ORIGINAL PAPER Evolution of the fast start response in the cavefish Astyanax mexicanus Kim Hoke & Adina Schwartz & Daphne Soares Received: 15 May 2012 / Revised: 28 May 2012 / Accepted: 30 May 2012 / Published online: 28 June 2012 # Springer-Verlag 2012 Abstract The extreme sensory environment of subterranean caves presents an adaptive challenge for troglobitic organisms. Although caves provide distinct selective pressures (e.g., dark- ness, food scarcity), advantages often come with living in isolated places. The cave-adapted (hypogean) form of the tele- ost Astyanax mexicanus lacks predators in its natural environ- ment, unlike its ancestral surface (epigean) form that inhabits the nearby rivers. This species provides a unique opportunity for comparative studies because A. mexicanus has invaded the cave environment multiple times, providing us with indepen- dent comparative adaptive experiments. Here, we examine kinematic parameters that characterize the escape responses of various forms of A. mexicanus. We demonstrate that popula- tions differ in details of fast start performance, but that surface fish do not perform better than cavefish in escape responses. Our results are more consistent with genetic drift or divergent selection among caves than with relaxation of selection in determining the evolution of escape behavior in this system. Keywords Cavefish . Fast start . Escape response . Relaxed selection . Genetic drift Introduction Organisms occasionally invade novel environments in which they experience substantial changes in selective pressures compared to those that have shaped their ancestors. A general prediction is that a relaxation of selection in a novel environ- ment will enable the degradation or loss of structures or functions shaped by those selective pressures in the ancestral environment. Nonetheless, traits may persist paradoxically over long periods of time either due to secondary functions, pleiotropy, low cost, or lack of genetic variation (reviewed in Coss 1999; Lahti et al. 2009). Caves are fruitful environments for the study of ecology, evolution, and speciation (Poulson and White 1969; Culver 1982; Christman and Culver 2001; Christman et al. 2005; Niemiller et al. 2008) because organ- isms have independently invaded caves and evolved with a common set of environmental conditions (lack of light, re- stricted food sources, and predation pressure; Howarth 1980). Here, we examine whether an anti-predator behavior has degraded or persisted after the Mexican cavefish Astyanax mexicanus invaded caves with reduced or absent predation pressure. One common strategy by which fishes avoid predation is to quickly evade an imminent attack (Webb 1977, 1978; Walker et al. 2005; Fuiman et al. 2006). The fast start escape response is an anti-predator strategy in which performance differs among fishes due to both morphological and behav- ioral variation (Webb 1978, 1984; Andraso 1997; Hale et al. 2002). This behavior, a rapid burst of movement away from a threatening stimulus, is highly stereotyped within a species (Hale 1999; Hale et al. 2002). When a fish senses a preda- tory attack, it responds by initially making a very fast (∼10 ms) simultaneous contraction to the muscles of one side of the body, bending the fish into a “C” shape (stage 1), away from the danger. A secondary stage follows (stage 2) in which there is a contraction of the opposite musculature to allow the fish to generate a tail beat and swim away (for a review, see Eaton and Hackett 1984; Domenici and Blake 1997). The fast start response is an effective means of avoiding a predator ’ s attack, and variation in fast start Communicated by T. Bakker K. Hoke Department of Biology, Colorado State University, Fort Collins, CO 80523, USA A. Schwartz : D. Soares (*) Biology Department, University of Maryland, College Park, MD 20742, USA e-mail: [email protected]Behav Ecol Sociobiol (2012) 66:1157–1164 DOI 10.1007/s00265-012-1368-6
Transcript
ORIGINAL PAPER
Evolution of the fast start response in the cavefish Astyanaxmexicanus
Kim Hoke & Adina Schwartz & Daphne Soares
Received: 15 May 2012 /Revised: 28 May 2012 /Accepted: 30 May 2012 /Published online: 28 June 2012# Springer-Verlag 2012
Abstract The extreme sensory environment of subterraneancaves presents an adaptive challenge for troglobitic organisms.Although caves provide distinct selective pressures (e.g., dark-ness, food scarcity), advantages often come with living inisolated places. The cave-adapted (hypogean) form of the tele-ost Astyanax mexicanus lacks predators in its natural environ-ment, unlike its ancestral surface (epigean) form that inhabitsthe nearby rivers. This species provides a unique opportunityfor comparative studies because A. mexicanus has invaded thecave environment multiple times, providing us with indepen-dent comparative adaptive experiments. Here, we examinekinematic parameters that characterize the escape responses ofvarious forms of A. mexicanus. We demonstrate that popula-tions differ in details of fast start performance, but that surfacefish do not perform better than cavefish in escape responses.Our results are more consistent with genetic drift or divergentselection among caves than with relaxation of selection indetermining the evolution of escape behavior in this system.
Organisms occasionally invade novel environments in whichthey experience substantial changes in selective pressures
compared to those that have shaped their ancestors. A generalprediction is that a relaxation of selection in a novel environ-ment will enable the degradation or loss of structures orfunctions shaped by those selective pressures in the ancestralenvironment. Nonetheless, traits may persist paradoxicallyover long periods of time either due to secondary functions,pleiotropy, low cost, or lack of genetic variation (reviewed inCoss 1999; Lahti et al. 2009). Caves are fruitful environmentsfor the study of ecology, evolution, and speciation (Poulsonand White 1969; Culver 1982; Christman and Culver 2001;Christman et al. 2005; Niemiller et al. 2008) because organ-isms have independently invaded caves and evolved with acommon set of environmental conditions (lack of light, re-stricted food sources, and predation pressure; Howarth 1980).Here, we examine whether an anti-predator behavior hasdegraded or persisted after the Mexican cavefish Astyanaxmexicanus invaded caves with reduced or absent predationpressure.
One common strategy by which fishes avoid predation isto quickly evade an imminent attack (Webb 1977, 1978;Walker et al. 2005; Fuiman et al. 2006). The fast start escaperesponse is an anti-predator strategy in which performancediffers among fishes due to both morphological and behav-ioral variation (Webb 1978, 1984; Andraso 1997; Hale et al.2002). This behavior, a rapid burst of movement away froma threatening stimulus, is highly stereotyped within a species(Hale 1999; Hale et al. 2002). When a fish senses a preda-tory attack, it responds by initially making a very fast(∼10 ms) simultaneous contraction to the muscles of oneside of the body, bending the fish into a “C” shape (stage 1),away from the danger. A secondary stage follows (stage 2)in which there is a contraction of the opposite musculatureto allow the fish to generate a tail beat and swim away (for areview, see Eaton and Hackett 1984; Domenici and Blake1997). The fast start response is an effective means ofavoiding a predator’s attack, and variation in fast start
Communicated by T. Bakker
K. HokeDepartment of Biology, Colorado State University,Fort Collins, CO 80523, USA
A. Schwartz :D. Soares (*)Biology Department, University of Maryland,College Park, MD 20742, USAe-mail: [email protected]
performance predicts survival in staged predator encounters(Walker et al. 2005; Langerhans 2009; O’Steen et al. 2010).Furthermore, fast start performance varies within species in amanner consistent with relaxation of selection by reducedpredation pressure; Trinidadian guppies and mosquito fish bothhave reduced fast start performance in populations with lowerabundance of predators (O’Steen et al. 2002; Ghalambor et al.2004; Langerhans 2009).
The fast start response behavior of fishes is mediated by asimple and well-described neural circuit (e.g., goldfish: Zottoliand Faber 1980; Faber et al. 1989; zebrafish: Kimmel et al.1982; Eaton 2001; puffer fish: Greenwood et al. 2010; weaklyelectric fish: Trujillo-Cenóz and Bertolotto 1990; Borde et al.1991). Sensory inputs from auditory, mechanosensory, tactile,and visual brain regions innervate a series of reticulospinalneurons, of which the most prominent are the Mauthner cells(M cells). This innervation occurs via mixed electrical andchemical synapses. These M cells, along with other reticulo-spinal cells, innervate muscles on the contralateral body mus-culature and inhibit each other. A single action potential in theM cell can initiate the escape response (Eaton and Farley1975; Zottoli 1977; Eaton et al. 1981). The response directionis often directed away from the stimulus if the fish is in freewater with an unobstructed escape path to both sides (Eatonand Emberley 1991).
Here, we examine how the cave adaptation has changedthe direction, angle, latency, and distance traveled of theescape responses in various forms of the Mexican tetra A.mexicanus. In A. mexicanus, we find a particularly goodwindow into the adaptation of behavior. This fish is under-going allopatric speciation and is extant in two major diver-gent forms: one river-dwelling form and various cave-dwelling forms (Fig. 1). Cavefish derived from river-dwelling ancestors, with different cavefish populations be-lieved to have originated in parallel (Dowling et al. 2002;
Strecker et al. 2003). Cave A. mexicanus lack functionaleyes as adults (Jeffery and Martasian 1998; Jeffery et al.2000; Jeffery 2009) and have lost the ancestral silver pig-mentation (McCauley et al. 2004; Protas et al. 2007), buthave more taste buds (Jeffery 2001) and are attracted tomechanosensory stimulation (Yoshizawa et al. 2010). Nopredators have been reported in caves harboring Astyanax,and cave A. mexicanus are likely at the top of the food chain(Niemiller and Poulson 2010; Poulson 2010).
The consistency of morphological and behavioralchanges across multiple evolutionary transitions in A. mex-icanus suggests that these parallel changes are consequencesof common selective pressures across cave environments,such as lack of light and low food availability. To determinewhether the lack of predators consistently drives or enableschanges in anti-predator behaviors, we examined escaperesponses to test two main hypotheses: (1) that cavefishshow modified escape performance due to the relaxationfrom selection by predators and (2) that cavefish populationsretain escape responses but differ in the details of escaperesponses consistent with divergent environmental condi-tions in each cave or the process of genetic drift.
Methods
A total of 51 adult fishes from our facility and the colony ofDr. William Jeffery were used in this study. Animals werebred in captivity from wild stock for a minimum of fourgenerations under approved IACUC protocol. Cavefisheshave been named after the cave from which the populationswere collected: Molino, Pachón, Tinaja, Jineo, Guerrero,Curva, Subterraneo and Los Sabinos. Surface fish popula-tions were collected from the nearby rivers. One cavefish,named Steinhardt, is the aquarium stores. We used surfacefish to represent the accepted ancestral form. Fish wereacclimated for 5 to 10 min in a custom-made arena at roomtemperature. To test fast start behavior, we used a vigoroustap to the side of the arena that triggered mechanosensoryand auditory information. The stimulus was attached to astring and a pivot, and raised to the same height for eachtrial to generate consistency in the impact, as in Greenwoodet al. (2010). The fast start responses of individuals wererecorded using high-speed videography (X-PRI camera; DelImaging, Cheshire, CT) at 500 frames per second usingAOS imaging light software (Del Imaging). Three to 11 fishper population were tested, with each individual stimulated20–25 times. We waited for a period of 3 to 5 min betweenstimulations. When the fish did not startle, we waited foranother acclimation period before continuing with the trial.We only quantified the responses in which the fish formed aC shape within 50 ms for the analyses presented here, for atotal of 279 trials.
Fig. 1 a Surface form of A. mexicanus and b cave form representativefrom the Pachón cave. Scale bar01 cm
1158 Behav Ecol Sociobiol (2012) 66:1157–1164
We digitized three points (head, midbody, and base oftail) for reference along the fish’s body using the imaginganalysis software Proanalyst (Xcitex, Boston, MA). Webased measurements for each fast start on the fish’s move-ments between the frame in which the stimulus struck andthe frame marking the end of the first tail beat, in which thetail returns to center (Fig. 2). In the frame showing thestimulus impact, we measured the stimulus distance as thedistance between the impact site and head, and the stimulusangle as the angle between lines connecting the fish headand midbody and connecting the fish midbody and stimulusposition. We measured the time interval between the stimu-lation and the onset of body curvature as the responselatency. We measured the angle between the two lines con-necting the midbody with the head and the midbody withthe tail in each frame, and report the greatest angle in eachfast start as the maximum turning angle. Proanalyst calcu-lated the speed of the head based on its distance movedbetween concurrent frames. We measured distance traveledby taking the integral of the speed of the head between theframe comprising the onset of body curvature and the finalframe of the first tail beat.
We tested for differences among populations in the laten-cy of response, maximum turning angle, and distance trav-eled during the first tail beat using mixed model procedures(Proc Mixed) in SAS (Cary, NC, USA). Parameters wereestimated by restricted maximum likelihood, and the stan-dard errors of the fixed effects as well as the degrees offreedom were corrected using the Kenward–Roger method.For all of our analyses, we included the individual fish as arandom effect so as to account for the repeated measures ofeach fish. All analyses tested for the fixed effects of popu-lation, and both the angle and distance between stimulus and
fish as covariates. We included two interaction terms, pop-ulation by stimulus angle and population by stimulus dis-tance, in the first round of analysis and compared the fullmodel with models in which nonsignificant interactionterms were omitted. In no cases did the reduced modelshave large differences in parameter estimates or leastsquares means estimates for population, so we present herethe reduced model retaining only those interaction termswith p<0.05.
We followed up significant main or interaction effects ofpopulation to test two hypotheses: (1) that cave populationsdiffered in a consistent manner from the surface populationand (2) that cave populations differed from each other. Weconducted F tests of these subhypotheses using the contraststatement in Proc Mixed of SAS and further explored differ-ences among cave populations by F tests that comparedeach population to the other eight.
In addition to the above analyses, we assessed whetherthe direction turned (toward or away from the stimulus)differed across populations. We used the generalized linearmodels procedure in SAS and assumed a binomial distribu-tion with a logit link function because the dependent vari-able had two categories. We included fixed effects ofpopulation of origin, the individual fish ID as a randomfactor, as well as two covariates: the distance between theanimal and stimulus, and the angle between the stimulus andthe fish. We included interaction terms of each covariatewith population for the first round of analysis and thencompared models that had significant interaction terms.We report here the models in which interaction terms withp<0.05 are omitted. We tested for significance of the fixedeffects using score statistics rather than Wald statistics dueto our small sample sizes.
Fig. 2 Diagram showing the measurements made for each fast startresponse. Three points mark the three locations (head, midbody, andbase of tail) used for measurements, with subsequent fish outlinesoverlaid in the main panel. Ipsilateral responses were considered thosethat turned the fish toward the stimulus, while contralateral turned thefish away. The insert shows the trajectory from the initiation of the fast
start (0), to the maximum turning angle (1) to the end of the first tailbeat (2). θs angle of stimulus (stim) in relationship to the body of thefish, measured at time 0, d distance from the head to the stimulus attime 0, t distanced traveled by the head throughout the first tail beat,from time 0 to 2, θr maximum turning angle of fish during theresponse, measured at time 1
Behav Ecol Sociobiol (2012) 66:1157–1164 1159
Results
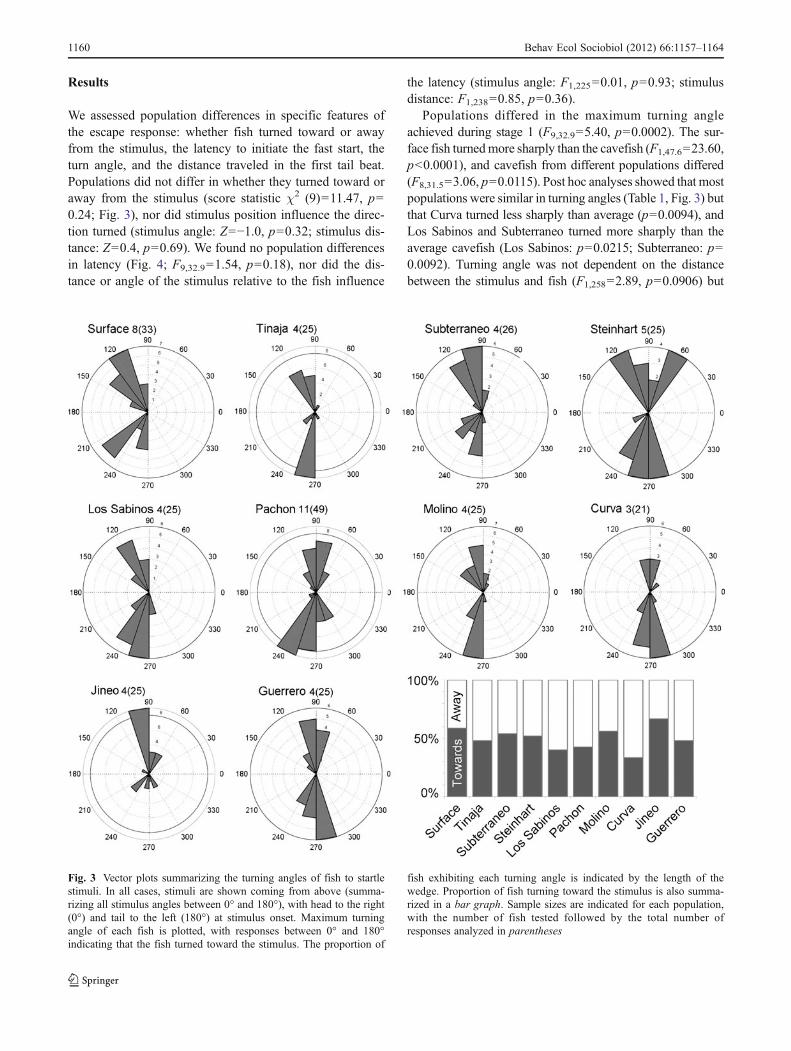
We assessed population differences in specific features ofthe escape response: whether fish turned toward or awayfrom the stimulus, the latency to initiate the fast start, theturn angle, and the distance traveled in the first tail beat.Populations did not differ in whether they turned toward oraway from the stimulus (score statistic χ2 (9)011.47, p00.24; Fig. 3), nor did stimulus position influence the direc-tion turned (stimulus angle: Z0−1.0, p00.32; stimulus dis-tance: Z00.4, p00.69). We found no population differencesin latency (Fig. 4; F9,32.901.54, p00.18), nor did the dis-tance or angle of the stimulus relative to the fish influence
the latency (stimulus angle: F1,22500.01, p00.93; stimulusdistance: F1,23800.85, p00.36).
Populations differed in the maximum turning angleachieved during stage 1 (F9,32.905.40, p00.0002). The sur-face fish turnedmore sharply than the cavefish (F1,47.6023.60,p<0.0001), and cavefish from different populations differed(F8,31.503.06, p00.0115). Post hoc analyses showed that mostpopulations were similar in turning angles (Table 1, Fig. 3) butthat Curva turned less sharply than average (p00.0094), andLos Sabinos and Subterraneo turned more sharply than theaverage cavefish (Los Sabinos: p00.0215; Subterraneo: p00.0092). Turning angle was not dependent on the distancebetween the stimulus and fish (F1,25802.89, p00.0906) but
Fig. 3 Vector plots summarizing the turning angles of fish to startlestimuli. In all cases, stimuli are shown coming from above (summa-rizing all stimulus angles between 0° and 180°), with head to the right(0°) and tail to the left (180°) at stimulus onset. Maximum turningangle of each fish is plotted, with responses between 0° and 180°indicating that the fish turned toward the stimulus. The proportion of
fish exhibiting each turning angle is indicated by the length of thewedge. Proportion of fish turning toward the stimulus is also summa-rized in a bar graph. Sample sizes are indicated for each population,with the number of fish tested followed by the total number ofresponses analyzed in parentheses
1160 Behav Ecol Sociobiol (2012) 66:1157–1164
was associated negatively with stimulus angle (F1,24305.35,p00.0215).
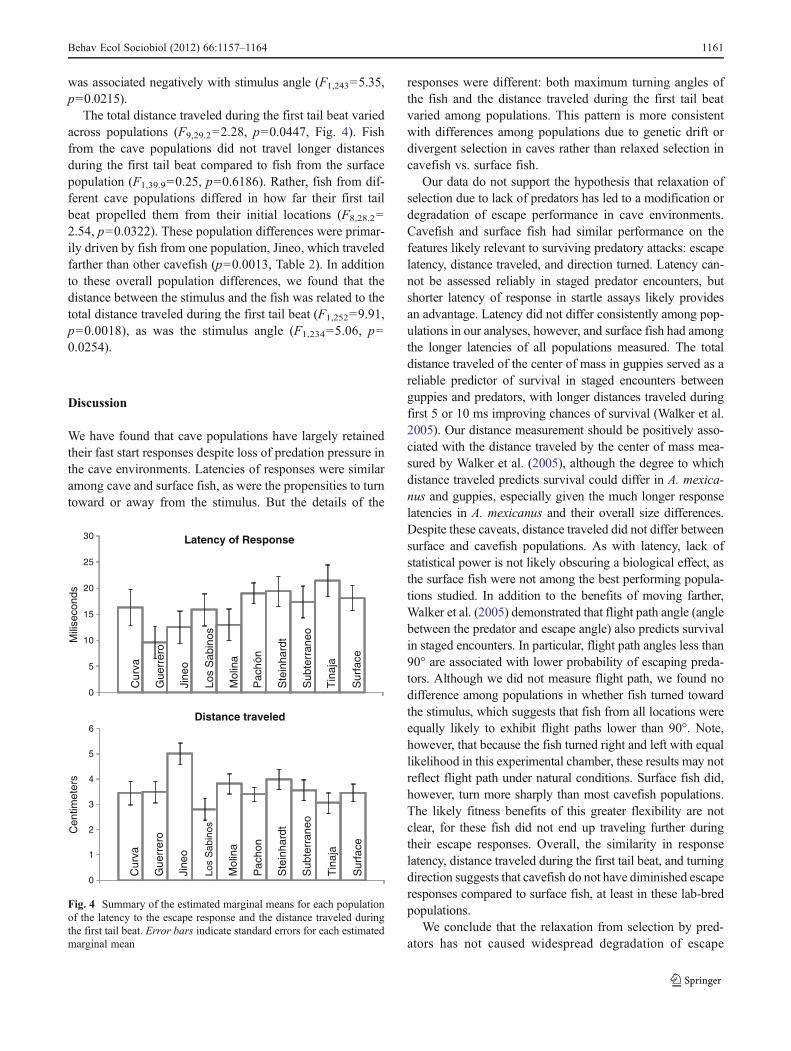
The total distance traveled during the first tail beat variedacross populations (F9,29.202.28, p00.0447, Fig. 4). Fishfrom the cave populations did not travel longer distancesduring the first tail beat compared to fish from the surfacepopulation (F1,39.900.25, p00.6186). Rather, fish from dif-ferent cave populations differed in how far their first tailbeat propelled them from their initial locations (F8,28.20
2.54, p00.0322). These population differences were primar-ily driven by fish from one population, Jineo, which traveledfarther than other cavefish (p00.0013, Table 2). In additionto these overall population differences, we found that thedistance between the stimulus and the fish was related to thetotal distance traveled during the first tail beat (F1,25209.91,p00.0018), as was the stimulus angle (F1,23405.06, p00.0254).
Discussion
We have found that cave populations have largely retainedtheir fast start responses despite loss of predation pressure inthe cave environments. Latencies of responses were similaramong cave and surface fish, as were the propensities to turntoward or away from the stimulus. But the details of the
responses were different: both maximum turning angles ofthe fish and the distance traveled during the first tail beatvaried among populations. This pattern is more consistentwith differences among populations due to genetic drift ordivergent selection in caves rather than relaxed selection incavefish vs. surface fish.
Our data do not support the hypothesis that relaxation ofselection due to lack of predators has led to a modification ordegradation of escape performance in cave environments.Cavefish and surface fish had similar performance on thefeatures likely relevant to surviving predatory attacks: escapelatency, distance traveled, and direction turned. Latency can-not be assessed reliably in staged predator encounters, butshorter latency of response in startle assays likely providesan advantage. Latency did not differ consistently among pop-ulations in our analyses, however, and surface fish had amongthe longer latencies of all populations measured. The totaldistance traveled of the center of mass in guppies served as areliable predictor of survival in staged encounters betweenguppies and predators, with longer distances traveled duringfirst 5 or 10 ms improving chances of survival (Walker et al.2005). Our distance measurement should be positively asso-ciated with the distance traveled by the center of mass mea-sured by Walker et al. (2005), although the degree to whichdistance traveled predicts survival could differ in A. mexica-nus and guppies, especially given the much longer responselatencies in A. mexicanus and their overall size differences.Despite these caveats, distance traveled did not differ betweensurface and cavefish populations. As with latency, lack ofstatistical power is not likely obscuring a biological effect, asthe surface fish were not among the best performing popula-tions studied. In addition to the benefits of moving farther,Walker et al. (2005) demonstrated that flight path angle (anglebetween the predator and escape angle) also predicts survivalin staged encounters. In particular, flight path angles less than90° are associated with lower probability of escaping preda-tors. Although we did not measure flight path, we found nodifference among populations in whether fish turned towardthe stimulus, which suggests that fish from all locations wereequally likely to exhibit flight paths lower than 90°. Note,however, that because the fish turned right and left with equallikelihood in this experimental chamber, these results may notreflect flight path under natural conditions. Surface fish did,however, turn more sharply than most cavefish populations.The likely fitness benefits of this greater flexibility are notclear, for these fish did not end up traveling further duringtheir escape responses. Overall, the similarity in responselatency, distance traveled during the first tail beat, and turningdirection suggests that cavefish do not have diminished escaperesponses compared to surface fish, at least in these lab-bredpopulations.
We conclude that the relaxation from selection by pred-ators has not caused widespread degradation of escape
0
5
10
15
20
25
30
Gue
rrer
o
Cur
va
Jine
o
Los
Sab
inos
Mol
ina on
Pac
h
Ste
inha
rdt
Tin
aja
Sub
terr
aneo
Sur
face
Latency of Response
0
1
2
3
4
5
6
Gue
rrer
o
Cur
va
Jine
o
Los
Sab
inos
Mol
ina
Pac
hon
Ste
inha
rdt
Tin
aja
Sub
terr
aneo
Sur
face
Distance traveled
Mili
seco
nds
Cen
timet
ers
Fig. 4 Summary of the estimated marginal means for each populationof the latency to the escape response and the distance traveled duringthe first tail beat. Error bars indicate standard errors for each estimatedmarginal mean
Behav Ecol Sociobiol (2012) 66:1157–1164 1161
responses in cave populations. This finding is consistentwith theory that behavior is more likely to persist underrelaxed selection than are morphological traits both due toits inherent plasticity and because of extensive pleiotropy(Lahti et al. 2009). Possible reasons for maintaining theescape response in cavefish are that false escapes incurlow costs or that the circuitry underlying fast starts couldbe important for other behaviors. If fish rarely produce faststarts in their natural environments, there would be minimalcost and thus little selective advantage to lose the escaperesponses. In addition, the Mauthner cell circuitry that ini-tiates the fast start response could be used in other behaviorssuch as prey capture (Canfield and Rose 1993), socialbehaviors such as courtship or aggression, or juvenile es-cape responses from conspecifics or other cave-dwellingspecies. Alternatively, the fast start circuitry could be linkeddevelopmentally to other hindbrain functions, such thatevolved loss of the Mauthner neuron would have pleiotropiceffects on other behaviors. Additional studies of hindbraindevelopment and the natural history of cavefish could helpexplain the maintenance of escape responses despite lackingpredators.
Because release from predation pressure does not explainpopulation differences in escape performance, we suggestthat phylogeny, genetic drift, and divergent selective pres-sures contribute to population differences in distance
traveled and turning angle. Unfortunately, the Astyanaxphylogeny has not been fully resolved, and existing phylog-enies (Dowling et al. 2002; Strecker et al. 2003; Ornelas-García et al. 2008) do not include all populations in ourstudy; thus, we do not analyze our data formally to test forinfluences of phylogenetic signal in escape performancetraits. Dowling et al. (2002) suggested a minimum of twodistinct lineages of Astyanax cavefishes, one lineage includ-ing Pachón and Subterraneo and closely related surfacefishes and another consisting of Los Sabinos, Tinaja, andCurva. Strecker et al. (2003) have basically concordantresults in this subset of populations. Our turning angleresults are not consistent with phylogeny driving currentpatterns of fast start variation. Fish from Subterraneo andLos Sabinos, populations in two separate lineages, had thelargest turning angles, whereas Curva had the lowest turningangle despite its relatedness to Los Sabinos. One possibleinfluence on turning angle is vertebral number (Breinerd andPatek 1998), but variation in intracranial vertebrae from asubset of these populations matches phylogenetic related-ness (Dowling et al. 2002), not our patterns of turning angle.The existing phylogenies are not informative to assess thedistance-traveled results, as Jineo is most distinct and hasnot been assessed in any published phylogenies. Thus, wefind little evidence for a strong signature of relatedness inshaping population differences in escape behavior; however,
Table 1 Contrasts comparingturning angles of fish from eachcave population to those of othercavefish
DF degrees of freedom
Population Numerator DF Denominator DF F value p value
Curva 1 27.3 7.81 0.0094
Guerreo 1 30.8 1.02 0.3205
Jineo 1 31.6 1.48 0.2323
Los Sabinos 1 31.4 5.86 0.0215
Molina 1 30.6 0.11 0.7380
Pachón 1 40.7 1.34 0.2534
Steinhardt 1 34.3 0.44 0.5119
Subterraneo 1 29.3 7.77 0.0092
Tinaja 1 30.2 1.74 0.1967
Table 2 Contrasts comparingdistances traveled during thefirst tail beat of fish from eachcave population to those of othercavefish
DF degrees of freedom
Population Numerator DF Denominator DF F value p value
Curva 1 25.1 0.14 0.7077
Guerreo 1 27.4 0.02 0.9018
Jineo 1 28.1 12.79 0.0013
Los Sabinos 1 27.6 5.40 0.0277
Molina 1 27.1 0.43 0.5187
Pachón 1 34.7 0.54 0.4667
Steinhardt 1 31.0 0.93 0.3434
Subterraneo 1 26.4 0.03 0.8581
Tinaja 1 27.5 2.59 0.1192
1162 Behav Ecol Sociobiol (2012) 66:1157–1164
we lack the fully resolved phylogeny required for testingthis hypothesis.
Cave populations have low genetic diversity, suggestingthat bottlenecks may have occurred and therefore that ge-netic drift could be a dominant evolutionary force (Panaramand Borowsky 2005). On the other hand, we lack detailedenvironmental data on these caves to propose alternativescenarios of divergent selective pressures that could accountfor the patterns of fast start responses. Thus, the patterns ofvariation in escape kinematics could be due to either geneticdrift or to pleiotropic effects of selective pressures that haveshaped other features of fish morphology, physiology, orbehavior. The cavefish from these populations differ instandard length and shape, but those patterns of variationin body morphology do not correspond to the differences inescape kinematics we describe (unpublished data). Futurestudies comparing escape performance and other factors thatinfluence performance kinematics, such as fin shape, bodystiffness, musculoskeletal system, and physiology, couldidentify which morphological or physiological features areresponsible for these population differences in the details ofthe fast start behavior.
Acknowledgments We would like to thank Dr. William Jeffery forsharing his animal colony and Amy Parkhurst for her help. We alsoacknowledge the helpful comments of two anonymous reviewers.
Ethical standards All experiments comply with the current laws ofthe USA.
Conflict of interest The authors declare that they have no conflict ofinterest.
References
Andraso G (1997) A comparison of startle response in two morphs ofthe brook stickleback (Culaea inconstans): further evidence for atrade-off between defensive morphology and swimming ability.Evol Ecol 11:83–90
Borde M, Pereda A, Morales F (1991) Electrophysiological character-istics of the Mauthner cell of the weakly electric fish Gymnotuscarapo. Brain Res 567:145–148
Breinerd EL, Patek SN (1998) Vertebral column morphology, C-startcurvature, and the evolution of mechanical defenses in tetradonti-form fishes. Copeia 1998:971–984
Canfield JG, Rose GJ (1993) Activation of Mauthner neurons duringprey capture. J Comp Physiol A 172:611–618
Christman MC, Culver DC (2001) The relationship between cavebiodiversity and available habitat. J Biogeogr 28:367–380
Christman MC, Culver DC, Madden MK, White D (2005) Patterns ofendemism of the eastern North American cave fauna. J Biogeogr32:1441–1452
Coss RG (1999) Effects of relaxed natural selection on the evolution ofbehavior. In: Foster SA, Endler JA (eds) Geographic variation inbehavior: perspectives on evolutionary mechanisms. Oxford Uni-versity Press, Oxford, pp 180–208
Culver D (1982) Cave life: evolution and ecology. Harvard UniversityPress, Cambridge
Domenici P, Blake RW (1997) The kinematics and performance of fishfast-start swimming. J Exp Biol 200:1165–1178
Dowling TE, Martasian DP, Jeffery WR (2002) Evidence for multiplegenetic forms with similar eyeless phenotypes in the blind cave-fish, Astyanax mexicanus. Mol Biol Evol 19:446–455
Eaton RC (2001) The Mauthner cell and other identified neurons of thebrainstem escape network of fish. Prog Neurobiol 63:467–485
Eaton RC, Emberley DS (1991) How stimulus direction determines thetrajectory of the Mauthner-initiated escape response in a teleostfish. J Exp Biol 161:469–487
Eaton RC, Farley R (1975) Mauthner neuron field potential in newlyhatched larvae of the zebrafish. J Neurophysiol 38:502–512
Eaton RC, Hackett J (1984) The role of Mauthner cells in fast-startsinvolving escape in teleost fish. In: Eaton RC (ed) Neural mech-anisms of startle behavior. Plenum Press, New York, pp 213–266
Eaton RC, Lavender W, Wieland C (1981) Identification of Mauthner-initiated response patterns in goldfish: evidence from simultaneouscinematography and electrophysiology. J Comp Physiol 144:521–531
Faber DS, Fetcho JR, Korn H (1989) Neuronal networks underlyingthe escape response in goldfish. Ann NYAcad Sci 563:11–33
Fuiman L, Rose K, Cowanjr J, Smith E (2006) Survival skills requiredfor predator evasion by fish larvae and their relation to laboratorymeasures of performance. Anim Behav 71:1389–1399
Ghalambor CK, Reznick DN, Walker JA (2004) Constraints on adap-tive evolution: the functional trade-off between reproduction andfast-start swimming performance in the Trinidadian guppy (Poe-cilia reticulata). Am Nat 164:38–50
Greenwood AK, Peichel CL, Zottoli SJ (2010) Distinct startleresponses are associated with neuroanatomical differences in puf-ferfishes. J Exp Biol 213:613–620
Hale ME (1999) Locomotory mechanics during early life history:effects of size and ontogeny on fast start perfomance of salmonidfishes. J Exp Biol 202:1465–1479
Hale ME, Long JH Jr, McHenry MJ, Westneat MW (2002) Evolutionof behavior and neural control of the fast-start escape response.Evolution 56:993–1007
Howarth FG (1980) The zoogeography of specialized cave animals: abioclimatic model. Evolution 34:394–406
Jeffery WR (2001) Cavefish as a model system in evolutionary devel-opmental biology. Dev Biol 231:1–12
Jeffery WR, Martasian DP (1998) Evolution of eye regression in thecavefish Astyanax: apoptosis and the Pax-6 gene. Integr CompBiol 38:685–696
Jeffery WR, Strickler AG, Guiney S, Heyser DG, Tomarev SI (2000)Prox 1 in eye degeneration and sensory organ compensationduring development and evolution of the cavefish Astyanax.Dev Genes Evol 210:223–230
Kimmel CB, Powell SL, Metcalfe WK (1982) Brain neurons whichproject to the spinal cord in young larvae of the zebrafish. J CompNeurol 20:112–127
Lahti DC et al (2009) Relaxed selection in the wild. Trends Ecol Evol24:487–496
Langerhans RB (2009) Morphology, performance, fitness: functionalinsight into a post-Pleistocene radiation of mosquitofish. Biol Lett5:488–491
McCauley DW, Hixon E, Jeffery WR (2004) Evolution of pigment cellregression in the cavefish Astyanax: a late step in melanogenesis.Evol Dev 6:209–218
Niemiller ML, Fitzpatrick BM, Miller BT (2008) Recent divergencewith gene flow in Tennessee cave salamanders (Plethodontidae:Gyrinophilus) inferred from gene genealogies. Mol Ecol17:2258–2275
Behav Ecol Sociobiol (2012) 66:1157–1164 1163
Niemiller ML, Poulson TL (2010) Studies of the Amblyopsidae: past,present, and future. In: Trajano E, Bichuette ME, Kapoor BG(eds) The biology of subterranean fishes. Science Publishers,Enfield, pp 169–280
Ornelas-García C, Domínguez-Domínguez O, Doadrio I (2008) Evo-lutionary history of the fish genus Astyanax Baird & Girard(1854) (Actinopterygii, Characidae) in Mesoamerica reveals mul-tiple morphological homoplasies. BMC Evol Biol 8:340
O’Steen S, Cullum AJ, Bennett AF (2002) Rapid evolution of escapeability in Trinidad guppies (Poecilia reticulata). Evolution56:776–784
O’Steen S, Eby SL, Bunce JA (2010) Dynamically honest displays:courtship locomotor performance indicates survival in guppies.Funct Ecol 24:1045–1053
Panaram K, Borowsky R (2005) Gene flow and genetic variability incave and surface populations of the Mexican Tetra, Astyanaxmexicanus (Teleostei: Characidae). Copeia 2005:409–416
Poulson TL (2010) Cavefish: retrospective and prospective. In: TrajanoE, Bichuette ME, Kapoor BG (eds) The biology of subterraneanfishes. Science Publishers, Enfield, pp 1–40
Poulson TL,WhiteW (1969) The cave environment. Science 165:971–981Protas M, Conrad M, Gross JB, Tabin C, Borowsky R (2007) Regres-
sive evolution in the Mexican cave tetra, Astyanax mexicanus.Curr Biol 17:452–454
Strecker U, Bernatchez L, Wilkens H (2003) Genetic divergence be-tween cave and surface populations of Astyanax in Mexico (Char-acidae, Teleostei). Mol Ecol 12:699–710
Trujillo-Cenóz O, Bertolotto C (1990) Mauthner cells in the medulla ofthe weakly electric fish Gymnotus carapo. Experientia 46:441–443
Walker JA, Ghalambor CK, Griset OL, McKenney D, Reznick DN(2005) Do faster starts increase the probability of evading preda-tors? Funct Ecol 19:808–815
Webb P (1977) Effects of median-fin amputation on fast-start perfor-mance of rainbow trout (Salmo gairdneri). J Exp Biol 68:123–135
Webb P (1978) Fast-start performance and body form in seven speciesof teleost fish. J Exp Biol 74:211–226
Webb P (1984) Form and function in fish swimming. Sci Am 251:72–82
Yoshizawa M, Gorički Š, Soares D, Jeffery WR (2010) Evolution of abehavioral shift mediated by superficial neuromasts helps cave-fish find food in darkness. Curr Biol 20:1631–1636
Zottoli SJ, Faber DS (1980) An identifiable class of statoacousticinterneurons with bilateral projections in the goldfish medulla.Neurosci 5:1287–1302
Zottoli SJ (1977) Correlation of the startle reflex and Mauthner cellauditory responses in unrestrained goldfish. J Exp Biol 66:243–254