Expression and function of clift in the development of somatic gonadal
precursors within the Drosophila mesoderm
Monica Boyle1, Nancy Bonini2 and Stephen DiNardo1,*1The Rockefeller University, 1230 York Avenue, NYC, NY 10021-6399, USA2The University of Pennsylvania, Department of Biology, 415 S. University Avenue, Philadelphia, PA 19104, USA
The gonad forms from cells of two lineages: the germlineand soma. The somatic gonadal cells generate the variouscell types within the testis or ovary that support gameto-genesis. These cells derive from embryonic mesoderm, buthow they are specified is unknown. Here, we describe anovel regulator of Drosophila gonadogenesis, clift,mutations in which abolish gonad formation. clift isexpressed within somatic gonadal precursors as these cellsfirst form, demonstrating that 9-12 cells are selected assomatic gonadal precursors within each of three posteriorparasegments at early stages in gonadogenesis. Despite thisearly expression, somatic gonadal precursors are specifiedin the absence of clift function. However, they fail tomaintain their fate and, as a consequence, germ cells do notcoalesce into a gonad. In addition, using clift as a marker,we show that the anteroposterior and dorsoventral positionof the somatic gonadal precursor cells within a paraseg-
ment are established by the secreted growth factor Wg,coupled with a gene regulatory hierarchy within themesoderm. While loss of wg abolishes gonadal precursors,ectopic expression expands the population such that mostcells within lateral mesoderm adopt gonadal precursorfates. Initial dorsoventral positioning of somatic gonadalprecursors relies on a regulatory cascade that establishesdorsal fates within the mesoderm and is subsequentlyrefined through negative regulation by bagpipe, a gene thatspecifies nearby visceral mesoderm. Thus, these studiesidentify essential regulators of gonadal precursor specifi-cation and differentiation and reveal novel aspects of thegeneral mechanism whereby distinct fates are allocatedwithin the mesoderm.
The gonad forms during embryogenesis from germline andsomatic cells. It is the interaction of these cells types withinthis organ that leads to the production of gametes in the adult.In many organisms, these two components are initially pos-itioned apart from one another due to the early segregation ofgerm cells away from the rest of the developing embryo. It isnot until a later stage that germ cells migrate back inside theembryo to contact somatic gonadal cells, with which they willcondense into a gonad. Therefore, gonadogenesis provides anopportunity to examine the mechanisms whereby cells ofseparate lineages are specified, recognize and adhere to oneanother, and assemble into a functional organ.
In Drosophila, the association of germ cells and somaticgonadal precursor cells (SGP cells) has been described(Brookman et al., 1992; Warrior, 1994; Boyle and DiNardo,1995). The germ cells, after an initial displacement at theposterior pole of the embryo, reenter the embyro by squeezingthrough gut endoderm (Sonnenblick, 1941; Jaglarz and Howard,1994, 1995; Warrior, 1994). Shortly afterwards, germ cellscontact mesoderm and adhere to SGP cells that lie within themesoderm of several posterior abdominal segments. Gonads
form, however, at the most anterior of these segments. Therefore,germ cells and SGP cells must migrate anteriorly. They do so inclose association with one another, eventually settling in the fifthabdominal segment and condensing into a gonad.
Although germ cells and SGP cells are associated through-out later stages of their migration to the gonad, germ cells aredispensible to gonadogenesis. In the absence of germ cells,SGP cells cluster into a gonad normally (Aboïm, 1945;Brookman et al., 1992; Warrior, 1994; Boyle and DiNardo,1995). In contrast, SGP cells are essential for ensuring theincorporation of germ cells into the gonad. Mutations thataffect SGP cell development disrupt germ cell migration to thegonad (Cumberledge et al., 1992; Brookman et al., 1992;Warrior, 1994; Boyle and DiNardo, 1995). Given this crucialrole in directing gonadogenesis, we have focused on the devel-opment of SGP cells. Uncovering the regulatory pathwaysresulting in their specification and differentiation is a first steptoward understanding the mechanisms of cellular recognition,guided migration and organogenesis used by these cells.
Despite the identification of the SGP cells and a descriptionof their association with germ cells into a gonad, little is knownabout how these cells are generated within the mesoderm. SGPcells derive from segments of the embryo whose identity is
972 M. Boyle, N. Bonini and S. DiNardo
defined by the homeotic gene abdominal A and indeed, abd Afunction is required for their development (Cumberledge et al.,1992; Brookman et al., 1992; Warrior, 1994; Boyle andDiNardo, 1995). However, the mechanism whereby abd Aprovides this function or what additional factors are required fortheir specification is not known. Recently, several studies haveidentified both intrinsic and extrinsic factors that act to determineparticular cell types within the mesoderm (Azpiazu and Frasch,1993; Staehling-Hampton et al., 1994; Frasch, 1995; Carmenaet al., 1995; Wu et al., 1995; Lawrence et al., 1995; Baylies etal., 1995; Baylies and Bate, 1996). To date, however, there hasbeen little consideration of the gonadal soma. This is due in partto the lack of markers that identify SGP cells or mutations thatspecifically affect their development. Although such mutationscould in theory be identified in screens for female and malesterility, no mutations that specifically affect the developmentand survival of the gonadal mesoderm have been isolated despiteextensive screening (Schupbach and Wieschaus, 1991; Castril-lon et al., 1993). Therefore, genes required for SGP developmentare likely to have other essential functions.
Based on these observations, we screened lethal collectionsto identify genes required for gonad formation. Here, wedescribe one such mutant, clift (cli, also known as eyes absent),mutations in which disrupt gonad formation. cli was originallyidentified as an embryonic lethal mutation with defects in headdevelopment (Nusslein-Volhard et al., 1984). It was later foundto be allelic to eyes absent, a viable cli mutation with defectsin photoreceptor cell differentiation and survival, and encodesa novel nuclear protein (Bonini et al., 1993). We find that clialso has an essential role in SGP cell development. In itsabsence, SGP cells do not differentiate and, as a consequence,germ cell migration is affected. Consistent with this require-ment, we show that cli expression identifies SGP cells as thesecells first form. Using cli expression as a marker, we identifyessential regulators that act to ensure the precise allocation ofSGP cells within the mesoderm.
MATERIALS AND METHODS
Fly stocksLine 68-77 (Fig. 1A), is a P element transgenic line, which expressesLacZ in all somatic cells of the embryonic gonad (Simon et al., 1990;Boyle and DiNardo, 1995).
The cliIID and cliIIE alleles were obtained from Bloomington andTubingen stock centers and are described in Nusslein-Volhard et al.(1984). The eyaDI allele, a presumed null allele, is described in Boniniet al. (1993). All stocks are maintained over a balancer that carries aen promoter-LacZ transgene inserted at wingless (Pw+ wg[1-en-11];Kassis, 1990). Phenotypes were examined in heteroallelic combina-tions, or over the defiency Df (2L)E(P)41, generated by the Boninilab. hsp70-EyaII line K58.1 was constructed by the Bonini lab.
The 1-7′-980 dpp LacZ transgenic line was provided by the Coureylab and is described in Huang et al. (1993).
The tin346 stock was provided to us by Manfred Frasch and isdescribed in Azpiazu and Frasch (1993). tin mutant embryos wereidentified by the absence of evenskipped staining in heart precursors(Azpiazu and Frasch, 1993). bagpipe null embryos were generated bycrossing Df(3R)eD7, P(tinre58)/TM3 ftzlacZ flies, carrying a tinrescue transgene, to Df(3R)eF1/TM3 ftzlacZ flies, also obtained fromthe Frasch lab.
Hs-Wg/TM3 is described in Noordermeer et al. (1992) and Hs-abdA obtained from Gines Morata. For ectopic expression of wg, cli or
abd A, embryos were collected for 1 hour and aged at 25°C for 3-4hours. Heat-shock treatments were administered by floating agarplates in a 37°C water bath for 30 minutes to 1 hour. Plates werereturned to 25°C and aged appropriately.
The UAS-ArmS10 (activated Arm) and UAS-Armwt stocks are fromMark Peifer, who has demonstrated that ectopic expression of ArmAct,but not Armwt, results in the activation of the Wingless signallingpathway in cells (L.-M. Pai, S. Orsulic, A. Bejsovic and M. Peifer,personal communication). The twist-Gal4 stock is described inBaylies et al. (1995).
Immunohistochemistry and RNA in situ hybridizationEmbryos were collected, fixed and processed in a standard manner(Kellerman et al., 1990). Anti-Vasa antibody, provided by theLehmann laboratory, was used at 1:3000; anti-Cli (10H6), generatedwithin the Bonini lab against a peptide fragment, was used at 1:1000;Fasciclin III (mAb 2D5), used at 1:50 (Snow et al., 1989); anti-MuscleMyosin, used at 1:500 (Kiehart and Feghali, 1986); anti-β-galactosi-dase used at 1:1000, Cappel.
Whole-mount in situ hybridization to RNA in embryos using digox-igenin-labeled probes was as described in Tautz and Peifle (1989).Modifications for double labelling with antibodies were as describedin Dougan and DiNardo (1992) in all figures, except for doublelabelling with Fas III or Cli in Fig. 7E-F and Fig. 8C, which was doneas in Azpiazu and Frasch (1993). For cli, an eya type II cDNA probe,described in Bonini et al. (1993), was used. tin and bap cDNA clonesare described in Azpiazu and Frasch (1993); the 412 retrotransposonis described in Brookman et al. (1992); nau cDNA probe is describedin Michelson et al. (1990). Stained embryos were dehydrated andembedded in Araldite for sectioning as described by Leptin andGrunewald (1990). Embryos were photographed using Kodak Ekta160 color film. Slides were scanned and images processed withPhotoshop 3.0. All panels were assembled using Pagemaker 6.0.
Mutant screenPreexisting lethal collections (Nusslein-Volhard et al., 1984; Bier et al.,1989; Karpen and Spradling, 1992) were screened for defects in gonadformation by Vasa and 412 expression. Candidate mutants were subse-quently assayed for global defects in mesodermal patterning by exam-ination of tinman, Fasciclin III and nautilus expression. Due to its rel-atively specific phenotype in SGP development, we focused on clift.
RESULTS
clift is required for the development of somaticgonadal precursorsThe embryonic gonad is composed of 8 to 15 germ cells (Fig.1A, arrowhead) intermingled with approximately 35 somaticmesodermal cells (Fig. 1A, arrow; Sonnenblick, 1941). Thegonad forms when germ cells, visualized with the vasaantibody (brown, Fig. 1B; Hay et al., 1988; Lasko andAshburner, 1990) and SGP cells, visualized by the expressionof the 412 retrotransposon (blue, Fig. 1B; Brookman et al.,1992) cluster together within parasegment (PS) 10 at 10 hoursafter egg laying (AEL). In a screen to identify genes thatregulate the development of SGP cells (see MATERIALS ANDMETHODS), we identified clift (cli), mutations in which leadto the scattering of germ cells throughout posterior regions ofthe embryo at late stages of embryogenesis and the absence ofmost 412-expressing cells (Fig. 1C). The few 412-expressingcells observed in cli mutants are often associated with germcells, indicating that some SGP cells do develop (Fig. 1C,arrowhead). However, 412-expressing cells and associatedgerm cells do not coalesce into a gonad, often remaining
973Somatic gonadal precursor cells in Drosophila
clumped in posterior regions of the embryo. A second SGPmarker, Dwnt-2 (Russell et al., 1992), is also not expressed inSGP cells in cli mutants (data not shown). These resultsdemonstrate that cli is essential for gonadogenesis.
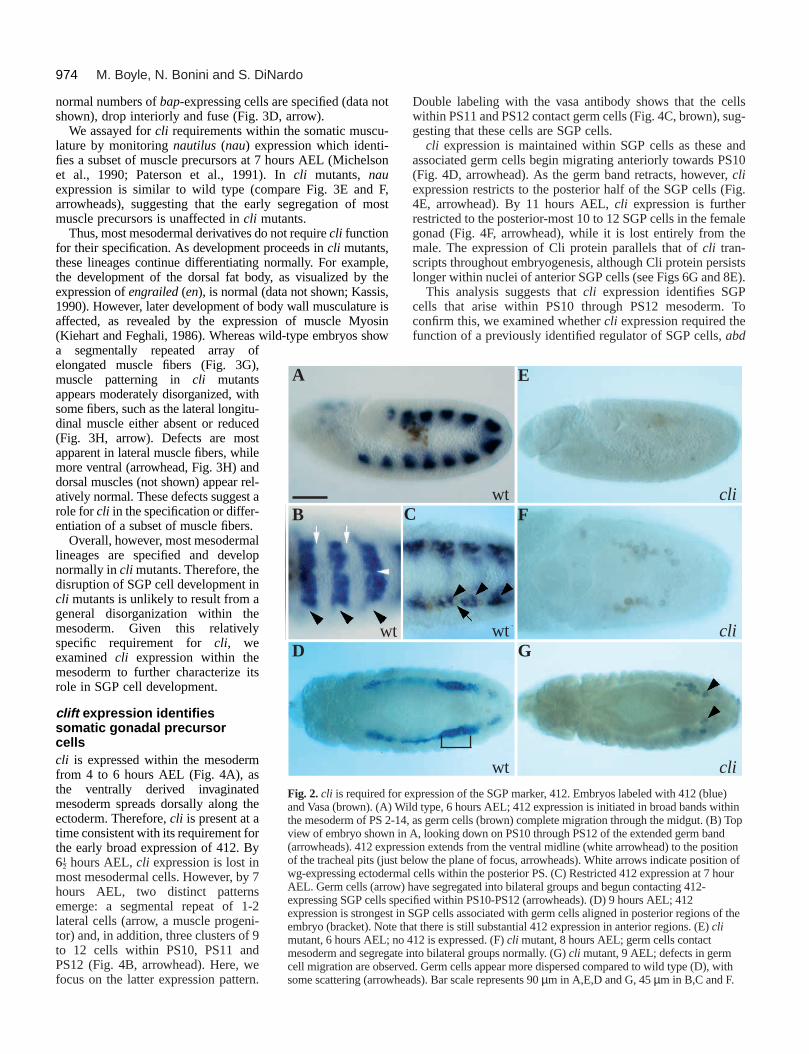
To determine the point at which gonad formation is affectedin cli mutants, we followed the development of germ cells andSGP cells throughout earlier stages in gonadogenesis. The originof the SGP cells, their association with germ cells and coales-cence into a gonad can be followed by 412 expression. At 6 hoursAEL, as germ cells complete migration through the midgut, 412is expressed in broad bands in the mesoderm of PS2-PS14 (Fig.2A). Initially, expression extends from the ventral midlinelaterally to approximately the level of the tracheal pit withinoverlying ectoderm (Fig. 2B). However, as germ cells contactmesoderm and segregate into bilateral groups, ventral-most cellswithin each PS lose 412 expression, such that by 7 hours AEL,only cells within dorsolateral regions of each PS continueexpressing. SGP cells derive from the patches of 412-expressingcells within PS10, PS11 and PS12 (Fig. 2C; arrowhead). Thosewithin PS11 and PS12 are already seen associating with germcells (Fig. 2C; arrow). As development proceeds, SGP cells
A
C
Bwt
wt
cli
Fig. 1. cli is required for somatic gonadal precursor development.Anterior is to the left in all panels. Embryos, 10-12 hours AEL.(A) Coalesced gonad dissected from an embryo carrying the enhancertrap line 68-77. Anti-β-galactosidase visualizes SGP cells (arrow)intermingled with unlabelled germ cells (arrowhead). (B,C) Embryosdouble labeled with the SGP marker 412 (blue) and the germ cellmarker, Vasa (brown). (B) 412-expressing SGP cells clustered withgerm cells at PS10 in wild type (bracket). (C) cli mutant, few 412-expressing SGP cells are present and many germ cells scatter. The fewSGP cells that do develop are often associated with germ cells(arrowhead). Bar represents 21 µm in A, 90 µm in B-C.
within PS11 and PS12, along with associated germ cells, migrateanteriorly towards PS10 and begin to condense. Throughout thisprocess, 412 expression in more anterior PSs diminishes, suchthat by 9 hours AEL, expression is strongest in SGP cells lyingalong PS10/PS11 mesoderm (Fig. 2D, bracket). Finally, by 10hours AEL, germ cells and 412-expressing SGP cells coalesceinto a gonad at PS10 (above). Therefore, the 412 SGP marker isinitially broadly expressed, then focusses down to SGP cellsderived from PS10 through PS12.
In cli mutants, little or no 412 is expressed when SGP cellsare normally segregating within the mesoderm (Fig. 2F). Thissuggests that cli function may be required for an early step inSGP cell development. Strikingly, cli function is also requiredfor the early broad expression of 412 (Fig. 2E), normallyobserved long before SGP cells form.
Despite these dramatic affects on 412 expression, initialgerm cell behavior is normal in cli mutants. Germ cells contactmesoderm and gather into bilateral groups in cli mutants as inwild type (Fig. 2F). They remain associated with lateralmesoderm throughout most of germ band retraction. Defectsin germ cell migration in cli mutants are apparent, however,during migration in early germ-band-retracted embryos.Whereas germ cells normally appear lined up, closely adheringto one another in wild type, germ cells in cli mutants appearmore dispersed, with some germ cells scattering throughoutposterior regions (Fig. 2G, arrowheads). The stage at whichthis occurs is later than that observed in abd A mutants, whichcompletely abolish SGP cells and result in scattering of germcells during retraction (Warrior, 1994). This suggests that SGPcells may form in cli mutants despite the absence of 412expression, thereby accounting for the initially normal germcell behaviour. However, SGP cells may fail to maintain theirfate and as a result, germ cells eventually disperse.
Most other mesodermal cell types develop normallyin clift mutantsOur analysis of gonadogenesis in cli mutants suggests that climay encode a specific regulator of SGP development. Alter-natively, defects in SGP development could result from globaldefects within the mesoderm, suggesting cli functions ingeneral mesodermal patterning. To distinguish these possibili-ties, we examined the specification and differentiation of othermesodermal cell types.
As SGP cells segregate within the mesoderm, progenitorsfor the heart, somatic and visceral musculature becomeapparent. For example, in wild type at 7 hours AEL, heart pro-genitors can be identified by the expression of tinman (tin) incells along the dorsal-most edge of the mesoderm (Fig. 3A,arrow; Azpiazu and Frasch, 1993; Bodmer, 1993). In climutants, tin expression is indistinguishable from wild type(Fig. 3B, arrow), showing that heart progenitors are specifiedin the absence of cli function.
Similarly, cli function is not required for the specification ofthe visceral musculature. Precursors of the visceral muscula-ture originate in patches of approximately 20 cells within eachabdominal PS and are identified by the expression the genebagpipe, (bap, Azpiazu and Frasch, 1993). Once specified,these cells drop internally and fuse to generate a sheet of cellsalong the length of the embryo. This latter step can be visual-ized by the expression of Fasciclin III (Fas III; Fig. 3C, arrow;Snow et al., 1989; Azpiazu and Frasch, 1993). In cli mutants,
974 M. Boyle, N. Bonini and S. DiNardo
C
wt
cli
cli
wt
cli
E
F
wt
G
expression of the SGP marker, 412. Embryos labeled with 412 (blue)ild type, 6 hours AEL; 412 expression is initiated in broad bands within
4, as germ cells (brown) complete migration through the midgut. (B) Topn A, looking down on PS10 through PS12 of the extended germ bandssion extends from the ventral midline (white arrowhead) to the positionbelow the plane of focus, arrowheads). White arrows indicate position ofal cells within the posterior PS. (C) Restricted 412 expression at 7 hour) have segregated into bilateral groups and begun contacting 412-cified within PS10-PS12 (arrowheads). (D) 9 hours AEL; 412 SGP cells associated with germ cells aligned in posterior regions of thehat there is still substantial 412 expression in anterior regions. (E) cli 412 is expressed. (F) cli mutant, 8 hours AEL; germ cells contact into bilateral groups normally. (G) cli mutant, 9 AEL; defects in germed. Germ cells appear more dispersed compared to wild type (D), withads). Bar scale represents 90 µm in A,E,D and G, 45 µm in B,C and F.
normal numbers of bap-expressing cells are specified (data notshown), drop interiorly and fuse (Fig. 3D, arrow).
We assayed for cli requirements within the somatic muscu-lature by monitoring nautilus (nau) expression which identi-fies a subset of muscle precursors at 7 hours AEL (Michelsonet al., 1990; Paterson et al., 1991). In cli mutants, nauexpression is similar to wild type (compare Fig. 3E and F,arrowheads), suggesting that the early segregation of mostmuscle precursors is unaffected in cli mutants.
Thus, most mesodermal derivatives do not require cli functionfor their specification. As development proceeds in cli mutants,these lineages continue differentiating normally. For example,the development of the dorsal fat body, as visualized by theexpression of engrailed (en), is normal (data not shown; Kassis,1990). However, later development of body wall musculature isaffected, as revealed by the expression of muscle Myosin(Kiehart and Feghali, 1986). Whereas wild-type embryos showa segmentally repeated array ofelongated muscle fibers (Fig. 3G),muscle patterning in cli mutantsappears moderately disorganized, withsome fibers, such as the lateral longitu-dinal muscle either absent or reduced(Fig. 3H, arrow). Defects are mostapparent in lateral muscle fibers, whilemore ventral (arrowhead, Fig. 3H) anddorsal muscles (not shown) appear rel-atively normal. These defects suggest arole for cli in the specification or differ-entiation of a subset of muscle fibers.
Overall, however, most mesodermallineages are specified and developnormally in cli mutants. Therefore, thedisruption of SGP cell development incli mutants is unlikely to result from ageneral disorganization within themesoderm. Given this relativelyspecific requirement for cli, weexamined cli expression within themesoderm to further characterize itsrole in SGP cell development.
clift expression identifiessomatic gonadal precursorcellscli is expressed within the mesodermfrom 4 to 6 hours AEL (Fig. 4A), asthe ventrally derived invaginatedmesoderm spreads dorsally along theectoderm. Therefore, cli is present at atime consistent with its requirement forthe early broad expression of 412. By6G hours AEL, cli expression is lost inmost mesodermal cells. However, by 7hours AEL, two distinct patternsemerge: a segmental repeat of 1-2lateral cells (arrow, a muscle progeni-tor) and, in addition, three clusters of 9to 12 cells within PS10, PS11 andPS12 (Fig. 4B, arrowhead). Here, wefocus on the latter expression pattern.
A
B
Dwt
Fig. 2. cli is required forand Vasa (brown). (A) Wthe mesoderm of PS 2-1view of embryo shown i(arrowheads). 412 expreof the tracheal pits (just wg-expressing ectodermAEL. Germ cells (arrowexpressing SGP cells speexpression is strongest inembryo (bracket). Note tmutant, 6 hours AEL; nomesoderm and segregatecell migration are observsome scattering (arrowhe
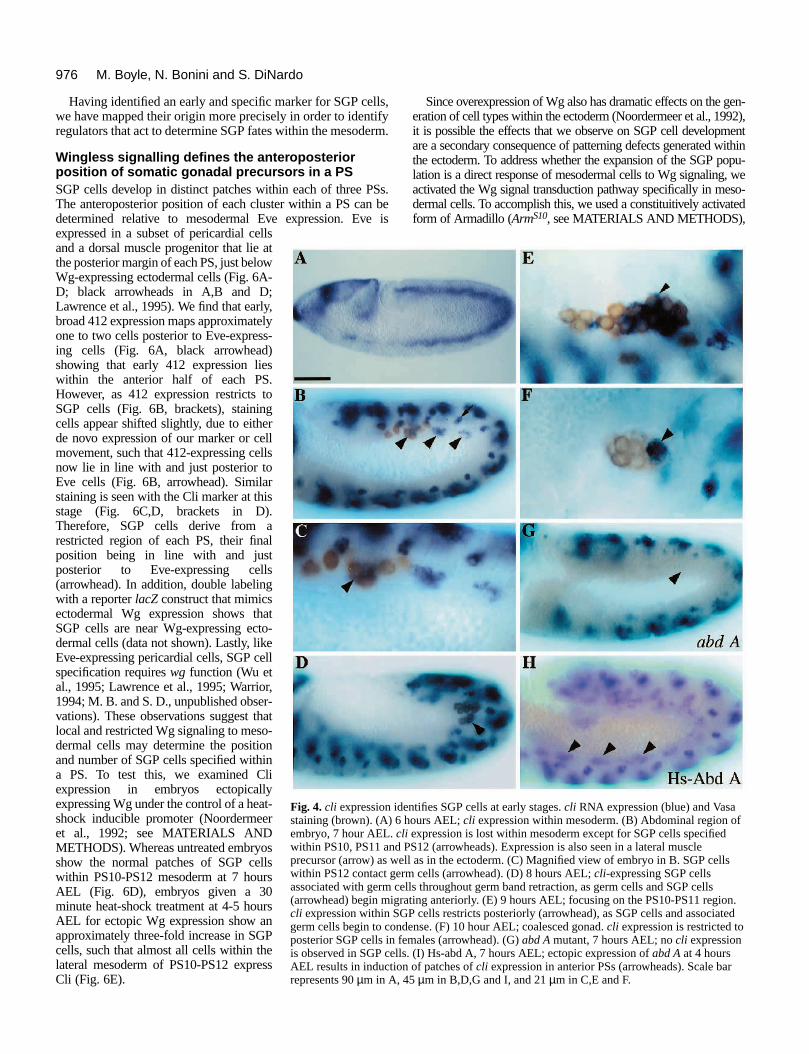
Double labeling with the vasa antibody shows that the cellswithin PS11 and PS12 contact germ cells (Fig. 4C, brown), sug-gesting that these cells are SGP cells.
cli expression is maintained within SGP cells as these andassociated germ cells begin migrating anteriorly towards PS10(Fig. 4D, arrowhead). As the germ band retracts, however, cliexpression restricts to the posterior half of the SGP cells (Fig.4E, arrowhead). By 11 hours AEL, cli expression is furtherrestricted to the posterior-most 10 to 12 SGP cells in the femalegonad (Fig. 4F, arrowhead), while it is lost entirely from themale. The expression of Cli protein parallels that of cli tran-scripts throughout embryogenesis, although Cli protein persistslonger within nuclei of anterior SGP cells (see Figs 6G and 8E).
This analysis suggests that cli expression identifies SGPcells that arise within PS10 through PS12 mesoderm. Toconfirm this, we examined whether cli expression required thefunction of a previously identified regulator of SGP cells, abd
975Somatic gonadal precursor cells in Drosophila
A
C
E
G H
F
B
wt cli
wt
wt
wt
D
cli
cli
cli
Fig. 3. cli function within the mesoderm isrelatively specific to SGP cell development.(A,B) Lateral view, 7 hours AEL, tin RNAexpression. (A) Wild type; tin expression inheart precursors at the dorsal crest of themesoderm (arrow). (B) cli mutant; tin-expressing heart precursors are specifiednormally (arrow). (C,D) Fas III expression indeveloping visceral mesoderm (arrow) in wildtype (C) and cli mutants (D). (E,F) nauexpression reveals a subset of developingmuscle precursors in wild type (E, arrowheadpoints to lateral cluster) and in cli mutants (F),indicating that most, if not all muscleprecursors develop normally. The slightvariability in staining is likely due to thedynamic expression of nau at this stage(Michelson et al., 1990). (G,H) 15 hours AEL;anti-Muscle Myosin staining reveals the finalmuscle pattern, repeated in each abdominal PS(G). Wild type; arrow points to the cluster offour lateral longitudinal muscle fibers. (H) climutant; the final muscle pattern is disorganizedin lateral regions. The normal cluster of fourlateral longitudinal muscles are either absent orreduced (arrow). In contrast, cli does not affectdorsal muscle fibers (not shown), or ventraloblique fibers (arrowhead in H), which areobscured in G by the β-galactosidase stainingfrom a balancer chromosome. Bar represents90 µm in A,B and 45 µm in C-H.
A (Lewis, 1978; Szabad and Nöthiger, 1992; Cumberledge etal., 1992; Brookman et al., 1992; Boyle and DiNardo, 1995;Greig and Akam, 1995). In abd A mutants, no cli expressionwithin PS10-12 is detected at 7 hours AEL (Fig. 4G), althoughearly, broad expression of cli within the mesoderm is unaf-fected (data not shown). In addition, when abd A is ectopicallyexpressed at 4 hours AEL, cli expression is induced in patchesof mesodermal cells in anterior PSs (Fig. 4H). These ectopiccli-expressing cells differentiate as SGP cells, since theyexpress the differentiation marker escargot at later stages (datanot shown; Boyle and DiNardo, 1995). Therefore, cli is anearly and specific marker of SGP cells.
clift is required to maintain somatic gonadalprecursor fates within the mesodermSince cli is initially expressed throughout the mesoderm, thenrestricts to those cells eventually selected as SGP cells and isrequired for their development, we wondered whether clifunction is required for the initial segregation of SGP cells. Toassay whether SGP cells fail to segregate in cli mutants, orsegregate but fail to maintain their fate, we examined cliexpression in embryos mutant for the cliIIE allele. This allele,
although a strong loss of function allele, produces a transcript,thereby allowing us to examine early stages of SGP cell devel-opment. In the absence of cli function, we see cli expressionin PS10-PS12 mesoderm at 7 hours AEL (Fig. 5B, arrow-heads). In addition, these cells are often observed contactinggerm cells (data not shown). Thus, cli function is not essentialfor the initial selection of SGP cells out of the mesoderm. Con-sistent with this, ectopic expression of cli by a heat-shockinducible promoter at various stages of development in wild-type embryos does not alter the number of SGP cells specified(data not shown). Therefore, cli function is not sufficient topromote SGP cell fates, nor is it an essential component of theinitial selection process. cli function is required, however, tomaintain SGP fates within the mesoderm. Examination of laterstages in development shows that cli expression drops off dra-matically within the mesoderm such that fewer cells can bedetected by 8 hours AEL compared to wild type (Fig. 5C,D).As seen with 412 expression, these few cli-positive cells oftenadhere to germ cells (not shown). However, germ cells andSGP cells fail to complete their migration and coalescence intoa gonad. Therefore, cli function is required to maintain SGPfates within the mesoderm.
976 M. Boyle, N. Bonini and S. DiNardo
entifies SGP cells at early stages. cli RNA expression (blue) and Vasa hours AEL; cli expression within mesoderm. (B) Abdominal region ofli expression is lost within mesoderm except for SGP cells specified PS12 (arrowheads). Expression is also seen in a lateral musclell as in the ectoderm. (C) Magnified view of embryo in B. SGP cellsrm cells (arrowhead). (D) 8 hours AEL; cli-expressing SGP cellsells throughout germ band retraction, as germ cells and SGP cellsrating anteriorly. (E) 9 hours AEL; focusing on the PS10-PS11 region.GP cells restricts posteriorly (arrowhead), as SGP cells and associateddense. (F) 10 hour AEL; coalesced gonad. cli expression is restricted toemales (arrowhead). (G) abd A mutant, 7 hours AEL; no cli expressions. (I) Hs-abd A, 7 hours AEL; ectopic expression of abd A at 4 hoursn of patches of cli expression in anterior PSs (arrowheads). Scale bar 45 µm in B,D,G and I, and 21 µm in C,E and F.
Having identified an early and specific marker for SGP cells,we have mapped their origin more precisely in order to identifyregulators that act to determine SGP fates within the mesoderm.
Wingless signalling defines the anteroposteriorposition of somatic gonadal precursors in a PSSGP cells develop in distinct patches within each of three PSs.The anteroposterior position of each cluster within a PS can bedetermined relative to mesodermal Eve expression. Eve isexpressed in a subset of pericardial cellsand a dorsal muscle progenitor that lie atthe posterior margin of each PS, just belowWg-expressing ectodermal cells (Fig. 6A-D; black arrowheads in A,B and D;Lawrence et al., 1995). We find that early,broad 412 expression maps approximatelyone to two cells posterior to Eve-express-ing cells (Fig. 6A, black arrowhead)showing that early 412 expression lieswithin the anterior half of each PS.However, as 412 expression restricts toSGP cells (Fig. 6B, brackets), stainingcells appear shifted slightly, due to eitherde novo expression of our marker or cellmovement, such that 412-expressing cellsnow lie in line with and just posterior toEve cells (Fig. 6B, arrowhead). Similarstaining is seen with the Cli marker at thisstage (Fig. 6C,D, brackets in D).Therefore, SGP cells derive from arestricted region of each PS, their finalposition being in line with and justposterior to Eve-expressing cells(arrowhead). In addition, double labelingwith a reporter lacZ construct that mimicsectodermal Wg expression shows thatSGP cells are near Wg-expressing ecto-dermal cells (data not shown). Lastly, likeEve-expressing pericardial cells, SGP cellspecification requires wg function (Wu etal., 1995; Lawrence et al., 1995; Warrior,1994; M. B. and S. D., unpublished obser-vations). These observations suggest thatlocal and restricted Wg signaling to meso-dermal cells may determine the positionand number of SGP cells specified withina PS. To test this, we examined Cliexpression in embryos ectopicallyexpressing Wg under the control of a heat-shock inducible promoter (Noordermeeret al., 1992; see MATERIALS ANDMETHODS). Whereas untreated embryosshow the normal patches of SGP cellswithin PS10-PS12 mesoderm at 7 hoursAEL (Fig. 6D), embryos given a 30minute heat-shock treatment at 4-5 hoursAEL for ectopic Wg expression show anapproximately three-fold increase in SGPcells, such that almost all cells within thelateral mesoderm of PS10-PS12 expressCli (Fig. 6E).
Fig. 4. cli expression idstaining (brown). (A) 6embryo, 7 hour AEL. cwithin PS10, PS11 andprecursor (arrow) as wewithin PS12 contact geassociated with germ c(arrowhead) begin migcli expression within Sgerm cells begin to conposterior SGP cells in fis observed in SGP cellAEL results in inductiorepresents 90 µm in A,
Since overexpression of Wg also has dramatic effects on the gen-eration of cell types within the ectoderm (Noordermeer et al., 1992),it is possible the effects that we observe on SGP cell developmentare a secondary consequence of patterning defects generated withinthe ectoderm. To address whether the expansion of the SGP popu-lation is a direct response of mesodermal cells to Wg signaling, weactivated the Wg signal transduction pathway specifically in meso-dermal cells. To accomplish this, we used a constituitively activatedform of Armadillo (ArmS10, see MATERIALS AND METHODS),
977Somatic gonadal precursor cells in Drosophila
Cwt
wt
cliIIE
cliIIE
A B
D Fig. 5. SGP cells are specified, but fail tomaintain their fate in cli mutants. (A) Wildtype, 7 hours AEL; cli is expressed in SGPcells (B) cliIIE mutant, 7 hours AEL; cli-expressing cells are present. (C) Wild type, 8hours AEL; cli expression is maintained inSGP cells (arrows). (D) cliIIE mutant, 8 hoursAEL; fewer cli-expressing SGP cells arepresent. Scale bar represents 45 µm in allpanels.
an essential component of the Wg signal transduction pathway (L.-M. Pai, S. Orsulic, A. Bejsovic and M. Peifer, personal communi-cation; Peifer et al., 1991). To drive expression from a UAS-ArmS10
construct specifically within the mesoderm, we used a twist-GAL4line (Staehling-Hampton et al., 1994; Baylies and Bate, 1996).Embryos carrying both constructs showed an increase in SGP cellnumber similar to that seen in hs-wg embryos (Fig. 6F). In contrast,similar expression of a UAS-Armwt construct had no effect (data notshown). Extra Cli-expressing cells generated by ectopic Wg pro-duction or by activating the Wg transduction pathway truly are SGPcells as they associate with germ cells and populate the gonad at laterstages of development (Fig. 6H, compare with G). We conclude thatmesodermal cells can respond directly to Wg signaling for the spec-ification of SGP cells within the PS. Since Wg is normally secretedfrom a narrow stripe of cells within overlying ectoderm, the restrictedsource of this ligand may define the anteroposterior position andpossibly the number of SGP cells within each PS.
Somatic gonadal precursor cells develop adjacent tovisceral mesodermWe next investigated the dorsoventral positioning of SGP cells.An early event in the establishment of distinct fates along thisaxis occurs with the split into dorsal and ventral domains. Thisregionalization relies upon induction by the transforming growthfactor β homologue dpp (Padgett et al., 1987; Staehling-Hampton et al., 1994; Frasch, 1995). dpp, expressed in dorsalectoderm, maintains the expression of tin in underlyingmesoderm, thereby allowing the specification of dorsal cell typessuch as heart and visceral mesoderm (diagramed in Fig. 7B;Azpiazu and Frasch, 1993; Bodmer, 1993). To identify regula-tors that control the dorsoventral aspect of SGP specification, wesectioned stained embryos to determine the position at whichSGP cells develop relative to other mesodermal derivatives.
cli is initially expressed throughout the mesoderm as theinvaginated mesoderm flattens into a monolayer and beginsspreading dorsally along the ectoderm (data not shown). As themigrating mesodermal cells extend to their dorsal limit (Fig.7C, arrowhead), cli expression is lost from dorsal-most cellssuch that, by 5 hours AEL, expression is limited to ventral andlateral mesodermal cells.
This restriction in cli expression coincides with the subdiv-ision of the mesoderm into dorsal and ventral domains throughinduction by dpp. To map the dorsal-most limit of cliexpression at this stage, we double-labeled embryos carryinga dpp-lac Z reporter gene (Huang et al., 1993). cli expressionwithin mesoderm extends to and overlaps slightly with dpp-expressing ectodermal cells (Fig. 7C, arrow). Subsequent tothis, cli expression within the mesoderm declines dramatically,except for expression in SGP cells, here shown contactinggerm cells (Fig. 7D, arrowhead).
These SGP cells are likely to derive from the dorsal-mostcli-expressing cells near overlying dpp-expressing ectodermalcells (Fig. 7B). This would place the SGP cells adjacent toprospective visceral mesoderm, a dorsal mesodermal cell type.To confirm this, we examined 412 expression, which initiatesexpression slightly later than cli but persists longer and, thus,can be mapped relative to the visceral mesoderm marker, FasIII. Indeed most Fas III-expressing cells are dorsal to the 412expression domain (Fig. 7E). Note, however, that at this stage,the patches of visceral mesodermal cells are in the process ofdropping interiorly, so that some Fas III-expressing cells lieadjacent and interal to 412-expressing cells. At the same time,412-expressing cells appear to extend further dorsally. In sub-sequent stages, high level 412 expression is maintained in SGPcells lying adjacent to Fas III-staining cells (Fig. 7F,arrowhead), now positioned interior and adjacent to themidgut. Although we cannot exclude the possibility of de novo412 expression, the simplest hypothesis is that the SGP cellsderive from the leading edge of 412-expressing cells. Thus, webelieve that SGP cells originate in regions just ventral to thevisceral mesoderm precursors, near the boundary of dppexpression in overlying ectodermal cells.
Somatic gonadal precursor development dependson tinmanSince tin appears to mediate the role of dpp in specifying dorsalmesoderm, we tested whether tin function is also required forSGP cell development. In tin mutants, few or no SGP cells aredetected at 7 hours AEL by cli expression (Fig. 8B), althoughearly, broad cli expression within the mesoderm is not affected
978 M. Boyle, N. Bonini and S. DiNardo
nteroposterior position of SGP cells within a PS. (A,B) 412 expressionstaining (brown). Embryos in A-D double labeled with Eved D) expressed in a subset of pericardial cells and a dorsal musclee dorsal posterior margin of each PS. (A) Embryo 7 hour AEL. Early,2 lies in the anterior half of each PS, one or two cells posterior to Eve- arrows indicate position of ectodermal Wg expression, while whitesition of tracheal pit. (B) Magnified view of PS10-PS12 mesoderm atrs AEL. Restricted 412 expression in SGP cells (brackets), now overlapping and just posterior to Eve-expressing cells. (C,D) Similarlynti-Cli staining. (D) Magnified view of boxed portion in C, focusing on. (E) Hs-Wg; ectopic wg expression induced at 4 hours AEL by heat- in increased numbers of Cli-staining SGP cells specified throughout(arrowheads). (F) twi-Gal 4/UAS-ArmS10, 7 hours AEL; a similaropulation (arrowheads) is observed. (G) Wild type, 12 hours AEL;
nad. (H) twi-Gal4/UAS-ArmS10, 12 hours AEL; coalesced gonad. Scalen A and C, 21 µm in B, D-F.
S10
S10
(data not shown). In addition, germ cells are scattered at latestages and no gonad forms (data not shown). Thus, SGP cells,like other dorsal mesodermal cell types, require tin function.
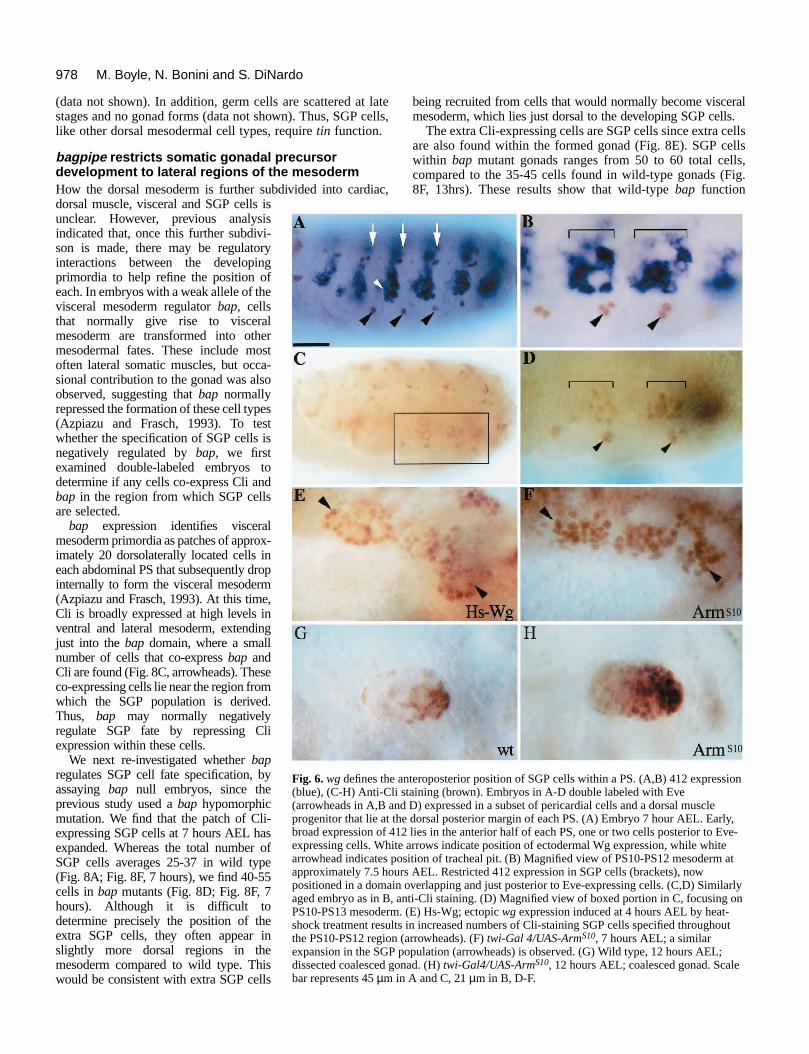
bagpipe restricts somatic gonadal precursordevelopment to lateral regions of the mesodermHow the dorsal mesoderm is further subdivided into cardiac,dorsal muscle, visceral and SGP cells isunclear. However, previous analysisindicated that, once this further subdivi-son is made, there may be regulatoryinteractions between the developingprimordia to help refine the position ofeach. In embryos with a weak allele of thevisceral mesoderm regulator bap, cellsthat normally give rise to visceralmesoderm are transformed into othermesodermal fates. These include mostoften lateral somatic muscles, but occa-sional contribution to the gonad was alsoobserved, suggesting that bap normallyrepressed the formation of these cell types(Azpiazu and Frasch, 1993). To testwhether the specification of SGP cells isnegatively regulated by bap, we firstexamined double-labeled embryos todetermine if any cells co-express Cli andbap in the region from which SGP cellsare selected.
bap expression identifies visceralmesoderm primordia as patches of approx-imately 20 dorsolaterally located cells ineach abdominal PS that subsequently dropinternally to form the visceral mesoderm(Azpiazu and Frasch, 1993). At this time,Cli is broadly expressed at high levels inventral and lateral mesoderm, extendingjust into the bap domain, where a smallnumber of cells that co-express bap andCli are found (Fig. 8C, arrowheads). Theseco-expressing cells lie near the region fromwhich the SGP population is derived.Thus, bap may normally negativelyregulate SGP fate by repressing Cliexpression within these cells.
We next re-investigated whether bapregulates SGP cell fate specification, byassaying bap null embryos, since theprevious study used a bap hypomorphicmutation. We find that the patch of Cli-expressing SGP cells at 7 hours AEL hasexpanded. Whereas the total number ofSGP cells averages 25-37 in wild type(Fig. 8A; Fig. 8F, 7 hours), we find 40-55cells in bap mutants (Fig. 8D; Fig. 8F, 7hours). Although it is difficult todetermine precisely the position of theextra SGP cells, they often appear inslightly more dorsal regions in themesoderm compared to wild type. Thiswould be consistent with extra SGP cells
Fig. 6. wg defines the a(blue), (C-H) Anti-Cli (arrowheads in A,B anprogenitor that lie at thbroad expression of 41expressing cells. Whitearrowhead indicates poapproximately 7.5 houpositioned in a domainaged embryo as in B, aPS10-PS13 mesodermshock treatment resultsthe PS10-PS12 region expansion in the SGP pdissected coalesced gobar represents 45 µm i
being recruited from cells that would normally become visceralmesoderm, which lies just dorsal to the developing SGP cells.
The extra Cli-expressing cells are SGP cells since extra cellsare also found within the formed gonad (Fig. 8E). SGP cellswithin bap mutant gonads ranges from 50 to 60 total cells,compared to the 35-45 cells found in wild-type gonads (Fig.8F, 13hrs). These results show that wild-type bap function
979Somatic gonadal precursor cells in Drosophila
F
B
g
Cli/Dpp
A
E
D
Cli/Vasa
412/FasIII 412/FasIII
C
Fig. 7. SGP cells arise just ventral to visceral mesoderm. (A) 5 hoursAEL; cli RNA expression; line indicates position of transversesections shown in C-F. (B) Diagram of transverse section of embryo,7 hours AEL, illustrating early subdivisions within the mesoderm.Due to the extended germ band, mesodermal cells lie within both thetop and bottom half of each section. (g) Midgut. The dorsal-mostmesodermal cells underlie dpp-expressing ectoderm, indicated by thethick black lines. dpp maintains tin expression within underlyingmesoderm, which subsequently generates heart precursors (red),visceral mesoderm (yellow) and dorsal muscles (not shown). SGPcells (blue) lie adjacent to visceral mesoderm. (C-F) 10 µm thicktransverse sections. (C) 5 hours AEL; cli RNA (blue) and dpp-lacZ(brown), which marks the extent of dpp expression within theectoderm (arrow). Arrowhead marks the dorsal limit of the extendedmesoderm; asterix marks the ventral midline. cli expression extendsto, and overlaps slightly with adjacent dpp-expressing ectoderm.(D) 7 hours AEL; cli expression (blue) within the mesodermdiminishes dramatically except for a small number of SGP cellswithin lateral regions (arrowhead) that begin contacting germ cells(brown). Arrows point to cli expression in CNS and muscleprecursors. (E,F) 412 (blue) and the visceral mesoderm marker FasIII (brown). (E) 6 hours AEL; 412-expressing cells are ventral andadjacent to visceral mesoderm (arrow) which is in the process ofdropping interiorly. (F) 7 hours AEL; 412 expression has restricted toSGP cells (arrowhead) in lateral mesoderm. These cells lie adjacentto visceral mesoderm which has dropped interiorly completely andnow contacts the gut. Bar represents 90 µm for A and 47 µm for C-F.
represses SGP cell development, confirming previous observa-tions (Azpiazu and Frasch, 1993). Thus, the dorsoventralposition of SGP specification is refined by negative interactionswith the dorsally adjacent visceral mesoderm precursors.
DISCUSSION
Gametogenesis entails a complex developmental program thatensures the continuous generation of mature gametes fromgermline stem cells. This differentiation program is supportedby a variety of somatic cell types, each of which derives fromthe population of SGP cells found clustered in the embryonicgonad. To begin to investigate how these cells differentiate andsustain the germline, we have been focusing on the develop-ment of SGP cells during embryogenesis. Our studies of thegeneration of SGP cells within the mesoderm not only uncoveressential regulators of their specification and differentiation butalso reveal novel aspects of the general mechanism wherebycell fates are allocated within the mesoderm.
Segregation of somatic gonadal precursor cellswithin the mesodermUntil recently, little has been known about the mechanismwhereby SGP fates are determined within the mesoderm.Previous analysis illustrated that they arise as patches of cellsfrom three posterior abdominal PS at mid embryogenesis(Brookman et al., 1992; Warrior, 1994; Boyle and DiNardo,1995; Greig and Akam, 1995). In addition, using 412expression as a marker, these studies established that the restric-tion of SGP cell specification to these PSs depends on thefunction of the homeotic gene abd A. Using the more specificmarker cli, we have confirmed these results and demonstrateda role for abd A in SGP cells as these cells first form. Thus,ectopic expression of abd A results in SGP cells being specifiedwithin more anterior PS, while loss of abd A expressionabolishes SGP cells. Thus, abd A function detemines the com-petence of a PS for SGP cell induction. abd A does not,however, instruct SGP cell number or position within a PS,since SGP cells induced in anterior segments still show reiter-ated patches of expression. Therefore, abd A function is per-missive. Here, we show that the anteroposterior and dorsoven-tral position of SGP cells within a PS are established byectodermal signals generated by the secreted growth factor Wg,coupled with a gene regualtory hierarchy within the mesoderm.
Wingless defines the anteroposterior position ofsomatic gonadal precursor cells within a PS Our results indicate that restricted Wg expression determines theposition at which SGP cells develop within a PS. While loss ofwg function abolishes SGP cells (Warrior, 1994; M. B. and S.D., unpublished observations), global expression of Wg, or acti-vation of its signal transduction pathway within the mesoderm,results in almost all dorsolateral mesodermal cells within the PSacquiring SGP fates and populating the gonad. Thus, Wg inputinstructs SGP fate within PS10-PS12 mesoderm as long as thesecells have acquired appropriate dorsoventral cues.
Wg is required in the generation of other cell types within themesoderm, although this requirement appears to be permissiverather than instructive (Lawrence et al., 1995; Baylies et al.,1995; Ranganayakulu et al., 1996; Park et al., 1996). Forexample, absence of wg function results in the loss of nau
980 M. Boyle, N. Bonini and S. DiNardo
D
bap
tinwt
d
v
wt
bap
B
F
pment requires tin function and is negatively regulated by bap. (A-wn). (A) Wild type, 7 hours AEL; Cli-expressing SGP cells (arrowhead).AEL; few or no SGP cells are specified (arrow points to single Cli-thin PS12). (C) Wild type, PS10-12 region, 6 hour AEL, double labeledp RNA (blue). Dorsal (d) is down, ventral (v), up. A small number of
t lie at the ventral border of the bap domain are doubly labeledull mutant, 7 hours AEL; increased numbers of SGP cells are specifiedull mutant, 13 hours AEL; dissected gonad showing extra SGP cells arermed gonad (arrow in E). (F) Histogram showing average number of3 wild-type (s.d.=6.93) and 13 bap mutant embryos (s.d.=5.68) at 7umbers are also found incorporated into the gonad at 13 hours AELnts (s.d.=4.98) with 10 wild-type embryos (s.d.=6.05). One-tailed t-testsents 45 µm in A, B and D; 20 µm in C and E.
expression in a subset of muscle progenitors, while uniform Wgexpression restores them to wg mutant embryos. Ectopic Wgexpression, however, has no affect on the number or position ofnau-expressing muscle progenitors specified (Baylies et al.,1995). Wg also appears to function permissively in the devel-opment of Eve-expressing heart precursors and a dorsal muscleprogenitor. Ectopic Wg expression expands the population ofEve-expressing cells (Lawrence et al., 1995), just as for SGPcells. However, unlike for SGP cells, ectopic Eve-expression istransient and remains restricted to the posterior-most cells withineach PS. Thus, Wg input is not sufficient to position these cellfates within a PS. Indeed, more recent evidence suggests that Wginput determines only the competence of a region for the speci-fication of Eve cells, allowing other signaling pathways tospecify cell fate (A. Michelson, personal communication).
Since our results demonstrate that Wg signaling can be instruc-tive, at least for SGP cell fates, taken together these observationsextend our understanding of how different cell types aregenerated within the mesoderm. One signaling molecule, Wg,can act as a competence factor for one celltype while instructing the fate of another.
Although it is not known what regulatesthe different responses to Wg within thesemesodermal lineages, in the epidermis,Wg signaling at different times in devel-opment has distinct outputs. Early, Wgsignaling maintains the expression of atarget gene, engrailed, whereas later Wginput specifies engrailed cell fate (Douganand DiNardo, 1992). Therefore, it ispossible that the distinct responses to Wgsignalling within the mesoderm are aresult of Wg action at different times indevelopment. However, Wg appears to berequired during the same early time periodto specify heart precursors as SGP cells(Wu et al., 1995; M. B. and S. D., unpub-lished observations).
Given that our mapping places someSGP cells several cell diameters posteriorto Wg-expressing cells, perhaps Wg actsindirectly, generating a secondary signalto determine SGP fates. If so, such asignal must originate within themesoderm, since activation of the Wgpathway specifically within mesoderm issufficient to specify SGP fates. Alterna-tively, Wg may signal directly todetermine SGP position within a PS,spreading across several cell diameters. Itis possible, however, that spread of Wgprotein is not required. SGP cells may bemore closely positioned to Wg-express-ing cells earlier, when Wg is required togenerate SGP fate. Rearrangementswithin the mesoderm may have takenplace to position SGP cells further fromWg-expressing cells by the time thesecells can be identified by our markers.
The SGP population expandsprimarily along the anteroposterior axis
C
Cli/Bap
A
E
Fig. 8. SGP cell develoE) anti-Cli staining (bro(B) tin mutant, 7 hours expressing SGP cell wiwith Cli (brown) and baCli-expressing cells tha(arrowheads). (D) bap n(arrowheads). (E) bap nalso found within the foSGP cells specified in 1hours AEL. Increased ncomparing 10 bap mutaP<0.05. Scale bar repre
in response to global wg expression. Therefore, additionalregulators act along the dorsoventral axis to specify SGP cells.
Positioning somatic gonadal precursors along thedorsoventral axis requires tinman function and isnegatively regulated by bagpipeOur results show that SGP cells, like other dorsal cell typessuch as visceral mesoderm and heart precursors, requires tinfunction (Bodmer, 1993; Azpiazu and Frasch, 1993). Theseresults suggest that the initial positioning of SGP cells alongthe dorsoventral axis relies on a regulatory cascade that estab-lishes a domain of mesodermal cells competent to developdorsal cell fates. This regionalization is determined by aninductive signal by overlying dpp-expressing ectodermal cells(Staehling-Hampton et al., 1994; Frasch, 1995; Maggert et al.,1995). dpp maintains tin expression in underlying mesoderm,thereby allowing the specification of dorsal cell types.
Once this initial subdivision has occurred, distinct progenitorsfor each of the cell types derived from this region are precisely
981Somatic gonadal precursor cells in Drosophila
defined. How this occurs is unclear. Perhaps there is a continuedrequirement for dpp, acting in a second step to establish differentfates along the dorsoventral axis in a concentration-dependentmanner, such as has been proposed for the determination of cellfates by dpp in the ectoderm (Ferguson and Anderson, 1992;Wharton et al., 1993). At present, however, there is littleevidence of such a mechanism acting within the mesoderm. Thepositioning of visceral precursors within dorsal mesodermappears to be determined simply by the extent of dpp expressionin overlying ectoderm. The ventral limit of the visceralmesoderm coincides with that of ectodermal dpp and broaden-ing dpp expression within ectoderm results in the concomitantexpansion of visceral mesoderm (Staehling-Hampton et al.,1994; Frasch, 1995). The enlarged dpp expression domain,however, has no effect on the number or position of cardiac ordorsal muscle precursors (Frasch, 1995). Therefore, positioningof these progenitors must rely on additional regulators to furthersubdivide the mesoderm. In fact, some determinants for cardiacmesoderm are being identified, indicating that a heirarchy ofsignals act in combination with dpp to determine these fates (A.Michelson, personal communication).
In our analysis, the SGP cells are unambiguously identifiedfor the first time by the specific expression of Cli in smallclusters within PS10, PS11 and PS12 at 7 hours AEL. Theirposition is within dorsal mesoderm, adjacent to the internallylocated visceral mesoderm. However, we place the origin ofSGP cells just ventral to visceral mesoderm. First, SGP cells arefound at the same anteroposterior register as visceral mesoderm.Second, initial mesodermal expression of both the 412 and climarkers extends from ventral mesoderm laterally to the domainof visceral mesoderm cells, as marked by bap and Fasciclin IIIexpression. As visceral mesoderm drops internally, the leadingedge of 412-expressing cells extends further dorsally, leavingthe final position of the SGP cells at these slightly more dorsalpositions. Although it remains possible that there is de novo 412and Cli expression at these more dorsal positions, we view thisas less likely. Recent data indicate that all other cells in thedorsal mesoderm at this anteroposterior register are allocated tovisceral or heart progenitor fate, leaving no mesodermal cellsto take on the SGP fate (Azpiazu et al., 1996).
At present, it is still unclear what might specify SGP cellfate at the ventral border of visceral mesoderm. Our results do,however, indicate that cross regulatory interactions areimportant in refining this patterning. A significant increase inthe number of SGP cells specified is observed when thedorsally adjacent visceral mesoderm is eliminated. Such cross-regulatory interactions are central to patterning within manytissues (Kellerman et al., 1990; Brook and Cohen, 1996; Jiangand Struhl, 1996). Thus, we view it likely that the repressionof SGP fate by bap reveals a common mechanism usedbetween nearby developing mesodermal primordia to ensurethe precise allocation of each within the mesoderm.
clift function within somatic gonadal precursors Our analysis of cli expression shows that, after an initial burstthroughout the mesoderm, cli expression restricts to SGP cells.Despite this broad cli expression and its requirement for the earlyexpression of 412, our results indicate that cli is not essential forthe initial selection of SGP cells within the mesoderm. In theabsence of cli function, cli-expressing SGP cells do form andinitially associate with germ cells, indicating that early steps in
their development have occurred. Therefore, it is unclear whatrole, if any, is played by early mesodermal expression of cli.Perhaps cli function at this stage is functionally redundant withan as yet unidentified gene that, like cli, acts to establish SGPcell fates, but does so independently of initiating 412 expression.
Our results do, however, reveal an essential requirement forcli in the maintenance of SGP cells. Shortly after SGP cells arespecified in cli mutants, we observe a reduction in the numberof cli-expressing SGP cells, such that by germ band retraction,few SGP cells remain. Those few SGP cells that are present,however, fail to complete their migration to the position of gonadformation. As a consequence, germ cells are not incorporatedinto a gonad, remaining scattered throughout posterior regionsof the embryo. Although our experiments do not allow us todetermine the ultimate fate of the mutant SGP cells, perhapsSGP cells fail to differentiate properly and as a consequence die.
The mechanism whereby cli maintains SGP cell fates isunclear. The molecular nature of cli provides little insight intoits function, as it encodes a novel nuclear protein with no obviousmotifs (Bonini et al., 1993). Previous studies have shown that clifunctions elsewhere, for instance, in head morphogenesis(Nusslein-Volhard et al., 1984), and for the differentiation and/orsurvival of cells within the eye imaginal disk (Bonini et al.,1993). In viable cli mutants, the onset of photoreceptor celldifferentiation fails to occur. As a consequence, cells die by pro-grammed cell death. The targets of cli function, however, withinthese other tissues, as in SGP cells, are currently unknown.
At present, we do know a little about the regulatory pathwayleading to the specification and differentiation of SGP cells.Previous studies have suggested that abd A function is requiredin a second step in gonadogenesis, distinct from its earlier rolein competence. In embryos carrying mutations in the iab-4 cis-regulatory region of abd A, germ cells initially associate withSGP cells, but SGP cells fail to coalesce into a gonad at PS10and germ cells scatter throughout the embryo at early stages ofretraction (Warrior, 1994; Boyle and DiNardo, 1995). Theextent of germ cell scattering in iab-4 mutants is more severethan that observed in cli mutants, suggesting that cli may actdownstream of late abd A function in gonadogenesis. Giventhat ectopic expression of cli alone has no affect on gonadformation, it is unlikely to be the sole target of late abd Afunction, suggesting that other genes required for the differen-tiation and function of SGP cells remain to be identified.
We are grateful to Manfred Frasch and Mark Peifer for sharingmaterials prior to publication. In addition, we thank members of theLehmann laboratory for helpful discussions and reagents, GinesMorata for the Hs-abdA stock, Alan Michelson for sharing unpub-lished results and Lee Cohen-Gould of the Cornell MicroscopyFacility for teaching us sectioning. Members of the DiNardo labora-tory and Alex Langston provided helpful comments on the manu-script. Bloomington, Bowling Green and Tübingen sent us fly stocks.In addition, we thank Jen LaBella for help with the screen and KerryMonks for additional technical assistance. Finally, we thank ananonymous reviewer for comments that helped improve the manu-script. This work was supported by NSF grant IBN9418271.
REFERENCES
Aboïm, A. N. (1945). Dévelopement embryonnaire et postembryonnaire desgonades normales et agamétiques de Drosophila melanogaster. Revue Suissede Zoologie 52, 53-154.
Azpiazu, N. and Frasch, M. (1993). tinman and bagpipe: two homeo box
982 M. Boyle, N. Bonini and S. DiNardo
genes that determine cell fates in the dorsal mesoderm of Drosophila. GenesDev. 7, 1325-40.
Azpiazu, N., Lawrence, P. A., Vincent, J. P. and Frasch, M. (1996).Segmentation and specification of the Drosophila Mesoderm. Genes Dev. 10,3183-3194.
Baylies, M. K., Martinez Arias, A. and Bate, M. (1995). wingless is requiredfor the formation of a subset of muscle founder cells during Drosophilaembryogenesis. Development 121, 3829-37.
Baylies, M. K. and Bate, M. (1996). twist: a myogenic switch in Drosophila .Science 272, 1481-4.
Bier, E., Vaessin, H., Shepherd, S., Lee, K., McCall, K., Barbel, S.,Ackerman, L., Carretto, R., Uemura, T., Grell, E., Jan, L. Y. and Jan, Y.N. (1989). Searching for pattern and mutation in the Drosophila genome witha P-lacZ vector. Genes Dev. 3, 1273-1287.
Bodmer, R. (1993). The gene tinman is required for specification of the heartand visceral muscles in Drosophila. Development 118, 719-29.
Bonini, N. M., Leiserson, W. M. and Benzer, S. (1993). The eyes absent gene:genetic control of cell survival and differentiation in the developingDrosophila eye. Cell 72, 379-95.
Boyle, M. and DiNardo, S. (1995). Specification, migration and assembly ofthe somatic cells of the Drosphila gonad. Develoment 121, 1815-1825.
Brook, W. J. and Cohen, S. M. (1996). Antagonistic interactions betweenWingless and Decapentaplegic responsible for dorsal-ventral pattern in theDrosophila leg. Science 273, 1373-1377.
Brookman, J. J., Toosy, A. T., Shashidhara, L. S. and White, R. A. H.(1992). The 412 retrotransposon and the development of gonadal mesodermin Drosophila. Development 116, 1185-1192.
Carmena, A., Bate, M. and Jimenez, F. (1995). Lethal of scute, a proneuralgene, participates in the specification of muscle progenitors duringDrosophila embryogenesis. Genes Dev. 9, 2373-83.
Castrillon, D. H., Gönczy, P., Alexander, S., Rawson, R., Eberhart, C. G.,Viswanathan, S., DiNardo, S. and Wasserman, S. A. (1993). Toward amolecular genetic analysis of spermatogenesis in Drosophila melanogaster:Characterization of male-sterile mutants generated by single P elementmutagenesis. Genetics 135, 489-505.
Cumberledge, S., Szabad, J. and Sakonju, S. (1992). Gonad formation anddevelopment requires the adb-A domain of the bithorax complex inDrosophila melanogaster. Development 115, 395-402.
Dougan, S. T. and DiNardo, S. (1992). wingless generates cell type diversityamong engrailed expressing cells. Nature 360, 347-350.
Ferguson, E. L. and Anderson, K. V. (1992). decapentaplegic acts as amorphogen to organize dorsal-ventral pattern in the Drosophila embryo. Cell71, 451-461.
Frasch, M. (1995). Induction of visceral and cardiac mesoderm by ectodermalDpp in the early Drosophila embryo. Nature 374, 464-7.
Greig, S. and Akam, M. (1995). The role of homeotic genes in thespecification of the Drosophila gonad. Current Biology 5, 1057-62.
Hay, B., Ackerman, L., Barbel, S., Jan, L. Y. and Jan, Y. N. (1988).Identification of a component of Drosophila polar granules. Development103, 625-640.
Huang, J. D., Schwyter, D. H., Shirokawa, J. M. and Courey, A. J. (1993).The interplay between multiple enhancer and silencer elements defines thepattern of decapentaplegic expression. Genes Dev. 7, 694-704.
Jaglarz, M. K. and Howard, K. R. (1994). Primordial germ cell migration inDrosophila melanogaster is controlled by somatic tissue. Development 120,83-89.
Jaglarz, M. K. and Howard, K. R. (1995). The active migration of Drosophilaprimordial germ cells. Development 121, 3495-503.
Jiang, J. and Struhl, G. (1996). Complementary and mutually exclusiveactivities of Decapentaplegic and Wingless organize axial patterning duringDrosophila leg development. Cell 36, 401-409.
Karpen, G. H. and Spradling, A. C. (1992). Analysis of subtelomericheterochromatin in the Drosophila minichromosome Dp1187 by single P-element insertional mutagenesis. Genetics 132, 737-753.
Kassis, J. A. (1990). Spatial and temporal control elements of the Drosophilaengrailed gene. Genes Dev. 4, 433-443.
Kellerman, K. A., Mattson, D. M. and Duncan, I. M. (1990). Mutationsaffecting the stability of the fushi tarazu protein of Drosophila. Genes Dev. 4,1936-1950.
Kiehart, D. P. and Feghali, R. (1986). Cytoplasmic myosin from Drosophilamelanogaster. J. Cell Biol. 103, 1517-25.
Lasko, P. and Ashburner, M. (1990). Posterior localization of vasa correlateswith, but is not sufficient for, pole cell development. Genes Dev. 4, 905-921.
Lawrence, P. A., Bodmer, R. and Vincent, J. P. (1995). Segmental patterningof heart precursors in Drosophila. Development 121, 4303-8.
Leptin, M. and Grunewald, B. (1990). Cell shape changes during gastrulationin Drosophila. Development 110, 73-84.
Lewis, E. B. (1978). A gene complex controlling segmentation in Drosophila.Nature 276, 565-570.
Maggert, K., Levine, M. and Frasch, M. (1995). The somatic-visceralsubdivision of the embryonic mesoderm is initiated by dorsal gradientthresholds in Drosophila. Development 121, 2107-16.
Michelson, A. M., Abmayr, S. M., Bate, M., Arias, A. M. and Maniatis, T.(1990). Expression of a MyoD family member prefigures muscle pattern inDrosophila embryos. Genes Dev. 4, 2086-2097.
Noordermeer, J., Johnston, P., Rijsewik, F., Nusse, R. and Lawrence, P. A.(1992). The consequences of ubiquitous expression of the wingless gene inthe Drosophila embryo. Development 116, 711- 719.
Nüsslein-Volhard, C., Wieschaus, E. and Kluding, H. (1984). Mutationsaffecting the pattern of the larval cuticle in Drosophila melanogaster. I.Zygotic loci on the second chromosome. Roux’s Arch. Dev. Biol. 193, 267-282.
Padgett, R. W., St. Johnston, R. D. and Gelbart, W. M. (1987). A transcriptfrom a Drosophila pattern gene predicts a protein homologous to thetransforming growth factor-β family. Nature 325, 81-84.
Park, M., Wu, X., Golden, K., Axelrod, J. D. and Bodmer, R. (1996). TheWingless Signaling Pathway is Directly Involved in Drosophila HeartDevelopment. Dev. Biol. 177, 104-116.
Paterson, B. M., Walldorf, U., Eldridge, J., Dubendorfer, A., Frasch, M.and Gehring, W. J. (1991). The Drosophila homologue of vertebratemyogenic-determination genes encodes a transiently expressed nuclearprotein marking primary myogenic cells. Proc. Natl. Acad. Sci. USA 88,3782-3786.
Peifer, M., Rauskolb, C., Williams, M., Riggleman, B. and Wieschaus, E.(1991). The segment polarity gene armadillo interacts with the winglesssignalling pathway in both embryonic and adult pattern formation.Development 111, 1029-1043.
Ranganayakulu, G., Schulz, R. A. and Olson, E. (1996). Wingless SignalingInduces nautilus Expression in the Ventral Mesoderm of the DrosophilaEmbryo. Dev. Biol. 176, 143-148.
Russell, J., Gennissen, A. and Nusse, R. (1992). Isolation and expression oftwo novel Wnt/wingless gene homologues in Drosophila. Development 115,475-485.
Schupbach, T. and Wieschaus, E. (1991). Female sterile mutations on thesecond chromosome of Drosophila melanogaster. II. mutations blockingoogenesis or altering egg morphology. Genetics 129, 1119-1136.
Simon, J., Peifer, M., Bender, W. and O’Connor, M. (1990). Regulatoryelements of the bithorax complex that control expression along the anterior-posterior axis. EMBO J. 9, 3945-3956.
Snow, P. M., Bieber, A. J. and Goodman, C. S. (1989). Fasciclin III: a novelhomophilic adhesion molecule in Drosophila. Cell 59, 313-323.
Sonnenblick, B. P. (1941). Germ cell movements and sex differentiation of thegonads in the Drosophila embryo. Proc. Natn. Acad. Sci. USA 26, 373-381.
Staehling-Hampton, K., Hoffmann, F. M., Baylies, M. K., Rushton, E. andBate, M. (1994). dpp induces mesodermal gene expression in Drosophila.Nature 372, 783-786.
Szabad, J. and Nöthiger, R. (1992). Gyandomorphs of Drosophila suggest onecommon primordium for the somatic cells of the female and male gonads inthe region of abdominal segments 4 and 5. Development 115, 527-533.
Tautz, D. and Peifle, C. (1989). A non-radioactive in situ hybridization methodfor localization of specific RNAs in Drosophila embryos reveals translationalcontrol of the segmentation gene hunchback. Chromosoma 98, 81-85.
Warrior, R. (1994). Primordial germ cell migration and the assembly of theDrosophila embryonic gonad. Dev. Biol. 166, 180-94.
Wharton, K. A., Ray, R. P. and Gelbart, W. M. (1993). An activity gradient ofdecapentaplegic is necessary for the specification of dorsal pattern elementsin the Drosophila embryo. Development 117, 807-22.
Wu, X., Golden, K. and Bodmer, R. (1995). Heart development in Drosophilarequires the segment polarity gene wingless. Dev. Biol. 169, 619-28.