THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 269, No. 7, Issue of February 18, pp. 5358-5363,1994 Printed in U.S.A. Expression and Characterization of Biologically Active Human Extracellular Superoxide Dismutase in Milk of Transgenic Mice* (Received for publication, June 29, 1993, and in revised form, September 24, 1993) Lennart HanssonSf, Michael EdlundS, Anders EdlundS, Thore JohanssonS, Stefan L. Marklundll, Sigurd FrommII, Mats StromqvistS, and Jan Tornell** From SSymbicom AB, P.O. Box 1451, S-901, 24 Umed, Sweden, the nDepartment of Clinical Chemistry, Umed University Hospital, S-901 85 Umed, Sweden, the IlLaboratory of Molecular Embryology, University of Oslo, P.O. Box 1052 Blindern, N-0316 Oslo 3. Norway, and the **Department of Physiology, University of Goteborg, Medicinaregatan 11, S-413 90 Goteborg, Swiden . - We have targeted the expression of recombinant hu- man extracellular superoxide dismutase, a glycosy- lated tetrameric metalloprotein, to the mammary gland of transgenic mice. This was achieved by using regu- latory elements from either the murine whey acidic protein gene or the ovine &lactoglobulin gene to con- trol expression of humanextracellular superoxide dis- mutase cDNA. Whey acidic protein regulatory se- quences directed high level mammary gland-specific expression of the recombinant gene and secretion of biologically active extracellularsuperoxide dismutase into the milk. The produced recombinant protein was fully active, it was in tetrameric form, it showed hep- arin affinity, and its mass was similar to that of the native enzyme. In addition, the in vivo plasma clear- ance in a rabbit model was similar to the previously studied native and recombinant forms. To our knowl- edge, this is the first example of efficient production of a tetrameric, protease-susceptible metalloprotein in milk of transgenic animals. Production at equivalent levels in transgenic farm animals would yield suffi- cient extracellular superoxide dismutase for therapeu- tic purposes. The secretory extracellular superoxide dismutase (EC- SOD)’ is one of three different SOD isoenzymes present in mammals (1). It isthe major SOD isoenzyme in plasma, lymph, and synovial fluid (2-5) but exists primarily in the interstitial space of tissues (6-8). The other two isoenzymes are the dimeric CuZn-SOD, which is an intracellular cytosolic enzyme, and the tetrameric Mn-SOD, which is found in the mitochondrial matrix (9-11). The human EC-SOD cDNA has been isolated and sequenced (12), and the recombinant pro- tein has been produced in mammalian cells (13). The protein is composed of four equal subunits, each containing 222 amino acids with a calculated molecular mass of 24.2 kDa (13). Each subunit binds one copper and one zinc atom and has a single * This work wasfinancially supported by Symbicom AB and Grant 9204 from the Swedish Natural Science Research Council. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “adver- tisement’’in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 8 To whom correspondence should be addressed. Tel.: 46-90- 190120; Fax: 46-90-192332. l The abbreviations used are: EC-SOD, extracellular superoxide dismutase; SOD, superoxide dismutase; bp, base pair(s); WAP, whey acidic protein; kb, kilobase pair(s); MOPS, 3-(N-morpholino)pro- panesulfonic acid; CHO, Chinese hamster ovary; PAGE, polyacryl- amide gel electrophoresis. N-glycosylation site at asparagine 89 (13, 14). The glycan of the native protein has not been the subject of any thorough investigation, but its lectin binding properties have been studied (1,13). A fundamental and distinguishing property of EC-SOD is its affinity for some glycosaminoglycans such as heparin and heparan sulfate (1,3,4,15, 16). The latter, which exists in the glycocalyx of cell surfaces and in the connective tissue matrix, is the important physiological ligand of EC- SOD. Because of this affinity, EC-SOD forms in the vascu- lature an equilibrium between the plasma phaseand the glycocalyx of the endothelium (3, 4, 17, 18). In tissues, vir- tually all EC-SOD exists anchored to heparan sulfate in the interstitial space and on the cell surfaces (8). The positively charged carboxyl-terminal end of the enzyme is responsible for the heparin binding (19). This domain is sensitive to proteolysis (20) resulting in EC-SOD forms with reduced or absent heparin affinities in vivo (8). Upon intravenous injection, EC-SOD rapidly binds to the surface of the endothelium and displays a long half-life (15- 20 h) in the vasculature (15,18). CuZn-SOD (parenterally administered) has been shown to exert a host of therapeutic actions (21-25). The experience with EC-SOD is in compari- son small (26-33), butthis isoenzyme appears even more potent. The high efficacy is most certainly related to the special pharmacokinetic properties of the enzyme. Genetic control elementsfrom different milk protein genes have been used to direct production of recombinant proteins in the milk of transgenic animals (34-36). The use of such transgenic techniques in farm animals offers the potential to produce pharmaceutical proteins in the milk. The potential advantages are the capability to produce recombinant, com- plex post-translationally modified, human proteins in large quantities in a highly cost-effective system without detriment to the producing animal. The temporal and spatial distribution of recombinant gene expression directed by milk protein gene regulatory fragments from various species seems to be very conserved and general among all species studied (37,38). Despite the fact that many cDNAs and genomic fragments now have been evaluated in mammary gland expression systems in transgenic animals, it is hard to define the factors that affect the level of expression. The expression levels reported are for some vector constructs very high, whereas they are disappointingly low for others (35). The qualitative aspects related to detailed analysis of post-translational modifications of recombinant proteins pro- duced in themilk of transgenic animals have received limited attention. In this study, the expression and functionality of a recombinant human glycosylated multimeric metalloprotein expressed in mouse milk are described. 5358
Transcript
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 7, Issue of February 18, pp. 5358-5363,1994 Printed in U.S.A.
Expression and Characterization of Biologically Active Human Extracellular Superoxide Dismutase in Milk of Transgenic Mice*
(Received for publication, June 29, 1993, and in revised form, September 24, 1993)
Lennart HanssonSf, Michael EdlundS, Anders EdlundS, Thore JohanssonS, Stefan L. Marklundll, Sigurd FrommII, Mats StromqvistS, and Jan Tornell** From SSymbicom AB, P.O. Box 1451, S-901, 24 Umed, Sweden, the nDepartment of Clinical Chemistry, Umed University Hospital, S-901 85 Umed, Sweden, the IlLaboratory of Molecular Embryology, University of Oslo, P.O. Box 1052 Blindern, N-0316 Oslo 3. Norway, and the **Department of Physiology, University of Goteborg, Medicinaregatan 11, S-413 90 Goteborg, Swiden
. -
We have targeted the expression of recombinant hu- man extracellular superoxide dismutase, a glycosy- lated tetrameric metalloprotein, to the mammary gland of transgenic mice. This was achieved by using regu- latory elements from either the murine whey acidic protein gene or the ovine &lactoglobulin gene to con- trol expression of human extracellular superoxide dis- mutase cDNA. Whey acidic protein regulatory se- quences directed high level mammary gland-specific expression of the recombinant gene and secretion of biologically active extracellular superoxide dismutase into the milk. The produced recombinant protein was fully active, it was in tetrameric form, it showed hep- arin affinity, and its mass was similar to that of the native enzyme. In addition, the in vivo plasma clear- ance in a rabbit model was similar to the previously studied native and recombinant forms. To our knowl- edge, this is the first example of efficient production of a tetrameric, protease-susceptible metalloprotein in milk of transgenic animals. Production at equivalent levels in transgenic farm animals would yield suffi- cient extracellular superoxide dismutase for therapeu- tic purposes.
The secretory extracellular superoxide dismutase (EC- SOD)’ is one of three different SOD isoenzymes present in mammals (1). It is the major SOD isoenzyme in plasma, lymph, and synovial fluid (2-5) but exists primarily in the interstitial space of tissues (6-8). The other two isoenzymes are the dimeric CuZn-SOD, which is an intracellular cytosolic enzyme, and the tetrameric Mn-SOD, which is found in the mitochondrial matrix (9-11). The human EC-SOD cDNA has been isolated and sequenced (12), and the recombinant pro- tein has been produced in mammalian cells (13). The protein is composed of four equal subunits, each containing 222 amino acids with a calculated molecular mass of 24.2 kDa (13). Each subunit binds one copper and one zinc atom and has a single
* This work was financially supported by Symbicom AB and Grant 9204 from the Swedish Natural Science Research Council. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “adver- tisement’’ in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
8 To whom correspondence should be addressed. Tel.: 46-90- 190120; Fax: 46-90-192332.
l The abbreviations used are: EC-SOD, extracellular superoxide dismutase; SOD, superoxide dismutase; bp, base pair(s); WAP, whey acidic protein; kb, kilobase pair(s); MOPS, 3-(N-morpholino)pro- panesulfonic acid; CHO, Chinese hamster ovary; PAGE, polyacryl- amide gel electrophoresis.
N-glycosylation site at asparagine 89 (13, 14). The glycan of the native protein has not been the subject of any thorough investigation, but its lectin binding properties have been studied (1,13). A fundamental and distinguishing property of EC-SOD is its affinity for some glycosaminoglycans such as heparin and heparan sulfate (1,3,4,15, 16). The latter, which exists in the glycocalyx of cell surfaces and in the connective tissue matrix, is the important physiological ligand of EC- SOD. Because of this affinity, EC-SOD forms in the vascu- lature an equilibrium between the plasma phase and the glycocalyx of the endothelium (3, 4, 17, 18). In tissues, vir- tually all EC-SOD exists anchored to heparan sulfate in the interstitial space and on the cell surfaces (8). The positively charged carboxyl-terminal end of the enzyme is responsible for the heparin binding (19). This domain is sensitive to proteolysis (20) resulting in EC-SOD forms with reduced or absent heparin affinities in vivo (8).
Upon intravenous injection, EC-SOD rapidly binds to the surface of the endothelium and displays a long half-life (15- 20 h) in the vasculature (15, 18). CuZn-SOD (parenterally administered) has been shown to exert a host of therapeutic actions (21-25). The experience with EC-SOD is in compari- son small (26-33), but this isoenzyme appears even more potent. The high efficacy is most certainly related to the special pharmacokinetic properties of the enzyme.
Genetic control elements from different milk protein genes have been used to direct production of recombinant proteins in the milk of transgenic animals (34-36). The use of such transgenic techniques in farm animals offers the potential to produce pharmaceutical proteins in the milk. The potential advantages are the capability to produce recombinant, com- plex post-translationally modified, human proteins in large quantities in a highly cost-effective system without detriment to the producing animal.
The temporal and spatial distribution of recombinant gene expression directed by milk protein gene regulatory fragments from various species seems to be very conserved and general among all species studied (37,38). Despite the fact that many cDNAs and genomic fragments now have been evaluated in mammary gland expression systems in transgenic animals, it is hard to define the factors that affect the level of expression. The expression levels reported are for some vector constructs very high, whereas they are disappointingly low for others (35). The qualitative aspects related to detailed analysis of post-translational modifications of recombinant proteins pro- duced in the milk of transgenic animals have received limited attention. In this study, the expression and functionality of a recombinant human glycosylated multimeric metalloprotein expressed in mouse milk are described.
5358
Expression of Human EC-SOD in Transgenic Mice 5359
MATERIALS AND METHODS
Construction of Transgenes-The human EC-SOD cDNA (12) was subcloned as a 1396-bp EcoRI fragment into pUC18. The cDNA was modified in the 5’ end by insertion of a Hind11 linker 32 bp upstream of the start codon and at the 3’ end by insertion of a SalI linker downstream of the translational stop. To facilitate introduction of human EC-SOD cDNA under control of the WAP promoter, a plas- mid harboring the 7.2-kb EcoRI fragment containing the murine WAP gene (39) was digested with KpnI, and a HindIII linker was inserted. The resulting plasmid was digested with HindIII and SalI, thereby removing the sequence between KpnI in exon I and Sal1 located in exon I11 of the WAP gene. This fragment was then ligated with the HindIII/SalI human EC-SOD cDNA fragment. The result- ing vector, pS172, was then digested with EcoRI, and a fragment of about 5.9 kb (Fig. 1) was isolated and purified by electroelution before injection into mouse embryos.
A second expression vector, pS315, was a variant of pS172 having a longer 5’ WAP of about 4.5 kb, and the translational start of EC- SOD was inserted directly downstream of the KpnI site in exon I of the murine WAP gene. The WAP/EC-SOD recombinant gene was isolated, purified by electroelution, and injected as described below.
A third expression vector called pS387, containing 1.8-kb ovine @- lactoglobulin upstream regulatory sequence (40) in front of human EC-SOD cDNA, was also constructed. The ovine @-lactoglobulin promoter fragment was cloned as a BamHI/PuuII fragment into BamHIISmaI-digested pUC19. From this plasmid, the promoter frag- ment was then isolated as a 1.8-kb HindIIIIKpnI (partial digestion) fragment. This @-lactoglobulin promoter fragment was then ligated to a human EC-SOD cDNA that was modified by insertion of a KpnI site in front of the start codon. Downstream of the EC-SOD stop codon, a murine 4.6-kb WAP genomic fragment was inserted, provid- ing the mRNA processing signals. This 4.6-kb WAP fragment extends from the KpnI site, which was changed to a SalI, in the first exon to the EcoRI site located about 1.6 kb downstream of the last exon. The @-lactoglobulin/EC-SOD/WAP fragment was isolated from pS387 and injected. The milk protein gene sequences were kindly provided by Dr. Lothar Hennighausen.
Generation and Screening of Transgenic Mice-An EcoRI fragment was isolated from the plasmid pS172 and used for injection (Fig. 1). Alternatively, the WAP/EC-SOD fragment of pS315 or B-lactoglob- ulin/EC-SOD/WAP fragment of pS358 was isolated and injected. The isolated fragments were injected into the pronucleus of C57B1/ 6JxCBA-fi or C57B1/6JxDBA/2J-fZ embryos obtained after priming of donor female mice with 5 IU of pregnant mare’s serum gonadotro- pin followed by 5 IU of human chorion gonadotropin 48 h later for superovulation of the mice. The C57B1/6JxCBA-fl animals were obtained from Bomholtgird Breeding and Research Centre Ltd. (Ry, Denmark). The injections were performed using Narishigi hydraulic micromanipulators and a Nikon inverted microscope (41). After in- jection the embryos were implanted in pseudopregnant C57B1/ 6JxCBA-f1 recipients. Mice having integrated the transgene were identified with polymerase chain reaction analysis of DNA from tail biopsy specimens obtained 3 weeks after the birth of the animals. Biopsies were incubated in 0.6 ml of urea lysis buffer containing 1 M urea, 50 mM Tris-HC1, pH 8.0, 0.1 M NaCl, 0.5% SDS, 5 mM EDTA, and 200 mg/ml proteinase K. Samples were incubated overnight at 37 “c. Polymerase chain reaction analysis was performed in 50 mM KCI, 10 mM Tris-HC1, pH 8.4, 1.5 mM MgCl2, 0.25 mM dNTP, 2.5 units of Taq polymerase (Life Technologies, Inc.), using primers complementary to WAP (5”CTGTGTGGCCAAGAAGGAAGTTT- GT-3’), @-lactoglobulin (5’-GCCTGAGGATGAGCCAAGTG-3’), and EC-SOD (5’-GTCCAGCGTGGCCGACGGCTGCACCTG-3’) sequences, respectively. Positive results were confirmed by Southern blot analysis using the human EC-SOD cDNA as a probe (42).
RNA Isolation and Analysis-Total RNA from various tissues of transgenic lactating female mice was isolated (42). Samples of 10 pg of total RNA were fractionated on MOPS/formaldehyde-agarose gels, transferred to GeneScreen Plus membranes (DuPont NEN), and subsequently hybridized to a 32P-labeled probe according to the sup- plier’s protocol (Amersham, Little Chalfont, United Kingdom).
Milk Collection-Mice were anesthetized with 0.017 ml of Avertin (2.5%)/g of body weight (41) and given 0.2 IU of oxytocin (Ferring, Lund, Sweden) intraperitoneally 10 min before milk collection. The milk was collected using a milk apparatus that collected the milk by continuous suction into chilled 1.5-ml Eppendorf tubes. To inhibit proteases present in the milk or released from cells disrupted by freeze/thawing, a mixture of protease inhibitors was immediately
added to some milk samples. The final concentrations were 0.1 pg/ ml aprotinin, 0.5 pg/ml E-64, 50 pg/ml chymostatin, 10 pM (4- amidinopheny1)methanesulfonyl fluoride, and 0.1 mM EDTA (all inhibitors were obtained from Boehringer Mannheim, Mannheim, Germany). Milk was stored frozen at -70 “C.
Partial Purification of EC-SOD Produced in Transgenic Mice- Milk collected from mice was diluted 10 times with distilled water and thereafter centrifuged at 8000 X g for 5 min. The lipid layer at the surface was carefully removed, and the supernatant was collected. The skimmed milk was diluted 1:l with 50 mM sodium phosphate, 0.5 M NaCl, pH 7.0 (buffer A) and thereafter applied to a column with a monoclonal antibody directed against native human EC-SOD. The column was washed with buffer A and eluted with 50 mM 2- amino-2-methyl-1-propanol-HCl, 1 M KSCN, pH 9.0. The eluate was ultrafiltrated and transferred to distilled water and stored frozen at -20 “C. The protein concentration of collected fractions was meas- ured using the method of Bradford (43) with recombinant EC-SOD produced in CHO cells as standard.
Quantification of EC-SOD-SOD enzymatic activity was deter- mined with the direct spectrophotometric method using KO, (44) as modified by Marklund (45). One unit corresponds to 8.6 ng of human EC-SOD (13). The concentration of EC-SOD antigen was determined by enzyme-linked immunosorbent assay (17). As reference substances native umbilical cord and recombinant CHO cell-produced EC-SOD were used (13).
Determination of Apparent Molecular Masses by Means of Size- exclusion Chromatography-The chromatographies were carried out at 4 “C on a calibrated Sephacryl S-300 (Pharmacia, Uppsala, Swe- den) column (1.6 X 89 cm), equilibrated and eluted with 10 mM potassium phosphate, pH 7.4, containing 0.15 M NaC1, as previously detailed (4).
SDS-PAGE and Zmmunoblotting-SDS gels were run in a Midget electrophoresis unit (Pharmacia LKB Biotechnology Inc.) according to the discontinuous buffer system described by Laemmli (46). Gels were blotted in 48 mM Tris, 39 mM glycine, 1.3 mM SDS, 20% methanol, in a Trans-Blot SD unit (Bio-Rad) under constant voltage (0.8 mA/cm2). EC-SOD was detected by the use of polyclonal anti- bodies prepared in rabbits using the native protein for immunization and immobilized EC-SOD for affinity purification. Alkaline phospha- tase anti-rabbit IgG (Dakopatts, Copenhagen, Denmark) was used as a secondary antibody.
Carbohydrate Comparison-The binding of the different SODS to lectins was analyzed using the lectin-link kit (Genzyme, Cambridge, MA). The proteins were first run on SDS-PAGE, electroblotted, and developed using the different lectins, according to the vendor’s in- structions.
Analytical Separation of EC-SOD on Heparin-Sepharose-The chromatographies were carried out as previously detailed (4) at room temperature in a 1-ml heparin-Sepharose (Pharmacia) column with 15 m~ sodium cacodylate, 50 mM NaC1, pH 6.5, as equilibration buffer and eluent. Bound material was eluted with a NaCl gradient.
Plasma Clearance of EC-SOD in Rabbits-Recombinant EC-SOD from mouse milk was partially purified by chromatography on hepa- rin-Sepharose equilibrated with 50 mM sodium phosphate, pH 7.4. Bound EC-SOD was eluted with a gradient of NaCl in the same buffer. The central two-thirds of the high heparin-affinity fraction (cf. Fig. 5 ) was collected and used for plasma clearance studies. About 10 pg of EC-SOD/kg of body weight, dissolved in 50 mM potassium phosphate, pH 7.4, containing 0.2% bovine serum albumin, was injected into ear veins of 34-kg chinchilla ram rabbits. Blood samples were tapped at the times indicated in Fig. 4 into tubes containing EDTA as an anticoagulant. The experiments were terminated at 1, 6, or 24 h (cf. Fig. 4) by intravenous injection of heparin, 2500 IU/kg of body weight, followed by tapping of blood samples at 2, 5, and 15 min thereafter. The plasma volumes varied between 34 and 38 rnl/kg of body weight. The rabbits were used only once.
RESULTS
Generation of Transgenic Animals-To direct expression of recombinant human EC-SOD to the mammary gland of trans- genic mice, a hybrid WAP/EC-SOD gene, pS172, was con- structed. This recombinant gene comprises about 2.3 kb of upstream regulatory sequences including the first 24 bp of the transcribed but untranslated WAP sequence in front of the human EC-SOD cDNA. Downstream of the cDNA, a WAP genomic fragment of about 4 kb extending from the Sal1 site
5360 Expression of Human EC-SOD in Transgenic Mice
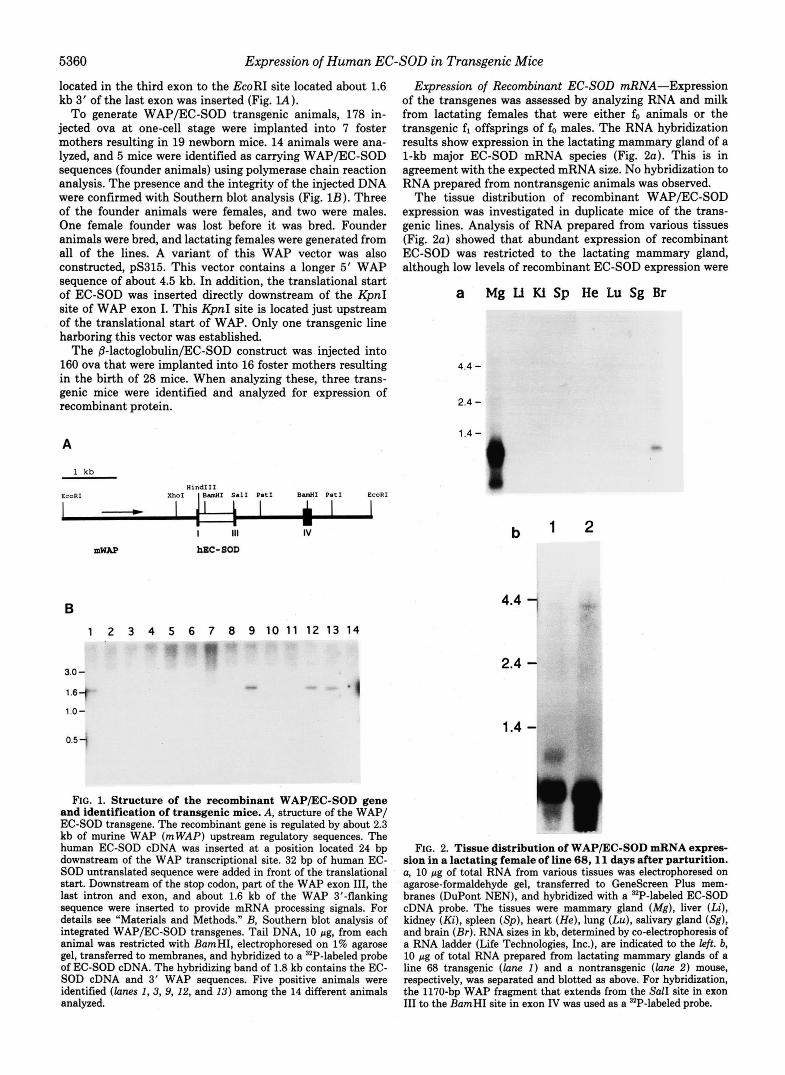
located in the third exon to the EcoRI site located about 1.6 kb 3‘ of the last exon was inserted (Fig. IA).
To generate WAP/EC-SOD transgenic animals, 178 in- jected ova at one-cell stage were implanted into 7 foster mothers resulting in 19 newborn mice. 14 animals were ana- lyzed, and 5 mice were identified as carrying WAP/EC-SOD sequences (founder animals) using polymerase chain reaction analysis. The presence and the integrity of the injected DNA were confirmed with Southern blot analysis (Fig. 1B). Three of the founder animals were females, and two were males. One female founder was lost before it was bred. Founder animals were bred, and lactating females were generated from all of the lines. A variant of this WAP vector was also constructed, pS315. This vector contains a longer 5‘ WAP sequence of about 4.5 kb. In addition, the translational start of EC-SOD was inserted directly downstream of the KpnI site of WAP exon I. This KpnI site is located just upstream of the translational start of WAP. Only one transgenic line harboring this vector was established.
The &lactoglobulin/EC-SOD construct was injected into 160 ova that were implanted into 16 foster mothers resulting in the birth of 28 mice. When analyzing these, three trans- genic mice were identified and analyzed for expression of recombinant protein.
A
1 kb
EcoRI Hind111
XhoI BamHI Sa11 P.tI BamHI PstI EcoRI
I - I , I A I I r I ‘ I I 111 IV
mWAP hEC-SOD
B 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4
3.0 - 1.6-
1.0-
0.5-
ErG. 1. Structure of the recombinant WAP/EC-SOD gene and identification of transgenic mice. A, structure of the WAP/ EC-SOD transgene. The recombinant gene is regulated by about 2.3 kb of murine WAP (mWAP) upstream regulatory sequences. The human EC-SOD cDNA was inserted at a position located 24 bp downstream of the WAP transcriptional site. 32 bp of human EC- SOD untranslated sequence were added in front of the translational start. Downstream of the stop codon, part of the WAP exon 111, the last intron and exon, and about 1.6 kb of the WAP 3”flanking sequence were inserted to provide mRNA processing signals. For details see “Materials and Methods.” B, Southern blot analysis of integrated WAP/EC-SOD transgenes. Tail DNA, 10 pg. from each animal was restricted with BamHI, electrophoresed on 1% agarose gel, transferred to membranes, and hybridized to a 32P-labeled probe of EC-SOD cDNA. The hybridizing band of 1.8 kb contains the EC- SOD cDNA and 3’ WAP sequences. Five positive animals were identified (lanes I , 3, 9, 12, and 13) among the 14 different animals analyzed.
Expression of Recombinant EC-SOD mRNA-Expression of the transgenes was assessed by analyzing RNA and milk from lactating females that were either fo animals or the transgenic fi offsprings of fo males. The RNA hybridization results show expression in the lactating mammary gland of a 1-kb major EC-SOD mRNA species (Fig. 2a). This is in agreement with the expected mRNA size. No hybridization to RNA prepared from nontransgenic animals was observed.
The tissue distribution of recombinant WAP/EC-SOD expression was investigated in duplicate mice of the trans- genic lines. Analysis of RNA prepared from various tissues (Fig. 2a) showed that abundant expression of recombinant EC-SOD was restricted to the lactating mammary gland, although low levels of recombinant EC-SOD expression were
a Mg U Ki Sp
4.4 - 2.4 -
1.4-
I b 1
He Lu Sg Br
2
4.4 -
2.4 -
1.4 -
FIG. 2. Tissue distribution of WAP/EC-SOD mRNA expres- sion in a lactating female of line 6 8 , l l days after parturition. a, 10 pg of total RNA from various tissues was electrophoresed on
branes (DuPont NEN), and hybridized with a 32P-labeled EC-SOD agarose-formaldehyde gel, transferred to GeneScreen Plus mem-
cDNA probe. The tissues were mammary gland (Mg), liver (Li), kidney (Ki), spleen (Sp ) , heart (He), lung (Lu), salivary gland (Sg), and brain (Br) . RNA sizes in kb, determined by co-electrophoresis of a RNA ladder (Life Technologies, Inc.), are indicated to the left. b, 10 pg of total RNA prepared from lactating mammary glands of a line 68 transgenic ( l a n e I ) and a nontransgenic ( l a n e 2 ) mouw, respectively, was separated and blotted as above. For hybridization, the 1170-bp WAP fragment that extends from the Sal1 site in exon 111 to the BamHI site in exon IV was used as a 3ZP-labeled probe.
Expression of Human EC-SOD in Transgenic Mice 5361
TABLE I Production of human EC-SOD in milk of transgenic mice
Vector Transgenic line EC-SOD wlml
pS172 65 90 66,67 UD" 68 710,000
pS315 90 15 pS387 607,702 UD
716 8 None Control 0-0.75
a UD, undetectable, within the range of controls.
also detected in brain. In additional experiments, where the same WAP regulatory sequences were used to direct expres- sion of other human cDNAs, no recombinant gene expression was detected in the brain.'
Northern blots of mammary gland RNA of transgenic line 68 and controls were hybridized with a mouse WAP exon I11 and exon IV probe, which hybridizes to the same extent to both endogenous WAP mRNA and to recombinant WAP/ EC-SOD mRNA. The endogenous WAP mRNA level was found to be significantly higher than the level of recombinant WAP/EC-SOD mRNA (Fig. 2b). The p-lactoglobulin/EC- SOD transgenic lines were not analyzed for mRNA expres- sion.
Secretion of Recombinant EC-SOD into Mouse Milk-EC- SOD was primarily determined with enzyme-linked immu- nosorbent assay. The production varied a lot between trans- genic lines but was overall highest in the WAP transgenes (Table I). One line, 68, displayed a very high productivity, around 0.7 mg/ml. This level of EC-SOD corresponds to about 1% of the total milk protein.
Because the SOD activity of control milk was about 100 units/ml, enhancements in activity in the milk samples with low EC-SOD production could not be distinguished with certainty. However, in line 68 the SOD activity in the milk was very high (Fig. 3), and the specific activity (units/ng EC- SOD) was apparently identical to that of native and CHO- produced recombinant EC-SOD (13). Production of recombi-
L. Hansson, M. Edlund, A. Edlund, T. Johansson, S. Marklund, S. Fromm, M. Stromqvist, and J. Tornell, unpublished observation.
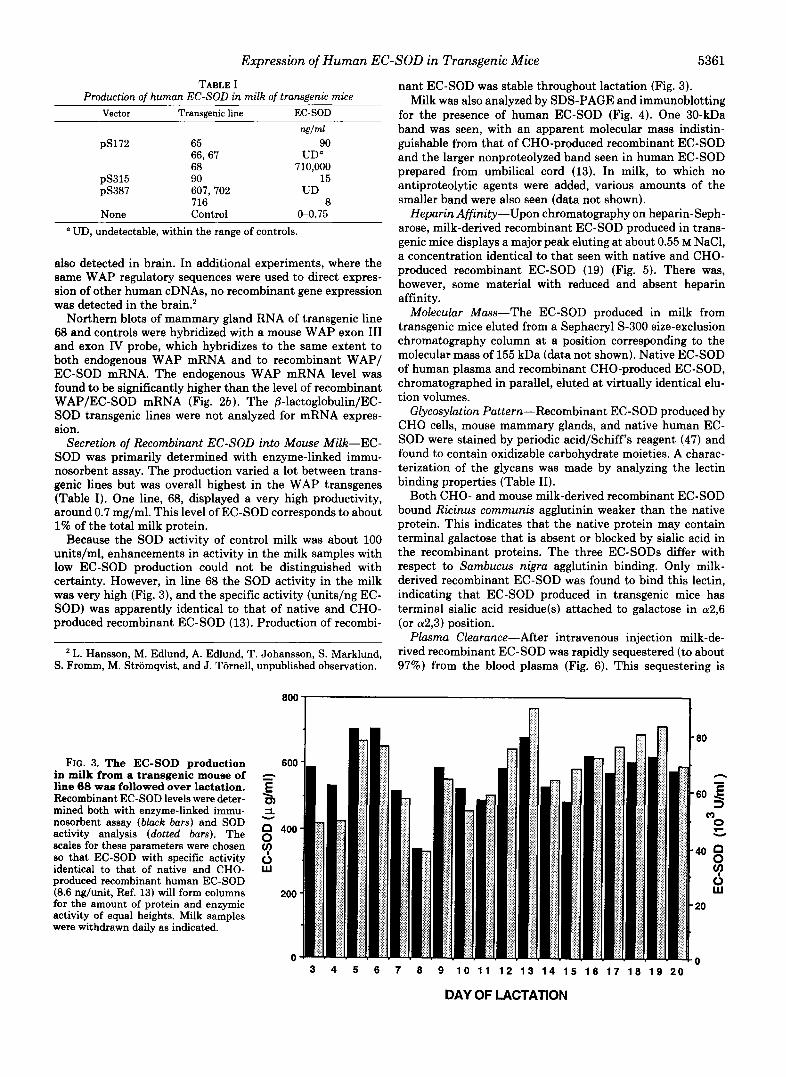
nant EC-SOD was stable throughout lactation (Fig. 3). Milk was also analyzed by SDS-PAGE and immunoblotting
for the presence of human EC-SOD (Fig. 4). One 30-kDa band was seen, with an apparent molecular mass indistin- guishable from that of CHO-produced recombinant EC-SOD and the larger nonproteolyzed band seen in human EC-SOD prepared from umbilical cord (13). In milk, to which no antiproteolytic agents were added, various amounts of the smaller band were also seen (data not shown).
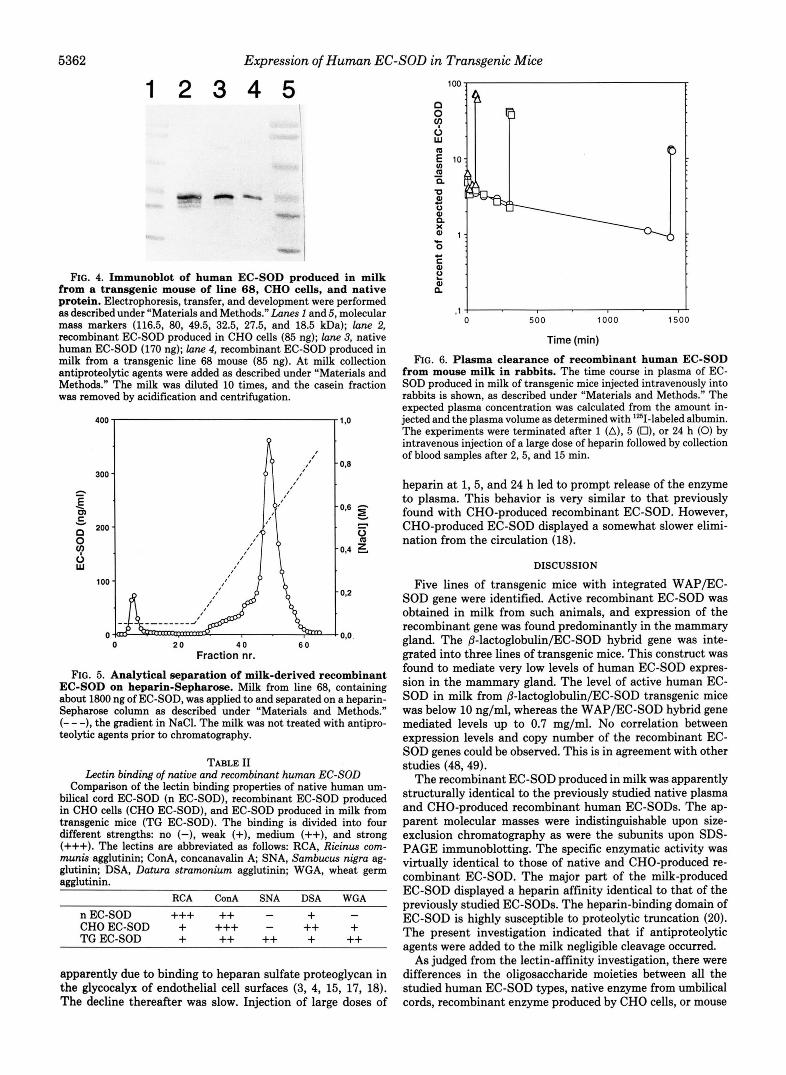
Heparin Affinity-Upon chromatography on heparin-Seph- arose, milk-derived recombinant EC-SOD produced in trans- genic mice displays a major peak eluting at about 0.55 M NaC1, a concentration identical to that seen with native and CHO- produced recombinant EC-SOD (19) (Fig. 5). There was, however, some material with reduced and absent heparin affinity.
Molecular Mass-The EC-SOD produced in milk from transgenic mice eluted from a Sephacryl S-300 size-exclusion chromatography column at a position corresponding to the molecular mass of 155 kDa (data not shown). Native EC-SOD of human plasma and recombinant CHO-produced EC-SOD, chromatographed in parallel, eluted at virtually identical elu- tion volumes.
Glycosylation Pattern-Recombinant EC-SOD produced by CHO cells, mouse mammary glands, and native human EC- SOD were stained by periodic acid/Schiff's reagent (47) and found to contain oxidizable carbohydrate moieties. A charac- terization of the glycans was made by analyzing the lectin binding properties (Table 11).
Both CHO- and mouse milk-derived recombinant EC-SOD bound Ricinus communis agglutinin weaker than the native protein. This indicates that the native protein may contain terminal galactose that is absent or blocked by sialic acid in the recombinant proteins. The three EC-SODS differ with respect to Sambucus nigra agglutinin binding. Only milk- derived recombinant EC-SOD was found to bind this lectin, indicating that EC-SOD produced in transgenic mice has terminal sialic acid residue(s) attached to galactose in a2,6 (or a2,3) position.
Plasma Clearance-After intravenous injection milk-de- rived recombinant EC-SOD was rapidly sequestered (to about 97%) from the blood plasma (Fig. 6) . This sequestering is
FIG. 3. The EC-SOD production in milk from a transgenic mouse of line 68 was followed over lactation. Recombinant EC-SOD levels were deter- mined both with enzyme-linked immu- nosorbent assay (black bars) and SOD activity analysis (dotted bars). The scales for these parameters were chosen so that EC-SOD with specific activity identical to that of native and CHO- produced recombinant human EC-SOD (8.6 ng/unit, Ref. 13) will form columns for the amount of protein and enzymic activity of equal heights. Milk samples were withdrawn daily as indicated.
FIG. 4. Immunoblot of human EC-SOD produced in milk from a transgenic mouse of line 68, CHO cells, and native protein. Electrophoresis, transfer, and development were performed as described under “Materials and Methods.” Lunes 1 and 5, molecular mass markers (116.5, 80, 49.5, 32.5, 27.5, and 18.5 kDa); lane 2, recombinant EC-SOD produced in CHO cells (85 ng); lane 3, native human EC-SOD (170 ng); lune 4, recombinant EC-SOD produced in milk from a transgenic line 68 mouse (85 ng). At milk collection antiproteolytic agents were added as described under “Materials and Methods.” The milk was diluted 10 times, and the casein fraction was removed by acidification and centrifugation.
0 20 4 0 6 0 Fraction nr.
FIG. 5. Analytical separation of milk-derived recombinant EC-SOD on heparin-Sepharose. Milk from line 68, containing about 1800 ng of EC-SOD, was applied to and separated on a heparin- Sepharose column as described under “Materials and Methods.” (- - -), the gradient in NaCl. The milk was not treated with antipro- teolytic agents prior to chromatography.
TABLE I1 Lectin binding of native and recombinant human EC-SOD
Comparison of the lectin binding properties of native human um- bilical cord EC-SOD (n EC-SOD), recombinant EC-SOD produced in CHO cells (CHO EC-SOD), and EC-SOD produced in milk from transgenic mice (TG EC-SOD). The binding is divided into four different strengths: no (-), weak (+), medium (++), and strong (+++). The lectins are abbreviated as follows: RCA, Ricinus com- munis agglutinin; ConA, concanavalin A, SNA, Sambucus nigra ag- glutinin; DSA, Datura stramonium agglutinin; WGA, wheat germ agglutinin.
apparently due to binding to heparan sulfate proteoglycan in the glycocalyx of endothelial cell surfaces (3, 4, 15, 17, 18). The decline thereafter was slow. Injection of large doses of
.SOD in Transgenic Mice
-1 4 L
0 500 1000 1500
Time (min)
FIG. 6. Plasma clearance of recombinant human EC-SOD from mouse milk in rabbits. The time course in plasma of EC- SOD produced in milk of transgenic mice injected intravenously into rabbits is shown, as described under “Materials and Methods.“ The expected plasma concentration was calculated from the amount in- jected and the plasma volume as determined with 1251-labeled albumin. The experiments were terminated after 1 (A), 5 (O), or 24 h (0) by intravenous injection of a large dose of heparin followed by collection of blood samples after 2,5, and 15 min.
heparin at 1,5, and 24 h led to prompt release of the enzyme to plasma. This behavior is very similar to that previously found with CHO-produced recombinant EC-SOD. However, CHO-produced EC-SOD displayed a somewhat slower elimi- nation from the circulation (18).
DISCUSSION
Five lines of transgenic mice with integrated WAPPC- SOD gene were identified. Active recombinant EC-SOD was obtained in milk from such animals, and expression of the recombinant gene was found predominantly in the mammary gland. The &lactoglobulin/EC-SOD hybrid gene was inte- grated into three lines of transgenic mice. This construct was found to mediate very low levels of human EC-SOD expres- sion in the mammary gland. The level of active human EC- SOD in milk from 8-lactoglobulin/EC-SOD transgenic mice was below 10 ng/ml, whereas the WAPPC-SOD hybrid gene mediated levels up to 0.7 mg/ml. No correlation between expression levels and copy number of the recombinant EC- SOD genes could be observed. This is in agreement with other studies (48,49).
The recombinant EC-SOD produced in milk was apparently structurally identical to the previously studied native plasma and CHO-produced recombinant human EC-SODS. The ap- parent molecular masses were indistinguishable upon size- exclusion chromatography as were the subunits upon SDS- PAGE immunoblotting. The specific enzymatic activity was virtually identical to those of native and CHO-produced re- combinant EC-SOD. The major part of the milk-produced EC-SOD displayed a heparin affinity identical to that of the previously studied EC-SODS. The heparin-binding domain of EC-SOD is highly susceptible to proteolytic truncation (20). The present investigation indicated that if antiproteolytic agents were added to the milk negligible cleavage occurred.
As judged from the lectin-affinity investigation, there were differences in the oligosaccharide moieties between all the studied human EC-SOD types, native enzyme from umbilical cords, recombinant enzyme produced by CHO cells, or mouse
Expression of Human EC-SOD in Transgenic Mice 5363
mammary glands. However, the finding does not necessarily indicate that the recombinant EC-SODS contain oligosaccha- ride moieties that do not exist bound to EC-SOD in the human body. The oligosaccharides are known to be hetero- geneous, even as produced by a single cell type (50). The umbilical cord native EC-SOD studied is probably mainly produced by fibroblasts. However, EC-SOD is also secreted by glia cells (51), smooth muscle cells: and possibly other not yet identified cell types. It is well known that the glycosylation of a protein may vary widely when it is produced by different cell types in the human body (52).
Finally, the recombinant EC-SOD from mouse milk was found to behave similarly to native and recombinant CHO- produced EC-SOD (17, 18) after intravenous injection into rabbits. There was a prompt binding to the vascular endothe- lium as well as a rapid release after injection of heparin. Possibly, the mouse milk-produced EC-SOD was a little more rapidly eliminated from the circulation than the previously studied forms.
To conclude, our study shows that by using transgenic techniques the mammary gland can be used as a bioreactor for high yield production of biologically active human EC- SOD, a tetrameric, protease-susceptible, copper- and zinc- containing glycoprotein.
Acknowledgments-We thank Jan-Olov Andersson, Asa Skytt, and Agneta C)berg for excellent technical assistance and Drs. Gunnar Skogman and Anders Bergman for generous support.
REFERENCES
2. Marklund, S. L., Holme, E., and Hellner, L. (1982) Clin. Chim. Acta 126 , 1. Marklund, S. L. (1982) Proc. Natl. Acad. Sci. U. S. A . 7 9 , 7634-7638
3. Karlsson, K., and Marklund, S. L. (1987). Biochem. J . 2 4 2 , 55-59 4. Karlsson. K., and Marklund, S. L. (1988) Biochem. J . 266,223-228 5. Marklund, S. L., Bjelle, A., and Elmqvist, L.-G. (1986) Ann. Rheum. Dis.
6. Marklund, S. L. (1984) J. Clin. Inuest. 74 , 1398-1403 7. Marklund, S. L. (1984) Biochem. J. 2 2 2 , 649-655 8. Sandstrom, J., Karlsson, K., Edlund, T., and Marklund, S. L. (1993)
9. McCord, J. M., and Fridovich, I. (1969) J. Biol. Chem. 2 4 4 , 6049-6055
41-51
46,847-851
Biochem. J. 294,853-857
10. Keele, B. B., Jr., McCord, J. M., and Fridovich, I. (1970) J. Biol. Chem.
11. Weisiger, R. A,, and Fridovich, 1. (1973) J. Biol. Chem. 2 4 8 , 4793-4796 12. Hjalmarsson, K., Marklund, S. L., Engstrom, A,, and Edlund, T. (1987)
13. Tibell, L., Hjalmarsson, K., Edlund, T., Skogman, G., Engstrom, A,, and
246,6176-6181
P m . Natl. Acad. Sci. U. S. A . 84,6340-6344
Marklund, S. L. (1987) Proc. Natl. Acad. Sci. U. S. A . 84,6634-6638
S. Marklund, unpublished observation.
14. Stromqvist, M., Holgersson, J., and Samuelsson, B. (1991) J. Chromatogr.
15. Karlsson, K., Llndahl, U., and Marklund, S. L. (1988) Biochem. J. 2 5 6 , 648,293-301.
29-23 16. Markfund, S. L., and Karlsson, K. (1989) Lab. Inuest. 60, 659-666 17. Karlsson, K., and Marklund, S. L. (1988) J. Clin. Inuest. 8 2 , 762-766 18. Sandstrom, J., Edlund, A., Karlsson, K., and Marklund, S. L. (1993) Free
19. Sandstrom, J. , Carlsson, L., Marklund, S. L., and Edlund, T. (1992) J. Biol.
20. Karlsson, K., Edlund, A., Sandstrom, J., and Marklund, S. L. (1993)
21. Ferrari, R., Ceconi, C., Curello, S., Ghielmi, S., and Albertini, A. (1989)
22. Schoenberg, M. H., and Beger, H. G. (1990) Chem. Biol. Interuct. 76,141-
23. Sanfey, H., Bulkley, G. B., and Cameron, J. L. (1984) Ann. Surg. 200 ,
24. Bannister, J. V., Bannister, W. H., and Rotilio, G. (1987) CRC Crit. Reu.
25. Omar, B. A,, Flores, S. C., and McCord, J. M. (1992) Adu. Pharmacol. 2 3 ,
26. Johansson, M., Deinum, J., Marklund, S. L., and SjMuist, P.-0. (1990)
27. Erlansson, M., Berg vist, D , Marklund, S. L., Persson, N. H., and Svensjo,
28. Sjoquist, P.-O., Carlsson, L., Jonasson, G., Marklund, S. L., and Abra-
29. S’oquist, P.-O., and Marklund, S. L. (1992) Cardiouusc. Res. 26,347-350 30. Abrahamsson, T., Brandt, U., Marklund, S. L., and Sjoquist, P.-0. (1992)
31. Hatori, N., S‘oquist, P.-0.. Marklund, S. L., Pehrsson, S. K., and Ryden,
32. Hatori, N., Sj” uist P 0 Marklund, S. L., and Ryden, L. (1992) Free
33. Wahlund, G., Marklund, S. L., and Sjoquist, P.-0. (1992) Free Rad. Res.
34. Hennighausen, L. (1990) Protein E x ression Pur$ 1: 3-8. , ,
35. Hennighausen, L., Ruiz, L., and Waf, R. (1990) urr Oprn Btotechnol. 1,
Radical Biol. & Med. 14 , 185-190
Chem. 267,18205-18209
Biochem. J. 290,623-626
Pharmacol. Res. 2 1,57-65
161
405-412
Biochem. 22,111-179
109-161
Cardiouusc. Res. 24,500-503
E. (1990) Free Ro%ical Bibl. & Med. 9, 59-65
hamsson, T. (1991) J. Cardiouasc. Phormacol. 17,678-683
Circ. Res. 70, 264-271
L. (1992) dree Radical B d . & Med. 13 , 137-142
Radical Bwl% MLd. 13;‘221-230
Commun. 17,41-47
74-7x 36. Wilmui, A,, Archibald, A. L., McClenaghan M., Simons, J. P Whitelaw,
C. B. A,, and Clark, A. J. (1991) Experienh (Basel) 47,905L912 37. Simons, J. P., McClenaghan, M., and Clark, A. J. (1987) Nature 328,530-
532 38. WafirR. J. Pursel, V. G., Shamay, A., McKni ht R A Pittius, C. W., and
Henni iausen L (1991) Proc Natl Acad. 8ci; S:k. 88,1701-1705 39. Campbelf S. M.,’Rosen, J. M., Hennighausen, L. G., Strech-Jurk, U., and
Sippe1,’A. E. (1984) Nucleic Acids Res. 12 8685-8697 40. Harris, S., Ali, S., Anderson, S., Archibald, ’A. L., and Clark, A. J. (1988)
Nucleic Acids Res. 1 6 , 10379-10380 41. Ho an, B., Constantini, F., and Lacy, E. (1986) in Manipulating the Mouse
.&&yo: A Luborato Manual, pp. 153-182, Cold Spring Harbor L a b -
42. A::LYF. M. fjrent R. Kin ston R. E. Moore, D. D., Seidman, J. G. , Smith’, J. A.,’and Sthhi , K. fleds)’(1992)’Current Protocols in Molecular
43. Bradfort M. &. (1976) A p l . Biochem. 72,2481254 Bwlog cha ters 2 and 4, John Wiley & Sons Inc., New York
44. Marklund, S. (1976) J. Btol. Chem. 2 5 1 , 7504-7507 45. Marklund, S. L. (1985) in Handbook ofMethods for Oxygen RadicalResearch
46. Laemmli, U. K. (1970) Nature 227,680-685 47. Stromqvist, M., and Gmffman, H. (1992) Biotechniques 13 , 744-749 48. Palmiter, R. D., and Brinster, R. L. (1986) Annu. Reu. Genet. 20,465-499 49. Pittius, W. C., Hennighausen, L., Lee, E., Westphal, H., Nicols E Vitale,
50. Rademacher R. W., Parekh, R. B., and Dwek, R. A. (1988) Annu. Reu. J., and Gordon, K. (1988) Proc. Natl. Acad. Sci. U. S. A. 86, b8?4-5878
51. Marklund, S. L. (1990) Biochem. J. 260 , 213-219 Biochem. 5 7 , 785-838
52. Moss, D. W., and Whitaker, K. B. (1985) Enzyme (Busel) 3 4 , 212-216
Cold S ring 8arbor, NY
(Greenwald, R. A., ed) pp. 249-255, CRC Press Inc., Boca Raton, FL