EXPORT PRODUCTION DEPTH OF EMILI ANI A HUXLEY/IN THE GULF OF CALIFORNIA: AN EVALUATION OF THE ALKENONE UK'37 PALEOSST-PROXY A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAI'IIN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE IN OCEANOGRAPHY DECEMBER 2007 By Amanda S. Pontius Thesis Committee: Brian Popp, Chairperson Fred Mackenzie Robert Bldigare
Transcript
EXPORT PRODUCTION DEPTH OF EMILI ANI A HUXLEY/IN THE GULF OF
CALIFORNIA: AN EVALUATION OF THE ALKENONE UK'37 PALEOSST-PROXY
A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAI'IIN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN
OCEANOGRAPHY
DECEMBER 2007
By Amanda S. Pontius
Thesis Committee:
Brian Popp, Chairperson Fred Mackenzie Robert Bldigare
We certify that we have read this thesis and that, in our opinion, it is
satisfactory in scope and quality as a thesis for the degree of Master
of Science in Oceanography.
THESIS COMMITTEE
ii
iii TABLE OF CONTENTS
List of Tables ........................................................................................ iv List of Figures ....................................................................................... v Chapter 1: Introduction ........................................................................... 1 Chapter 2: Samples and Methods ............................................................ 12
Chapter 3: Results and Discussion ........................................................... 17 Water column properties ............................................................... 17 Depth of alkenone export .............................................................. 17 Depth of maximum alkenone concentration and production .................. 29 Comparison of UK
'37 and in situ temperatures .................................... 38 Chapter 4: Conclusion ........................................................................... 44 References ......................................................................................... 46
iv LIST OF TABLES
1. Alkenone Temperature Calibrations Compiled from the Literature ........... 3
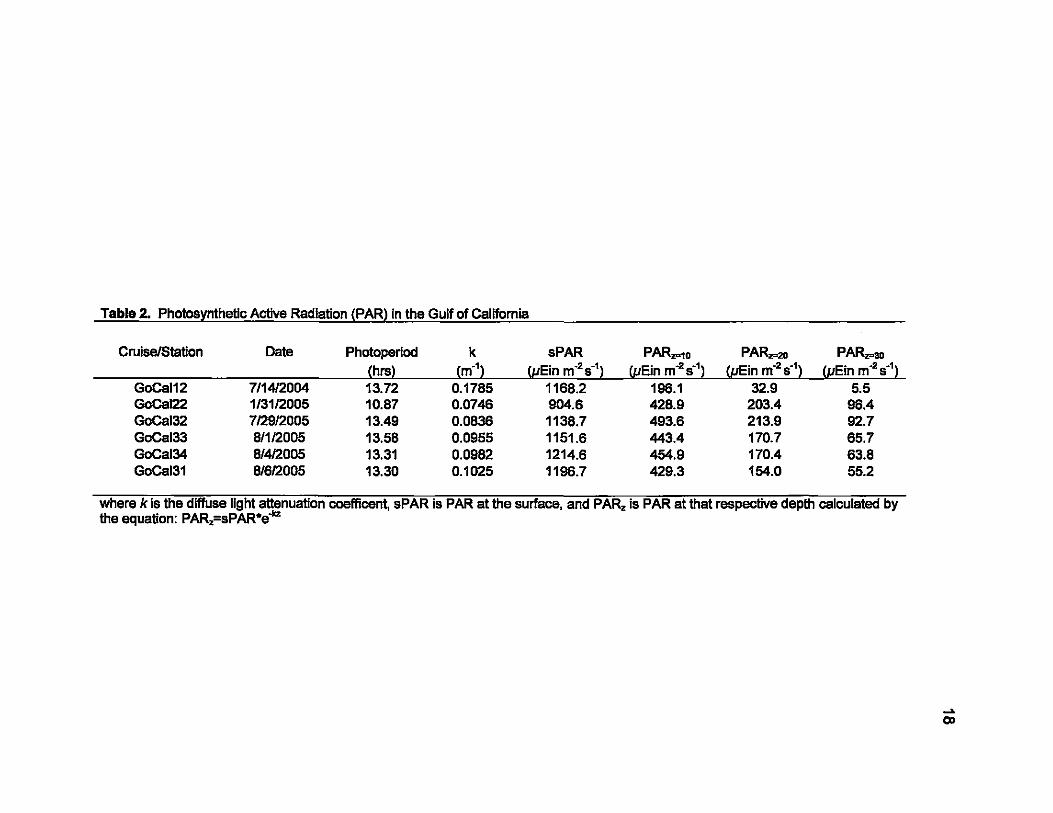
2. Photosynthetic Active Radiation (PAR) in the Gulf of Califomia ............... 18
3. UK'37 and li37:2 Pattems in the Gulf of Califomia ................................... 22
4. Alkenone Export in the Gulf of Califomia .......................................... 27
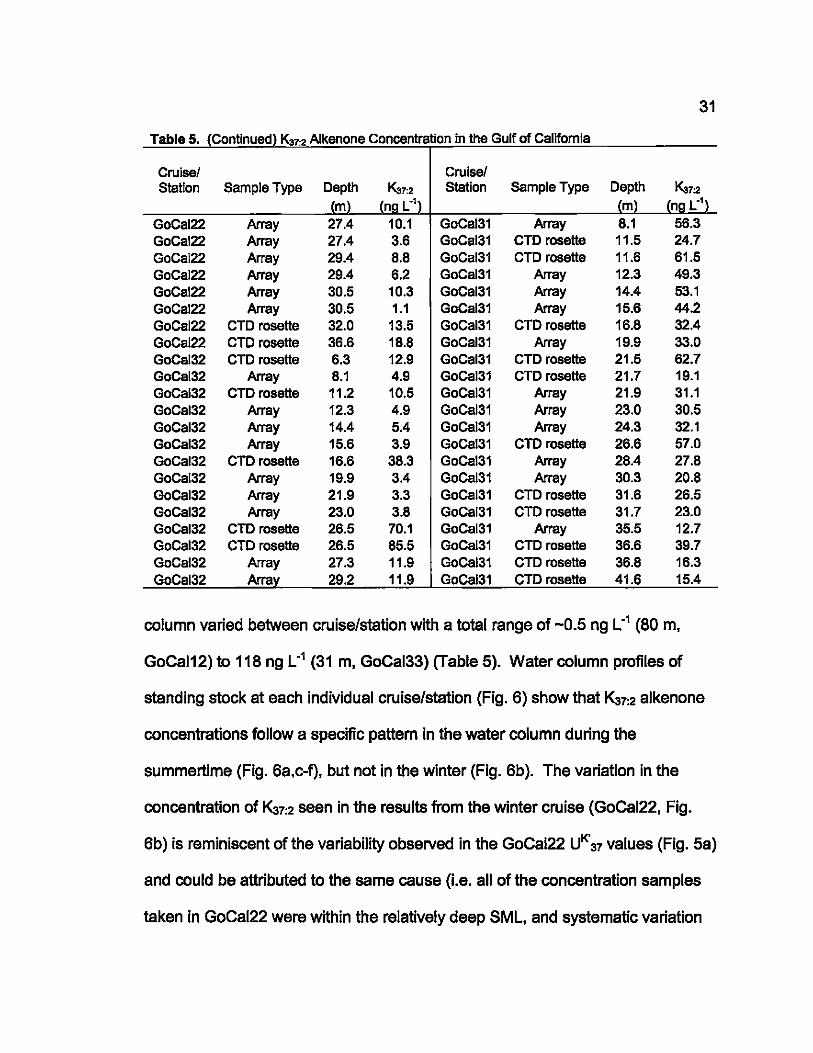
5. K:!7:2 Alkenone Concentration in the Gulf of Califomia ......................... 30
6. K:!7:2 Production in the Gulf of California ........................................... 34
7. A Comparison of Export Depth to the Depths of Maximum Concentration and Production in the Gulf of California ........................ 36
8. Integrated K:!7:2 Concentration and Production in the Gulf of California .................................................................................... 37
9. A Comparison of UK'37 Predicted Temperature with in situ Temperature ............................................................................. .41
v LIST OF FIGURES
Figure
1. Muller et al.. 1998 Core-top Dataset.: .............................................. 3
2. Map ofthe Gulf of Califomia .......................................................... 7
3. Goni et al.. 2001 Time-series Dataset... ........................................... 8
4. Water Column Profiles ................................................................ 19
K' 5. Depth Profiles of U 37 and 5i<37:2 ................................................... 20
6. Depth Profiles of Alkenone i<37:2 Concentration ................................. 32
7. Depth Profiles of i<37:2 Production Rates .......................................... 32
8. UK'37 vs. in situ temperatures ......................................................... 39
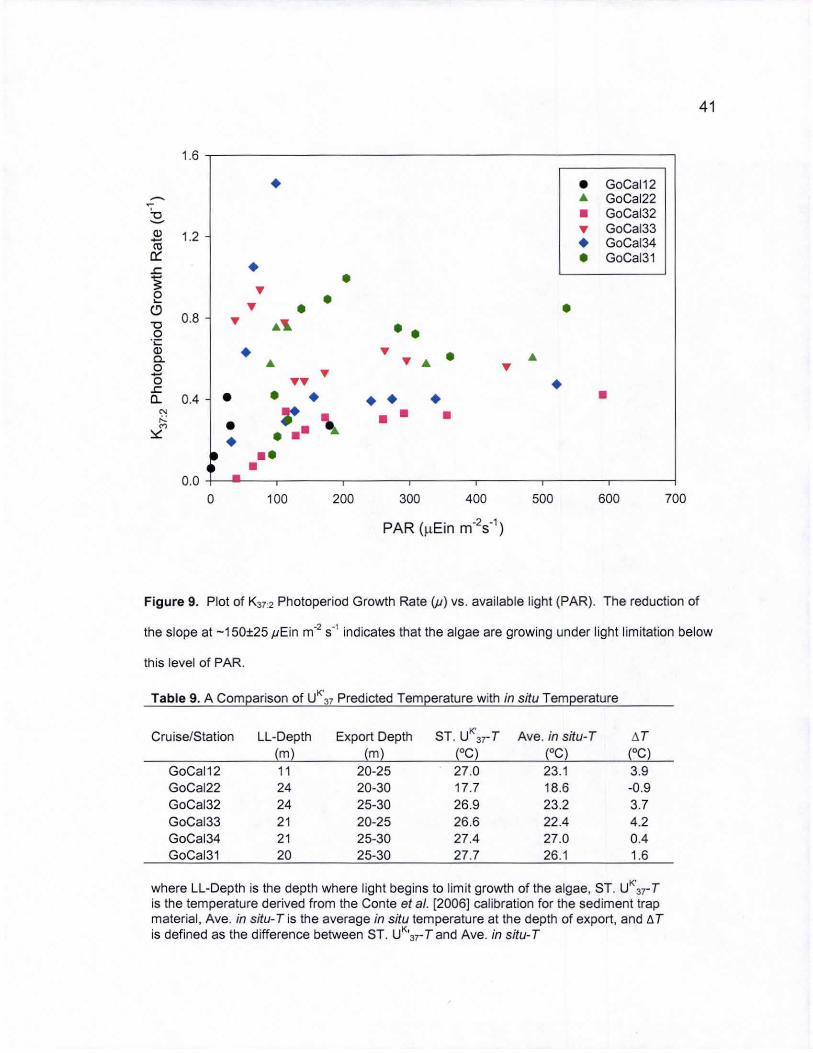
9. i<37:2 Photoperiod Growth Rate (P) vs. available light (PAR) ................. 41
CHAPTER 1 INTRODUCTION
1
Since their discovery and identification in sediments collected from Walvis
Ridge off West Africa [Boon et al. 1978] and the Black Sea [deLeeuw et al .•
1980]. alkenones (Car-Cas di-. trio. and tetraunsaturated methyl and ethyl
keytones) have been found to possess many characteristics that lend themselves
to be used as biomarkers. A1kenones in the open ocean are thought to be
produced exclusively by members of the Class Haptophyceae; most notably the
cosmopolitan species Emiliania huxleyi and the closely related Gephyrocapsa
oceanica [Marlow et al .• 1984a. b]. Unlike the majority of biologically synthesized
compounds. the chemical structure of alkenones contains the 'trans' double bond
configuration. which may help to prevent biodegradation in sediments [Rechka
and Maxwell. 1988a. b]. Most importantly for paleoceanographic applications.
the relative abundance of the double bonds in the compound has been found to
vary linearly with growth temperature (gT) of the biosynthesizing algae in
laboratory cultures [Prahl and Wakeham. 1987; Prahl at al.. 1988] and in the field
[Conte and Eglinton. 1993; Temois at al. 1997]. Focusing on the Ca7 alkenones.
Brassel et al. [1986] defined an alkenone unsaturation index (UKa7) that is
calculated from the relative abundances of the C37 methyl alkenones containing
where k is the diffuse light attenuatlon-coefficent, sPAR is PAR at the surface, and PAR. is PAR at that respective depth calculated by the equation: PAR.=sPAR*e.Jcz
..... 00
(a)
o
20
I 40
'" -a 60 ~
C
80
Temperature (DC)
1416182022 24 2628 30 32
. N~N '(~';'O'I k~ -1') ,
o 5 10 15 20 25
• ~ • •
- PAR ---4- N+N
100 1# • Temperature
(d)
20
40
;; g- 60 c
80
100
o 200 400 600 800 1000
PAR (~E i n m-2 s-l )
Temperature (ee )
1214 16 1820 22 24 26 28 30 32
' N~N (~OI 'k9~ 1 ) ' ,
o
•
•
5 10 15 20 25
•
~ -• - • . -
o 200 400 600 800 1000
PAR (J,LEin m·2 5.1)
(b)
o
20
40
I a 60 • c
80
100
(e)
Temperature ('"C)
14 15 16 17 18 19 20
N+~ (~I k~·1 ) o 5 10 15 20 25
- PAR ~ N+N
• Array2 e. . . Array1
o 200 400 600 BOO
PAR ().lEin mo2 s·1)
Temperature ('C)
14 16 1820 22 24 26 28 30 32
20
I 40
'" C. 60 c3
60
100
o
N+N (';"'~ I kg-l) ,
• •
5 10 15 20 25
•
. 1· ...: ••• -• ••
o 200 400 600 800 1000
PAR (~E in m-2 s-l)
(c)
20
80
Temperature (DC)
14 16 18 20 22 24 26 28 30 32
N+N (.mol kg-1)
o 5 10 15 20 25
•
•
. ----100 ..
(I)
o 200 400 600 800 1000
PAR (jlEin m-2 s·1)
Temperature (0C)
16 18 20 22 24 26 28 30 32
N~N (~Ol 'k9:1) ,
-2024681012141618 o •
r 20
I 40
'" C. 60 ~
C
80
lOa ••
• •
•
-• •• ...
o 200 400 600 800 1000
PAR (.ein m-2 s-l )
Figure 4. Water column profiles of available light (PAR), dissolved inorganic nitrogen (NO; +
NO,-) and temperature for GoCal12 (a), GoCal22 (b), GoCal32 (c) , GoCal33 (d), GoCal34 (e),
and GoCal31 (f). The temperature profile in GoCal22 was slightly different for each incubation
experiment.
19
parameter are necessary to constrain the depth of alkenone export. For each
station/cruise in the present study, at least one of the two parameters shows a
sufficient range in values for comparison to the sediment trap values with the
exception of the winter cruise (GoCaI22) (Fig. 5). We assumed that UK'37 and
OK372 values measured in the sediment trap material reflect the depth of
alkenone export production , Additional assumptions are that there are no
changes in these parameters during export and that lateral inputs of alkenones
where NA /j37:2 is the smoothed neturel abundance alkenone-specific carbon isotopic pattern determined from the water column profiles (Figure 5). A!Sa7:2 Is the difference between the Naturel Abundance profile and the isotopically labeled profile from the incubation experiments. and Kar:2 PR is the Production Rate of the 1<07'2 alkenones
cruise/station were greatest during cruise 3 (Fig. 7b). and lowest during cruises 1
and 2 (Fig. 7a). Given the results of the Goni et a/. [20011 sediment-trap time
series that found alkenone flux maximizing in the summe~ and minimizing in the
winter, low production rates during the winter cruis, (GoCaI22) are to be
expected. The cause of the low observed production rates during the summer of
2004 (GoCa112, Fig. 7a) is much less straightforward. Sampling resolution
during GoCal12 was reduced with respect to the other cruise/stations, and the
maximum production peak could have been missed (e.g. compare GoCal32 to
GoCal12 from 20-30 m in Fig. 7a). Physical differences in the water column
between the two summers could also explain a reduced production rate in 2004.
Maximum standing stock concentrations in GoCal12 were approximately 50% of
the maximum standing stock observed in GoCal32 (Fig. 6a,c). Concentration
also maximized at the surface during GoCal12 (Fig. 6a), while subsurface
36
maxima was seen in all concentration depth profiles during the following summer
(cruise 3, Fig. 6c-f). This difference could be related to the availability of light.
With respect to depth, PAR diminished much quicker in GoCal12 than in
GoCal32 (Fig. 4a,c) due to the higher attenuation of light. If the availability of
light were limiting production during GoCal12, favorable growth conditions would
be found closer to the surface, and could explain the observed concentration and
production discrepancies.
The depths of maximum Ka7:2 concentration and production rate appear to
coincide with previously determined depths of alkenone export production (Table
7). All depths of maximum Ka7:2 concentration and production rate are within
-5m of the inferred depth of alkenone export production. These results suggest
that alkenones preserved in sediments would likely record the gT of water at 20-
30 m, which during the summer, is well below the SML and is therefore not
indicative of SST at that time.
Table 7. A Comparison of Export Depth to the Depths of Maximum Concentration and Production in the Gulf of California
Cruise/Station Export Depth 1<:.7:2 Maxima PRMaxima
(m) (m) (m)
GoCal12 20-25 10.4 21.5
GoCal22 20-30 11.6-36.6 21.9
GoCal32 25-30 26.5 27.3
GoCal33 20-25 18.5-31.4 21.9
GoCal34 25-30 28.4-36.8 28.4
GoCal31 25-30 21.5 24.3
37
Depth integrated alkenone concentration and production rates were
determined for each station (Table 8). The inventory of K37:2 in the upper water
column (0-40 m) was greatest when a subsurface maximum was present (Cruise
3) and lowest when absent (Cruises 1 and 2) (Table 8). Integrated KJ7:2
production rates for the upper water column (0-35 m) varied with respect to
cruise/station and differed in relative magnitude to the integrated concentrations.
Highest integrated concentration was seen in GoCal33, while highest integrated
production rate was seen in GoCal32 (Table 8). Integrated concentration and
production rate were equal and low during GoCal12 and GoCa122.
Concurrent measurements of integrated KJ7:2 production and sediment
trap accumulation rate allow calculation of the efficiency of alkenone export
production. Using the surface area of the collection cups and the duration of
deployment, alkenone concentrations collected with the sediment trap are used
to calculate flux and compared to integrated production measured in the
overlying water column (Table 8). Trapping efficiencies ranged from 9-34% with
an average efficiency of -20%, indicating that -80% of the alkenones produced
Table 8. Int rated Ka7:2 Concentration and Production in the Gulf of califomia
CrulsefStetion Array # IC IP IP Trap lKa7:i1 % Eff. 119 m·2 11 m·2 cr' 119 d·' 11 cr'
where IC and IP are the Integrated concentration and production of the upper 4Om, and Trap lKa7:2l is the flux of Ka7:2 alkenones to the sediment traps
38
in the euphotic zone are not represented by the material collected with the
sediment traps. Previous studies on the trapping efficiencies of this type of
sediment trap deployed at four different locations (BATS, HOT, Dabob Bay, and
the Baltic Sea) ranged from 44-69% [Buesseler at al., 2007 and references
therein] indicating that a significant fraction of the lost alkenone export can be
attributed to the limitations of sediment trap measurements (Le. alkenones
degrading within the collection cup or failing to remain in the collection cup due to
turbulence at the sediment cup-water interface [Buesseler et al., 2007]).
Assuming that the collection efficiency of the sediment traps deployed in the
present study is similar to those discussed in Buesseler et al. [2007] (i.e. -50%),
then the -20% collected with the sediment traps represents export of -40% of
the total production. This leaves -60% of the alkenones produced in the
euphotic zone unaccounted for. Possible factors contributing to this loss include
alkenones being recycled in the upper water column, .subjected to lateral
transport, or degraded during export. Additionally, because of the significant
fraction of alkenone production that is not exported, it appears unlikely that
accurate estimates of paleoproduction could be determined by measuring
alkenone concentrations or accumulation rates in sediments.
3.4. Comparison of If37 and in situ temperatures
UK'37 measured in the SPM collected throughout the water column is
compared to in situ temperatures in order to determine how well the most recent,
sediment-based, UK' 37"temperature relationship of Conte et al. [2006] predicts gT
39
in the Gulf of California (Fig. 8). Results shown in Figure 8 appear qualitatively
similar to the corresponding plot of Goni et a/. [2001] (Fig. 2) in that both
relationships are relatively linear until in situ temperatures reach -26°C. The
reduction in slope at temperature extremes has been documented for other water
column samples [Conte et a/. [2006], and references therein] and has been
attributed to the cell 's alkenone-based adaptation to temperature limitation at the
extremes of its growth temperature range [Conte et al. 1998]. However, when
the Conte et al. [2006] UK'3rSST relationship is superimposed onto Figure 8,
1.0 ,---------------:._~,.._e4 ... ~t_--__, " .. -., I 0.9 •
where LL-Depth is the depth where light begins to limit growth of the algae, ST . UK'37- T
is the temperature derived from the Conte et a/. [2006] calibration for the sediment trap material, Ave. in situ-T is the average in situ temperature at the depth of export, and l!. T is defined as the difference between ST . UK
'3], T and Ave. in situ- T
42
150±25 pEin m"2 s"' fell within the depth range of alkenone export production only
during the winter cruise. A comparison of the sediment trap-derived UK"3T"Twith
the average in situ temperature at the depth of export shows that in situ
temperatures are indeed overestimated by UK·37 temperature predictions when
light is limiting growth (Table 9).
The magnitude at which UK" 3rderived growth temperatures overestimate
in situ temperatures during the summer cruises varies with respect to location
(-4°C at stations 2 and 3 and -1°C at stations 4&1) and is probably influenced by
the depth of the nutricline at that given station. Dissolved inorganic nitrogen
concentrations were below detection limits throughout the range of incubation
experiment depth (0-40 m) for GoCal34 and GoCal31 (see Fig. 4e-f). Assuming
that the alkenone-producing algae growing at these two stations are being
affected by nutrient limitation as well as light limitation, the overestimating effect
of light limitation is partially canceled out due to the underestimating effect of
nutrient limitation, resulting in a relatively small overestimate of in situ
temperature (-1°C). Dissolved inorganic nitrogen concentrations are low, but
detectable in GoCal12, GoCal32, and GoCal33 and therefore growth of the
alkenone-producing algae was not likely nutrient-limited, resulting in a relatively
large overestimate of in situ temperature (-3°C).
This overestimate of in situ temperature explains why UK·3T" Tfrom the
sediment traps did not underestimate SML-T as severely as predicted based on
the temperature difference between the SML and the depth of export (Section
43
3.2). The magnitude of the SML-T underestimate was reduced because light
limitation caused the UK' 3r T to overestimate in situ growth temperature. Since
favorable conditions for alkenone growth are often found below the SML
throughout the world ocean [Prahl et aI., 1993 and 2001; Temois et a/., 1997;
Ohkouchi et aI., 1999], the effects we document here may extend to other sites in
the worlds ocean.
CHAPTER 4 CONCLUSION
44
The depth of alkenone export production in the Gulf of California was
constrained to -20-30 m, which is well below the SML during the summer. As a
consequence, SST is systematically underestimated by UK' 37 collected in shallow
sediment traps. The magnitude of the underestimate falls within the range of
variability observed in most UK' 3r T relationships based on core-top samples
[Muller et a/. 1998; Conte et a/. 2006]. However, the discrepancy between UK'3r
temperature recorded in sediment trap material and SST was smaller than
expected based on the temperature gradient through the upper water column
and the depth of alkenone export production. It appears that the physiological
effect of growth under light limitation and its affect on UK' 37 values helped to
minimize the difference between UK'3rtemperature recorded in sediment trap
material and SST. Therefore, relatively accurate SST estimates can be obtained
from UK'37 measurements, even when the alkenone producing algae are
exporting from depths below the SML (within -±3°C). On the other hand,
recognition of the depth of export in ancient record could substantially improve
paleotemperature estimates using the alkenone unsaturation index.
Concurrent measurements of integrated production in the water column
and flux to shallow sediment traps allow for an estimate of the efficiency of
alkenone export production. Efficiencies ranged from 9-34% and averaged
-20%. Assuming an -50% loss due to limitations in the sediment trap collection
method, it is estimated that the remaining -60% of the alkenone production is
45
recycled through grazing or lost during transport (presumably to degradation or
lateral transport). Unfortunately. the relatively large fraction of production that is
not exported indicates that accurate estimates of paleoproduction could not be
made by measuring alkenone accumulation rates in sediments.
46 REFERENCES
1. Armstrong, F. A., Steams, J.R., and Strickland, J.H., 1976. The measurement of upwelling and subsequent biological processes by means of the Technicon AutoAnalyzer™ and associated equipment, Deep Sea Research 114: 381-389.
2. Atlas, E.L., Hager, S.W., Gordon, LI., and Park, P.K., 1971. A practical manual for the use of the Technicon AutoAnalyzer™ in seawater nutrient analysis: Revised, Tech Rep. 215 Ref. 71-22,48 pp., Dep. Of Oceanogr., Oregon State Univ., Corvallis.
3. Boon, J.J., Meer, F.W.V.D., Schuyl, P.J.W., deleeuw, J.W., and Schenck, P.A.,1978. Organic geochemical analysis of core samples from site 362 Walvis Ridge, DSDP leg 40. In Initial reports of the deep sea drilling project, Vol. 40, Bolli, H.M, et a/., editors, US Government Printing Office, Washington, pp. 627-637.
4. Brassell, S. C., Eglinton, G., Marlowe, I. T., Pflaumann, U., and Samthein, M., 1986. Molecular stratigraphy: a new tool for climatic assessment, Nature 320:129-133.
5. Buesseler et aI., 2007. An assessment of the use of sedimet traps for estimating upper ocean particle fluxes, Joumal of Marine Research 65:345-416.
6. Christie, W.W., 1973. Upid Analysis: Isolation, Separation. Identification. and structural Analysis of Upids, Elsevier, New York.
7. Conte, M.H. and Eglinton, G., 1993. Alkenone and alkenoate distributions within the euphotic zone of the eastern North Atlantic: correlation with production temperature, Deep Sea Research 140: 1935-1961.
8. Conte, M. H., Thompson, A., lesley, D., and Harris, R., 1998. Genetic and physiological influences on the alkenoneJalkenoate versus growth temperature relationship in Emiliania huxleyi and Gephyrocapsa oceanica, Geochim. Cosmochim. Acta, 62:51-68.
9. Conte, M.H., Sicre, M., Ruhlemann, C., Weber, J.C., Schulte, S., Schulz-Bull, D., Blanz, T., 2006. Global temperature calibration of the alkenone unsaturation index (UK'37) in surface waters and comparison with surface sediments, G3 vol. 7 no. 2. doi: 10.1 02912005GC001 054
10.Deines, P., langmuir, D., and Harmon, R., 1974. Stable carbon isotope ratios and the existence of a gas phase in the evolution carbonate ground waters, Geochim. Cosmochim. Acta. 38:1147-1164.
11.deleeuw, J.W., Meer, F.W.V.D., Rijpstra, W.I.C., and Schenk, P.A., 1980. On the occurrence and structural identification of long chain unsaturated alkenones and hydrocarbons in sediments. In: Advances in organic geochemistry 1979, Douglas, A.G. and Maxwell, J.R., editors, Pergamon Press, Oxford, pp. 211-217.
12. Dickson, A. G., and Millero. F. J., 1987. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media, DeepSea Research 34:1733-1743.
47
13. Dickson, A.G., 1990a. Standard potential of the reaction: AgClis) + 1.2H2(g) = Ag(s) + 11 CI(aq), and the standard acidity constants of the ion HSO-4 in synthetic seawater 273.15 to 318.15°K, J. Chem. Thermodyn.22:113-127.
14. Dickson, A.G., 1990b. Thermodynamics of the dissociation of boric acid in synthetic seawater 273.15 to 318.15°K, Deep Sea Research, Part A 37:755-766.
15. Epstein, B. L., d'Hondt, S., and Hargraves, P. E., 2001. The possible metabolic role of C37 alkenones in Emiliania hux/eyi, Org. Geochem. 32:867-875.
16. Epstein, B. L., d'Hondt, S., Quinn, J. G., Zhang, J., and Hargraves, P. E., 1998. An effect of dissolved nutrient concentrations on alkenone-based temperature estimates, Paleoceanography 13:122-126.
17. Gaxiola-Castro, G., Alvarez-Borrego, S. A., Lavin, M. F., Zirino, A., and Najera-Martinez, S., 1999. Spatial variability of the photosynthetic parameters and biomass of the Gulf of California phytoplankton, Jouma/of Plankton Research 21:231-245.
18. Grice, K. et a/., 1998. Effects of zooplankton herbivory on biomarker proxy records, Paleoceanography 13:686-693.
19.Goni, M. A., Hartz, D. M., Thunell, R. C., and Tappa, E., 2001. Oceanographic considerations for the application of the alkenone-based paleotemperature UK" 37 index in the Gulf of California, Geochim. Cosmochim. Acta 65:545-557.
20.Gran, G., 1952. Determination of the equlivance point in potentiometric titrations, II. Analyst 77:661-671.
21.Hama, T., Hama, J., and Handa, N., 1993. 13C tracer methodology in microbial ecology with special reference to primary production processes in aquatic environments. In Advances in Microbial Ecology (ed. J. GWYNFRYN), pp. 39-83. Plenum Press.
22. Hamanaka, J., Sawada, K., and Tanoue, E., 2000. Production rates of C37 aiken ones determined by 13C-labeling technique in the euphotic zone of Sagami Bay, Japan, Organic GeochemistI}' 31:1095-1102.
23. Haxo, F. T., 1985. Photosynthetic action spectrum of the coccolithiophorid, Emiliania huxleyi (Haptophyceae): 19'-hexanolyoxyfucoxanthin as antenna pigment, Joumal of Phycology 21 :282-287.
24. Hayes, J. M., Freeman, K. H., Popp, B. N., and Hoham, C. H., 1990. Compound-specific isotopic analyses: a novel tool for reconstruction of ancient biogeochemical processes, Organic GeochemistI}' 16:1115-1128.
25.Johnson, K.M., Willis, K.D., Butler, WK, Johnson, WK, and Wong, C.S., 1993. Colulometric total carbon dioxide analysis for marine studies: maximizing the performance of an automated gas extraction system and coulometric detector, Mar. Chem.44:167-188.
48
26. Karl, D.M., Winn, C.D., Hebel, D.V.W., and Letelier, R, 1990. Hawaii Ocean Time- series Program Field and Laboratory Protocols, September 1990. School of Ocean and Earth Science and Technology, Univ. of Hawaii, Honolulu, HI, 72 pp.
27. Knappertsbusch, M., 1993. Geographic distribution of living and Holocene coccolithiophores in the Mediterranean Sea, Marine Micropa/entology 21:219-247.
28. Knauer, GA, Martin, J.H., and Bruland, K., 1979. Fluxes of particulate carbon, nitrogen, and phosphorous in the upper water column of the Northeast Pacific Ocean, Deep-Sea Research 26:97-108.
29.Kroopnick, P., 1985. The distribution of l3C in rC02 in the world oceans, Deep Sea Research 132:57-84.
30.Laws, EA, Popp, B.N., Bidigare, RR, Kennicutt, M.C., Macko, SA, 1995. Dependence of phytoplankton carbon isotopic composition on growth rate and [C02]aq: Theoretical considerations and experimental results, Geochim. Cosmochim. Acta. 59:1131-1138.
31.Lewis, E., and Wallace, D., 1998. Program Developed for C02 System Calculations. ORNUCDIAC-105. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy, Oak Ridge, Tennessee.
32. Malinvemo, E., Prahl, F.G., Popp, B.N., Ziveri, P., in prep. Alkenone abundance and its relationship to the coccolithiophore in Gulf of California surface waters.
33. Marlowe, I. T., Brassell, S. C., G. Eglinton, G., and Green, J. C., 1984a. Long chain unsaturated ketones and esters in living algae and marine sediments, in Advances in Organic Geochemistry 1983 (P. A. Schenck, J. W. de Leeuw, and G. M. W. Lijmbach, eds.), Organic Geochemistry, 6:135-141.
34.Marlowe, I. T., Green, J. C., Neal, A. C., Brassell S. C., Eglinton, G., and Course, P. A., 1984b. Long chain (n-C3rC39) alkenones in the Primnesiophyceae. Distribution of alkenones and other lipids and their taxonomic significance, Br. Phyco/. J., 19:203-216.
35. Mehrbach, C., C. H. Culberson, J. E. Hawley, and R M. Pytkowicz, . Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheriC pressure, Limnology and Oceanography 18, 897-907 (1973).
37.Muller, P. J., Kirst, G., Ruhland, G., von Storch, I., and Rosell-Mele, A., 1998. Calibration of the alkenone paleotemperature index UK' 37 based on coretops from the eastern South Atlantic and the global ocean (600N-600S), Geochim. Cosmochim. Acta 62:1757-1772.
49
38.0hkouchi, N., Kawamura, K., Kawahata, H., Okada, H., 1999. Depth ranges of alkenone production in the central Pacific Ocean. Global Biogeochemical Cycles 13:695-704.
39. Pond, D. W., Harris, R P., 1996. The lipid composition of the coccolithophore Emiliania huxleyi and its possible ecophysiological significance. Joumal of the Marine Biological Association of the United Kingdom 76:579-594.
40.Popp, B. N., Prahl, F. G., Wallsgrove, R. J., and Tanimoto, J., 2006a. Seasonal patterns of alkenone production in the subtropical oligotrophic North Pacific, Paleoceanography, 21:P Al 004, doi: I 0.1029/200SP A00116S.
41. Popp, B.N. et a/., 2006b. A new method for estimating growth rates of alkenone-producing haptophytes, Umnologyand Oceanography: Methods 4:xx-xx.
42.Popp, B.N., Kenig, F., Wakeham, S.G., Laws, E.A., and Bidigare, R.R, 1998. Does growth rate affect ketone unsaturation and intracellular carbon isotopic variability in Emiliania hux/eyn Paleoceanography 13:35-41.
43. Prahl F. G., Pilskaln C. H., and Sparrow M. A., 2001. Seasonal record for alkenones in sedimentary particles from the Gulf of Maine, Deep-sea Res. 48:515-528.
44. Prahl, F. G., and Wakeham, S. G., 1987. Calibration of unsaturation patterns in long-chain ketone compositions for palaeotemperature assessment, Nature 330:367-369.
45. Prahl, F. G., Popp, B. N., Karl, D. M., and Sparrow, M. A., 2005. Ecology and biogeochemistry of alkenone production at subtropical North Pacific Station ALOHA, Deep-Sea Research 152:699-719
46. Prahl, F. G., Wolfe, G. V., Sparrow, M. A., 2003. Physiological Impacts on Alkenone Paleothermometry, Paleooceanography 18:1025-1032.
47. Prahl, F.G., Collier, RB., Dymond, J., Lyle, M., Sparrow, M.A., 1993. A biomarker perspective on prymnesiophyte productivity in the northeast Pacific Ocean, Deep-Sea Research 140:2061-2076.
48. Prahl, F.G., Muehlhausen, L.A., Lyle, M., 1989. An organic geochemical assessment of oceanographic conditions at MANOP Site C over the past 26,000 years, Paleoceanography 4:495-51 O.
49. Prahl, F.G., Rontani, J., Volkman, J.K., Sparrow, M.A., Royer, I.M., 2006. Unusual C35 and C36 alkenones in a paleoceanographic benchmark strain of Emiliania huxleyi, Geochimica et Cosmochimica Acta 70:2856-2867.
50. Rechka, J. A., and Maxwell, J. R, 1988a. Characterization of alkenone temperature indicators in sediments and organisms, in Advances in Organic Geochemistry 1987 (L. Mattavelle and L. Novelli, ads.) Organic Geochemistry 13:727-734.
51. Rechka, J. A., and Maxwell, J. R, 1988b. Unusual long chain keytones of algal origin, Tetrahedron Letters, 29:2599-2600.
52. Rost, B., Riebsell, U., Burkhardt, S., and Sultemeyer, D., 2003. Carbon acquisition of bloom-forming marine phytoplankton, Umno/. Oceanogr. 48:55-67.
50
53. Rost, B., Zondervan, I., Riebesell, U., 2002. Light-dependent carbon isotopic fractionation in the coccolithophorid Emiliania huxleyi, Umno/. Oceanogr. 47:120-128.
54.Schlenk, H., and Gellerman, J., 1960. Esterfication offatty acids with diazomethane on a small scale, Anal. Chem., 32:1412-1414.
55. Shin, K.-H., Tanaka, N., Harada, N., and Marty, J.-C., 2002. Production and turnover rates of C37 alkenones in the eastern Bearing Sea: Implications for the mechanism of a long duration of Emiliania huxleyi bloom, Prog. Oceanogr.55:113-129.
56. Strickland, J. and Parsons, T., 1972. A Practical Handbook of Seawater Analysis, 2nd ed., bull. 167,310 pp., Fish. Res. Board of Can., Ottawa, Ont.
57. Temois, Y., Sicre, M.-A., Boireau, A., Conte, M.H., Eglinton, G., 1997. Evaluation of long-chain alkenones as paleotemperature indicators in the Mediterranean Sea, Deep-Sea Research 144:271-286.
58. Thumell, R.C., 1998. Seasonal and annual variability in particle fluxes in the Gulf of California: A response to climate forcing, Deep Sea Research I 45:2059-2083.
59. Versteegh, G., Riegman, R., de Leeuw, J., Jansen, J., 2001. UK'37 values for Isochrysis galbana as a function of culture temperature, light intensity, and nutrient concentrations, Organic Geochemistry 32:785-794.
60.Ziveri, P., and Thurnell, R., 2000. Coccolithiophore export production in Guaymas Basin, Gulf of California: Response to climate forcing, DeepSea Research 1147:2073-2100.