Excretion of Flagellin by a Short-Flagella Mutant ofSalmonella typhimurium
TAKESHI IKEDA,'* RITSU KAMIYA,1 AND SHIGERU YAMAGUCHI2Institute of Molecular Biology, Faculty of Science, Nagoya University, Nagoya 464, Japan,1 and Department
of Biology, School of Education, Waseda University, Shinjuku-ku, Tokyo 160, Japan2
Received 21 June 1982/Accepted 9 October 1982
A nonmotile mutant of Salmonella typhimurium, SJW1254, has very shortflagella (<0.1 pm long) due to a mutation in the structural gene of flagellin (H2).When ammonium sulfate was added to the culture medium of SJW1254 grown tothe late-log phase, a large amount of protein precipitated. Gel electrophoresis andimmunodiffusion showed that more than 90% (wt/wt) of the precipitated proteinwas flagellin. The mutant flagellin appeared to be excreted in the monomericform, in an amount comparable to the amount in the flagellar filaments of wild-type bacteria. No such precipitate was obtained from the medium of wild-typebacteria. The mutant flagellin had the same apparent molecular weight (55,000)and isoelectric point (5.3) as the wild-type flagellin, but differed in mobility inpolyacrylamide gel electrophoresis under nondenaturing conditions. Moreover,the mutant flagellin did not polymerize in vitro under various conditions in whichwild-type flagellin polymerized. These results suggested that the mutant bacteriaexcreted flagellin because the flagellin polymerized poorly and therefore could notbe trapped at the tip of the flagellar filament. This short-flagella mutant should beuseful for studying the mechanism of flagellin transport.
The flagellar filaments of Salmonella typhi-murium are tubular structures with a diameter ofabout 20 nm and a variable length of up to 20p.m. They grow in vivo by the addition ofglobular protein subunits (flagellin) to the distalends of the filaments (7, 10, 17). Thus, during theprocess of flagellar elongation, the flagellinmolecules should be transported to the tip, pos-sibly through the central hole of the filament (4,15, 16, 19).Recently, Yamaguchi has isolated a mutant of
S. typhimurium which is nonmotile owing to itsextremely short flagella (<0.1 p.m in length).Genetic analysis has shown that this strain has asingle mutation in the flagellin structural gene(H2). Being interested in how the mutation in theflagellin gene leads to the production of shortflagella, we tried to examine the nature of themutant flagellin. In the course of this study, wefound that the mutant bacteria excreted a largeamount of defective flagellin into the culturemedium.
MATERIALS AND METHODS
Bacterial strains. The short-flagella mutant of S.typhimurium, SJW1254, is a nonmotile strain derivedby spontaneous mutation from a motile strain,SJW806, which produces flagella in an H2-monophasicmanner (8). The flagellar antigen type of these strainsis e, n, x. Both the mutant and the parent bacteria
aggregated when anti-e, n, x-flagellin antibody wasadded to the medium.
Preparation of flagellin. The flagellin of the parentstrain was prepared by the method of Asakura et al. (2,3). Briefly, the bacteria, grown in 1% polypeptone and1% yeast extract, were harvested in the late-log phase.Flagellar filaments were detached from the cell bodiesby shaking concentrated bacterial suspensions andpurified by differential centrifugation and polymeriza-tion-depolymerization cycling. Monomeric flagellinwas obtained by heating the flagellar solution at 65°Cfor 3 min.The flagellin of the short-flagella mutant was isolat-
ed in the following manner. The mutant bacteria,grown in M9 medium (13), were removed in the late-log phase by centrifugation at 10,000 x g for 30 min.Solid ammonium sulfate was dissolved in this mediumto 40% saturation (about 1.8 M) to precipitate theflagellin. After standing overnight at 4°C, this solutionwas centrifuged at 10,000 x g for 30 min, and the pellet(AS-precipitate) was dissolved in a solution containing0.15 M NaCl and 10 mM sodium phosphate buffer (pH7.0; standard buffer solution). After dialysis againststandard buffer and centrifugation at 100,000 x g for60 min to remove insoluble materials, the solution wassubjected to column chromatography with SephacrylS-200 superfine gel (Pharmacia Fine Chemicals, Pis-cataway, N.J.), and a peak fraction that eluted at aposition corresponding to that of monomeric flagellinwas collected as the final sample.
Isolation of short flagella. The mutant bacteria grownto late-log phase were centrifuged at 10,000 x g for 20min, and the pellet was suspended in an equal volumeof standard buffer solution. This suspension was
FIG. 1. Electron micrographs of detached flagellar filaments from the short-flagella mutant SJW1254 (a, b,and c) and the parent strain SJW806 (d) of S. typhimurium. The filaments were negatively stained with 1% uranylacetate. The sample in (c) was reacted with antibody raised against e, n, x-flagellin. The portion indicatedbetween arrows is the flagellar hook. Bar, 0.1 ,um.
blended in a homogenizer (Nihon Seiki, Co., Tokyo) atabout 15,000 rpm for 10 min. Cell debris was removedby centrifugation at 20,000 x g for 15 min. The shortflagella in the supernatant were pelleted at 100,000 x gfor 60 min, resuspended in standard buffer, and usedas the sample of mutant flagella.
Electrophoresis. Polyacrylamide gel electrophoresiswas carried out by the method of Davis (6), except thata slab-gel apparatus was used. Sodium dodecyl sulfate(SDS)-polyacrylamide gel electrophoresis was per-formed by the method of Weber and Osborn (20).Isoelectric focusing was carried out by the method ofO'Farrell (14), with the destaining method of Vester-berg et al. (18).Other methods. Anti-e, n, x-flagellin antibody was
obtained by injecting purified flagella from SJW806into a rabbit. Immunodiffusion was performed by themethod of Clausen (5). Protein concentrations weredetermined by the modified biuret method of Gornallet al. (9) or by the method of Lowry et al. (12). A JEM100C microscope (JEOL Co. Ltd., Tokyo) was usedfor electron microscopy.
RESULTSElectron microscopy of mutant flagella. The
mutant flagella attached to the cell body weredifficult to observe by electron microscopy be-cause they were very short. Thus, we observedthe flagella after detaching them by treatment of
cell suspensions with a blender (see above). Theflagella were fairly uniform in length, and almostall of the filament seemed to be composed of thebasal hook portion (Fig. la and b). When theseflagella reacted with anti-e, n, x-flagellin anti-body, a small portion at the tip of the filamentswas always labeled with the antibodies (Fig. lc).From these images, we estimated that flagellinconstituted only 10 to 20 nm of the filament,whereas the hook was about 70 nm long, as in anormal flagellum (Fig. ld). The filaments in Fig.1 probably were not fragments of longer fila-ments, because almost all of the flagella had thebasal hook.
Excretion of flagellin. Genetic studies haveshown that the short-flagella mutant has a pointmutation in the flagellin structural gene (H2)(Yamaguchi, unpublished data). We expectedthat mutant and normal bacteria would synthe-size equivalent amounts of flagellin. If this wastrue, the flagellin should be present either insideor outside the cell body. To examine whetherthe flagellin was excreted by the mutant, weused the method of ammonium sulfate precipita-tion, since we knew that Salmonella flagellin canbe precipitated by ammonium sulfate at 40%saturation (1).
FIG. 2. SDS-polyacrylamide gel electrophoresis ofthe AS-precipitate and flagellins. Lane 1, purifiedflagellin of SJW806; lane 2, the AS-precipitate fromthe culture medium of SJW1254; lane 3, the SJW1254flagellin sample purified as described in the text.
The mutant bacteria were grown in M9 medi-um and removed from the medium by centrifu-gation in late-log phase. When ammonium sul-fate was added to this bacteria-free medium to40% saturation, the medium became turbid, ow-ing to production of precipitate. In contrast,almost no precipitate was produced when theparent strain was cultured and treated as above.Figure 2 shows the SDS-polyacrylamide gelelectrophoresis pattern of the AS-precipitatefrom the medium of the mutant. There is anessentially single band with an apparent molecu-lar weight of 55,000, which is the same molecu-lar weight as that of the parent strain flagellin.From a densitometric scan of the pattern, weestimated that this band amounted to more than90% of the total protein precipitated in 40%-saturated ammonium sulfate. The AS-precipi-tate cross-reacted with anti-e, n, x-flagellin anti-body to form a single precipitin line (Fig. 3).From these results, we concluded that the short-flagella mutant excreted a large amount of flagel-lin into the medium.
Table 1 shows the amounts of protein ob-tained from the medium by various treatments.The amount of AS-precipitate from the mediumof the mutant was comparable to the amount of
parent bacteria flagella obtained by blendertreatment (Table 1, lines 1 and 4); the smalldifference between them could be explained bythe difference in the method used to obtainflagellin. The AS-precipitate from the medium inwhich the mutant was grown did not increasenoticeably when ammonium sulfate was saturat-ed to 100%. Hence, we concluded that almost allof the flagellin in the medium was recovered inthe AS-precipitate. The second line of Table 1shows that little precipitate was obtained whenthe medium of the mutant was centrifuged at100,000 x g for 120 min without the addition ofammonium sulfate. This result indicated that theflagellin in the medium of the mutant existed inmonomeric or oligomeric forms, since centrifu-gation of such an extent is sufficient to sedimentflagellar fragments as small as the basal hook. Incontrast to the mutant, only a minute amount ofAS-precipitate was obtained from the medium ofthe parent strain (Table 1, line 3).
Purification of mutant flagellin. The mutantflagellin precipitated with ammonium sulfatewas purified as described above. After the AS-precipitate was dissolved in and dialyzed againststandard buffer (see above), the flagellin becameunsedimentable by centrifugation at 100,000 x gfor 120 min. This is in good contrast to theparent strain flagellin, which polymerizes in thepresence of a high concentration (0.8 to 1.2 M)of ammonium sulfate and remains polymeric instandard buffer (1). As electron microscopy didnot show ordered polymeric structures in theinitial AS-precipitate, it is likely that the mutantflagellin did not polymerize into filaments in40%-saturated (1.8 M) ammonium sulfate solu-tion.
Figure 4 shows the elution profile of the gelfiltration at the last step of the purification. Onexamination by SDS-polyacrylamide gel electro-phoresis, the peak fraction was found to containflagellin as an almost single component. Hencewe kept this fraction as the final sample of the
FIG. 3. Immunodiffusion of the AS-precipitate andflagellin. Wells 1, 2, and 3 contained anti-e, n, x-flagellin antibody, purified SJW806 flagellin, and theAS-precipitate from the medium of SJW1254, respec-tively.
a Protein was precipitated by centrifugation at100,000 x g for 120 min. UC, Ultracentrifuge.
b Flagellar filaments were obtained by the blendertreatment used for the isolation of SJW1254 flagella(see text).
mutant flagellin. The purity of flagellin in thissample was estimated from the densitometricscan of the gel to be more than 98% (Fig. 2). Thefinal yield was 5 to 10 mg of flagellin per 1 liter ofculture; this value was compatible with the yieldof flagella purified from the parent strain by anordinary method (2). In Fig. 4, the positionwhere the mutant flagellin was eluted was thesame as that of the monomeric parent flagellin(data not shown). This result indicates that themutant flagellin assumed a monomeric formunder these conditions. Since we did not useprocedures tending to depolymerize flagellathroughout the purification steps, it is reason-able to conclude that the mutant flagellin wasexcreted in the monomeric form.
Characterization of mutant flagellin. As stated,the mutant flagellin had the same apparent mo-lecular weight as the parent flagellin on SDS-polyacrylamide gel electrophoresis. This resultis consistent with the genetic evidence that themutant carries a point mutation in the flagellin
loF
o.s
I
I1
.I:p
Au 1) D a Q 50 a ' a
FRACDON Ni?BERFIG. 4. Chromatography of the AS-precipitate on
Sephacryl S-200 superfine. The arrow indicates thevoid volume. The shaded area indicates the fractioncontaining the flagellin. OD280, Optical density at 280nm.
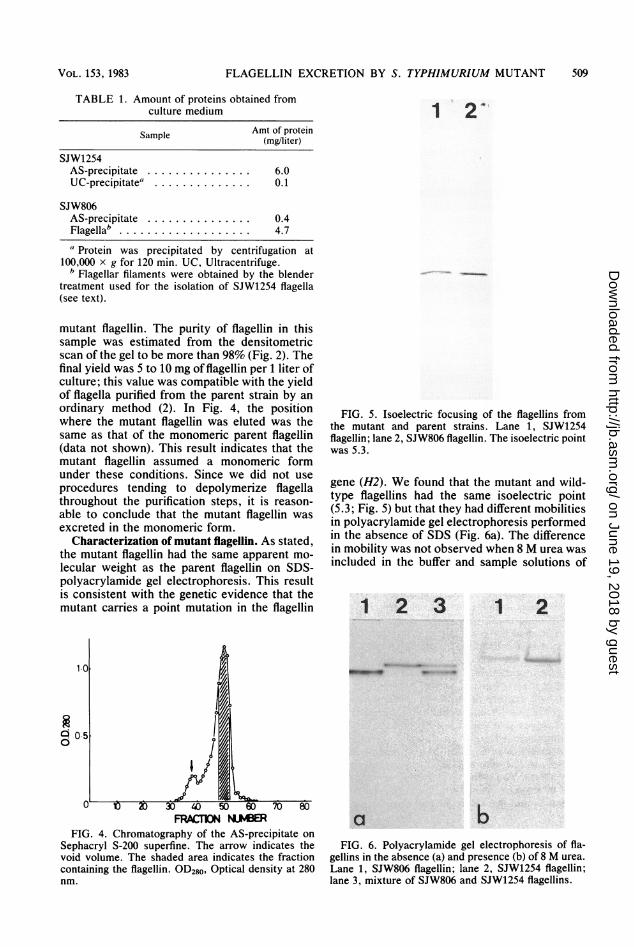
FIG. 5. Isoelectric focusing of the flagellins fromthe mutant and parent strains. Lane 1, SJW1254flagellin; lane 2, SJW806 flagellin. The isoelectric pointwas 5.3.
gene (H2). We found that the mutant and wild-type flagellins had the same isoelectric point(5.3; Fig. 5) but that they had different mobilitiesin polyacrylamide gel electrophoresis performedin the absence of SDS (Fig. 6a). The differencein mobility was not observed when 8 M urea wasincluded in the buffer and sample solutions of
1 2 3 1 2
a b
FIG. 6. Polyacrylamide gel electrophoresis of fla-gellins in the absence (a) and presence (b) of 8 M urea.Lane 1, SJW806 flagellin; lane 2, SJW1254 flagellin;lane 3, mixture of SJW806 and SJW1254 flagellins.
the electrophoresis (Fig. 6b). These results sug-gest that the mutant flagellin is different from thewild-type flagellin in tertiary structure, chargedistribution, or both.We tried to polymerize the mutant flagellin in
vitro under various conditions in which normalflagellin is polymerized, but these attempts wereunsuccessful. Instead, our preliminary studiesshowed that the mutant flagellin had an inhibi-tory effect on the polymerization of wild-typeflagellin: when the mutant flagellin was mixedwith the wild-type flagellin in a protein ratio of1:25, polymerization was almost completely in-hibited. Studies on the mechanisms of the inhibi-tion of polymerization are now in progress andwill be reported elsewhere.
DISCUSSIONWe showed above that a short-flagella mutant
of Salmonella excreted monomeric flagellin intothe medium. Of all the proteins present in themedium, the excreted flagellin was present in byfar the largest amount. Since the density of thebacteria at the final stage of growth in theseexperiments was about 109 cells per ml, theresults in Table 1 suggest that the amount offlagellin obtained from one cell should be about6 x 10-15 g in the mutant and 5 x 10-15 g in theparent bacteria. A 1-pum flagellar filament isconstructed of 2,200 flagellin molecules, eachwith a molecular weight of 55,000. Thus, theflagellin excreted by a mutant cell could make a30-p.m-long flagellar filament, or three 10-,um-long filaments. From these considerations, weconclude that flagellin is produced in a normalamount in the mutant bacteria and that most ofthe flagellin is excreted.Our preliminary studies suggested that the
mutant flagellin polymerized very poorly. It iseasy to understand why such a defective flagel-lin is excreted if we assume that the transporta-tion system for flagellin is working normally inthe mutant. By analogy, we may expect thatsome fla mutants that have defects in flagellarmorphogenesis, but not in the synthesis of fla-gellin, will excrete flagellin. In agreement withthis prediction, Yamaguchi (unpublished data)has observed that flagellin is excreted in certainfla mutants that have been shown to be normalin the synthesis of flagellin (11). Those mutants,as well as the short-flagella mutant reportedhere, should be useful in clarifying the mecha-nism of flagellin transportation, especially theenergetics and the exact path of flagellin secre-tion.
ACKNOWLEDGMENTS
We thank S. Asakura for helpful advice and encouragementthroughout the present study, K. Owaribe for help in prepara-
tion of the antiserum, and E. Shioi for excellent technicalassistance.This study was supported by a grant from the Ministry of
Education, Science and Culture of Japan.
LITERATURE CITED
1. Ada, G. L., J. V. Nossal, J. Pye, and A. Abbot. 1963.Behaviour of active bacterial antigens during the induc-tion of the immune response. (1) Properties of flagellarantigens from Salmonella. Nature (London) 199:1257-1259.
2. Asakura, S., G. Eguchi, and T. Iino. 1964. Reconstitutionof bacterial flagella in vitro. J. Mol. Biol. 10:42-56.
3. Asakura, S., and T. lino. 1972. Polymorphism of Salmo-nella flagella as investigated by means of in vitro copoly-merization of flagellins derived from various strains. J.Mol. Biol. 64:251-268.
4. Bode, W., J. Engel, and D. Winklair. 1972. A model ofbacterial flagella based on small-angle X-ray scatteringand hydrodynamic data which indicate an elongated shapeof the flagellin protomer. Eur. J. Biochem. 26:313-327.
5. Clausen, J. 1972. Immunochemical techniques for theidentification and estimation of macromolecules, p. 397-572. In T. S. Work and E. Work (ed.); Laboratorytechniques in biochemistry and molecular biology, vol. 1,part 3. Elsevier/North-Holland Publishing Co., Amster-dam.
6. Davis, B. 1964. Disc electrophoresis. Ann. N.Y. Acad.Sci. 121:404-427.
7. Emerson, S. U., K. Tokuyasu, and M. I. Simon. 1970.Bacterial flagella: polarity of elongation. Science 169:190-192.
8. Fujita, H., S. Yamaguchi, T. Taira, and T. lino. 1981. Asimple method for the isolation of flagellar shape mutantsin Salmonella. J. Gen. Microbiol. 125:213-216.
9. Gornall, A. G., C. J. Bardawill, and M. M. David. 1949.Determination of serum proteins by means of the biuretreaction. J. Biol. Chem. 177:751-766.
10. Iino, T. 1969. Polarity of flagellar growth in Salmonella. J.Gen. Microbiol. 56:227-239.
11. Komeda, Y., and T. Iino. 1979. Regulation of expressionof the flagellin gene (hag) in Escherichia coli K-12:analysis of hag-lac gene fusions. J. Bacteriol. 139:721-729.
12. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
13. Miller, J. H. 1972. Experiments in molecular genetics.Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
14. O'Farrell, P. H. 1975. High resolution two-dimensionalelectrophoresis of proteins. J. Biol. Chem. 250:4007-4021.
15. Shirakihara, Y., and T. Wakabayashi. 1979. Three-dimen-sional image reconstruction of straight flagella from amutant Salmonella typhimurium. J. Mol. Biol. 131:485-507.
16. Sleytr, U. B., and A. M. Glauert. 1973. Evidence for anempty core in a bacterial flagellum. Nature (London)241:542-543.
17. Suzuki, T., and T. Iino. 1977. Appearance of straightflagellar filaments in the presence of p-fluorophenylalan-ine in Pseudomonas aeruginosa. J. Bacteriol. 129:527-529.
18. Vesterberg, O., L. Hansdn, and A. Sjosten. 1977. Stainingof proteins after isoelectric focusing in gels by newprocedures. Biochim. Biophys. Acta 491:160-166.
19. Wagenknecht, T., D. DeRosier, L. Shapiro, and A. Weiss-born. 1981. Three-dimensional reconstruction of the fla-gellar hook from Caulobacter crescentus. J. Mol. Biol.151:439-465.
20. Weber, K., and M. Osborn. 1969. The reliability ofmolecular weight determinations by dodecyl sulfate-poly-acrylamide gel electrophoresis. J. Biol. Chem. 244:4406-4412.




![Excretion [2015]](https://static.documents.pub/doc/80x56/55d39c87bb61eb05278b46dd/excretion-2015-55d47f0693bf7.jpg)
















