Exposure to Heat Shock Affects Thermosensitivity of the Locust Flight System R. M. Robertson, * H. Xu, K. L. Shoemaker, and K. Dawson-Scully Department of Biology, Queen's University, Kingston, Ontario, Canada, K7L 3N6 SUMMARY The natural habitat of the migratory locust, Locustu mi- grutoriu, is likely to result in locusts being heat stressed during their normal adult life. It is known that locusts exhibit a heat-shock response: exposure to 45°C for 3 h induces thermotolerance and the expression of heat- shock proteins. We investigated the effects of exposure to heat-shock conditions on the thermosensitivity of flight rhythm generation in tethered, intact animals and in deafferented preparations. Heat shock had no effect on wingbeat frequency measured at the start of flight se- quences, nor did it affect the postimaginal maturation of this parameter. During sustained flight, heat shock slowed the characteristic asymptotic reduction of wing- beat frequency. Wingbeat frequency of heat-shocked an- imals was less sensitive to temperature in the range 24" to 47°C than that of control animals, and the upper tem- perature limit, above which flight rhythms could not be produced, was 6" to 7°C higher in heat-shocked animals. These results were mirrored in the response of deaffer- ented preparations, indicating that modifications in the properties of the flight neuromuscular system were in- volved in mediating the response of the intact animal. We propose that exposure to heat shock had the adaptive consequences of reducing thermosensitivity of the neural circuits in the flight system and allowing them to operate at higher temperatures. Keywords: insect, flight, temperature, heat shock, wing- beat frequency. 0 1996 John Wiley & Sons, Inc. INTRODUCTION Neural operation within mammalian nervous sys- tems is to a large extent protected from variations in the physical characteristics of the environment. However, many other animal species lack homeo- static mechanisms that allow independence from environmental constraints. This is particularly true for poikilotherms whose internal temperatures are not physiologically regulated but are allowed to vary as ambient temperature fluctuates. Given the profound influence of temperature on neural pro- cesses (for reviews, see Montgomery and Macdon- ald, 1990; Janssen, 1992), this raises the problem of how the nervous system maintains adaptive function under different thermal regimens. The Received August 29, 1995; accepted November 2, 1995 Journal of Neurobiology, Vol. 29, No. 3, pp. 367-383 ( 1996) 0 1996 John Wiley &Sons, Inc. CCC 0022-3034/96/030367- I? * To whom correspondence should be addressed. problem is most pressing for animals that experi- ence large, rapid temperature changes as a conse- quence of their ecology. Behavioral adaptations for thermoregulation can mitigate, but usually not eradicate, the effects of experiencing extremes in ambient temperature. It is important to determine the neural adaptations that compensate for tem- perature variability, and those that permit opera- tion at extremely high temperatures. The migratory locust, Locusta Pnigratoriu, is na- tive to semiarid regions of equatorial Africa and is thus normally exposed to a range of high environ- mental temperatures (Uvarov, 1966). Locusts of- ten fly when ambient temperature exceeds 30°C and the heat produced by working flight muscles significantly elevates thoracic temperature above ambient by between 6" and 10°C (Weis-Fogh, 1956, 1964). Under laboratory conditions the per- missive lower and upper limits of thoracic temper- ature for flight are around 24" and 42°C (Weis- Fogh, 1956; Neville and Weis-Fogh, 1963). As in other insects (May, 1981; Oertli, 1989), the wing- 367
Transcript
Exposure to Heat Shock Affects Thermosensitivity of the Locust Flight System
R. M. Robertson, * H. Xu, K. L. Shoemaker, and K. Dawson-Scully
Department of Biology, Queen's University, Kingston, Ontario, Canada, K7L 3N6
SUMMARY
The natural habitat of the migratory locust, Locustu mi- grutoriu, is likely to result in locusts being heat stressed during their normal adult life. It is known that locusts exhibit a heat-shock response: exposure to 45°C for 3 h induces thermotolerance and the expression of heat- shock proteins. We investigated the effects of exposure to heat-shock conditions on the thermosensitivity of flight rhythm generation in tethered, intact animals and in deafferented preparations. Heat shock had no effect on wingbeat frequency measured a t the start of flight se- quences, nor did it affect the postimaginal maturation of this parameter. During sustained flight, heat shock slowed the characteristic asymptotic reduction of wing- beat frequency. Wingbeat frequency of heat-shocked an-
imals was less sensitive to temperature in the range 24" to 47°C than that of control animals, and the upper tem- perature limit, above which flight rhythms could not be produced, was 6" to 7°C higher in heat-shocked animals. These results were mirrored in the response of deaffer- ented preparations, indicating that modifications in the properties of the flight neuromuscular system were in- volved in mediating the response of the intact animal. We propose that exposure to heat shock had the adaptive consequences of reducing thermosensitivity of the neural circuits in the flight system and allowing them to operate a t higher temperatures. Keywords: insect, flight, temperature, heat shock, wing- beat frequency.
0 1996 John Wiley & Sons, Inc.
INTRODUCTION
Neural operation within mammalian nervous sys- tems is to a large extent protected from variations in the physical characteristics of the environment. However, many other animal species lack homeo- static mechanisms that allow independence from environmental constraints. This is particularly true for poikilotherms whose internal temperatures are not physiologically regulated but are allowed to vary as ambient temperature fluctuates. Given the profound influence of temperature on neural pro- cesses (for reviews, see Montgomery and Macdon- ald, 1990; Janssen, 1992), this raises the problem of how the nervous system maintains adaptive function under different thermal regimens. The
Received August 29, 1995; accepted November 2, 1995 Journal of Neurobiology, Vol. 29, No. 3, pp. 367-383 ( 1996) 0 1996 John Wiley &Sons, Inc. CCC 0022-3034/96/030367- I ?
* To whom correspondence should be addressed.
problem is most pressing for animals that experi- ence large, rapid temperature changes as a conse- quence of their ecology. Behavioral adaptations for thermoregulation can mitigate, but usually not eradicate, the effects of experiencing extremes in ambient temperature. It is important to determine the neural adaptations that compensate for tem- perature variability, and those that permit opera- tion at extremely high temperatures.
The migratory locust, Locusta Pnigratoriu, is na- tive to semiarid regions of equatorial Africa and is thus normally exposed to a range of high environ- mental temperatures (Uvarov, 1966). Locusts of- ten fly when ambient temperature exceeds 30°C and the heat produced by working flight muscles significantly elevates thoracic temperature above ambient by between 6" and 10°C (Weis-Fogh, 1956, 1964). Under laboratory conditions the per- missive lower and upper limits of thoracic temper- ature for flight are around 24" and 42°C (Weis- Fogh, 1956; Neville and Weis-Fogh, 1963). As in other insects (May, 1981; Oertli, 1989), the wing-
367
beat frequency ( WBF) of tethered locusts increases with increasing thoracic temperature. At least part of the increase is due to the effects of temperature on the neuronal circuits in the central nervous sys- tem (Foster and Robertson, 1992). Much informa- tion has been gathered about the central nervous circuits controlling locust flight (for reviews, see Robertson, 1986, 1989, 1995), and computer modeling has demonstrated that the known cir- cuitry is sufficient for generating robust flight rhythms (Grimm and Sauer, 1995). The locust flight system thus provides an excellent, experi- mentally tractable, system for investigating ther- mal adaptations of nervous tissue. Recent studies have shown that the membrane properties oflocust flight neurons (Xu and Robertson, 1994), and the parameters of synaptic potentials recorded in the flight neuropile (Robertson, 1993), are affected by temperature. A consequence of temperature that has not yet been investigated in this system is the result of an exposure to heat-shock conditions.
Many organisms, both plant and animal, be- come thermotolerant of nonpermissive tempera- tures after previous exposure to high, sublethal temperatures for a period of time. In most cases this form of thermotolerance is coincident with the enhanced expression of a set of proteins known as heat-shock proteins (HSPs) (Ritossa, 1962), which are thought to protect protein function by acting as molecular chaperones (for reviews, see Lindquist and Craig, 1988; Craiget al., 1993; Geor- gopoulos and Welch, 1993; Harrington et al., 1994). There is mounting evidence that thermotol- erance is imparted by HSPs (for reviews, see Kam- pinga, 1993; Moseley, 1994). Indeed, there is a cor- relation between the characteristic temperature of the ecological niche of some species of lizards and the level of expression of HSPs at normal tempera- tures, without heat shock (Ulmasov et al., 1992). Similarly, certain particularly thermotolerant ant species ( Cataglyphis), may synthesize HSPs in an- ticipation of foraging in the desert where surface temperatures exceed 50°C (Gehring and Wehner, 1995). L. migrcitoria also lives in such inhospitable regions (Uvarov, 1966), and a heat-shock response has been demonstrated: exposure to 45°C for 0.5 to 4.5 h induces expression of HSPs and permits survival on subsequent exposure to a normally le- thal temperature of 50°C (Whyard et al., 1986).
We are interested in determining whether exposure to heat-shock conditions might protect neural function rather than just prevent death and thus permit behavior at elevated temperatures. We used WBF as a convenient measure of the opera-
tion of the flight system and we tested whether exposure to heat shock had any effect on the tem- perature sensitivity of this parameter of flight. We also investigated the effect of heat shock on flight rhythms recorded from deafferented preparations in order to confirm that our results from intact an- imals reflected changes in the operation of the flight circuitry. We were particularly interested in effects on the upper temperature limits for tethered flight rhythms and for flight rhythm generation in deafferented nervous systems. Our prediction was that induction of thermotolerance by exposure to heat-shock conditions would have the adaptive effect of raising the upper temperature limit for flight rhythm generation.
MATERIALS AND METHODS
Animals
Adult female and male locusts ( L . migratoria) were ob- tained from a colony in the Department of Biology at Queen's University immediately after the final moult and maintained at 29 k 1 "C, on a 16:8 h light/dark cycle. Only one sex was used within individual experiments to reduce intersex variability. Locusts for separate experi- ments were raised together. When necessary, locusts were individually marked with a felt-tipped permanent marker prior to testing.
Heat-Shock Exposure
Locusts were placed in 2 L ventilated plastic tubs in an incubator at 45°C for 3 h. Control locusts were placed in similar containers at 29°C for 3 h. Both groups of locusts were allowed to recover for at least 1 h at room tempera- ture before being used in experiments.
Test of Thermotolerance
To test the efficacy of the heat-shock treatment at induc- ing thermotolerance, male locusts were collected 1 day, 1 week, 2 weeks, or 3 weeks following their imaginal moult and half were heat shocked as just described. Fol- lowing the l-h recovery period, heat-shocked and control locusts were placed together in a single container and ex- posed to 53°C in the incubator for 1.5 to 3 h. When ap- proximately half of the locusts in the container appeared dead, the container was removed and surviving locusts were counted after a 24 h recovery period. A locust was considered a survivor if strong behavioral function (standing, walking, or wing movement) was observed. A locust was deemed moribund or dead, if it was not stand- ing, if no movement was observed (excluding small twitches ofthe appendages and mouthparts), and ifthere
Flight of Heat-Shocked Locusts 369
was no reflex antenna1 withdrawal to mechanical stimu- lation. We repeated the experiment with a 1 -week recov- ery period (except for the I-day old locusts, which were given a 2-week recovery period). When possible, the to- tal number of animals used in this study was reduced by using locusts in the WBF experiments first (described next) and then using them in the thermotolerance test.
Wingbeat Frequency Measurement
WBF was measured stroboscopically. Each locust was tethered at the pronotum with wax to a copper rod and suspended in the center of a cylindrical wind tunnel (wind speed: 2.5 to 3.0 m/s). When the locust wings ap- peared stationary in the flashing light of the stroboscope, the frequency of flashing was recorded as the WBF. This method of measuring WBF is noninvasive and requires no time to prepare the animal, thereby allowing a large number of animals to be tested rapidly. However, stro- boscopic visual stimulation has been shown capable of entraining WBF. Waldron ( 1968) demonstrated phase locking of the wingbeat to a constant frequency flashing light but only within a narrow frequency range. Gee and Robertson (unpublished observations) have tested the stroboscopic method of WBF determination and found that when there is no attempt to entrain with the strobo- scope, there is no significant difference between electro- myographic (EMG) measurement of WBF and subse- quent stroboscopic measurement. In addition, we are concerned here less with absolute values of frequency than with comparisons between the heat-shocked and control animals.
In some experiments WBF was measured electro- myographically. An EMG electrode (0.1 mm diameter copper wire, insulated except at the tip) was inserted through the cuticle into the forewing first basalar muscle (M97) (after Snodgrass, 1929) and secured with a drop of wax. Muscle activity was amplified and recorded con- ventionally. This method of measuring WBF, although requiring some initial set up, was immediate and thus preferable for continuous flight experiments in which thoracic temperature was being manipulated. There was no obvious difference in the WBF data obtained using either of the two methods.
Initial WBF Experiment
Locusts were divided into four age groups ( I-day-old, I - week-old, 2-week-old, and 3-week-old adults). Each group contained 40 animals. WBF was measured prior to the heat-shock treatment. Each group was then di- vided into two groups so that the mean WBF for each group was approximately equal (each group had an equal number of fast and slow fliers). One of the two groups was then arbitrarily chosen to receive the heat- shock treatment. Following recovery, the WBF was mea- sured for all locusts every day for 6 or 13 days. Animals were flown at the same time each day and WBF was mea-
sured within the first minute of flight. All animals were tested at room temperature (22" to 25°C).
Maintained WBF Experiment
We used adult locusts 2 weeks past their final moult for these experiments. Sixteen locusts were selected and half were heat shocked as already described. Locusts were flown in pairs, one heat-shocked and one control, in a steady wind stream at room temperature (22" to 25°C). At certain times, each animal was transferred to a second wind tunnel and WBF was measured stroboscopically. After the WBF measurement, the locust was transferred back to the original wind stream. Measurements were taken at 30 s, 2 min, and every 5 min thereafter until the 60-min flight time was complete.
Thermosensitivity of Intact Flight
Four separate repetitions of the same experiment were undertaken to test the effect of heat shock on the upper temperature limit for, and thermosensitivity of, tethered flight. For each experiment, 10 locusts were heat shocked as already described and 10 served as the control group. Individual locusts were tethered and flown in an air- stream whose temperature could be varied. For two rep- etitions of the experiment, female locusts were used and the airstream was provided by a 1600 W hair dryer with two heater settings (as well as a setting without heat). The hair dryer was clamped to a retort stand 40 cm from the head of the locust (the airstream was turbulent and at a higher speed than that provided by the wind tunnel, approximately 8 m/s). For the other two repetitions of the experiment, male locusts were used and the airstream was provided by a 1500 W fan heater with 3 heater set- tings (as well as a setting without heat). In these two rep- etitions, the animal was located 6 cm from the fan heater so that it experienced a wind speed of 3 m/s at its head (measured at the position of its head with a hot-wire anemometer). No difference in the results obtained us- ing either of the two methods was apparent. Locusts seemed undisturbed by either method of wind stimula- tion and sustained flight for the duration of the experi- ments. The wingbeats appeared normal and WBFs, monitored electromyographically, before heat stimula- tion were within the normal range of values obtained from experiments using the wind tunnel. Thoracic tem- perature was monitored using a Sensortek thermometer (model BAT- 12) with a copper/constantan thermocou- ple inserted through the arthodial membrane at the base of the right metathoracic leg and directed anteriorly 4 mm. Room temperature was 22" to 24°C and thoracic temperature during flight without a heat stimulus was 25" to 28°C. Turning on each heating element of the hair dryer or fan heater raised thoracic temperature by 5" to 10°C. These experiments were completed within the 10 min before WBF decreases during sustained flight se- quences (Foster and Robertson, 1992).
370 Robertson et a1
EMG signals and a voltage proportional to the tho- racic temperature provided by the BAT- 12 thermometer were stored on videotape (using a Neurocorder model DR-890) for subsequent digitizing and analysis using mi- crocomputer software (Experimenter's Workbench, Da- tawave Technologies, Longmont, CO).
A single brief puff of air to the head was used to reini- tiate flight in those locusts that stopped flying when the temperature was initially raised. About half the locusts stopped flying and there was no difference between heat- shocked and control animals in this regard. Our criterion for heat-induced failure of flight was that, after flight had ceased at high temperatures, three similar puffs of air to the head could not restart flight. Flight was defined as apparently full amplitude wingbeating, at an appropriate frequency (approximately the frequency just before flight stopped), and sustained for at least 5 s (around 100 wingbeats).
From the original 10 animals in each group, we se- lected only animals that appeared vigorous after the heat- shock treatment and recovery. For some animals, it was not possible to obtain a sufficiently clean EMG signal for the computer analysis. Animals that were reluctant to fly were discarded. Data from animals with WBFs below 16 Hz before heat stimulation were also discarded. There was no difference between the results obtained from the four repetitions of the experiment; therefore, the data were pooled. Thirteen control animals and 19 heat- shocked animals met all of the criteria and provided the dataset for comparison of the upper temperature limit for tethered flight.
EMG and temperature records from each animal were analyzed in the following way. At the time of occur- rence of an EMG potential, WBF was computed as the reciprocal ofthe period between that EMG potential and the previous one. At the same time, a reading ofthe tem- perature monitor was taken. One 500 s trial thus gave approximately 10,000 simultaneous data points for WBF and thoracic temperature. It proved unmanageable to use all of these data points to determine the relation- ship between thoracic temperature and WBF. Further- more, thoracic temperature was relatively stable between the times the heater elements were turned on. Therefore, 100 data points (from about 5 s of flight) of WBF and temperature, taken just before each temperature transi- tion, were averaged to provide a mean WBF at three or four different thoracic temperatures for each animal. Only the data from animals with at least three different thoracic temperatures were included in subsequent com- parisons. Eleven control animals and 17 heat-shocked animals provided the dataset for comparison of the ther- mosensitivity of tethered WBF.
Central Flight Rhythm Frequency Measurement
A standard deafferented flight preparation of the locust (Robertson and Pearson, 1982) was used. The wings and
legs were removed and the locust was pinned dorsal sur- face uppermost to a cork board. The thorax and anterior portion of the abdomen was opened with a dorsal, mid- line incision and the thoracic nervous system was ex- posed by removing the gut and the overlying tissue. Usu- ally nerves 1 and 3 (Campbell, 196 1 ) were cut to reduce sensory input and cross-talk of muscle activity. The prep- aration was superfused with locust saline (Robertson and Pearson, 1982). Saline temperature was controlled with a heating coil around a glass pipette that directed the sa- line into the thoracic cavity. A copper/constantan ther- mocouple was placed next to the mesothoracic ganglion to monitor thoracic temperature.
Flight sequences were initiated by blowing on the head of the animal. To obtain a monitor of the centrally generated flight rhythm, an EMG electrode was inserted into a subalar ( M 129) or a dorsal longitudinal (M 1 12) muscle of the hindwing (Snodgrass, 1929). The rhythm frequency was measured as an average of the instanta- neous frequency (reciprocal of EMG period) of 10 con- secutive cycles in four flight sequences. Intracellular re- cordings were made using 1 M potassium acetate filled glass microelectrodes (resistance around 40 MR meg- ohms) inserted into the neuropilar processes of flight motor neurons in the mesothoracic and metathoracic ganglia. All recordings were stored on videotape for sub- sequent analysis.
Thermosensitivity of the Central Flight Rhythm
Twenty-seven 2-week-old adult locusts ( 14 control and 13 heat shocked) were used to test the effect of heat shock on thermosensitivity of deafferented flight rhythm fre- quency in the range from 20°C to around 50"C, and the upper temperature limit for the generation of central flight rhythms. For each animal, the thoracic tempera- ture was gradually raised until flight rhythms as mea- sured by the depressor muscle EMG could no longer be induced by a wind stimulus delivered to the head of the animal. Central rhythm frequencies were measured from the depressor muscle EMG trace obtained during short deafferented flight sequences. Thoracic temperature was manipulated as already described and the preparation was allowed to stabilize to each new temperature for I min so that the thoracic ganglia and surrounding tissues could equilibrate. Each preparation was exposed to at least three temperatures. For technical reasons, it was not possible to raise the temperature by small enough incre- ments to get an exact measure of the temperature at which rhythm generation fails. Thus, the upper temper- ature limit reported is the mean value of the highest tho- racic temperature that permitted flight sequences and the next highest thoracic temperature at which no flight rhythms could be induced. Rhythm frequency immedi- ately before failure is reported as that frequency that was measured at the highest recorded thoracic temperature permitting flight sequences.
Flight of Hat-Shocked Locusts 371
n
S! 80 - * 60 - 9 40 -
W
d)
2 3
LA il n
S! 80 -
* 60 -
9 40 -
W
0
2 - LA
- 0 I
ocked
P 1 day 1 week 2 weeks3 weeks 1 day 1 week 2 weeks3 weeks
Figure 1 Heat-shock-induced thermotolerance. Comparison of survival rates at the lethal temperature of 53°C for groups of control and heat-shocked locusts. Locusts at four different stages of maturation ( 1 day, 1 week, 2 weeks, and 3 weeks old) were heat shocked. Thermo- tolerance was tested: (A) 1 h after heat shock; (B) at least 1 week after heat shock (2 weeks after for 1 -day-old locusts). Survivors were counted after half of the tested animals appeared dead (see text for details).
In 10 animals intracellular recordings were taken from flight rnotoneurons to test whether the failure mea- sured at the periphery was mirrored by a failure of pat- terning within the central circuitry.
In addition to the experiments described, 16 adult lo- custs at least 2 weeks past the adult moult were used to test the effect of heat shock on the thermosensitivity of rhythm frequency in the 20" to 35°C range. The control group contained seven, and the heat-shocked group con- tained nine locusts.
Statistics
Statistical analysis was performed using a software pack- age for microcomputers (Sigmastat, Jandel Scientific, Corte Madera, CA). Data were tested for normality and equal variance and appropriate parametric or nonpara- metric tests were applied. Data from the thermosensitiv- ity experiments were fit with least-squares linear regres- sions because it has been found previously that linear re- gressions give the best fits for the relationships between temperature and the rates of neural phenomena in poi- kilotherms (Walker, 1975) and WBF in insects (May, 198 1 ), including L. rnigrutoriu (Foster and Robertson, 1992). Significance was assessed a t p < 0.05.
periment was terminated when approximately half of the animals in the container appeared dead or moribund. Interestingly, it was the 1 -day-old adult locusts that required 3 h to reach this criterion, whereas the mature adults needed only 1.5 h. When the thermotolerance test was performed 1 h after the exposure to heat-shock conditions, locusts of all ages apart from the 1-day-olds showed in- creased survival rates [ Fig. 1 ( A ) ] . When the ther- motolerance test was performed after allowing the animals to recover for at least 1 week, only animals that were 2 weeks old at the time of heat shock failed to show an increased survival rate [Fig. 1 (B)] . When survival rates were pooled without regard to the age of the animal, or the time of the thermotolerance test, there was a significant difference ( t test: t = -4.25; p < 0.001; d f = 14) between the survival of control animals (mean and standard error; 40.8 2 7.1% survival) and heat- shocked animals (80.9 f 6.3% survival). These data are used only to confirm that the heat-shock conditions reported in a previous study ( Whyard et al., 1986), and used here, were sufficient to induce thermotolerance with our animals and techniques.
RESULTS Heat Shock and Initial WBF
Thermotolerance
Locusts that were exposed to 45°C for 3 h showed improved chances of survival when subsequently exposed to 53°C for 1.5 to 3 h (Fig. 1 ). The variable length of exposure to 53°C resulted because the ex-
There was no significant effect of the heat-shock treatment on the WBF within the first minute of flight for any age of locust tested daily up to 6 or 12 days ( I-day-old at time of heat shock) after the treatment (Fig. 2) . Paired t tests compared the
372 Robertson rt al
A B
1 Week Old Locusts 30 1 1 Day Old Locusts
30 1 n
3 ;E/ 20
C
15
A I
w control I 0 heat shocked
10 I I I I I I I 1 10 0 2 4 6 8 1 0 1 2 1 4 0 1 2 3 4 5 6 7
D
3 Week Old Locusts 30 1 2 Week Old Locusts
30 1
15
2o 1 10 I, 10 I,
0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7
Time (days) Time (days) Figure 2 Heat shock did not affect initial WBF at any maturational stage for at least 1 week after treatment. In locusts that were heat shocked I day ( A ) , 1 week (B) 2 weeks (C), and 3 weeks ( D ) after the imaginal moult, WBF was measured within the first minute of tethered flight, before and after treatment, and every day thereafter for at least 1 week. Data points are mean * SE. The two values at time 0 were taken before and after heat shock.
means of WBF on each test day for heat-shocked and control groups for 1-day-old locusts ( t = - 1.194; p = 0.26; df = 9) , 1-week-old locusts ( t = - 1 . 6 5 ; ~ = 0.14; df= 7), and2-week-oldlocusts ( t = -0.36; p = 0.73; df= 7). The differences be- tween the mean values for the 3-week-old locusts were not normally distributed ( Kolmogorov-Smir- nov test, p < 0.05) and a Wilcoxon signed rank test showed no significant difference for this group ( W
WBF change reflecting the maturation of the flight system, which takes approximately 2 weeks to complete (Kutsch, 1974). For example the WBF of 1-day-old locusts increased daily over the 13 days following the treatment [Fig. 2( A ) ] . Thus, these data also demonstrate that there is no obvious effect of the treatment on maturation of the flight system when it is assessed using this parameter.
- -~
Heat Shock and Maintained WBF = -20, p = 0.20). The animals used in these exper- iments were subsequently used to obtain the sur- vival data displayed in Figure 1 ( B) , confirming that thermotolerance had been induced. Locusts of different ages showed a different time course of
During a flight sequence, WBF normally increases to a maximum within 5 to 10 min before settling gradually to a lower sustained WBF characteristic
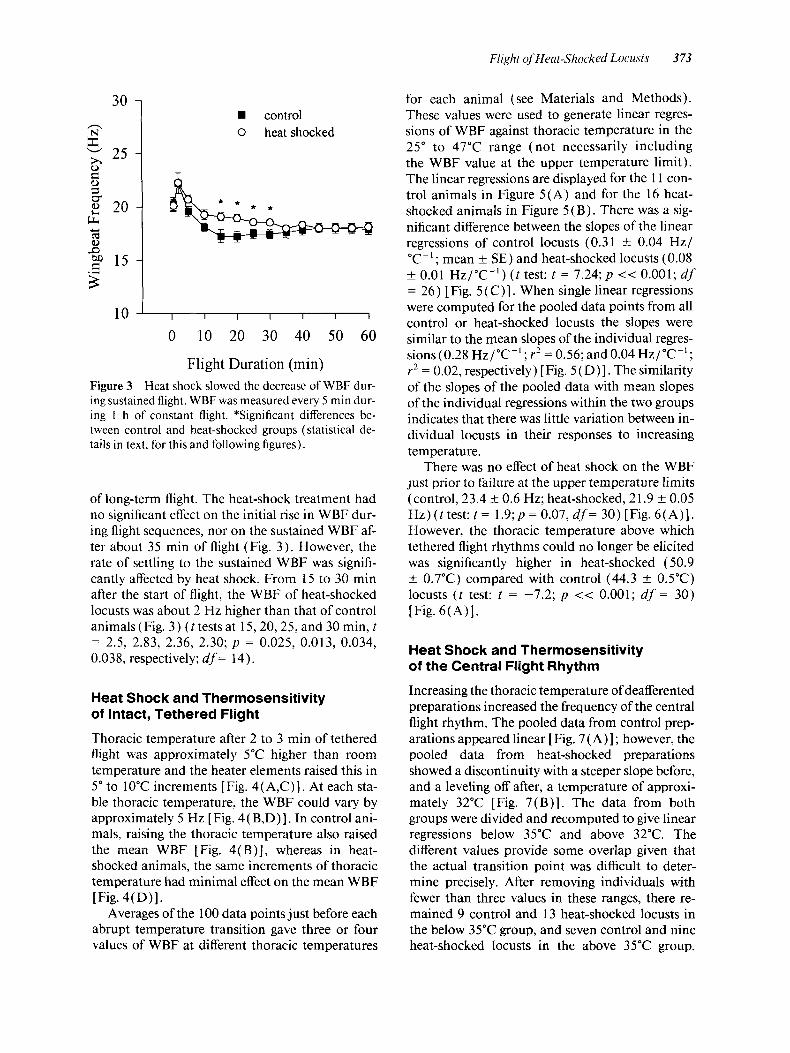
Flight of Heat-Shocked Locusts 373
30 - n
8 - 25 - 5
2 2 0 - f m c4 (d 0) P
C
2 1 5 - s
control 0 heat shocked
10 i 0 10 20 30 40 50 60
Flight Duration (min) Figure 3 Heat shock slowed the decrease of WBF dur- ing sustained flight. WBF was measured every 5 rnin dur- ing 1 h of constant flight. *Significant differences be- tween control and heat-shocked groups (statistical de- tails in text, for this and following figures).
of long-term flight. The heat-shock treatment had no significant effect on the initial rise in WBF dur- ing flight sequences, nor on the sustained WBF af- ter about 35 min of flight (Fig. 3 ) . However, the rate of settling to the sustained WBF was signifi- cantly affected by heat shock. From 15 to 30 rnin after the start of flight, the WBF of heat-shocked locusts was about 2 Hz higher than that of control animals (Fig. 3) ( t tests at 15,20,25, and 30 min, t = 2.5, 2.83, 2.36, 2.30; p = 0.025, 0.013, 0.034, 0.038, respectively; d f = 14).
Heat Shock and Thermosensitivity of intact, Tethered Flight
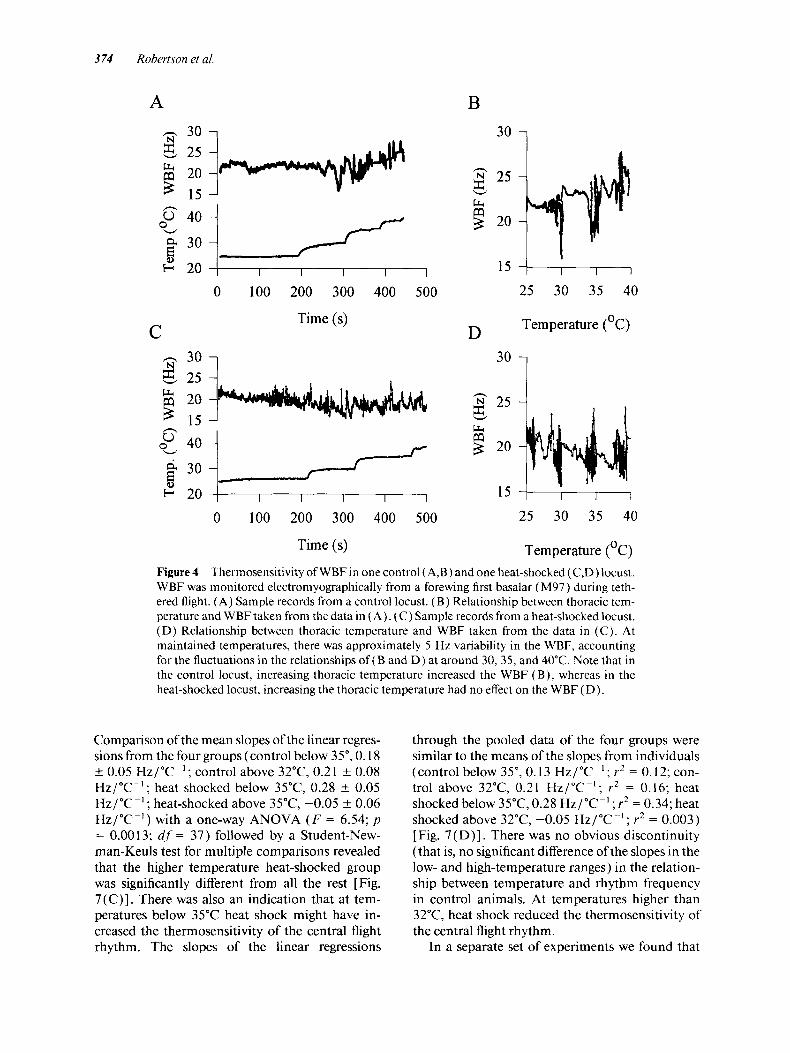
Thoracic temperature after 2 to 3 rnin of tethered flight was approximately 5°C higher than room temperature and the heater elements raised this in 5" to 10°C increments [Fig. 4( A,C)]. At each sta- ble thoracic temperature, the WBF could vary by approximately 5 Hz [Fig. 4( B,D)] . In control ani- mals, raising the thoracic temperature also raised the mean WBF [Fig. 4(B)], whereas in heat- shocked animals, the same increments of thoracic temperature had minimal effect on the mean WBF [Fig. 4(D)].
Averages of the 100 data points just before each abrupt temperature transition gave three or four values of WBF at different thoracic temperatures
for each animal (see Materials and Methods). These values were used to generate linear regres- sions of WBF against thoracic temperature in the 25" to 47°C range (not necessarily including the WBF value at the upper temperature limit). The linear regressions are displayed for the 1 1 con- trol animals in Figure 5 ( A ) and for the 16 heat- shocked animals in Figure 5 ( B). There was a sig- nificant difference between the slopes of the linear regressions of control locusts (0.31 5 0.04 Hz/ "C-' ; mean SE) and heat-shocked locusts (0.08
0.01 Hz/"C-') ( t test: t = 7.24; p << 0.001; df = 26) [Fig. 5 (C)]. When single linear regressions were computed for the pooled data points from all control or heat-shocked locusts the slopes were similar to the mean slopes of the individual regres- sions(0.28 Hz/"C-'; r2 = 0.56;andO.O4Hz/"C-'; r2 = 0.02, respectively) [Fig. 5 ( D ) ] . The similarity of the slopes of the pooled data with mean slopes of the individual regressions within the two groups indicates that there was little variation between in- dividual locusts in their responses to increasing temperature.
There was no effect of heat shock on the WBF just prior to failure at the upper temperature limits (control, 23.4 k 0.6 Hz; heat-shocked, 2 1.9 k 0.05 Hz) ( t test: t = 1.9; p = 0.07, df = 30) [Fig. 6( A)]. However, the thoracic temperature above which tethered flight rhythms could no longer be elicited was significantly higher in heat-shocked ( 50.9 k 0.7"C) compared with control (44.3 k 0.5"C) locusts ( t test: t = -7.2; p << 0.001; d f = 30) [Fig. 6(A)].
Heat Shock and Thermosensitivity of the Central Flight Rhythm
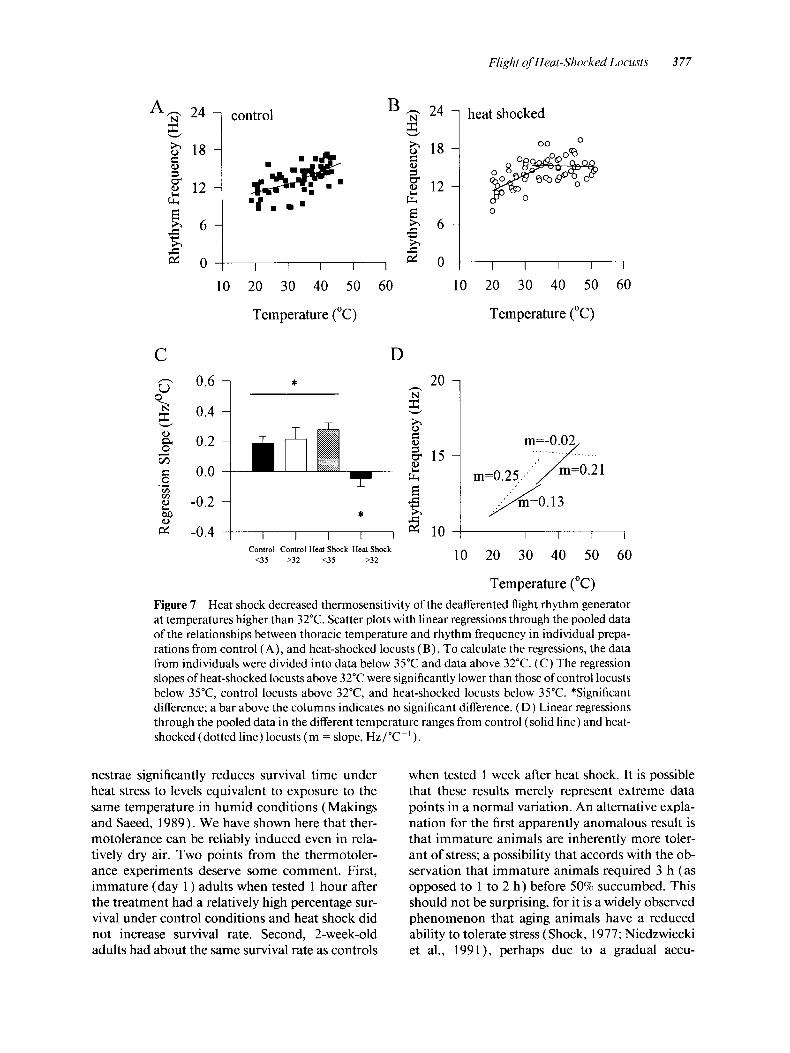
Increasing the thoracic temperature of deafferented preparations increased the frequency of the central flight rhythm. The pooled data from control prep- arations appeared linear [ Fig. 7 (A)] ; however, the pooled data from heat-shocked preparations showed a discontinuity with a steeper slope before, and a leveling off after, a temperature of approxi- mately 32°C [Fig. 7(B)]. The data from both groups were divided and recomputed to give linear regressions below 35°C and above 32°C. The different values provide some overlap given that the actual transition point was difficult to deter- mine precisely. After removing individuals with fewer than three values in these ranges, there re- mained 9 control and 13 heat-shocked locusts in the below 35°C group, and seven control and nine heat-shocked locusts in the above 35°C group.
374 Robertson el al
A B n 30 30
25
20
15
0 100 200 300 400 500 25 30 35 40
C Time (s)
N 25
Temperature (OC) D
30 1 h
25 W
g 20
15
0 100 200 300 400 500 25 30 35 40
Time (s) Temperature (OC) Figure 4 Thermosensitivity of WBF in one control ( A,B) and one heat-shocked (C,D) locust. WBF was monitored electromyographically from a forewing first basalar (M97) during teth- ered flight. ( A ) Sample records from a control locust. ( B ) Relationship between thoracic tem- perature and WBF taken from the data in ( A ) . (C) Sample records from a heat-shocked locust. ( D ) Relationship between thoracic temperature and WBF taken from the data in (C) . At maintained temperatures, there was approximately 5 Hz variability in the WBF, accounting for the fluctuations in the relationships of ( B and D) at around 30, 35, and 40°C. Note that in the control locust, increasing thoracic temperature increased the WBF ( B), whereas in the heat-shocked locust, increasing the thoracic temperature had no effect on the WBF (D).
Comparison of the mean slopes of the linear regres- sions from the four groups (control below 35", 0.18 -+ 0.05 Hz/"C-'; control above 32"C, 0.21 2 0.08 Hz/"C-'; heat shocked below 35"C, 0.28 f 0.05 Hz/"C-I; heat-shocked above 35T, -0.05 k 0.06 Hz/"C-') with a one-way ANOVA ( F = 6.54; p = 0.001 3; df = 37) followed by a Student-New- man-Keuls test for multiple comparisons revealed that the higher temperature heat-shocked group was significantly different from all the rest [Fig. 7 (C)] . There was also an indication that at tem- peratures below 35°C heat shock might have in- creased the thermosensitivity of the central flight rhythm. The slopes of the linear regressions
through the pooled data of the four groups were similar to the means of the slopes from individuals (control below 35", 0.13 Hz/"C-'; r2 = 0.12; con- trol above 32"C, 0.21 Hz/"C-'; r 2 = 0.16; heat shocked below 35T, 0.28 Hz/"C-'; r2 = 0.34; heat shocked above 32"C, -0.05 Hz/"C-'; r2 = 0.003) [Fig. 7 (D)] . There was no obvious discontinuity (that is, no significant difference of the slopes in the low- and high-temperature ranges) in the relation- ship between temperature and rhythm frequency in control animals. At temperatures higher than 32"C, heat shock reduced the thermosensitivity of the central flight rhythm.
In a separate set of experiments we found that
Flight of Heat-Shocked Locusts 375
30
25
20
15
0.5
0.4
0.3
0.2
0.1
0.0
control B
20 30 40
Temperature (OC)
T
50
D
n N z, B
-- -
Control Heat Shock
30
25
20
15
20
30
25
20
15
heat shocked
30 40 50
Temperature (OC)
m=0.28
20 30 40 50
Temperature (OC) Figure 5 Heat shock decreased the thermosensitivity of WBF during tethered flight. Linear regressions of the relationships between thoracic temperature and WBF in individual control ( A ) and heat-shocked locusts (B) . (C) The regression slopes of heat-shocked locusts were sig- nificantly less than those of control locusts. *Significant difference. (D) Linear regressions through the pooled data from control (solid line) and heat-shocked (dotted line) locusts (m = slope, Hz/"C-').
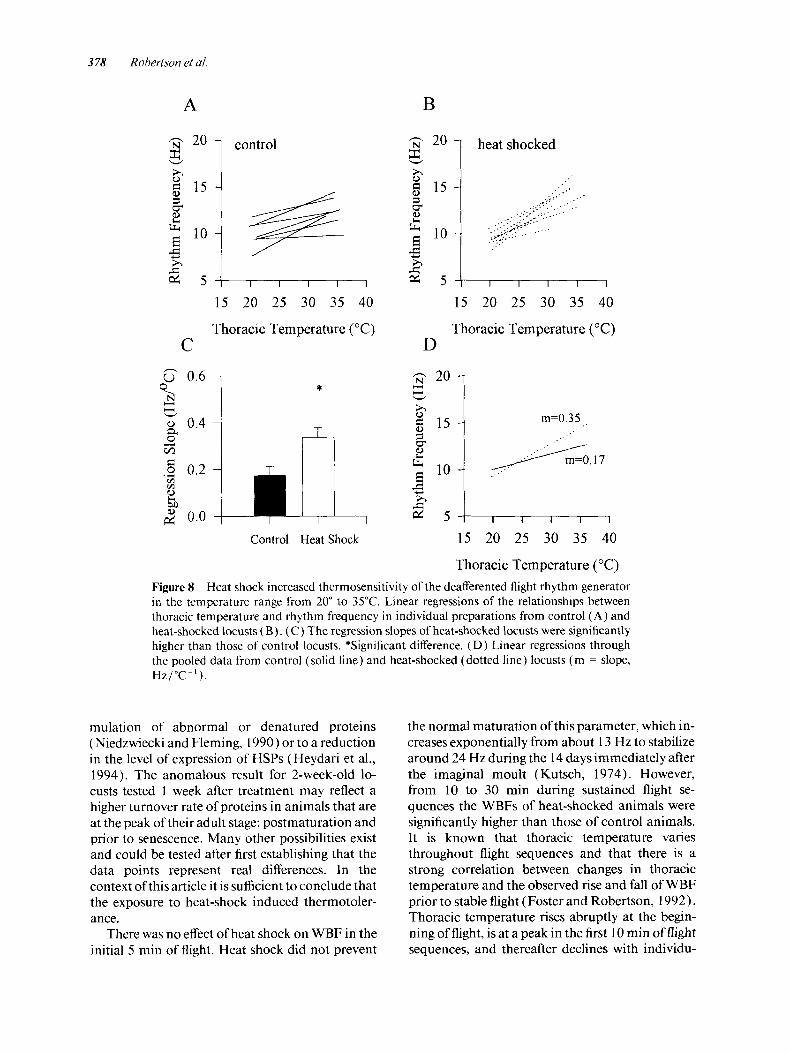
increases in thoracic temperature within the tem- perature range of 20" to 35°C increased the fre- quency of the central flight rhythm [Fig. 8 (A)]. The data from individual preparations were fitted with linear regressions [Fig. 8(A,B)], and the mean slopes of these regressions were significantly different in the control and heat-shocked groups ( t test: t = -2.82; p = 0.014; df = 14) [Fig. 8(C)]. The heat-shocked group was more sensitive to in- creasing temperature (mean slope, 0.34 Hz/"C-') than the control group (mean slope, 0.17 Hz/ OC-I). When the data from individual prepara- tions were pooled and fitted with a single linear re- gression, the slope of the control group was 0.17 Hz/"C-' ( r 2 = 0.32), and the slope of the heat-
shocked group was 0.35 Hz/"C-' ( r2 = 0.75) [ Fig. 8( D)] . At temperatures lower than 35"C, heat shock increased the thermosensitivity of the central flight rhythm.
The rhythm frequency expressed by deaffer- ented preparations just before heat-induced failure was not significantly different in heat-shocked and control groups (control, 14.62 f 0.50 Hz; heat shocked, 15.36 f 0.58 Hz) ( t test: t = 0.98; p = 0.34; df= 25) [Fig. 9(A)]. The heat-shock treat- ment significantly increased the upper temperature limit for the production of central flight rhythms ( t test: t = 3.7; p = 0.001; df= 25) [Fig. 9 (3 ) ] . Preparations in the control group failed at mean temperature of 42.59 * 0.85"C, whereas prepara-
376 Robertson et al
A 30
25 n
W 4
E 2o 15
10
B 5 5
9 50
35
*
T
-r- 1 Control Heat Shock Control Heat Shock
Figure 6 Heat shock had no effect on the WBF at heat-induced inability to fly but increased the upper temperature limit at which tethered flight failed. ( A ) The WBF at heat-induced failure was the same in control and heat-shocked locusts (bar above columns indicates no significant difference). ( B ) The mean temperature at which tethered flight failed was 6.6"C higher in heat-shocked locusts. *Significant difference.
tions in the heat-shocked group failed at a mean temperature of 47.64 f 1.06"C.
Failure to produce flight rhythms that are mon- itored at the periphery could result from a failure of communication from the head to the thoracic nervous system or from there to the muscle im- planted with an EMG electrode. Ten preliminary intracellular investigations showed that failure in both control and heat-shocked preparations was associated with a failure of neural patterning within the central nervous system (Fig. 10). Synaptic effects of blowing wind on the head persisted; small excitatory postsynaptic potentials were evident and motor neurons could still generate action poten- tials ( intracellular and dorsal longitudinal traces at 52°C in Fig. 10). However, the central flight cir- cuitry was incapable of rhythm generation.
DISCUSSION
In an environment where ground temperatures in unshaded areas can reach as high as 80°C (Hardy, 1979) and in which gregarious L. migratoria hop- pers sitting in the sun can achieve an internal tem- perature of 45"C, while the air temperature is only 30°C ( Uvarov, 1966 ) , it is a reasonable expectation that adult L. migratoria will experience heat stress at some point in their lives. Moreover, heat treat- ments of only 39°C are sufficient to induce thermo- tolerance (Whyard et a]., 1986), which is a charac-
teristic physiological response to heat stress. It is equally reasonable to expect that heat stress will be more likely during times of increased activity, such as during flight, when working muscles generate heat that may not be dissipated rapidly or effectively. It has been established that I,. migruto- ria exhibit a classic heat-shock response with the expression of HSPs, and concurrent induction of thermotolerance ( Whyard et al., 1986). The focus of this article is the expression of flight motor pat- terns in intact and deafferented locusts after exposure to heat-shock conditions. We were inter- ested in whether the induction of thermotolerance had adaptive consequences for nervous system function and for the ability to behave at higher temperatures as well as simply increasing the upper lethal temperature.
Our initial experiments confirmed the results of Whyard et a]. ( 1986) and showed that thermotol- erance could be induced by a prior exposure to an extreme, but sublethal temperature (45°C) for 3 h. Exposure to heat shock doubled the survival rate of locusts that were subsequently exposed to a nor- mally lethal temperature. In the experiments of Whyard et al. the humidity of the heat-shock envi- ronment was maintained at 50% using a water bath. This would have had the effect of compro- mising the mechanisms of heat loss via evaporative water loss (Makings, 1987) through the fenestrae in the abdominal and thoracic cuticle [Slifer's patches; Slifer ( 195 1 )] . In dry air, blocking the fe-
Flight of Heat-Shocked Locusts 377
*a 2 4 , W
C n x a a 0 m *
control
1
8 12
c4 Ox
heat shocked
00 O
0
I I I I 1
10 20 30 40 50 60 10 20 30 40 50 60
Temperature ("C) Temperature ("C)
0.6
0.4
0.2
0.0
-0.2
-0.4
* 1 1 -
D
m=-0.02 . . . . . . . . ../ .-.-.......
m=O .2 5. : ' . m=0.21
b 0 . 1 3
10 20 30 40 50 60
Temperature ("C)
Control Control Heat Shock Heat Shock <35 >32 <35 >32
Figure 7 Heat shock decreased thermosensitivity of the deafferented flight rhythm generator at temperatures higher than 32°C. Scatter plots with linear regressions through the pooled data of the relationships between thoracic temperature and rhythm frequency in individual prepa- rations from control ( A ) , and heat-shocked locusts ( B ) . To calculate the regressions, the data from individuals were divided into data below 35°C and data above 32°C. (C) The regression slopes of heat-shocked locusts above 32°C were significantly lower than those of control locusts below 35"C, control locusts above 32T, and heat-shocked locusts below 35°C. *Significant difference; a bar above the columns indicates no significant difference. (D) Linear regressions through the pooled data in the different temperature ranges from control (solid line) and heat- shocked (dotted line) locusts (m = slope, Hz/"C-').
nestrae significantly reduces survival time under heat stress to levels equivalent to exposure to the same temperature in humid conditions (Makings and Saeed, 1989). We have shown here that ther- motolerance can be reliably induced even in rela- tively dry air. Two points from the thermotoler- ance experiments deserve some comment. First, immature (day 1 ) adults when tested 1 hour after the treatment had a relatively high percentage sur- vival under control conditions and heat shock did not increase survival rate. Second, 2-week-old adults had about the same survival rate as controls
when tested 1 week after heat shock. It is possible that these results merely represent extreme data points in a normal variation. An alternative expla- nation for the first apparently anomalous result is that immature animals are inherently more toler- ant of stress; a possibility that accords with the ob- servation that immature animals required 3 h (as opposed to 1 to 2 h) before 50% succumbed. This should not be surprising, for it is a widely observed phenomenon that aging animals have a reduced ability to tolerate stress (Shock, 1977; Niedzwiecki et al., 1991), perhaps due to a gradual accu-
378 Robertson et a/.
A
I I I I 1
15 20 25 30 35 40
Thoracic Temperature ("C) C 0.6
0 -a z P) 0.4 W
8 .I 0.2
ko d" 0.0
- m
cn cn P)
T
*
T
I I Control Heat Shock
heat shocked
15 20 25 30 35 40
Thoracic Temperature ("C)
m=0.35 .
/ m=O. 17
I I I I I
15 20 25 30 35 40
Thoracic Temperature ("C) Figure 8 Heat shock increased thermosensitivity of the deafferented flight rhythm generator in the temperature range from 20" to 35°C. Linear regressions of the relationships between thoracic temperature and rhythm frequency in individual preparations from control ( A ) and heat-shocked locusts ( B). (C) The regression slopes of heat-shocked locusts were significantly higher than those of control locusts. *Significant difference. (D) Linear regressions through the pooled data from control (solid line) and heat-shocked (dotted line) locusts ( m = slope, Hz/"C-' ).
mulation of abnormal or denatured proteins (Niedzwiecki and Fleming, 1990) or to a reduction in the level of expression of HSPs (Heydari et al., 1994). The anomalous result for 2-week-old lo- custs tested 1 week after treatment may reflect a higher turnover rate of proteins in animals that are at the peak of their adult stage: postmaturation and prior to senescence. Many other possibilities exist and could be tested after first establishing that the data points represent real differences. In the context ofthis article it is sufficient to conclude that the exposure to heat-shock induced thermotoler- ance.
There was no effect of heat shock on WBF in the initial 5 min of flight. Heat shock did not prevent
the normal maturation of this parameter, which in- creases exponentially from about 13 Hz to stabilize around 24 Hz during the 14 days immediately after the imaginal moult (Kutsch, 1974). However, from 10 to 30 min during sustained flight se- quences the WBFs of heat-shocked animals were significantly higher than those of control animals. It is known that thoracic temperature vanes throughout flight sequences and that there is a strong correlation between changes in thoracic temperature and the observed rise and fall of WBF prior to stable flight (Foster and Robertson, 1992). Thoracic temperature rises abruptly at the begin- ning of flight, is at a peak in the first 10 min of flight sequences, and thereafter declines with individu-
Flight of'Heut-Shocked Locusts 379
5 .
I
I
T
I
B 55
0 50 O-
5 45 a
t-r 40
I 35 Control Heat Shock
T
* T
r
Control Heat Shock Figure 9 Heat shock had no effect on the rhythm frequency at failure but increased the upper temperature limit at which deafferented flight rhythms could no longer be generated. ( A ) The rhythm frequencies at heat-induced failure were the same in control and heat-shocked locusts (bar above columns indicates no significant difference). (B) The mean temperature at which deafferented flight rhythms failed to be induced was 5°C higher in heat-shocked locusts. *Sig- nificant difference.
ally varying rates to a stable level (Foster and Rob- ertson, 1992). Our preliminary interpretation of the above finding is that the difference in WBF from 15 to 30 min during sustained flight in control and heat-shocked animals reflects the changed thermosensitivity of the flight system after heat shock (see later). The drop in thoracic temperature after 10 min offlight (Foster and Robertson, 1992) may reflect the spread of heat to the abdomen and its subsequent dissipation from the abdominal fe- nestrae. Control animals, being more thermosensi- tive, may have been affected to a greater extent by this loss of heat. This idea could be tested by exam- ining the effect of blocking the abdominal fenes- trae.
The most interesting findings of this study relate to the thermosensitivity ofthe flight system. In con- trol animals the relationship between thoracic tem- perature and WBF appeared linear with a slope of 0.28 Hz/"C-'. This confirms previous work that showed that between 25" and 36°C there is a slope of around 0.3 Hz/"C-' in the linear relationship (Foster and Robertson, 1992). After heat shock, the relationship changed so that in the temperature range of 25" to 47°C the slope decreased to 0.04 Hz /"C -' and the WBF appeared temperature com- pensated (that is, thoracic temperature had mini- mal effect on WBF). It appears, therefore, that heat shock can induce the flight system to be relatively insensitive to changes in thoracic temperature
around its normal operating temperature. This is particularly interesting. Initial studies, using the desert locust (Schistocercu greguria), on the tem- perature sensitivity of WBF demonstrated inde- pendence from air temperature in the 25" to 35°C range (Weis-Fogh, 1956), which would result in thoracic temperatures in the 32" to 42°C range. Pre- viously, it was proposed that the difference with the results obtained from the migratory locust ( L. rnigratoria) may have been a species-specific difference. With the findings that there are species- specific differences in the heat-shock response, de- pending on the thermal ecology of the species, and that certain species normally express basal levels of HSPs or their cognates (Ulmasov et al., 1992; Gehring and Wehner, 1995), this proposal be- comes more compelling. Alternatively, if the ex- perimental animals had experienced high body temperatures, either as a result of conditions in the laboratory colony or as a result of a prior flight ex- periment, then it is possible that their flight systems had already been conditioned for operation with temperature compensation. Furthermore, animals taken from the wild for experiments would be likely to have preconditioned flight systems.
It is abundantly clear from the work reported here that heat shock extends the mean upper tem- perature limit for flight motor pattern generation in intact animals by 6.6"C. Some individual ani- mals were still capable of generating flight motor
380 Robertson el al.
E 23 "C
. . 37°C
Figure 10 Heat-induced failure to generate deaffer- ented flight rhythms was associated with failure of the flight central pattern generator. Intracellular recordings from a mesothoracic elevator motoneuron (E) of a heat- shocked locust during wind-induced flight rhythms at temperatures ranging from 23" to 5 1°C. The time of de- pression in the rhythm was monitored with an electro- myographic electrode in the dorsal longitudinal muscle (DL). Note that at 52°C a flight rhythm could not be generated, although wind-evoked postsynaptic poten- tials were still evident and the dorsal longitudinal moto- neurons could still generate action potentials.
patterns with thoracic temperatures of 52°C and higher. We do not yet have any information about how long wingbeating could be sustained at these high temperatures, or whether force production by the wing muscles is compromised, thus preventing flight. Nevertheless, it is difficult to think that this is anything other than an adaptive effect of heat shock to enable locusts to continue flying in their harsh environment. Insect flight muscles require high operating temperatures for optimal perfor- mance (Josephson, 198 1, Stevenson and Joseph- son, 1990). In honeybees the peak of force produc- tion is attained at thoracic temperatures around 40°C and declines to zero around 50°C (Coelho, 199 1 ). In insects average thoracic temperatures during flight are often maintained around 40°C (Heinrich, 198 1 ) . These high normal operating
temperatures are close to the lethal core tempera- tures of many other animals ( Schmidt-Neilsen, 1990). Furthermore, during flight in hot condi- tions in the field, thoracic temperatures in insects have been recorded as high as 49°C (Chappell, 1982, 1984; Coelho, 199 1 ) without obviously de- bilitating the insects. Given these observations, it would be surprising if insects living in extremely hot conditions did not possess adaptations of their neuromuscular systems that allow them to behave with high body temperatures.
We have demonstrated that the effects of heat shock observed in intact locusts are mirrored in deafferented preparations. In control animals there was a linear relationship between thoracic temper- ature and the frequency of central flight rhythms. In heat-shocked animals there was an increase in the slope of the linear relationships at relatively low temperatures (20" to 35°C). However, above about 32°C the relationship leveled off so that subse- quent increases in temperature had little effect on the frequency of the rhythms. Interestingly, the maximum frequency generated by deafferented preparations was not affected by the heat-shock treatment. We conclude that heat shock can condi- tion the central circuitry to become temperature insensitive at thoracic operating temperatures that are normal for flight. Furthermore, heat shock in- creased the upper temperature limit for the produc- tion of central flight rhythms by 5°C as monitored both in the periphery and by intracellular recording from flight motoneurons. Heat shock enabled cen- tral patterning at higher temperatures. Therefore, it is likely that the adaptive effects of heat shock we observed in intact animals were caused by condi- tioning of the operation of the neuromuscular sys- tem. There is some information available about the effects of temperature on the cellular properties of flight neurons (Xu and Robertson, 1994) and on synaptic interactions in the flight circuitry (Robertson, 1993). It is important now to repeat these experiments in heat-shocked animals to de- termine which neural processes are modified to cause the effects on thermosensitivity we describe here. One possibility is that the levels of neuromod- ulators in the hemolymph are altered by the treat- ment and this could affect operation of the flight circuit. For example, octopamine is known to play a role in coordinating the neural and metabolic re- quirements of flight (Orchard et al., 1993). In par- ticular, treatment with octopamine can release flight rhythms in deafferented preparations ( Som- bati and Hoyle, 1984) and can promote rhyth- micity by inducing the generation of plateau
Flight of’ Heal-Shocked Loc3irst.s 381
potentials in flight neurons (Ramirez and Pear- son, 199 l ). Heat stress promotes an increase in the concentration of circulating octopamine in lo- custs (Davenport and Evans, 1984); however, this could not account for our results with deaffer- ented preparations in which the hemolymph was replaced with superfused saIine. An alternative possibility is that the channel proteins that un- derlie the generation of action potentials and syn- aptic potentials become stabilized in the mem- brane and Iess sensitive to changes in temperature; an effect that may result in a favorable shift of the relationships between synaptic potential parame- ters and temperature.
Although it may be premature to speculate on the role of HSPs in the phenomena described here, and noting that in some instances thermotolerance has been reported to be independent of the synthe- sis and persistence of HSPs (Carretero et al., 199 1 ; Yocum and Denlinger, 1992), we can reasonably infer from previous work using animals from the same colony (Whyard et al., 1986) that our ther- motolerant animals were expressing HSPs. There is a considerable and rapidly expanding literature on HSPs, including studies of the molecular roles of HSPs and descriptions of HSP expression in different tissues after different treatments. How- ever, there is very little information on any possible physiological roles of HSPs. Within nervous tissue, most studies have correlated HSP expression with various pathological conditions and speculated on their possible protective function ( Lowenstein et al., 1991; Rordorf et al., 1991; Simon et al., 1991; Wong et al., 1992; Manzerra and Brown, 1993; Koroshetz and Bonventre, 1994; Passarelli et al., 1994). We believe it to be an exciting possibility that HSPs could be involved in mediating the effects described in this article.
Our general conclusion is that prior heat experi- ence, which may be a normal, inevitable conse- quence of the ecology of the animal, conditions the flight system of L. rnigratoria, including the central circuitry, so that it is able to operate at higher tem- peratures and to do this with minimal temperature sensitivity.
We thank J. W. Dawson, J. R. Gray, and V. K. Walker for critically reviewing a previous version of the manuscript. Supported by grants from the Natural Sci- ences and Engineering Research Council of Canada and the School of Graduate Studies and Research at Queen’s University.
REFERENCES
CAMPBELL, T. I. ( 1961 ). The anatomy of the nervous system of the mesothorax of Locustu migrutoriu mi- grutorioides (R. and F.). Proc. Zool. Soc. (Lond.) 137:
CARRETERO, M. T., CARMONA, M. J., and DIEz, J. L. ( 1991 ). Thermotolerance and heat shock proteins in Chironomus. J. Insect Physiol. 37:239-246.
CHAPPELL, M. A. (1982). Temperature regulation of carpenter bees (Xylocopu calqornicu) foraging in the Colorado desert of southern California. Physiol. Zool. 55267-280.
CHAPPELL, M. A. (1984). Temperature regulation and energetics of the solitary bee Centrispullida during for- aging and intermittent mate competition. Physiol.
COELHO, J. R. ( I99 1 ). The effect of thorax temperature on force production during tethered flight in honeybee (Apis mellfera) drones, workers, and queens. Physiol. Zool. 64:8 2 3 -8 3 5.
CRAIG, E. A,, GAMBILL, B. D., and NELSON, R. J. (1993). Heat shock protein: molecular chaperones of protein biogenesis. Microhiol. Rev. 57:402-4 14.
DAVENPORT, A. P. and EVANS, P. D. (1984). Stress-in- duced changes in the octopamine levels of insect haemolymph. Insect Biochem. 14:135-143.
FOSTER, J. A. and ROBERTSON, R. M. ( 1992). Temper- ature dependency of wing-beat frequency in intact and deafferented locusts. J. E,rp. Biol. 162:295-3 12.
GEHRING, W. J. and WEHNER, R. (1995). Heat shock protein synthesis and thermotolerance in Catuglyphis, an ant from the Sahara desert. Proc. Null. Acad. Sci. USA 92:2994-2998.
GEORGOPOULOS, C. and WELCH, W. J. ( 1993). Role of the major heat shock proteins as molecular chaper- ones. Annu. Rev. Cell Biol. 9:601-634.
GRIMM, K. and SAUER, A. E. ( 1995). The high number of neurons contributes to the robustness of the locust flight-CPG against parameter variation. Biol. Cyhern.
HARDY, R. N. ( 1979). Temperature and Animal Life. University Park Press, Baltimore.
HARRINGTON, H. M., DASH, S., DHARMASIRI, N., and DHARMASIRI, S. (1994). Heat-shock proteins: a search for functions. Aust. J. Plant Physiol. 212343- 855.
HEINRICH, B. ( 198 I ). Ecological and evolutionary per- spectives. In: Insect Thcrmoregulution. B. Heinrich, Ed. John Wiley & Sons, New York, pp. 235-302.
HEYDARLA. R., TAKAHASHI, R.,GUT-SMANN, A., You, S., and RICHARDSON, A. ( 1994). HSP and aging. Ex- perientia 50:1092-1098.
JANSSEN, R. ( 1992). Thermal influences on nervous sys- tem function. Neurosci. Biohehav. Rev. 16:399-4 13.
JOSEPHSON, R. K. ( 1981 ). Temperature and the me- chanical performance of insect muscle. In: Insect Thermoregulation. B. Heinrich, Ed. John Wiley & Sons, New York, pp. 19-44.
403-432.
Zed. 5 7 ~ 2 15-225.
72~329-335.
382 Robertson et ul
KAMPINGA, H. H. (1993). Thermotolerance in mam- malian cells: protein denaturation and aggregation and stress proteins. J. Cell Sci. 104: 1 I - 17.
KOROSHETZ, W. J. and BONVENTRE. J. V. ( 1994). Heat shock response in the central nervous system. Expe- rientia 50: 1085- 109 l.
KUTSCH, W. ( 1974). The development of the flight pat- tern in locusts. In: Experimental Analysis of Insect Be- haviour. L. Barton-Brown, Ed. Springer Verlag, New York, pp. 149-158.
LINDQUIST, S. and CRAIG, E. A. ( 1988). The heat shock proteins. Annu. Rev. Gene[. 22:63 1-677.
LOWENSTEIN, D. H., CHAN, P. H., and MILES, M. P. ( 1991 ). The stress protein response in cultured neu- rons: characterization and evidence for a protective role in excitotoxicity. Neuron 7:1053-1060.
MAKINGS, P. ( 1987 ). Survival value of Slifer’s patches for locusts at a high temperature. J. Insect Physiol. 33:
MAKINGS, P. and SAEED, F. N. ( 1989). Influence of Slif- er’s patches on survival and behaviour of locusts ( Locustu migratoria migrutorioides) at a high temper- ature. J. In.cect P/qaioI. 35:363-368.
MANZERRA, P. and BROWN, 1. R. ( 1993). Temporal and spatial distribution of heat shock mRNA and protein (HSP70) in the rabbit cerebellum in response to hy- perthermia. J . Neurosci. Rex 36:480-490.
MAY, M. L. (1981). Wingstroke frequency of dragon- flies (0donata:Anisoptera) in relation to temperature and body size. J . Comp. Physiol. (‘4) 144:229-240.
MONTGOMERY. J. C. and MACDONALD, J. A. (1990). Effects of temperature on nervous system: implica- tions for behavioral performance. Am. J . Physiol. 259: R I9 I-R 196.
Mostxeu, P. L. ( 1994). Mechanisms of heat adaptation: thermotolerance and acclimatization. J. Lab. Clin.
NEVILLE, A. C. and WEIS-FOGH, T. (1963). The effect of temperature on locust flight muscle. J. Exp. Biol. 40:l 11-121.
NIEDZWIECKI, A. and FLEMING, J. E. (1990). Changes in proteins turnover after heat shock are related to ac- cumulation of abnormal proteins in aging Drosophila m elu n ogust er. Mech. Ageing Dcv. 5 2: 2 9 5 - 304.
NIEDZWIECKI, A,, KONGPACHITH, A. M., and FLEMING, J. E. ( 199 I ). Aging affects expression of 70-kDa heat shock proteins in Drosciphilu. J . Biol. Chem. 266:
OERTLI, J. J. (1989). Relationship of wing beat fre- quency and temperature during take-off flight in tem- perate-zone beetles. J . Exp. Biol. 14532 1-338.
ORCHARD, I. , RAMIREZ, J.-M., and LANCE, A. B. (1993). A multifunctional role for octopamine in lo- cust flight. Annu. Rev. Entomol. 38~227-249.
PASSARELLI, F., AUGELETTI, B., ORRU, D., ORZI, F., and D’AMBROSIO, E. ( 1994). Effect of electroconvul- sive shock on the levels of HSPs70 and HSC73 mRNA in the rat brain. Neurosci. Lett. 177: 147- 150.
8 15-822.
Med. 123~48-52.
9 332-9338.
RAMIREZ, J.-M. and PEAKSON, K. G. ( 1991 ). Octopam- ine induces bursting and plateau potentials in insect neurones. Brain Rex 549:332-337.
RITOSSA, F. ( 1962). A new puffing pattern induced by heat shock and DNP in Drosophila. Experientia 18:
ROBERTSON, R. M. ( 1986). Neuronal circuits control- ling flight in the locust: central generation of the rhythm. Trends Neurosci. 9:278-280.
ROBERTSON, R. M. ( 1989). Idiosyncratic computational units generating innate motor patterns: neurones and circuits in the locust flight system. In: The Computing Nrzirone. R. Durbin. R. C. Miall, and G. Mitchison, Eds. Addison-Wesley, London, pp. 262-277.
ROBERTSON, R. M. (1993). Effects of temperature on synaptic potentials in the locust flight system. J. Neu- rophysiol. 70:2 197-2204.
ROBERTSON, R. M. (1995). Locust flight: components and mechanisms in the motor. In: The Handbook of Bruin Theory und Neural Networks. M. A. Arbib, Ed. MIT Press, Cambridge, MA, pp. 556-560.
ROBERTSON, R. M. and PEARSON, K. G. ( 1982). A prep- aration for the intracellular analysis of neuronal activ- ity during flight in the locust. J . Comp. Physiol. (A)
RORDORF, G., KOROSHETZ, W. J., and BONVENTRE, J. V. ( 1991). Heat shock protects cultured neurons from glutamate toxicity. Neuron 7: 1043- I05 1.
SCHMIDT-NEILSEN, K. ( 1990). Animal Physiology. Ad- uptution und Environment. Cambridge University Press, New York.
SHOCK, N. W. (1977). Systems integration. In: Hand- book of’ the Biology o f ’ Aging. C. E. Finch and L. Hayflick, Eds. Van Nostrand Reinhold, New York,
SIMON, R. P., CHO, H., GWINN, R., and LOWENSTEIN, D. H. ( 1991 ). The temporal profile of 72-kDa heat shock protein expression following global ischemia. J. Neurosci. 11:881-889.
SLIFER, E. H. (1951). Some unusual structures in Lo- czisia migratoria migratorioides and their probable function as thermoreceptors. Proe. R . Soc. Lond. [ B i d ] 138:4 14-437.
SNODCRASS, R. E. ( 1929). The thoracic mechanism ofa grasshopper, and its antecedents. Smithsonian M i x . Coll. 82: 1 - 1 1 1.
SOMBATI, S. and HOYLE, G. ( 1984). Generation ofspe- cific behaviours in a locust by local release into neu- ropil of the natural neuromodulator octopamine. J. Neurobiol. 15:48 1-506.
STEVENSON, R. D. and JOSEPHSON, R. K. (1990). Effects of operating frequency and temperature on me- chanical power output from moth flight muscle. J. Exp. Biol. 149:61-78.
ULMASOV, K. A., SHAMMAKOV, S., KARAEV, K., and EVGEN’EV, M. B. (1992). Heat shock proteins and thermoresistance in lizards. Proc. Natl. Acud. Sci. USA
57 1-573.
146:311-320.
pp. 639-665.
89: 1666- 1670.
Flight QfHeat-Shocked Locusts 383
UVAROV, B. ( 1966). Grasshoppers and Locusts: Anal- omy, Physiology, Development, Phase Polymorphism, Introduction to Taxonomy. Cambridge University Press, London.
WALDRON, I. ( 1968). The mechanism ofcoupling ofthe locust flight oscillator to oscillatory inputs. Z. Vergl. Physiol. 57:331-347.
WALKER, T. J . ( 1975). Effects of temperature on rates in poikilotherm nervous systems: evidence from the calling song of meadow katydids (Orthoptera: Tetti- goniidae: Orchelimum) and reanalysis of published data. J. Comp. Physiol. (A) 10157-69.
WEIS-FOGH, T. (1956). Biology and physics of locust flight-11. Flight performance of the desert locust (Schistocerca gregaria). Phil. Trans. R. SOC. Lond. [Biol.] 239:459-5 10.
WEIS-FOGH, T. (1964). Biology and physics of locust
flight. VIII. Lift and metabolic rate of flying locusts. J. Exp. Bid . 41:257-271.
WHYARD, S., WYATT, G. R., and WALKER, V. K. ( 1986). The heat shock response in Locustu migruto- ria. J. Comp. Physiol. ( B ) 1569 13-8 17.
WONG, M. L., WEISS, S. R. B., GOLD, P. W., Dor, S. Q., BANERIEE, S., LIANIO, J., LAD, R., POST, R. M., and SMITH, M. A. ( 1992). Induction of constitutive heat shock protein 73 mRNA in the dentate gyrus by sei- zures. Mol. Bruin Res. 13:19-25.
Xu, H. and ROBERTSON, R. M. ( 1994). Effects of tern- perature on properties of flight neurons in the locust. J. Comp. Physiol. (A) 175: 193-202.
YOCUM, G. D. and DENLINGER, D. L. (1992) . Induc- tion and decay of thermosensitivity in the flesh fly, Surcophuga crassipalpis. J. Comp. Physiol. (B ) 163: 113-1 17.















![SHOCK[1] - Hypovolemic Shock](https://static.documents.pub/doc/80x56/58edc1bc1a28abae538b4711/shock1-hypovolemic-shock.jpg)














