EVIDENCE THAT RENAL SODIUM EXCRETION BY NORMAL HUMAN SUBJECTS IS REGULATED BY ADRENAL CORTICAL ACTIVITY1 By ALEXANDER LEAF AND WILLIAM T. COUTER WITH THE TECHNICAL ASSISTANCE OF MARION LUTCHANSKY AND ANN REIMER (From the Department of Internal Medicine, The Medical School, University of Michigan, Ann Arbor, Mich.) (Received for publication February 25, 1949) When a normal human being is placed on a very low sodium intake, the urinary sodium decreases to very small amounts. Contrariwise, with large sodium intakes, the urinary sodium increases pro- portionately. In both cases, the normal body content of sodium is quickly reestablished. How is this conservation of body sodium effected? The depletion of body sodium that occurs in adrenalectomized animals and in patients suffer- ing from Addison's disease, and the retention of sodium with reestablishment of balance that ac- companies the administration of adrenal cortical extracts, leaves no doubt about the fundamental position of a cortical hormone in the regulation of body sodium in the abnormal states just men- tioned (1-5). ' Since there is a paucity of information regarding the regulation of body sodium content in normal subjects who receive large variations in intake of sodium, our 'studies were designed to learn 'the role of adrenal cortical activity in this regulation in normal human beings. Even though it is not possible to determine di- rectly the concentration of the specific hormone in the blood, nevertheless a number of methods that serve as indirect indices of adrenal cortical activity are available. It was surmised that by creating a need to conserve body sodium in the presence of large shifts in intake, evidence of other adrenal cor- tical effects, especially on protein metabolism, might be elicited. If such evidence of adrenal cortical activity can be obtained by the require- ment to conserve sodium, such data would greatly strengthen the hypothesis that in the normal sub- ject this conservation of body sodium is in fact mediated through an increased or decreased adrenal cortical activity (6). 1 This study was aided by a grant from the United States Public Health Service. EXPERIMENTAL Three normal young adults were subj ects for this study: one male resident physician (W. C.), age 32; one male medical student (E. W.), age 22; and one female graduate student (M. L.), age 22. The plan of study was to place the subject on an adequate caloric and protein dietary intake with a very low sodium con- tent (approximately 200 mgs. or 9 mEq. daily) through- out the study, and at definite periods to administer orally large amounts of sodium as sodium chloride or sodium citrate. This was accomplished by using a constant daily diet. The water intake was uncontrolled as its sodium content was found to be insignificant by analysis. The subjects went about their usual daily activities avoiding severe exertion and sweating. Urine and stool collections were made daily and the nude body weight was recorded each morning before breakfast and after emptying the bladder. A special human balance having a sensitivity of 5 gms. was used. Each 24 hour urine was collected with toluene as a preservative and was analyzed separately on the day following the collection. Stools were promptly treated with alcohol and made acid with glacial acetic acid or sulfuric acid, dried in an oven at 500 to 60' C., weighed, blended in a Waring blender and combined into two or three day periods with suitable aliquots taken for analy- ses. Subjects were fed from the same lot of food throughout the period of study and a sample day's diet was dried, blended and analyzed. When a constant body weight was achieved and the urine sodium and chloride excretions had reached a minimum on this low sodium intake, 11.0 gms. or 478 mEq. of sodium were administered orally as the chloride (28.0 gms. of sodium chloride) or citrate (46.88 gms. of sodium citrate) daily for three days. Fasting blood samples were obtained with a minimum of stasis at the beginning and end of the study, and before and after each period of sodium administration. Samples for potas- sium and carbon dioxide content were drawn under oil and the former kept refrigerated until the serum was separated from the cells with no hemolysis occurring. Hematocrit (packed red cell volume) was estimated by the method of Wintrobe (7). Sodium analyses were done on serum, urine, diet, and stools by means of the Butler and Tuthill (8) modification of the uranyl zinc acetate method of Barber and Kolthoff. For urines of very low sodium content a modification involving 50 cc. 1067
Transcript
EVIDENCE THAT RENAL SODIUM EXCRETION BY NORMALHUMANSUBJECTS IS REGULATEDBY ADRENAL
CORTICAL ACTIVITY1
By ALEXANDERLEAF AND WILLIAM T. COUTERWITH THE TECHNICAL ASSISTANCE OF MARIONLUTCHANSKYAND ANN REIMER
(From the Department of Internal Medicine, The Medical School, University of Michigan,Ann Arbor, Mich.)
(Received for publication February 25, 1949)
Whena normal human being is placed on a verylow sodium intake, the urinary sodium decreasesto very small amounts. Contrariwise, with largesodium intakes, the urinary sodium increases pro-portionately. In both cases, the normal bodycontent of sodium is quickly reestablished. Howis this conservation of body sodium effected?
The depletion of body sodium that occurs inadrenalectomized animals and in patients suffer-ing from Addison's disease, and the retention ofsodium with reestablishment of balance that ac-companies the administration of adrenal corticalextracts, leaves no doubt about the fundamentalposition of a cortical hormone in the regulation ofbody sodium in the abnormal states just men-tioned (1-5). '
Since there is a paucity of information regardingthe regulation of body sodium content in normalsubjects who receive large variations in intake ofsodium, our 'studies were designed to learn 'therole of adrenal cortical activity in this regulationin normal human beings.
Even though it is not possible to determine di-rectly the concentration of the specific hormone inthe blood, nevertheless a number of methods thatserve as indirect indices of adrenal cortical activityare available. It was surmised that by creating aneed to conserve body sodium in the presence oflarge shifts in intake, evidence of other adrenal cor-tical effects, especially on protein metabolism,might be elicited. If such evidence of adrenalcortical activity can be obtained by the require-ment to conserve sodium, such data would greatlystrengthen the hypothesis that in the normal sub-ject this conservation of body sodium is in factmediated through an increased or decreasedadrenal cortical activity (6).
1 This study was aided by a grant from the UnitedStates Public Health Service.
EXPERIMENTAL
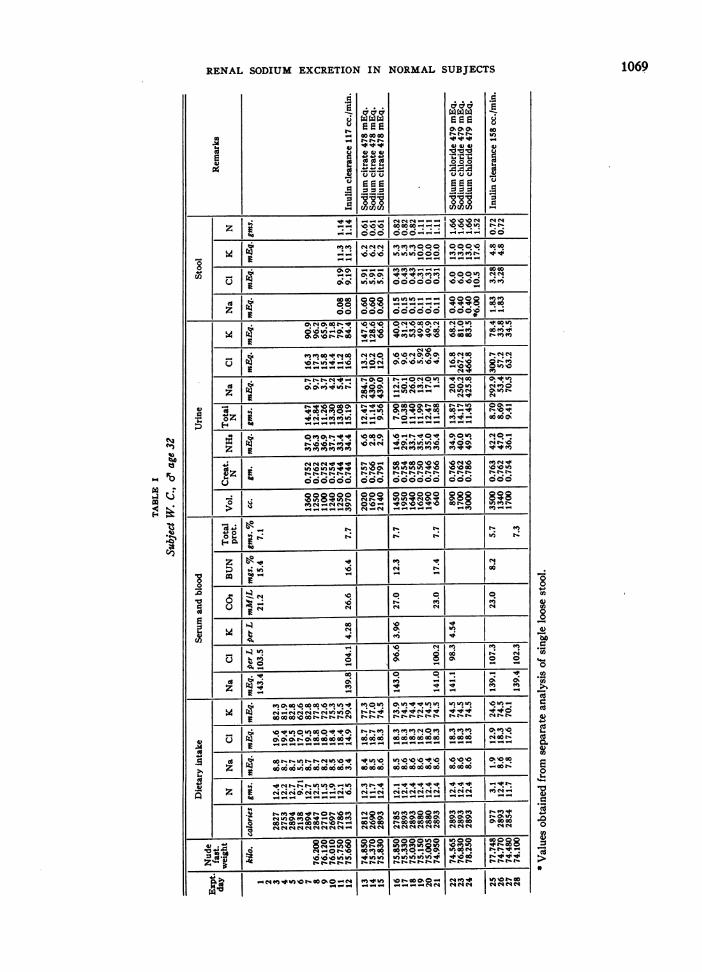
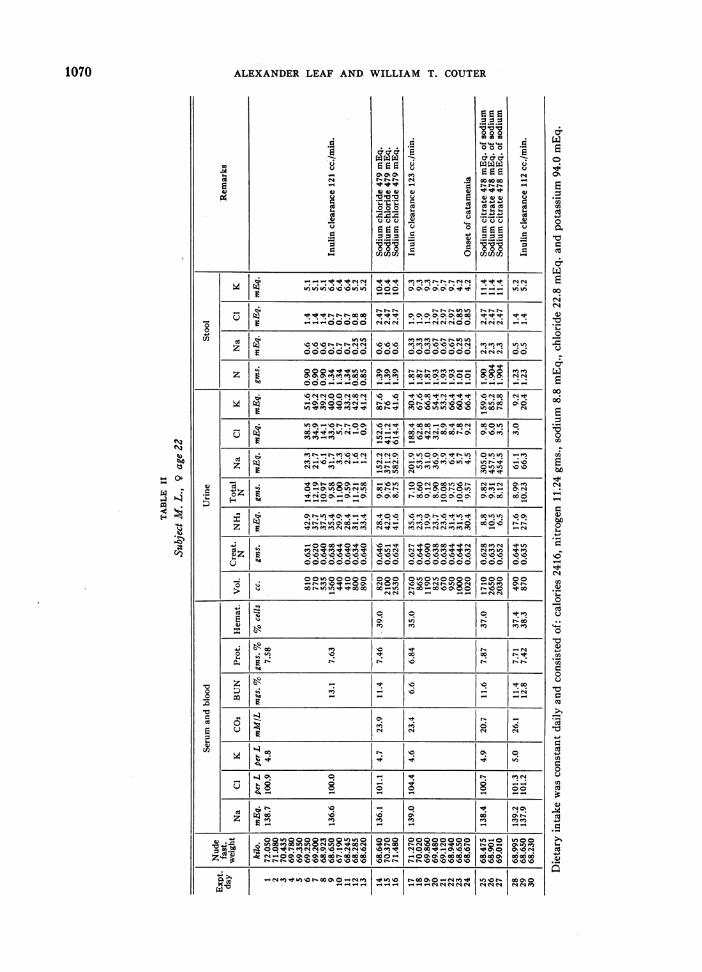
Three normal young adults were subj ects for thisstudy: one male resident physician (W. C.), age 32;one male medical student (E. W.), age 22; and onefemale graduate student (M. L.), age 22. The plan ofstudy was to place the subject on an adequate caloricand protein dietary intake with a very low sodium con-tent (approximately 200 mgs. or 9 mEq. daily) through-out the study, and at definite periods to administer orallylarge amounts of sodium as sodium chloride or sodiumcitrate.
This was accomplished by using a constant daily diet.The water intake was uncontrolled as its sodium contentwas found to be insignificant by analysis. The subjectswent about their usual daily activities avoiding severeexertion and sweating. Urine and stool collections weremade daily and the nude body weight was recorded eachmorning before breakfast and after emptying the bladder.A special human balance having a sensitivity of 5 gms.was used. Each 24 hour urine was collected with tolueneas a preservative and was analyzed separately on the dayfollowing the collection. Stools were promptly treatedwith alcohol and made acid with glacial acetic acid orsulfuric acid, dried in an oven at 500 to 60' C., weighed,blended in a Waring blender and combined into two orthree day periods with suitable aliquots taken for analy-ses. Subjects were fed from the same lot of foodthroughout the period of study and a sample day's dietwas dried, blended and analyzed.
When a constant body weight was achieved and theurine sodium and chloride excretions had reached aminimum on this low sodium intake, 11.0 gms. or 478mEq. of sodium were administered orally as the chloride(28.0 gms. of sodium chloride) or citrate (46.88 gms.of sodium citrate) daily for three days. Fasting bloodsamples were obtained with a minimum of stasis at thebeginning and end of the study, and before and aftereach period of sodium administration. Samples for potas-sium and carbon dioxide content were drawn under oiland the former kept refrigerated until the serum was
separated from the cells with no hemolysis occurring.Hematocrit (packed red cell volume) was estimated
by the method of Wintrobe (7). Sodium analyses weredone on serum, urine, diet, and stools by means of theButler and Tuthill (8) modification of the uranyl zincacetate method of Barber and Kolthoff. For urines ofvery low sodium content a modification involving 50 cc.
1067
ALEXANDERLEAF AND WILLIAM T. COUTER
aliquots and removal of potassium as well as phosphatewas utilized (9). Potassium was determined in serum,urine, diet and stool by a modification of the platinicchloride micro colorimetric method of Shohl and Bennett(10). Chloride analyses of serum were done using themethod of Van Slyke and Sendroy (11). Urine, stool,and diet chloride determinations were done by a modifica-tion of the Volhard-Harvey titration (12). Urinary am-
monia was determined by the aeration method of VanSlyke and Cullen (13). Total urine, diet and stool nitro-gen were determined by the Kjeldahl method for macro
analysis (14). Urine creatinine was measured by thealkaline picrate method of Folin (15) and in subjectE. W., also by the modification of Bonsnes and Taussky(16). Agreement between the very simple method ofFolin and the more refined latter method was surprisinglygood. The modification of Bonsnes and Taussky was alsoadopted for serum creatinine determinations. Blood urea
nitrogen was estimated by the method of Gentzkow (17)while for urine urea the method of Barker (18) was
utilized. Urine uric acid determination was done by thetotal color method of Buchanan, Block, and Christman(19). Carbon dioxide content of the serum was deter-mined by the method of Van Slyke and Neill (20).Serum protein analyses were done by the micro Kjeldahlmethod and the steam distillation of Goebel (21). Urine,stool and diet were analyzed for phosphate by the colori-metric method of Fiske and Subbarow (22).
The aliquots of diet and stools were dry ashed in sucha manner that analyses could be made for sodium, chlo-ride, potassium, and phosphorus on a single preparation.This was accomplished by mixing the aliquot with an
excess of calcium hydroxide until definitely alkaline,drying and then ashing in the muffle furnace at 400° C.for 48-72 hours. The residue was washed with waterand filtered. The filtrate containing the sodium, chloride,and potassium was made up to volume in a volumetricflask from which aliquots were taken for analysis. Thewashed residue was later dissolved with dilute nitric acidto yield phosphate. Values for chloride of diet and stoolobtained by this method were the same as values obtainedby wet ashing the sample.
Inulin clearances were done by the Harrison modifica-tion of the method of Alving, Rubin, and Miller (23)following a single intravenous injection of 10-20 gms.of inulin.
The values for dietary intake of calories, carbohydrate,fat and protein were obtained from Bowes and Church(24) and U. S. Department of Agriculture CircularNumber 549. The calculated values for sodium, chloride,potassium and phosphorus content of foods were obtainedfrom McCance and Widdowson (25). In addition, analy-ses were made on complete diets (with the exception ofthe butter) for total nitrogen, total phosphorus, sodium,chloride, and potassium. The agreement between actualanalyses and calculated values was surprisingly good.Thus in subject M. L.'s diet the calculated and analyzedcomposition was respectively: nitrogen-12.3 and 11.2
gms.; sodium-8.7 and 8.3 mEq.; chloride 2-21.0 and22.8 mEq.; and potassium-89.7 and 94.0 mEq. Simi-larly in subject E. W.'s diet the calculated and analyzedcomposition was respectively: nitrogen-14.3 and 15.0gms.; sodium-9.1 and 9.4 mEq.; chloride-24.4 and 24.9mEq.; and potassium-100 and 100 mEq. The dietaryintake figures recorded for subjects M. L. and E. W.in Tables II and III are values obtained by our analysesfor sodium, chloride, nitrogen, potassium and phosphorusas no diet changes were made and all the food was eaten.In subject W. C. who frequently rejected portions of hisdiet, such portions were weighed and their composition asobtained from the above-mentioned food compositiontables was subtracted from the total calculated contentof the diet, and each day's intake is recorded in Table I.
RESULTS
The complete data of these studies are shownin Tables I, II and III.
The electrolyte and weight changes are the sub-ject of a separate report (6). However, it isimportant to the present discussion to point outthat with the curtailment of sodium intake toabout 9 mEq. daily there occurred a sharp initialweight loss of about 2 kilograms in each subject.This weight loss was associated with a negativesodium and chloride balance (Figure 1 illustratesthese changes in subject E. W.). After approxi-mately one week of this low sodium regimen theurinary loss of sodium and chloride was reduced toextremely minute amounts (1 to 2 mEq. of sodiumin the 24 hour urine specimen) and the weight be-came stabilized. Under the stimulus of such severesodium restriction the kidneys conserve sodiummaximally.
On administration of the large amounts of so-dium (480 mEq. daily for three days) as the so-dium chloride or sodium citrate salts there waswith each a significant weight gain. None of thesubjects showed clinical edema, however. Theurinary excretion of sodium lagged in every caseso that only by the last day of the sodium adminis-tration had the output attained the daily intakelevel.
With these changes in weight and in urinary ex-cretion of sodium and -chloride, accompanying
2 Interestingly the diet analyses, not including "Lona-lac," yielded nearly equivalent values for sodium andchloride. The addition of "Lonalac" (Mead Johnson andCo., Evansville, Indiana), which contains supplementarypotassium chloride, resulted in the actual higher valuesof chloride.
1068
RENAL SODIUM EXCRETION IN NORMALSUBJECTS
3
UcC;
j-u4d
1-
00. ..4141I C
)co aCco
* -* v
U Us
s c- c-
I2VCQt
F .E
010,0104 1414
00
Uu
A
U
4)UU
coo.oo o-lo000 000V @O.
UjS o o o~~ I o o o Io oo 4 E -_ito tooooerO~~~~~~~~~~~~~~~~~~~~~~~~~N_O14M_M~n~o"Ie
FIG. 1. To SHOWCHANGESIN BODY WEIGHT, SERUMSODIUM AND CHLORIDE CONCENTRATIONS,AND SODIUM BALANCEIN SUBJECT E. W.
changes were noted in extracellular fluid composi-tion. The sodium restriction uniformly caused asmall but significant lowering of serum sodiumconcentration and elevations occurred on adminis-tration of sodium. Serum chloride values followeda similar trend though less uniformly. Adminis-tration of the sodium citrate caused a rise in serumsodium in each case, a decrease in serum chloridein two cases and a rise in serum CO2 content. Aparallel rise in serum sodium and chloride followedsodium chloride administration. With the sodiumcitrate administration urinary sodium excretionrose sharply to high levels but urinary chloride losswas, if anything, further decreased. This is ad-ditional evidence that the kidneys can deal sepa-rately with these two most important ions of theextracellular fluid.
There is slight variability in stool sodium andchloride excretion. But considering the drasticchanges in oral sodium and chloride intake fromabout 10 to 500 mEq. daily the stool losses re-mained surprisingly fixed and insignificantly small.
Fecal potassium excretion also showed no note-worthy variation.
Subject W. C. had one loose stool associatedwith abdominal cramps on the last day of sodiumchloride administration. This one stool was col-lected and analyzed separately. Its sodium andchloride content was much higher than the re-maining stools of that period.
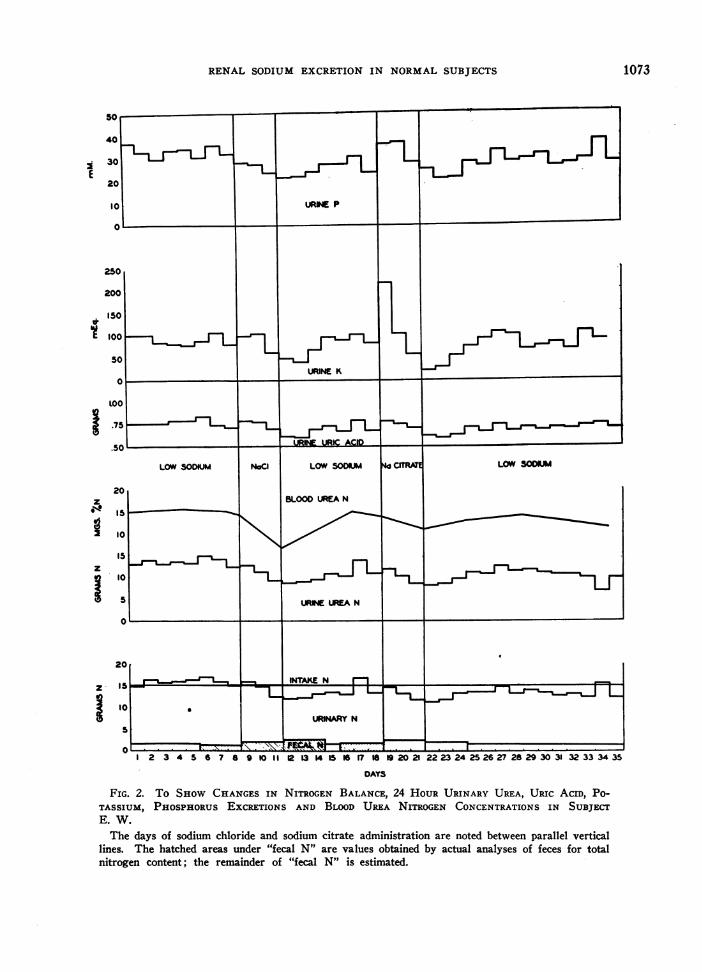
Of greatest importance to this discussion arethe observed changes in protein metabolism thatoccurred with variation of sodium intake. Figure2 shows graphically the changes in nitrogen bal-ance, urine urea excretion, blood urea nitrogenconcentration, urine total uric acid excretion,urine potassium and phosphorus excretions insubject E. W. Exactly similar changes were ob-served in our other two subjects but because dataare more complete in our last study on E. W.his results have been graphed. Sodium re-striction is shown to produce slightly negativenitrogen balance during the first low sodium pe-riod. Following both periods of sodium adminis-tration the urine plus fecal nitrogen is considerably
1072
RENAL SODIUM EXCRETION IN NORMALSUBJECTS
20
10 URINE P
*SISOeD
FIG. 2.T HWCAGSI IRGNBLNE,2 ORUIAYUEUICDoURINE-
.5UINTAEURIC ACV
TIUM PHOSPHOR ECRETIN AND BLOOD UREAN IOIO IN SBE
E. W.The days of sodium chloride and sodium citrate administration are noted between parallel vertical
lines. The hatched areas under "fecal N" are values obtained by actual analyses of feces for totalnitrogen content; the remainder of "fecal N" is estimated.
1073
ALEXANDERLEAF AND WILLIAM -T. COUTER
less than the intake of nitrogen so that nitrogen isstored in the body. The fecal nitrogen reveals thatthis decreased urinary nitrogen excretion was notthe result of impaired nitrogen absorption fromthe alimentary tract.
Creatinine and ammonia excretion remained es-sentially constant so that the changes in urinarynitrogen excretion could be expected to be theresult of changes in "metabolic" nitrogen. Theurine urea excretion is seen to confirm this ex-pectation since all changes in total urinary nitro-gen are due almost entirely to changes in urineurea nitrogen (the difference between total urinenitrogen and urine urea nitrogen remaining veryconstant throughout the study).
That the changes in total and urea nitrogenexcretion are not simply the result of storage ofurea in the body fluids is indicated by the dramaticchanges seen in the blood urea nitrogen. At thetime when the urinary excretion of urea is at itslowest the blood urea concentrations are likewiselowest. Correcting the blood urea concentrationfor the dilution that might occur as the result ofthe increased body fluid volume produced by thesodium administration causes only insignificantchanges. The serum creatinine was measured inthe subject E. W. and though it shows minorfluctuations it in no way parallels changes inblood urea level. It has been amply proven thatthe serum creatinine concentration is independentof the level of protein catabolism (26). As therehas occurred no impairment of gastrointestinalabsorption of nitrogen these changes can be in-terpreted only as the result of a decreased rateof urea production or a decreased catabolism ofprotein associated with administration of sodiumto individuals previously maintained on low so-dium diets. These effects on protein metabolismoccur following both sodium citrate and sodiumchloride administration and therefore are the re-sult of the sodium rather than of the accompany-ing anion.
Urine uric acid excretion, urine potassium andphosphorus excretion mimic the changes in nitro-gen metabolism exactly except for the very highrate of potassium excretion on the first day of so-dium citrate administration. This change is be-lieved to be due to effects other than those on pro-tein metabolism (6). Serum potassium concen-trations failed to show any evidence of increasing
with the periods of low urine potassium excretionthough unfortunately no serum potassiums weretaken on the third or fourth days after the highsodium periods when this effect might have beenpresent.
A point to be stressed is that there occurred adefinite lag in the renal excretion of sodium duringeach high sodium period. Not until the third andlast day of each high sodium period did the urinaryexcretion reach the level of the intake. Our inter-pretation of this lag is discussed below but here itshould be noted that all the effects associated withthe depressed protein catabolism were most pro-nounced immediately following rather than duringthe periods of high sodium administration, thusparalleling in this respect the lag in sodium ex-cretion.
Urines were tested daily for reducing substanceby the qualitative Benedict's test on subjects M.L. and E. W. but no evidence of glycosuria wasnoted. Glucose tolerance tests were done on sub-jects M. L. and E. W. M. L. showed no evidenceof impaired glucose tolerance on the 11th day ofthe low sodium intake. (Standard glucose toler-ance test after 1.75 gms. of glucose per kilogramof body weight revealed: fasting blood sugar-75mgs.%; one hour-71 mgs.%; two hours-82mgs.%; three hours-51 mgs.%; four hours-56 mgs.%o.) E. W. on the 12th day of the finallow sodium intake period showed: fasting bloodsugar-89 mgs.%o; after one hour-160 mgs.%second hour-100 mgs.%o; third hour-56 mgs.%o.A repeat glucose tolerance test on E. W. twomonths after completion of this study and whileon a normal dietary sodium intake revealed: fast-ing blood sugar-81 mgs.%o; after one hour-153mgs.o; after two hours-100 mgs.%o; and afterthree hours-64 mgs.%.
DISCUSSION
Changes in nitrogen metabolism similar to thosereported here have been noted by others. Instudying experimental sodium deficiency pro-duced by a low sodium intake and sweating innormal subjects McCance (27) found that his sub-jects developed negative nitrogen balances withrise in blood urea concentration. This observationwas striking enough to be emphasized by the au-thor. In a subsequent publication, McCance andWiddowson (28) conclude that the main cause
1074
- RENAL SODIUM EXCRETION IN NORMALSUBJECTS
of this elevation of blood urea concentration withsodium depletion is the associated reduction inglomerular filtration rate. In another study Mc-Cance (29) observed that the administration oflarge doses of sodium chloride and sodium bi-carbonate depressed the serum urea concentrationsignificantly in previously dehydrated subjects.The electrolyte balance study of Albright andBauer (30) on an edematous nephritic likewiseshows an effect of administered sodium chlorideand sodium bicarbonate on the nitrogen balance.Both sodium salts produced positive nitrogen bal-ance. Landis et al. (31) reported striking changesin blood urea nitrogen levels in three subjectswith varying degrees of renal insufficiency whenthe daily amount of sodium chloride intake wasaltered. Sodium chloride restriction caused in-creased blood urea nitrogen concentrations whileadministration of this salt lowered the concentra-tion. These workers did 24 hour urea and creati-nine clearances finding some increase in theseclearances during the periods of sodium chlorideadministration. The decrease in blood urea ni-trogen with administration of sodium chloride wasassociated with the increased renal clearances.
It is to be expected that administration of so-dium chloride will increase glomerular filtrationrate and hence urea clearance. Contrariwise, re-striction of sodium intake in non-edematous pa-tients with renal insufficiency is likely to producesevere dehydration through excessive renal lossof sodium with shrinkage of extracellular fluidvolume. Hence, the increase in glomerular filtra-tion rate with sodium administration is especiallyto be anticipated in patients with renal insufficiency(as were those of Landis et al.) who may be de-hydrated prior to administration of the sodium.
The gain in weight observed in our subjectswhen sodium chloride was administered was al-most entirely the result of an increase in extra-cellular fluid volume (as calculated from the gainin "chloride space" from the observed chloridebalance). In such acute experiments as ours,changes in the hematocrit reflect changes in in-travascular volume. Table IV clearly shows thatsodium chloride administration results in a de-crease in cell hematocrit with hemodilution. Thisincreased intravascular volume may be expected,other factors remaining constant, to increase renalplasma flow with resulting increased glomerular
TABLE IV
E. W.Hemodilution with increased rate of glomerular filtration
following sodium chloride but not sodium citrate
Total serum 24 hour 24 hour ureaDay Hematocritpoen
filtration rate. Another aspect of the hemodilu-tion is the decrease in total serum protein concen-tration also shown in Table IV. This decreasewould cause a decreased colloid osmotic pressureof the serum which also, other factors remainingconstant, would favor an increased rate of glo-merular filtration. The creatinine clearance fig-ures in Table IV taken as a measure of glomerularfiltration rate (32, 33) in E. W. show this ex-pected increase when sodium chloride is adminis-tered. An even proportionately greater increasein the 24 hour urea clearance is seen. This indi-cates a reduced rate of tubular reabsorption ofurea in addition to the increase in glomerular fil-tration rate. This is to be expected with thegreater rate of urine formation that followed thesodium chloride administration. Inulin clearancemeasurements of glomerular filtration rate in sub-ject W. C. on the low sodium intake and immedi-ately following the high sodium chloride periodreveal exactly similar changes in glomerular fil-tration rate (117 cc./min. after 12 days of lowsodium intake and 158 cc./min. the day after thesodium chloride administration) with the sameevidence of increased plasma volume and hemo-dilution. Female subject M. L. however showedno significant change in inulin clearance (121 cc./min. on the ninth day of low sodium intake and 123
1075
ALEXANDERLEAF AND WILLIAM T. COUTER
cc./min. and 112 cc./min. on the days followingthe sodium chloride and sodium citrate period re-spectively). She also showed much less evidenceof hemodilution than did the other two subjects.Sodium citrate produced no noteworthy changesin renal clearances in any subject.
Thus sodium chloride administration may tosome degree lower the blood urea concentrationby producing increased renal clearances in somecases. In our normal subjects the effect of al-tered renal dynamics on the blood urea concen-tration was, however, of lesser importance. Inspite of the increased urea clearance in subjectsE. W. and W. C. following sodium chloride, the24 hour urea excretion was markedly reduced.The only way an increased clearance alone candecrease blood concentrations of a substance isby an increased rate of removal of that substancefrom the body fluids. But our data in all casesreveal a sharp decrease in the rate of urea andnitrogen excretion just at the time of lowest bloodconcentrations. Sodium citrate, which failed to in-crease the glomerular filtration rate, also pro-duced a depression in blood urea concentration.In subject M. L. no noticeable increase in glo-merular filtration rate was apparent on the dayfollowing administration of sodium chloride.However, the blood urea nitrogen depressionstill occurred.
In the presence of normal alimentary absorptionof nitrogen and decreased renal excretion of it,the drop in blood concentrations of urea in our sub-jects must have been the result of a decreased rateof catabolism of protein. This accounted for thepositive nitrogen balance resulting from adminis-tration of sodium. Figure 2 clearly shows thatsuch an interpretation is justified.
The purpose of this study was to learn whetherthe physiological regulation of body sodium con-tent is controlled by adrenal cortical activity innormal persons who ingest variable amounts ofsodium. It has been amply demonstrated that theadrenal cortex produces hormones which are ca-pable of decreasing renal sodium excretion (1, 5,34) and increasing catabolism of protein (35, 36).The former has a desoxycorticosterone-like actionand has been termed the "salt-and-water" hor-mone (or hormones) while the latter is proteincatabolic or anti-anabolic and will be called the"protein catabolic" hormone in this discussion.
This does not prove, however, that in the normalsubject the desoxycorticosterone-like hormone ac-tually controls sodium balance. But if these twoapparently unrelated effects of adrenal cortical ac-tivity (i.e., on sodium and on protein) are elicitedby the single stimulus to conserve body sodium,this will afford good evidence that the adrenalcortex is importantly involved in regulation ofnormal body sodium.
With these two actions of the adrenal cortex inmind, the changes noted in Figure 2 lend them-selves readily to explanation on the basis of altera-tions of adrenal cortical activity as follows. In thefirst eight days of sodium restriction renal ex-cretion of sodium decreased to a minimum underthe stimulus of increased adrenal salt-and-waterhormone activity. At the same time the rate ofprotein catabolism resulting from increased proteincatabolic hormone activity caused a negative nitro-gen balance with the levels of excretion of urea, uricacid and potassium as charted. Administrationof large amounts of sodium chloride on the ninth,tenth and 11th days removed the requirement forrenal conservation of sodium. The adrenal re-sponded by decreasing the activity of its salt-and-water hormone output allowing the kidneys to in-crease their rate of sodium excretion enormously.At the same time the protein catabolic hormonewas likewise depressed with decreased proteincatabolism, a positive nitrogen balance, decreaseduric acid, urea, potassium and phosphorus excre-tions. Further sodium restriction from the 12thto 19th day resulted again in a decreased urinesodium excretion (increased adrenal activity).The following three day period of sodium citrateadministration reproduces the same effects of de-creased adrenal cortical function that occurred withsodium chloride administration, followed finallyby increased adrenal cortical activity when sodiumintake was again restricted during the final 14 daysof the study.
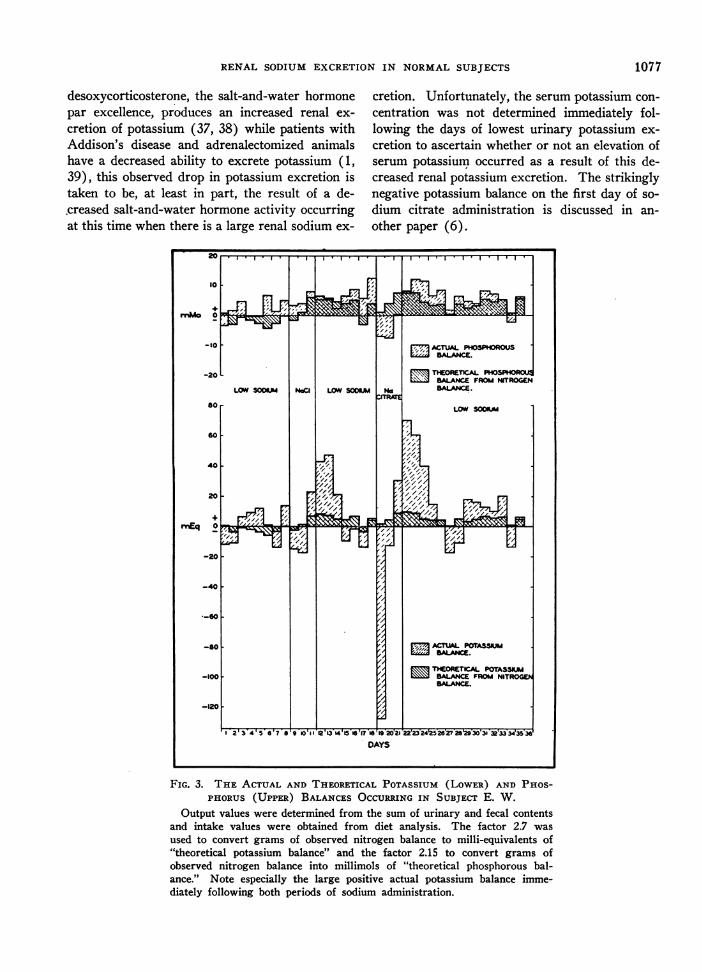
Figure 3 was constructed from the observed po-tassium and phosphorus balances of subject E. W.The theoretical potassium and phosphorus balanceswere calculated from the observed nitrogen bal-ance. It is noted from this figure that the de-creased potassium excretion that followed eachperiod of sodium administration can only partiallybe accounted for by the decreased catabolism ofprotein that occurred at that time. Knowing that
1076
RENAL SODIUM EXCRETION IN NORMALSUBJECTS
desoxycorticosterone, the salt-and-water hormonepar excellence, produces an increased renal ex-cretion of potassium (37, 38) while patients withAddison's disease and adrenalectomized animalshave a decreased ability to excrete potassium (1,39), this observed drop in potassium excretion istaken to be, at least in part, the result of a de-.creased salt-and-water hormone activity occurringat this time when there is a large renal sodium ex-
cretion. Unfortunately, the serum potassium con-centration was not determined immediately fol-lowing the days of lowest urinary potassium ex-cretion to ascertain whether or not an elevation ofserum potassium occurred as a result of this de-creased renal potassium excretion. The strikinglynegative potassium balance on the first day of so-dium citrate administration is discussed in an-other paper (6).
FIG. 3. THE ACTUAL AND THEORETICAL POTASSIUM (LOWER) AND PHOS-PHORUS (UPPER) BALANCESOCCURRINGIN SUBJECT E. W.
Output values were determined from the sum of urinary and fecal contentsand intake values were obtained from diet analysis. The factor 2.7 wasused to convert grams of observed nitrogen balance to milli-equivalents of"theoretical potassium balance" and the factor 2.15 to convert grams ofobserved nitrogen balance into millimols of "theoretical phosphorous bal-ance." Note especially the large positive actual potassium balance imme-diately following both periods of sodium administration.
1077
ALEXANDERLEAF AND WILLIAM T. COUTER
A comparison of the phosphorus and nitrogenbalances, also illustrated in Figure 3, reveals nosignificant differences between actual and theoreti-cal phosphorus balance. The theoretical phos-phorus balance was not, however, corrected forpossible changes in bone metabolism.
Our data show more distinct evidence of de-pression of adrenal cortical activity with adminis-tration of sodium than stimulation of activity byrestriction of sodium. However, with more severesodium depletion produced by sweating in ad-dition to very low sodium dietary intake, one ofMcCance's subjects had a negative nitrogen bal-ance of 45 gms. in 11 days of sodium depletionand McCance states (27) ". . . this illustratedrather well the fact that the nitrogen balancesteadily became more negative as sodium chloridewas withdrawn from the body." Conn (40) foundthat sodium restriction in normal subjects under-going acclimatization to humid heat results innegative nitrogen balance. This investigator dem-onstrated that it was the sodium restriction ratherthan a nonspecific effect of the hot environmentwhich caused the negative nitrogen balance.Thus when acclimatization to the hot environmentwas complete in his subjects while they were on afixed sodium intake, nitrogen balance was re-gained. Further restriction of sodium, withoutchanging the exposure to the hot environment orotherwise altering the diet, was again followed by anegative nitrogen balance, which disappearedgradually as reacclimatization was accomplishedon the lower sodium intake. Thus, it seems thatthe amount of evidence of increased protein cata-bolic hormone activity that is elicited in responseto sodium restriction is dependent upon the in-tensity of the requirement to conserve bodysodium.
It is also evident from our data that with pro-longation of the period of low sodium intake theeffects on nitrogen metabolism attributed toadrenal activity (protein catabolic hormone action)disappear. This occurs in spite of the continuedrenal conservation of sodium. In all three sub-jects the protein catabolic effects were most evi-dent early in the study. As mentioned above,Conn (40) found that during acclimatization to ahot environment negative nitrogen balance occursearly and gradually disappears. It is of interestin this connection to note that Swanson and Smith
(41) found that rats kept on a "salt-free" dietshowed a definite increase in weight of the adre-nals followed by regression, with adrenal weightsafter 90 days only slightly greater than in the con-trol rats receiving salt.
Apparently then, with continuation of the re-quirement to conserve body sodium, the proteincatabolic hormone effect disappears. This could.be the result of a decreased adrenal cortical out-put of protein catabolic hormones or of a decreasedresponse on the part of the tissues to the action ofan increased protein catabolic hormone in thebody. Our data give no clue as to whether one ofthe two or both possibilities actually occurs.
A further point of interest should be mentioned.That is the definite lag that occurred in the renalexcretion of sodium when it was administered toour subjects following the low sodium intake, andthe lag in the renal conservation of sodium that oc-curred when the sodium intake was again sharplyreduced. Because the changes noted in nitrogenbalance and renal potassium excretion with sodiumadministration show exactly the same lag, it ap-pears that the sudden changes in sodium intakerequire some time to produce an adrenal response.
We are well aware that this interpretation ofour results suffers considerably from the lack ofadrenal cortical hormone assays which wouldmore directly define the role of the adrenal cortex."However at the present time there is no methodof measuring directly adrenal salt-and-water hor-mone production. Forsham et al. have found in-creased uric acid excretion to be a more consistentand more marked result of prolonged administra-tion of 11 and 11-17 oxycorticosteroids than isthe nitrogen balance (42). Further studies bythis group of uric acid excretion in response toadrenocorticotrophin administration (43) sup-port the view that uric acid excretion reflectschanges in 1 1-oxycorticosteroid activity. Theobserved changes in total uric acid excretion insubject E. W. on a constant purine diet (see Figure2) clearly then support our interpretation of al-tered adrenal cortical activity in response to vary-ing sodium intake.
8 Space does not permit a discussion of possible changesin androgenic and protein anabolic hormone activity ofthe adrenal cortex and our data show no positive evidencethat such changes occurred.
1078
RENAL SODIUM EXCRETION IN NORMALSUBJECTS
Because evidence of both protein catabolic hor-mone activity and of salt-and-water hormone ac-tivity has been elicited by the need to conservebody sodium it is suggested that the adrenal re-ceived its direct stimulus via the anterior pitui-tary. Studies with adrenocorticotropic hormoneadministration to human beings support this viewsince this hormone causes stimulation of proteincatabolic effects as well as salt-and-water hor-mone effects (40, 43).
The fact that the changes observed were similarfollowing both sodium chloride and sodium citrateadministration indicates that the sodium cation isof importance rather than the anion. Just whatchange occurs in the body when sodium intakeis restricted or high, which serves to initiate theadrenal cortical activity, is not revealed. Changesin concentration of extracellular fluid, sodium,chloride, CO2 content, tonicity of extracellularfluid or volume of extracellular fluid perhapsthrough its effect on circulating intravascularvolume, all suggest themselves, but a multitudeof other changes which occur in the extra- andintracellular fluids with variation in sodium intakemight as likely act as the actual stimulus. Thework of Selye on the general adaptation syndrome(44) suggests that our observations might simplycover an adaptation to sodium restriction as thestress, but this would only further describe ratherthan explain our results. Our findings and inter-pretation are in agreement with those of Conn (40),who demonstrated an increased adrenal corticalactivity in response to the requirement to conservesalt during acclimatization to humid heat.
SUMMARYANDCONCLUSIONS
Some effects of marked variation in sodium in-take in three normal young adult subjects werestudied. These subjects were maintained on fixeddiets containing adequate calories and protein withvery low sodium and chloride content. At definiteperiods the sodium intake was abruptly increasedby the addition of 480 mEq. of sodium as sodiumchloride or sodium citrate.
With reduction from usual intakes of sodium to9 mEq. of it daily there occurred an initial nega-tive balance of sodium and chloride followed byprogressive reduction of urinary losses to minimalamounts in about five days. This initial loss ofbody sodium and chloride was associated with a
loss of about 2 kilograms of weight with subse-quent stabilization of weight as urine sodium andchloride loss became very small. Slight decreasein serum sodium and chloride levels also occurred.
Administration of sodium as sodium chlorideor as sodium citrate was accompanied by a sharpincrease in body weight much more pronouncedwith sodium chloride than with the citrate. Therealso occurred a hemodilution as evidenced by de-crease in serum proteins and hematocrit and anincreased glomerular filtration rate. These lattereffects were much more pronounced after sodiumchloride administration than after sodium citrate.
A definite lag in the renal excretion of sodiumwith the shift from the low to the high sodium in-take and a delay in the renal conservation of so-dium with change from the high to the low sodiumintake occurred.
With this variation in sodium intake definitechanges in protein metabolism occurred. Ad-ministration of sodium after sodium restrictionproduced positive nitrogen balance while sodiumrestriction increased the rate of protein catabolism.These changes were reflected in increased 24 hoururea, uric acid, phosphorus and potassium excre-tions with sodium restriction and depression ofthese urinary excretions with sodium administra-tion. In addition a definite decrease in blood ureaconcentration occurred with each period of sodiumadministration which could not be accounted forby alteration in renal clearance of urea but wasthe result primarily of a decrease in the rate ofprotein catabolism.
The decreased urinary potassium excretion fol-lowing the periods of sodium administration couldnot be entirely accounted for by the decrease inprotein catabolism that also occurred at this timeand this effect is discussed.
We believe that all the observed effects arebest explained by changes in adrenal cortical ac-tivity. In this sense the requirement for conserva-tion of body sodium in the presence of a very lowsodium intake was met by increased adrenal cor-tical activity of a desoxycorticosterone-like hor-mone, accompanied by increase in activity of adre-nal protein catabolic hormone. Administration oflarge quantities of sodium abolished this need toprevent renal loss of sodium with resultant de-crease in adrenal cortical activity. Our findingsindicate that in the normal subject, the conserva-
1079
ALEXANDERLEAF AND WILLIAM T. COUTER
tion of body sodium by variation of renal excre-tion of sodium is the result of increased adrenalcortical activity in response to low sodium intakeand decreased adrenal cortical activity when largeamounts of sodium are administered.
ACKNOWLEDGMENT
The authors wish to acknowledge gratefully their ap-preciation to Dr. L. H. Newburgh whose interest andwise guidance made this study possible.
BIBLIOGRAPHY
1. Harrison, H. E., and Darrow, D. C., Renal functionin experimental adrenal insufficiency. Am. J.Physiol., 1939, 125, 631.
2. Loeb, R. F., Atchley, D. W., Benedict, E. M., andLeland, J., Electrolyte balance studies in adrenal-ectomized dogs with particular reference to theexcretion of sodium. J. Exper. Med., 1933, 57, 775.
3. Loeb, R. F., Effect of sodium chloride in treatmentof patient with Addison's disease. Proc. Soc.Exper. Biol. & Med., 1933, 30,, 808.
4. Harrop, G. A., Weinstein, A., Soffer, L. J., andTrescher, J. H., Diagnosis and treatment of Addi-son's disease. J. A. M. A., 1933, 100, 1850.
5. Thorn, G. W., Garbutt, H. T., Hitchcock, F. A., andHartman, F. A., The effect of cortin on the so-dium, potassium, chloride, inorganic phosphorusand total nitrogen balance in normal subjects andin patients with Addison's disease. Endocrinology,1937, 21, 202.
6. Leaf, A., Couter, W. T., and Newburgh, L. H., Someeffects of variation of sodium intake in normalsubjects. J. Clin. Invest., 1949, 28, 1082.
7. Wintrobe, M. M., Clinical Hematology. Lea &Febiger, Philadelphia, 1942, p. 201.
8. Butler, A. M., and Tuthill, E., An application of theuranyl zinc acetate method for determination ofsodium in biological material. J. Biol. Chem.,1931, 93, 171.
9. Peters, J. P., and Van Slyke, D. D., QuantitativeClinical Chemistry. Methods, Vol. II, p. 735.Williams & Wilkins Co., Baltimore, 1932.
10. Shohl, A. T., and Bennett, H. B., A micro methodfor the determination of potassium as iodoplatinate.J. Biol. Chem., 1928, 78, 643.
11. Van Slyke, D. D., The determination of chlorides inblood and tissues. J. Biol. Chem., 1923, 58, 523.
12. Harvey, S. C., The quantitative determination ofchlorides in the urine. Arch. Int. Med., 1910, 6, 12.
13. Peters, J. P., and Van Slyke, D.D., (Reference No.9 above), p. 550.
14. Ibid., p. 516.15. Ibid., p. 602.16. Bonsnes, R. W., and Taussky, H. H., On colori-
metric determination of creatinine by Jaffe reac-tion. J. Biol. Chem., 1945, 158, 581.
17. Gentzkow, C., Accurate method for determination ofblood urea nitrogen by direct nesslerization. J.Biol. Chem., 1942, 143, 531.
18. Barker, S. B., Direct colorimetric determination ofurea in blood and urine. J. Biol. Chem., 1944,152, 453.
19. Buchanan, 0. H., Block, W. D., and Christman, A.A., The metabolism of the methylated purines. I.The enzymatic determination of urinary uric acid.J. Biol. Chem., 1945, 157, 181.
20. Van Slyke, D. D., and Neill, J. M., The determina-tion of gases in blood and other solutions by vac-uum extraction and manometric measurement. J.Biol. Chem., 1924, 61, 523.
21. Peters, J. P., and Van Slyke, D. D., (Reference No.9 above), p. 530.
22. Fiske, C. H., and Subbarow, Y., The colorimetricdetermination of phosphorus. J. Biol. Chem., 1925,66, 375.
23. Goldring, W., and Chasis, H., Hypertension andHypertensive Disease. Commonwealth Fund, NewYork, 1944, p. 198.
24. Bowes, A. deP., and Church, C. F., Food Values ofPortions Commonly Used. A. deP. Bowes, Phila-delphia, 1946.
25. McCance, R. A., and Widdowson, E. M., The Chem-ical Composition of Foods. Chemical PublishingCo., Inc., New York, 1947.
26. Barrett, E., and Addis, T., The serum creatinineconcentration of normal individuals. J. Clin.Invest., 1947, 26, 875.
27. McCance, R. A., Experimental sodium chloride de-ficiency in man. Proc. Roy. Soc., Series B, 1936,119, 245.
28. McCance, R. A., and Widdowson, E. M., The secre-tion of urine in man during experimental saltdeficiency. J. Physiol., 1937, 91, 222.
29. McCance, R. A., The excretion of urea, salts andwater during periods of hydropaenia in man. J.Physiol., 1945, 104, 196.
30. Albright, F., and Bauer, W., The action of sodiumchloride, ammonium chloride and sodium bicar-bonate on the total acid-base balance of a case ofchronic nephritis with edema. J. Clin. Invest.,1929, 7, 465.
31. Landis, E. M., Elsom, K. A., Bott, P. A., and Shiels,E., Observations on sodium chloride restriction andurea clearance in renal insufficiency. J. Clin.Invest., 1935, 14, 525.
32. Steinitz, K., and Turkand, H., The determination ofthe glomerular filtration rate by the endogenouscreatinine clearance. J. Clin. Invest., 1940, 19, 285.
33. Brod, J., and Sirota, J. H., The renal clearance ofendogenous "creatinine" in man. J. Clin. Invest.,1948, 27, 645.
34. Thorn, G. W., Engel, L. L., and Eisenberg, H., Ef-fects of corticosterone and related compounds onrenal excretion of electrolytes. J. Exper. Med.,1938, 68, 161.
1080
RENAL SODIUM EXCRETION IN NORMALSUBJECTS
35. Kendall, E. C., Hormones of the adrenal cortex.Endocrinology, 1942, 30, 853.
36. Long, C. N. H., A discussion of the mechanism ofaction of adrenal cortical hormones on carbohy-drate and protein metabolism. Endocrinology,1942, 30, 870.
37. Thorn, G. W., Howard, R. P., and Emerson, K., Jr.,Treatment of Addison's disease with desoxycorti-costerone acetate, a synthetic adrenal cortical hor-mone (preliminary report). J. Clin. Invest., 1939,18, 449.
38. Thorn, G. W., Desoxycorticosterone. J. Mt. SinaiHospital, 1942, 8, 1177.
39. Wilder, R. M., Kendall, E. C., Snell, A. M., Kepler,E. J., Rynearson, E. H., and Adams, M., Intakeof potassium, an important consideration in Addi-son's disease. Arch. Int. Med., 1937, 59, 367.
40. Conn, J. W., The mechanism of acclimatization toheat, in: Advances in Internal Medicine. Inter-science Publishers, Inc., New York, 1949.
41. Swanson, P. P., and Smith, A. H., Effect of restric-tion of inorganic salts in the diet on organ growth.Am. J. Physiol., 1936, 116, 516.
42. Forsham, P. H., Thorn, G. W., Bergner, G. E., andEmerson, K., Jr., Metabolic changes induced bysynthetic 11-dehydrocorticosterone acetate. Am. J.Med., 1946, 1, 105.
43. Forsham, P. H., Thorn, G. W., Prunty, F. T. G., andHills, A. G., Clinical studies with pituitary adreno-corticotrophin. J. Clin. Endocrinol., 1948, 8, 15.
44. Selye, H., The general adaptation syndrome and thediseases of adaptation. J. Clin. Endocrinol., 1946,6, 117.