A: Date: F = 0.3, df = 4, P = 0.85; Host: F = 41.2, df = 6, P < 0.01; Date*Host: F = 1.3, df = 24, P = 0.36
(two-way ANOVA).B: Date: F = 2.4, df = 3, P = 0.09; Host: F = 107.9, df = 6, P < 0.01; Date*Host: F = 5.1, df = 18, P < 0.01
(two-way ANOVA). C: Date: F = 1.9, df = 3, P = 0.15; Host: F = 26.0, df = 6, P < 0.01; Date*Host: F = 1.3, df = 18, P = 0.26
(two-way ANOVA).
Figure 4 Greenhouse recovery. (A) Neonate infested plants (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) were dissected after 355 degree-days for mid-development recovery or (B) after 790 degree-days for pupal recovery. (C) Plants infested with 3rd instars were dissected after 567 degree-days for pupal recovery. Bars with represent the number of individuals recovered at each dissection with SEM. Bars with different letters within one graph are significantly different (P < 0.05).
.
0
5
10
15
20
25
N SC C F GB SQ T
Aver
age
# R
ecov
ered
0
5
10
15
20
25
N SC C F GB SQ T
Aver
age
# R
ecov
ered
0
1
2
3
4
5
N SC C F GB SQ T
Aver
age
# R
ecov
ered
Host Plant
A.
C.
B.
a a
b b b b b
a
b
c c c c c
a a
b b b
b b
41
A: Date: F = 0.7, df = 1, P = 0.40; Host: F = 1.4, df = 6, P =0.22; Date*Host: F = 0.7, df = 6, P = 0.64 (two- way ANOVA).
B: Date: F = 3.5, df = 1, P = 0.07; Host: F = 1.7, df = 6, P = 0.14; Date*Host: F = 1.7, df = 6, P = 0.14 (two-way ANOVA).
C: Date: F = 3.5, df = 1, P = 0.06; Host: F = 3.2, df = 6, P < 0.01; Date*Host: F = 1.1, df = 6, P = 0.38 (two-way ANOVA).
Figure 5 Field recovery. (A) Neonate infested plant groupings (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) were dissected after 355 degree-days for a mid-development recovery or (B) after 790 degree-days for pupal recovery. (C) Plant groupings infested with 3rd instars were dissected after 567 degree-days for pupal recovery. Numbers did not differ for any trial (P > 0.05).
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
N SC C F GB SQ T
Aver
age
# R
ecov
ered
0
0.05
0.1
0.15
0.2
0.25
0.3
N SC C F GB SQ T
Aver
age
# R
ecov
ered
0
0.2
0.4
0.6
0.8
1
1.2
N SC C F GB SQ T
Aver
age
# R
ecov
ered
Host Plant
A.
C.
B.
42
Date: F = 2.2, df = 1, P = 0.17; Host: F = 11.0, df = 1, P < 0.01; Date*Host: F = 0.02, df = 1, P = 0.89 (two- way ANOVA).
Figure 6 Pupal weight comparison from third instar infested plants in the field.Average weight of pupae recovered from 3rd instar infested field experiments from non-Bt corn (sweet corn and near isoline) (n = 7) and tomato (n = 8). There was a significant difference between the two bars (P < 0.01). Error bars represent SEM.
0
20
40
60
80
100
Non Bt Corn Tomato
Aver
age
Pupa
l Wei
ght (
mg)
Host Plant
43
Chapter 3
EUROPEAN CORN BORER PLANT CONSUMPTION RATES AND PREFERENCE AMONG A RANGE OF AGRICULTURAL HOST CROP
SPECIES
Introduction
The European corn borer (ECB), Ostrinia nubialis (Hübner), was introduced to
North America in the early 1900s with the importation of broomcorn (Sorghum
technicum L.) from Italy and Hungary. At introduction, ECB had a preference for corn
(Zea mays L.), but was known to feed opportunistically on 223 crop and weedy host plant
species, earning the label of “generalist herbivore” (Hodgson 1928; Hüber et al. 1928).
Some common alternative hosts include bean, beat, cauliflower, celery, eggplant, pepper,
and tomato (Hodgson 1928). The largest impact of ECB has been observed in corn
agriculture where it is documented to cause more than 1 billion US dollars in annual yield
loss and management/prevention costs (Mason et al. 1996). Currently, ECB is controlled
without resistance by corn genetically modified with insecticidal crystalline (Cry)
proteins from the soil bacterial species Bacillus thuringensis (Fernandez-Cornejo et al.
2014).
Adult oviposition is the primary influence of host selection for larval Lepidoptera.
However, neonate ballooning and larval movement can also facilitate host selection (Ross
and Ostlie 1990). Specifically, larval movement is a concern in resistance management
with a mixed planting of Bt and non-Bt refuge in a cornfield. It has been documented
that 75% of ECB neonates abandon Bt natal plants and 42% of neonates abandon non-Bt
natal host plants (Razze et al. 2011). Young ECB larvae move around a great deal prior to
boring into the plant in the 3rd instar, potentially obtaining sub-lethal doses of Bt toxin as
44
older larvae and speeding up the development of Bt resistance. Larval movement also
opens potential for movement to alternate non-corn hosts, most likely field weed species
(Tate et al. 2006).
Previously, survival was observed on various early documented hosts and
oviposition deterrents (Hodgson 1928; Hüber et al. 1928; Tallamy et al. 1997), and it was
found that ECB most consistently succeeded on non-Bt corn hosts (non-Bt near isoline
corn and sweet corn), never succeeded on Cry1F Bt corn, and succeeded with variability
on cucurbits, green bean, and tomato (Chapter 2). Based on assessment of growth and
development on these hosts it was concluded that non-Bt corn hosts were the most
favorable host plants and all others tested are unfavorable or no longer acceptable hosts
since survival to maturity did not occur under experimental conditions.
No choice and choice tests are common methods for determining larval feeding
preferences and help predict pest occurrence in the presence of host plant options (Barnes
and Ratcliff 1967; Jackai 1991; Smith et al. 1994; Davis and Coleman 1997; Tate et al.
2006). In this part of the study, no choice and choice tests were conducted over the same
range of host plants previously tested for survival to determine if ECB feed more and
prefer host plants that yield better survival. Preference tests help to determine if ECB will
preferentially switch hosts if other options become available. This research addressed the
following objectives through laboratory studies: (1) evaluate amount of feeding on each
host plant based on leaf tissue consumption in a no choice 48-hour period by 3rd instar
ECB, and (2) determine feeding preferences based on leaf tissue consumption by 3rd
instar ECB when presented with a choice.
45
Materials and Methods
Insects and Plants
Larval, Z-race European corn borers (ECB) were received from the USDA-ARS
Corn Insects and Crop Genetics Research Unit (Ames, IA). Each year, field captured
ECB are incorporated into the colony to maintain field relevance and reduce inbreeding.
Third instars were shipped to the University of Delaware (Newark, DE) gregariously,
feeding on ECB diet and maintained in a Percival Scientific® (Perry, IA, USA) growth
chamber and maintained at 25°C, 40-60% humidity, and 16:8 L:D. All subsequent
experiments in the growth chamber were maintained at these same conditions.
Tested host plants were chosen based on early host plant literature documenting
presence of ECB eggs and larval feeding a few years after ECB introduction to North
America (Hodgson 1928; Hüber et al. 1928), and from more recent literature on
oviposition deterrents (Tallamy et al. 1997). To determine ECB’s ability to overcome
natural plant chemical defenses, specific crop types were selected to encompass a broad
range of feeding hosts based on known deterrent/antifeedant chemistry or minimal
defenses. The crops tested included corn encoded with Cry1F Bt toxin along with its near
green beans, Mountain Fresh Plus VFFN hybrid tomato, Fancipak pickling cucumber,
and Buttercup squash. Non-corn varieties selected for the experiments were listed in the
2013 Delaware Commercial Vegetable Production Recommendations as suggested
varieties for commercial farmers in the Delaware area.
Unfavorable host plants were identified as plants that yielded minimal survival in
previous growth chamber and greenhouse studies due to Bt toxin or chemical defenses
46
(Chapter 2). These plants included Cry1F Bt corn and tomato. Cry1F Bt corn is known to
kill ECB because of the presence of Bt toxin (Koziel et al. 1996). Tomato contains
tomatine, an alkaloid found in green plant parts, and is known to deter many insects, and
potentially other compounds (Buhr et al. 1958; Thorsteinson 1960; Boppre 1989; Isman
2002).
Intermediate hosts were those that have defensive chemistry, but ECB were
inconsistently able to survive under growth chamber and greenhouse conditions (Chapter
2). These hosts include cucumber, squash, and green bean. Cucumber and squash contain
varying concentrations of cucurbitacin, a bitter deterrent that is also specifically detected
and avoided by ovipositing ECB (Tallamy et al. 1997). Cucurbitacin content of squash is
0.02 mg/g of fresh weight in the leaves, while cucumber contains trace amounts (Metcalf
et al. 1982). Green bean was originally classified as a favorable host plant because it
offers a proper environment for ECB adults to rest during the day and mate at night, and
adults are known to oviposit on green bean (Mason et al. 1996). However, legumes
contain a diversity of secondary metabolites in the leaf tissues (Wink 2013; Abu-Reidah
2013). Cucumber, squash, and green bean provided inconsistent survival results (Chapter
2).
Favorable host plants were classified as those with minimal or lacking defensive
characteristics that previously provided the highest survival probability (Chapter 2).
These included non-Bt near isoline corn and sweet corn. Seedling corn contains high
concentrations of DIMBOA (2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one), a
compound that prevents feeding and is toxic to ECB neonates until corn reaches
vegetative stage 5 (V5) (Guthrie et al. 1986). Vegetative stage 6 (V6) corn was utilized
47
for experiments to avoid effects of DIMBOA. Near isoline corn is genetically identical
to Cry1F, except without the Bt toxin, and is used commonly as refuge corn. Sweet corn
was tested to determine if it is a more preferred host for ECB than near isoline.
Plants used were grown in the greenhouse maintained at 20-30°C, 40-60%
humidity, and 16:8 L:D supplemented during short day lengths. All plants were grown to
approximately 1000 accumulated degree-days to correspond with corn at V6. At this
point in development, plant leaf tissues were excised to feed ECB.
Consumption Rates and Preference
Choice and no choice tests are common methods for determining larval feeding
preferences (Barnes and Ratcliff 1967; Jackai 1991; Smith et al. 1994; Davis and
Coleman 1997; Tate et al. 2006). However, most previous experiments used an estimated
percentage of leaf material consumed to determine preference. The most accurate
measurement is taken by adjusting a known leaf area based on dry weight of the sample
(Barnes and Ratcliff 1967). After feeding assays, remaining leaf area (cm2) and dry
weight of remaining leaf material (mg) were measured to determine the amount of dry
biomass consumed (mg) based on original leaf area presented of each host type. ImageJ
computer imaging software (Rasband 2015) was used to measure leaf area before and
after feeding (Crawford et al. 2010; Cooney et al. 2012; Salazar and Marquis 2012).
First, no choice experiments were conducted to measure the amount of leaf tissue
consumed over a 48-hour period of exposure to one host plant at a time. A total of 25
replicates were conducted per host plant split evenly over five dates. Leaf disks (1.75 cm
in diameter) were cut using a brass cork borer (Humboldt®, Elgin, IL, USA). Four disks
48
of one host were photographed and placed in a 9 cm petri dish (Fisher Scientific®,
Waltham, WA, USA) containing 9 cm filter paper (Whatman®, Buckinghamshire, UK)
dampened with distilled water to retain moisture. One 3rd instar larva was weighed and
placed in the center of the four leaf disks. Each petri dish was wrapped in parafilm
(American Can Company®, Greenwich, CT, USA) to prevent escaping larvae as well as
desiccation of larvae or leaf tissue. Petri dishes were maintained in the growth chamber
for 48 hours. After 48 hours, the larva was removed and weighed. Leaf disks were
photographed, dried, and weighed. Larval weight change was calculated by subtracting
the weight before from the weight after. Using photographs and ImageJ software, leaf
area before and after consumption were calculated. After drying, the leaf material not
consumed of each leaf per host plant type was weighed. These measurements were used
to calculate the amount of dry biomass consumed by the individual over 48 hours.
To determine preference, leaf disks (1.75 cm in diameter) of two host species
were cut using a cork borer. Host comparisons included near isoline compared with sweet
corn, Cry1F, green bean, cucumber, or tomato and Cry1f compared with tomato. The
sample size for each combination was 25 leaf disk comparisons. Four leaf disks total
were photographed and placed in a 9 cm petri dish containing 9 cm filter paper dampened
with distilled water to retain moisture. Two disks of the same host type were placed
diagonally across from each other. One 3rd instar larva was placed in the center of the
four leaf disks. Each petri dish was wrapped in parafilm to prevent escaping larvae, as
well as desiccation of larvae or leaf tissue. Petri dishes were maintained in the growth
chamber for 48 hours. After 48 hours, the larva was removed. Leaf disks were
photographed, dried, and weighed. Using photographs and ImageJ software, leaf areas
49
before and after consumption were calculated. Total area of leaf tissue consumed was
calculated. After drying, the leaf material not consumed of each leaf per host plant was
weighed. These measurements were used to calculate the amount of dry biomass
consumed of each host by the individual over 48 hours.
Statistical Analysis
For the no choice experiments, the dry weight and the leaf area of material not
consumed were converted into dry biomass per cm2. Multiplying the dry biomass per cm2
by the area of the leaf disk before feeding produced the estimated dry biomass of the leaf
disk before feeding. Lastly, by subtracting the dry biomass after feeding from the
estimates dry biomass before feeding, the dry biomass consumed was calculated. This is
expressed in the following equations:
1.
2.
3.
Both larval weight change and dry biomass consumed were analyzed using two-way
ANOVAs followed by the Tukey-Kramer post hoc test in the program R 2.14 (R
Development Core Team). The nominal variables were date of experiment and host plant.
The measurement variables were either dry leaf weight consumed or larval weight
gain/loss.
For choice tests, dry weights of leaf tissue not consumed and leaf areas were used
to estimate the dry biomass of leaf tissue consumed of each host over a 48-hour period
using the equations provided previously. Choice data was analyzed as a proportion of one
50
host against the other host offered. The dry biomasses consumed were analyzed using a
one-sample t-test in Excel 2011.
Results
In no choice experiments, the most dry biomass was consumed of near isoline
followed by sweet corn (Figure 7). Cucumber and squash were consumed in intermediate
amounts, and very little Cry1F, green bean, or tomato tissue was consumed (Figure 7).
On average, larvae gained the most weight on non-Bt corn hosts (near isoline and sweet
corn) in comparison to other hosts tested (Figure 8). Individuals that consumed squash or
cucumber gained less weight while those that fed on green bean neither gained nor lost
weight (Figure 8). Individuals provided with Cry1F or tomato lost weight (Figure 8).
For choice trials, the comparisons that were not significantly different include
near isoline with sweet corn (Figure 9A), near isoline with Cry1F (Figure 9A), and Cry1F
with tomato (Figure 9B). Larvae consumed significantly more near isoline when provided
with a choice between near isoline and tomato or near isoline with green bean (Figure
9A). Larvae ate significantly more cucumber than near isoline (Figure 9A).
Discussion
Leaf tissue consumption and weight gain show similar trends (Figures 7 and 8).
Larvae gained the most weight and fed most when presented with only near isoline or
sweet corn. From the results here, sweet corn and near isoline remain suitable hosts. In
present-day agriculture, non-Bt corn provides an abundant food source that has minimal
51
defensive chemistry (Rosenthal and Dirzo 1997) and a large diameter stalk for larvae to
bore (Ehrlich and Raven 1964).
As for the intermediate hosts, when larvae were presented with green bean, they
consumed almost no plant material, but maintained their weight. Potentially, if given
more than 48 hours, larvae may feed more abundantly on green bean, or they may lose
weight. When larvae were presented with cucumber or squash, they consumed leaf tissue
and gained some weight within 48 hours. In the experiments provided here, cucumber
and squash were suitable hosts for third instars, providing weight gain and a moderate
consumption rate. These data also suggest that it would take a longer amount of time for
larvae on cucumber, squash, or green bean to develop than those consuming non-Bt corn
hosts. Slower developmental rate and obtaining greater resources for development are
criteria of less than ideal host plants (Timmins et al. 1988).
When larvae were presented with only Cry1F or tomato, there was little to no
feeding resulting in weight loss. Cry1F and tomato remain categorized as unfavorable
host plants. Cry1F corn is encoded with Bt toxin that is known to greatly reduce feeding
(Koziel et al. 1996). From observations, it was assumed that tomato would be a poor host
due to larval avoidance observed in preliminary studies. In addition, tomato is a highly
defended plant, with very few insects capable of detoxification (Boppre 1989). Neither
Cry1F nor tomato is a suitable host plant for ECB. By losing weight, larval growth is
regressing which will ultimately lead to death.
Preference was determined as a ratio of one host against the other host option.
There was no preference observed when larvae were given near isoline with sweet corn.
Sweet corn and near isoline were expected to be the same because they provide similar
52
nutrition value and minimal plant defenses (Chapter 2; Rosenthal and Dirzo 1997).
When presented with a choice between Cry1F and near isoline, there was a preference for
near isoline. A preference for near isoline was expected because it has been found that
ECB have a preference for near isoline when given a choice between Cry1AB and its
near isoline (Davis and Coleman 1997). No preference was detected when a choice was
offered between Cry1F and tomato. This was expected because they are both very poor
hosts causing the larvae to feed in very small amounts, lose weight, and have poor
survival (Koziel et al. 1996; Thorsteinson 1960). As anticipated, there was a preference
for near isoline when it was paired with tomato; tomato is a toxic host and near isoline is
a suitable host plant for ECB. There was also a preference for near isoline, as expected,
when it was paired with green bean. Based on the no choice data, green bean was an
intermediate host with minimal feeding, resulting in no change in weight over 48 hours.
An unexpected result from the choice experiments arose when a strong preference for
cucumber over near isoline was observed (Figure 9A).
Based on previous experiments, cucurbits provide poor survival for ECB (Chapter
2). Yet, from the results of the consumption analysis and preference tests, cucumber
appears to provide moderate consumption (Figure 7) and moderate weight gain (Figure
8), as well as a preference over near isoline (Figure 9). Since larvae did not survive to
pupation in the growth chamber, greenhouse, or field, there must be another explanation
for the behavior. One potential difference in these experiments pertains to the leaf tissue
present. In the previous greenhouse and field experiments, whole plants were utilized,
while in the no choice/choice experiments, excised leaf disks were provided. This
suggests that ECB may be inducing plant defenses when feeding on whole plants that are
53
not present with excised leaf tissue due to cutting off the connective elements to the leaf
tissue. It has been shown that the squash beetle girdles cucurbit leaves, cutting off
transfer of compounds in order to reduce interaction with induced defenses (Tallamy
1985). Additionally, when ECB larvae were fed cucumber leaf tissue throughout
development in the growth chamber, there was intermediate survival from 3rd to 4th instar
for individuals fed cucumber (Chapter 2). In the present experiment, leaf tissue was
removed from the plants, potentially preventing induced defenses from a fully
functioning plant resulting in greater survival on plant parts than on full plants. Induced
defenses of cucumber were briefly explored, however, no significant differences were
found (see Appendix A).
In summary, this study shows that near isoline field corn and sweet corn are the
only hosts of those tested that can be classified as favorable host plants based on high
consumption rates, weight gain, and high preference of the hosts evaluated here. Based
on low levels of the same criteria, Cry1F and tomato are classified as unsuitable host
plants. Green bean, cucumber, and squash remain intermediate hosts, however, the role
where cucurbits fit in as ECB host plants requires further investigation. Generalist insects
will accept many host plants and rarely exercise choice (Schoonhoven et al. 2005), which
is not the case for the Z-race ECB tested in this study. Aside from the cucumber data, all
of the experiments presented here further support claims of host plant specialization made
in Chapter 2. The criteria for host specialization include: (1) the hosts provided should
always be accepted or rejected, (2) the hosts in an expanded diet would be of low
suitability, and (3) the most suitable host is easy to find (Jaenike 1990; Charnov and
Stephens 1988; Futuyma 1983; Levins and MacArthur 1969; Rausher 1983). In support
54
of the first criterion, ECB feed more abundantly on the host plants on which they succeed
best (non-Bt corn hosts) and ECB preferentially select non-Bt corn as a larval host plant
(except for excised cucumber leaves). The second criterion is met by previous
survivorship studies presented in Chapter 2. Non-Bt corn hosts were found to be the most
suitable of the plant types tested, and all of the other plant type resulted in decreased
survival. The third criterion above is easily met by the agricultural corn system, despite
the abundance of Bt corn grown. Corn, the most suitable host for ECB, is one of the most
abundant crops grown in the United States making it easy for ECB to find. It also is much
more abundant than any other potential host plant in the Corn Belt (the abundant
soybeans are not a preferred ECB host), where historically the worst ECB infestations
have occurred (Brindley and Dicke 1963). From the data presented here, it appears that
ECB is becoming a corn specialist.
55
Date: F = 1.6, df = 4, P = 0.18; Host: F = 55.2, df = 6, P < 0.01; Date*Host: F = 1.3, df = 24, P = 0.19 (two-
way ANOVA).
Figure 7 Dry biomass consumed. Third instars fed one of the various hosts (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) for 48 hours. Bars represent the estimated amount of dry biomass consumed. Different letters are significantly different (P < 0.05). Error bars show SEM
0 2 4 6 8
10 12 14 16 18
N SC C F GB SQ T
Dry
Bio
mas
s Con
sum
ed (m
g)
Host Plant
a
cd c
b
d d d
56
Date: F = 7.8, df = 4, P < 0.01; Host: F = 39.5, df = 6, P < 0.01; Date*Host: F = 1.5, df = 24, P = 0.07 (two- way ANOVA).
Figure 8 Third instar larval weight change over 48 hours. Third instars fed one of the various hosts (N = near isoline, SC = sweet corn, C = cucumber, F = Cry1F, GB = green bean, SQ = squash, T = tomato) for 48 hours. Bars represent either gained (+) or lost (-) weight based on host. Different letters are significantly different (P < 0.05). Error bars show SEM.
-4
-2
0
2
4
6
8
N SC C F GB SQ T
Aver
age
Wei
ght C
hang
e (m
g)
Host Plant
a
a
b bc
cd d d
57
Figure 9 Preference determined by dry biomass consumed. Third instars given a choice between 2 host plants (near isoline (N) and either sweet corn (SC), Cry1F (F), tomato (T), green bean (GB), or cucumber (C), which are presented in figure 3A, or Cry1F (F) and tomato (T), presented in figure 3B) for 48 hours. Bars represent the estimated dry biomass consumed. Significant differences within one group are distinguished with an asterisk (*). Error bars show SEM.
B.
0
1
2
3
4
5
6
7
8
9
F T
Dry
Bio
mas
s Con
sum
ed (m
g)
Host Plant
0
1
2
3
4
5
6
7
8
9
SC F T GB C
Dry
Bio
mas
s Con
sum
ed (m
g)
Host Plant Compared to Near Isoline
N
Other
* *
* * A.
58
REFERENCES
Abu-Reidah, I. M., D. Arraez-Roman, J. Lazano-Sanchez, A. Segura-Carretero, and A. Fernandez-Gutierrez. 2013. Phytochemical characterization of green beans (Phaseolus vulgaris L.) by using high-performance liquid chromatography coupled with time-of-flight mass spectrometry. PCA. 24: 105-116.
Ali, J. B. and A. A. Agrawal. 2012. Specialist versus generalist insect herbivores and
plant defense. Trends Plant Sci. 17: 293-302. Awmack, C. S. and S. R. Leather. Host plant quality and fecundity in herbivorous insects.
Annu. Rev. Entomol. 47: 817-844. Baldwin, I. T., R. Halitschke, A. Paschold, C. C. von Dahl, and C. A. Preston. 2006.
Volatile signaling in plant-plant interactions: “talking trees” in the genomics era. Science 311: 812-815.
Barnes, D. K. and R. H. Ratcliffe. 1967. Leaf disk method of testing alfalfa plants for
resistance to feeding by adult alfalfa weevils. J. Econ. Entomol. 60: 1561-1565. Bartlett, B. R., C. P Clausen, P. DeBach, R. D. Goeden, E. F. Legner,J. A. McMurtry,
and E. R. Oatman. 1987. Introduced parasites and predators of arthropod pests and weeds: a world review. Washington, D.C.: United States Department of Agriculture, January 1978.
Beck, S. D. 1987. Developmental and seasonal biology of Ostrinia nubilalis. Agric. Zool.
Rev. 2:59-96. Benrey, B. and R. F. Denno. 1997. The slow-growth-high-mortality hypothesis: a test
using the cabbage butterfly. Ecology. 78: 987-999. Bentley, M. D., D. E. Leonard, and R. J. Bushway. 1984. Solanum alkaloids as larval
feeding deterrents for spruce budworm, Choristoneura fumiferana (Lepidoptera: Tortricidae). Ann. Entomol. Soc. Am. 77: 401-403.
Bernays, E. A. and M. Graham. 1988. On the evolution of host specificity in
phytophagous arthropods. Ecology. 69: 886-892. Boppre, M. 1990. Lepidoptera and pyrrolizidine alkaloids: exemplification of complexity
in chemical ecology. J. Chem. Ecol. 16: 165-185. Brindley, T. A. and F. F. Dicke. 1963. Significant developments in European corn borer
research. Annu. Rev. Entomol. 8: 155-176. Buchanan, B.B., W. Gruissen, and R. L. Jones. 2000. Biochemistry and molecular
biology of plants. American Society of Plant Physiologists.
59
Buhr, H., R. Toball, and K. Schreiber. 1958. Entomol. Exp. Appl. 1:209. Calcagno, V., Y. Thomas, and D. Bourguet. 2007. Sympatric host races of the European
corn borer: adaptation to host plants and hybrid performance. J. Evolution. Biol. 20: 1720-1729.
Charnov, E. L. and D. W. Stephens.1988. On the evolution of host selection in solitary
parasitoids. Am. Nat. 132:707- 22 Chen, Y., J. R. Ruberson, and D. M. Olson. 2008. Nitrogen fertilization rate affects
feeding, larval performance, and oviposition preference of the beet armyworm, Spodoptera exigua, on cotton. Entomol. Exp. Appl. 126:244–255.
Chen, Y., X. Ni, and G. D. Buntin. 2009. Physiological, nutritional, and biochemical
bases of corn resistance to foliage feeding fall armyworm. J. Chem. Ecol. 35:297-306.
Cohen, J. E. 2003. Human population: the next half century. Science. 302:1172-1175. Cooney, L. J., J. W. van Klink, N. M. Hughes, N. B. Perry, H. M. Schaefer, I. J. Menzies,
and K. S. Gould. 2012. Red leaf margins indicate increased polygodial content and function as visual signals to reduce herbivory in Pseurowintera colorata. New Phytol. 194: 488-497.
Crawford, K. M., J. M. Land, and J. A. Rudgers. 2010. Fungal endophytes of native
Davis, P. M. and S. B. Coleman. 1997. European corn borer feeding behavior and
survival on transgenic corn containing Cry1Ab protein from Bacillus thuringiensis. J. Kans. Entomol. Soc. 70: 31-38.
Dixon, A. F. G. 2000. Insect predator-prey dynamics: ladybird beetles and biological
control. New York: Cambridge University Press. Ehrlich, P. R. and D. D. Murphy. 1988. Plant chemistry and host range in insect
herbivores. Ecology. 69: 908-909. Ehrlich, P. R. and P. H. Raven. 1964. Butterflies and plants: a study of coevolution.
Evolution. 18: 586-608. Elsayed, G. 2011. Plant secondary substances and insects behaviour. Arch. Phytopath.
Plant Prot. 44: 1534-549.
60
Fernandez-Cornejo, J., S. Wechsler, M. Livingston, and L. Mitchell. 2014. Genetically engineered crops in the United States. United States Department of Agriculture. Economic Research Report Number 162.
Forister, M. L, V. Novotny, A. Panorska, L. Baje, Y. Basset, P. T. Butterill, L. Cizek, P.
D. Coley, F. Dem, I. R. Diniz, P. Dorozd, M. Fox, A. E. Glassmire, R. Hazen, J. Hrcek, J. P. Jahner, O. Kaman, T. J. Kozubowski, T. A. Kursar, O. T. Lewis, J. Lill, R. J. Marquis, S. E. Miller, H. C. Morais, M. Murakami, H. Nickel, N. A. Pardikes,R. E. Ricklefs, M. S. Singer, A. M. Smilanich, J. O. Stireman, S. Vilamarin-Cortez, S. Vodka, M. Volf, D. L. Wagner, T. Walla, G. D. Weiblen, and L. A. Dyer. 2015. The global distribution of diet breadth in insect herbivores. PNAS. 112: 442-447.
Fox, L. R., D. K. Letourneau, J. Eisenbach, and S. V. Nouhuys. 1990. Parasitism rates
and sex ratios of a parasitoid wasp: effects of herbivore and plant quality. Oecologia. 83:414–419.
Futuyma, D. J. 1983. Selective factors in the evolution of host choice by phytophagous
insects. In Herbivorous Insects, ed. S. Ahmad, pp. 227-44. New York: Academic Futuyma D. J. and F. Gould. 1979. Associations of plants and insects in a deciduous
forest. Ecol. Monogr. 49:33-50 Gill, S. S., E. A. Cowles, and P. V. Pietrantonio. 1992. The mode of action of Bacillus
thuringiensis endotoxins. Annu. Rev. Entomol. 37: 615-36. Glendinning, J. I. and F. Slansky Jr. Interactions of allelochemicals with dietary
constituents: effects on deterrency. Physiol. Entomol. 19: 173-186. Goldstein, J. A., C. E. Mason, and J. Pesek. 2010. Dispersal and movement behavior of
neonate European corn borer (Lepidoptera: Cambidae) on non-Bt and transgenic Bt corn. J. Econ. Entomol. 103: 331-339.
Gols, R., T. Bukovinszky, N. M. van Dam, M. Dicke, J. M. Bullock, and J. A. Harvey.
2008. Performance of generalist and specialist herbivores and their endoparasitoids differs on cultivated and wild Brassica populations. J. Chem. Ecol. 34: 132-143.
Gore, J., J. J. Adamczyk, and C. A. Blanco. 2005. Selective feeding of tobacco budworm
and bollworm on meridic diet with different concentrations of Bacillus thuringiensis proteins. J. Econ. Entomol. 98: 88-94.
Gould, F. 2000. Testing Bt refuge strategies in the field. Nature Biotechnol. 18: 266-267.
61
Guthrie, W. D., C. T. Tseng, W. A. Russell, J. R. Coats, J. C. Robbins, and J. J. Tollefson. 1986. DIMBOA content at seven stages of plant development in a maize synthetic cultivar. Journal of the Kansas Entomology Society 59: 356-60.
Grez, A. A. and R. H. Gonzalez. 1995. Resource concentration hypothesis: effect of host
plant patch size on density of herbivorous insects. Oecolgia. 103: 417-474. Hodgson, B. E. 1928. The host plants of the European corn borer in New England. U.S.
Dep. Agric. Tech. Bull. 77. 64 pp. Hüber, L. L., C. R. Neiswander, and R. M. Salter. 1928. The European corn borer and its
environment. Ohio Agriculture Experiment Station. 429: 13-151. Hutchison, W. D., E. C. Burkness, P. D. Mitchell, R. D. Moon, T. W. Leslie, J. Fleischer,
M. Abrahamson, K. L. Hamilton, K. L. Steffey, M. E. Gray, R. L. Hellmich, L. V. Kaster, T. E. Hunt, R. J. Wright, K. Pecinovsky, T. L. Rabaey, B. R. Flood, E. S. Raun. 2010. Area wide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science. 330: 222-225.
Isman, M. 2002. Insect antifeedants. Pesticide Outlook. August 2002: 152-157. Jackai, L.E.N. 1991. Laboratory and screenhouse assays for evaluating cowpea resistance
to the legume pod borer. Crop Prot. 10: 48-52. Jaenike, J. 1990. Host specialization in phytophagous Insects. Annu. Rev. of Ecol. Syst.
21: 243-273. Jermy, T. 1976. Insect-host-plant relationship - Coevolution or sequential evolution?
Symp. Biol. Hung. 16:109-113. Kingsolver, J. G. and R. B. Huey. 2008. Size, temperature, and fitness: three rules. Evol.
Ecol. Res. 10: 251-268. Kogan, M. and R. D. Goeden. 1970. The host-plant range of Lema trilineata daturaphila
(Coleoptera: Chrysomelidae). Ann. Entomol. Soc. Am. 63(4): 1175-1180. Koziel, M. 1996. Transgenic maize for the control of European corn borer and other
maize insect pests. Ann. NY Acad Sci. 792: 164-171. Letourneau, D. K. 1987. The enemies hypothesis: tritrophic interactions and vegetational
diversity in tropical agroecosystems. Ecology. 68(6): 1616-1622. Levins, R. and R. H. MacArthur. 1969. A hypothesis to explain the incidence of
monophagy. Ecology. 50:910-11
62
Liu, Y., B. E. Tabashnik, and M. W. Johnson. 1995. Larval age affects resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 88(4): 788-792.
Losey, J. E., D. D. Calvin, M. E. Carter, and C. E. Mason. 2001. Evaluation of noncorn
host plants as a refuge in a resistance management program for European corn borer (Lepidoptera: Crambidae) on Bt-corn. Environ. Entomol. 30(4) 728-735.
Losey, J. E., M. E. Carter, and S. A. Silverman. 2002. The effect of stem diameter of
European corn borer behavior and survival: potential consequences for IRP in Bt-corn. Entomol. Exp. Appl. 105: 89-96.
Mallet, J. and P. Porter. 1992. Preventing insect adaptation to insect-resistant crops: are
seed mixtures or refugia the best strategy? P. Biol. Sci. 250(1328): 165-169. Mason, C. E., M. E. Rice, D. D. Calvin, J. W. Van Duyn, W. B. Showers, W. D.
Hutchison, J. F. Witkowski, R. A. Higgins, D. W. Onstad, and G. P. Dively. 1996. European corn borer ecology and management. North Central Regional Extension Publication No. 327.
Mattson, W. J. Jr. 1980. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol.
Syst. 11:119-161. Maxwell, F. G. and P. R. Jennings. 1980. Breeding plants resistant to insects. Wiley. New
York. Metcalf, R. L., A. M. Rhodes, R. A. Metcalf, J. Ferguson, E. R. Metcalf, P. Lu. 1982. Cucurbitacin contents and diabroticite feeding upon cucurbita spp. Environ. Entomol. 11(4): 931-937
Microsoft. Microsoft Excel. Redmond, Washington: Microsoft, 2011. Mooney, E. H., E. J. Tiedeken, N. Z. Muth, and R. A. Niesenbaum. 2009. Differential
induced responses to generalist and specialist herbivores by Lindera benzoin (Lauraceae) in sun and shade. Oikos. 118: 1181-1189.
Niesenbaum, RA. 1996. Linking herbivory and pollination: Defoliation and selective
fruit abortion in Lindera benzoin. Ecology. 77(8): 2324-2331. Pepper, B. P. and L. A. Carruth. 1945. A new plant insecticide for control of the
European corn borer. J. Econ. Entomol. 38: 59-66. Prudic, K. L., J. C. Oliver, and M. D. Bowers. 2005. Soil nutrient effects on oviposition
preference, larval performance and chemical defense of a specialist insect herbivore. Oecologia 143:578–587.
63
Rasband, W.S., ImageJ, U. S. National Institutes of Health, Bethesda, Maryland, USA, http://imagej.nih.gov/ij/, 1997-2015.
Rasmann, S. and A. A. Agrawal. 2009. Plant defense against herbivory: progress in
identifying synergism, redundancy, and antagonism between resistance traits. Curr. Opin. Plant Biol. 12: 473-478.
Rausher, M. D. 1983. Ecology of host- selection behavior in phytophagous insects. In
variable plants and herbivores in natural and managed systems, ed. R. F. Denno, M. S. McClure, pp. 223-57. New York: Academic
Rausher, M. D. 2001. Coevolution and plant resistance to natural enemies. Nature. 411:
857-864. Razze, J. M., C. E. Mason, and T.D. Pizzolato. 2011. Feeding behavior of neonate
Ostrinia nubilalis (Lepidoptera: Cambidae) on Cry1Ab Bt Corn: implications for resistance management. J. Econ. Entomol. 104(3): 806-813.
Rosenthal, J. P. and R. Dirzo. 1997. Effects of life history, domestication and agronomic
selection on plant defense against insects: evident from maizes and wild relatives. Evol. Ecol. 11: 337-355.
Ross, S. E., and K. R. Ostlie. 1990. Dispersal and survival of early instars of European
corn borer (Lepidoptera: Pyralidae) in field corn. J Econ. Entomol. 83: 831-836. Salazar, D. and R. J. Marquis. 2012. Herbivore pressure increases toward the equator.
PNAS. 109: 12616-12620. Schoonhoven, L. M., J. J. A. van Loon, M. Dicke. 2005. Insect-plant biology. Oxford,
NY: Oxford University Press. Secchi, S., T. M. Hurley, B. A. Babcock, and R. L. Hellmich. “Managing European corn
borer resistance to Bt corn with dynamic refuges” Regulating Agricultural Biotechnology: Economics and Policy. New York: Springer Science and Business Media, 2006. 559-577.
Singer, M.S., E.A. Bernays, and Y. Carrière. 2002. The interplay between nutrient
balancing and toxin dilution in foraging by a generalist insect herbivore. Anim. Behav. 64:629-643.
Slansky, F. and P. Feeny. 1977. Stabilization of the rate of nitrogen accumulation by
larvae of the cabbage butterfly on wild and cultivated food plants. Ecol. Monogr. 47: 209-228.
Smith, C. M., D. J. Schotzko, R. S. Zemetra, and E. J. Souza. 1994. Categories of
resistance in plant introductions of wheat resistant to Russian wheat aphid
64
(Homoptera: Aphididae). J. Econ. Entomol. 85: 1480. Tallamy, D. W. 1985. Squash beetle feeding behavior: an adaptation against induced
cucurbit defenses. Ecology. 66: 1574-1579. Tallamy, D. W., J. Stull, N. P. Ehresman, P. M. Gorski, and C. E. Mason. 1997.
Cucurbitacins as feeding and oviposition deterrents to Insects. Environ. Entomol. 26: 678-83.
Tate, C. D., R. L. Hellmich, and L. C. Lewis. 2006. Evaluation of Ostrinia nubilalis
(Lepidoptera: Crambidae) neonate preferences for corn and weeds in corn. J. Econ. Entomol. 99: 1987-1993.
193-218. Timmins, W. A., K. Bellward, A. J. Stamp, and S. E. Reynolds. 1988. Food intake,
conversion efficiency, and feeding behavior of tobacco hornworm caterpillars given artificial diet of varying nutrient and water content. Physiol. Entomol. 13: 303-314.
United States Census Bureau. 2015. U.S. and World Population Clock.
(http://www.census.gov/popclock/?intcmp=home_pop) [accessed 9 May 2015]. United State Department of Agriculture. 2015. Crop Production 2014 Summary.
(http://www.usda.gov/nass/PUBS/TODAYRPT/cropan15.pdf) [accessed 9 May 2015]
Van Zandt, P. A. and A. A. Agrawal. 2004. Specificity of induced plant responses to
specialist herbivores of the common milkweed Asclepias syriaca. OIKOS. 104: 401-9.
Webb, D. R., C. J. Eckenrode, and M. H. Dickson. 1987. Variation among green and wax
beans in survival of larvae of a bivoltine-E Race of the European corn borer. J. Econ. Entomol. 80: 521-524
White, T. C. R. 1984. The abundance of invertebrate herbivores in relation to the
availability of nitrogen in stressed food plants. Oecologia. 63:90–105. Wink, M. 1988. Plant breeding: importance of plant secondary metabolites for protection
against pathogens and herbivores. Theor. Appl. Genet. 75: 225-233. Wink, M. 2013. Evolution of secondary metabolites in legumes (Fabaceae). S. Afr. J.
Bot. 89: 164-175.
65
World Hunger Education Service. 2015. 2015 World Hunger and Poverty Facts and Statistics.(http://www.worldhunger.org/articles/Learn/world%20hunger%20facts%202002.htm) [accessed 9 May 2015].
Woods, H. A. 1999. Patterns and mechanisms of growth of fifth-instar Manduca sexta
caterpillars following exposure to low- or high-protein food during early instars. Physiol. Biochem. Zool. 72:445–454.
Zalucki, M. P., A. R. Clarke, and S. B. Malcom. 2002. Ecology and behavior of first
After reviewing the results from the no choice/choice experiments in comparison
to the survivorship experiments, it was noticed that there was an inconsistency with
individuals fed cucumber. No individuals were recovered and there was very minimal
feeding damage on cucumber plants in any of the greenhouse trials where the insect was
provided with a full, intact plant (Figure 4), however, when provided with an excised leaf
disk, 3rd instar larvae fed abundantly (Figure 7). When larvae were given a choice
between near isoline and cucumber leaf disks, they fed significantly more on the
cucumber than corn (Figure 9A). One difference is that greenhouse experiments used the
whole plant while no choice and choice experiments used a leaf disk punched from the
plant.
This led to hypothesize that ECB induces chemical defenses on full cucumber
plants, therefore, preventing continual feeding. Punching out a leaf disk maybe artificially
trenching the plant, preventing the flow of defenses to the leaf tissue. This would allow
the insect to feed without harm. To test this hypothesis, the greenhouse and choice tests
were combined into one experiment. Cucumber plants were “induced” by infesting the
plants with larvae, and a choice test was conducted between leaf disks taken from
infested and non-infested cucumber plants. Hypothetically, the leaf disks from the
infested plant should have induced defenses to deter feeding while the leaf disks from the
non-infested plants should represent artificially trenched leaf tissue, yielding no plant
defenses.
67
In the greenhouse, twenty cucumber plants were raised until they accumulated
1384.5 degree-days after seeding in the greenhouse. Fifty third instars were placed in
individual empty 37 mL Solo® (Lake Forest, IL, USA) plastic cups to starve overnight
(approximately 18 hours). The next morning, five of the starved third instars were placed
on separate leaves of 10 of the cucumber plants to prevent crowding and initiate induced
defenses. The other 10 cucumber plants were left uninfested. All 20 Plants were covered
with a cage created from thin mesh fabric (JoAnn Fabric®, Hudson, OH, USA) and an 84
cm galvanized steel wire round tomato cages (Lowes®, Hebei, China) to keep conditions
consistent between treatments and keep the larvae on the plant. Plants were placed on
opposite sides of the greenhouse to prevent “talking”, communication via volatiles when
a defense is induced (Baldwin et al., 2006). Meanwhile, forty new 3rd instars were placed
in empty cups in the growth chamber and starved for three hours.
After three hours, four leaves were collected from each of the plants starting with
the oldest leaf and the next three in order. Two leaf disks from each leaf (1.75 cm in
diameter) were cut using a cork borer (Humboldt®, Elgin, IL, USA). A total of four disks
were photographed and placed in a 90 mm petri dish (Fisher Scientific®, Waltham, WA,
USA) containing 90 mm filter paper (Whatman®, Buckinghamshire, UK) dampened with
distilled water to retain moisture. Two disks from the same leaf were placed diagonally
across from each other. Infested plants were compared to uninfested plants using leaves
of the same age. One 3-hour starved 3rd instar larva was placed in the center of the four
leaf disks. Each petri dish was wrapped in parafilm (American Can Company®,
Greenwich, CT, USA) to prevent escaping larvae, as well as desiccation of larvae or leaf
tissue. After four hours, the arenas were opened and leaf disks were photographed. The
68
same larva and disks were placed back into the arena. Twenty-four and 48 hours after the
start of the experiment, the arenas were opened and photographed again.
After 48 hours, the individual was removed. Using photographs and ImageJ
software, total area of leaf tissue consumed was calculated. The leaf disks were dried and
weighed. Weight for the leaf area not consumed per host plant species was calculated to
estimate the dry biomass of leaf tissue consumed by the individual. Data were analyzed
as a proportion of biomass consumed of infested compared with non-infested and
analyzed using a one-sample t-test in the program Excel.
To analyze this data set, feeding at different time points was compared among the
leaf ages. All leaves of the same age were compared to one another. Since there was no
significant difference based on leaf age (P < 0.1354), we compiled all of the data into one
analysis to compare feeding abundance at each time point (Figure 10). After 4 hours,
there was very minimal feeding on either infested or non-infested disks (Figure 10A).
After 24 hours, there was a moderate amount of feeding overall (Figure 4B). After 48
hours, there was a good amount of feeding; some of the disks were 75-100% consumed
(Figure 4C). None of the experiments yielded significant results. Based on this data, it
was concluded that ECB is not inducing defenses in cucumber.
69
Figure 10 Cucumber induced defenses. 3rd instars given a choice between leaf tissue from an infested or non-infested plant for 4 hours (Figure 4A), 24 hours (Figure 4B), and 48 hours (Figure 4C). Bars represent the estimated amount of dry biomass consumed. Error bars show SEM; no significance was detected (P > 0.05).