Expression of Pendrin and the Pendred Syndrome (PDS)Gene in Human Thyroid Tissues*
JEAN-MICHEL BIDART, CATERINA MIAN, VLADIMIR LAZAR, DIEGO RUSSO,SEBASTIANO FILETTI, BERNARD CAILLOU, AND MARTIN SCHLUMBERGER
Departments of Clinical Biology (J.M.B., V.L.), Pathology (B.C.), and Nuclear Medicine (C.M., M.S.),Institut Gustave-Roussy, 94805 Villejuif, France; and Dipartimento di Medicina Sperimentale eClinica, Policlinico Mater Domini (D.R., S.F.), 88100 Catanzaro, Italy
ABSTRACTThe gene recently cloned that is responsible for the Pendred syn-
drome (PDS), an autosomal recessive disease characterized by goiterand congenital sensorineural deafness, is mainly expressed in thethyroid gland. Its product, designated pendrin, was shown to trans-port chloride and iodide. To investigate whether the PDS gene isaltered during thyroid tumorigenesis, PDS gene expression and pen-drin expression were studied using real-time kinetic quantitativePCR and antipeptide antibodies, respectively, in normal, benign, andmalignant human thyroid tissues. The results were then compared tothose observed for sodium/iodide symporter (NIS) expression.
In normal tissue, pendrin is localized at the apical pole of thyro-cytes, and this in contrast to the basolateral location of NIS. Immu-nostaining for pendrin was heterogeneous both inside and among
follicles. In hyperfunctioning adenomas, the PDS messenger ribonu-cleic acid level was in the normal range, although immunohistochem-ical analysis showed strong staining in the majority of follicular cells.In hypofunctioning adenomas, mean PDS gene expression was sim-ilar to that detected in normal thyroid tissues, but pendrin immu-nostaining was highly variable. In thyroid carcinomas, PDS geneexpression was dramatically decreased, and pendrin immunostainingwas low and was positive only in rare tumor cells. This expressionprofile was similar to that observed for the NIS gene and its proteinproduct. In conclusion, our study demonstrates that pendrin is locatedat the apical membrane of thyrocytes and that PDS gene expressionis decreased in thyroid carcinomas. (J Clin Endocrinol Metab 85:2028–2033, 2000)
THE GENE responsible for Pendred syndrome (PDS), anautosomal recessive disease characterized by goiter
and congenital sensorineural deafness, has recently beenidentified (1). The PDS gene is located on chromosome 7q31and encodes a 5-kb transcript that is mainly expressed in thethyroid gland (2, 3). The PDS gene product, pendrin, is ahighly hydrophobic transmembrane protein composed of780 amino acids. On the basis of sequence homology, it wasfirst suggested that pendrin might belong to the sulfate aniontransporter family, composed of the human diastrophic dys-plasia sulfate transporter (4) and the human sulfate trans-porter down-regulated in adenoma (5). More recently, pen-drin was shown to transport chloride and iodide, but notsulfate, thereby emphasizing a potential critical role for thisprotein in thyroid physiology (6, 7). This is in agreement withprevious in vitro and in vivo findings showing that iodineorganification was altered in thyroid cells obtained from apatient with PDS (8).
The iodine pathway comprises several steps in thyroidcells that involve an iodide transport mechanism via thesodium/iodide symporter (NIS) (9), iodide organificationinto the thyroglobulin (Tg) molecule at the apical pole that
is mediated by the thyroid peroxidase (TPO), and thyroidhormone synthesis and secretion (10, 11). Biochemical ab-normalities that affect the complex process of thyroid hor-mone synthesis have been demonstrated in differentiatedthyroid cancers (DTC) (12, 13). In vivo, DTC usually appearas hypofunctioning cold nodules on the thyroid scintiscan,indicating impaired radioiodine trapping ability, but theyretain the capacity to synthesize Tg and to respond to TSHstimulation (14). In vitro studies of tumor cells revealed a lowconcentration of intracellular iodine, reduced Tg iodination,decreased TPO activity, and a low rate of thyroid hormonesynthesis (15).
A defective iodide-trapping mechanism appears to be anearly and constant feature during oncogenic transformationof thyroid cells (16). Recent reports have shown that thisdefect is due to reduced NIS gene expression detected in bothmalignant and benign hypofunctioning thyroid tumors (17–21). Other changes in the expression of thyroid-specific genes(TPO, Tg, and TSH receptor) are associated with neoplastictransformation, but are present in only some thyroid tumorphenotypes (21, 22). In this respect, the demonstration thatpendrin serves as an iodide transporter prompted us to in-vestigate whether PDS gene expression may be altered dur-ing thyroid tumorigenesis.
We analyzed PDS gene expression using real-time kineticquantitative PCR (23) and pendrin expression using an im-munohistochemical method based on antipeptide antibodiesin a series of normal, benign, and malignant human thyroidtissues, in which we recently studied Tg, TPO, TSH receptor,and NIS gene expression (21).The results of PDS gene ex-
Received July 19, 1999. Revision received November 30, 1999. Ac-cepted December 20, 1999.
Address all correspondence and requests for reprints to: Prof. J. M.Bidart, Institut Gustave-Roussy, 39 rue Camille Desmoulins, 94805 Ville-juif, France. E-mail: [email protected].
* This work was supported by grants from the Comite du Val deMarne de la Ligue Nationale Contre le Cancer, the Electricite de France,the FEGEFLUC, LIPHA Sante, and the Associazione Italiana per laRicerca sul Cancro.
pression were then compared to those observed for NIS geneexpression, which have been previously reported (21).
Materials and MethodsTissue samples
Thyroid tissue samples were selected after histological examinationand classified according to WHO recommendations (24). Tissue samplesselected for real-time quantitative PCR were as follows (21): hyperfunc-tioning tissues, including Graves’ thyroid tissues (n 5 5) and toxicadenomas (n 5 8), benign hypofunctioning thyroid follicular adenomas(n 5 24), and differentiated thyroid carcinomas (n 5 43), includingpapillary carcinomas (n 5 38) and poorly differentiated and widelyinvasive follicular carcinomas (n 5 5). To obtain calibrator materials forreal-time PCR analysis, normal tissue counterparts (n 5 5) were carefullymicrodissected from thyroid glands obtained from five patients with
papillary carcinoma. For immunohistochemical analysis, normal thy-roid samples (n 5 4) and pathological thyroid tissue specimens wereselected from the above series, including Graves’ thyroid tissues (n 5 3),benign hypofunctioning adenomas (n 5 5), and papillary carcinomas(n 5 8). Specimens were frozen at 280 C in isopentane and stored inliquid nitrogen before use. Except for the patients with Graves’ diseaseor toxic adenoma, in whom serum TSH was undetectable, thyroid sam-ples were obtained from euthyroid subjects whose serum TSH concen-trations were in the normal range at the time of surgery. This study wasperformed in accordance with protocols previously approved by thelocal human studies committee.
Peptide synthesis, production, and characterization ofantipeptide antiserum to pendrin
A composite peptide spanning the N-terminal (1–16) and the C-terminal(765–780) regions of pendrin was synthesized by a conventional solid phasemethod using a PE Applied Biosystems model 431A peptide synthesizer(Foster City, CA). The identity and purity of the 1–16/765–780 peptide wereverified as previously described (18). The synthetic peptide was conjugatedto keyhole limpet hemocyanin using benzidine as the coupling agent on the765Tyr residue. Two rabbits were immunized by intradermal injections ofthe synthetic peptide-carrier conjugate. After two boosts at 3-week inter-vals, animals were bled, and their serum specimens were tested in anenzyme-linked immunosorbent assay. Antisera at various dilutions wereverified for their capacity to react with the pendrin synthetic peptide coatedon microtiter plates. Antibody binding was then revealed by peroxidase-labeled goat antirabbit antibody (Nordic, Tilburg, The Netherlands). Theproduction and characteristics of the antihuman NIS antiserum have beendescribed previously (18).
Immunohistochemistry
Serial frozen cryostat tissue sections (5 mm) were cut and fixed in acetonefor 10 min. These sections were then incubated for 30 min with either theantipendrin or anti-NIS antiserum, diluted at 1:75 and 1:500, respectively.Sections were then washed three times in Tris-HCl buffer for 5 min eachtime and incubated with a biotinylated antibody (K4017, EnVision LabeledPolymer, DAKO Corp., Carpinteria, CA). They were washed again threetimes and incubated with alkaline phosphatase-labeled streptavidin(DAKO-K674) for 10 min. After three additional washes, staining wascompleted after incubation with a substrate chromogen solution (DAKO-K699 Fast red). Negative controls were obtained by studying nonthyroidtissues and incubating thyroid sections with preimmune antiserum andimmune antiserum preabsorbed with excess corresponding peptide.
Determination of messenger RNA (mRNA) level using real-time PCR
Total RNA was isolated from tissue samples using the DNA/RNAextraction Midi kit according to the manufacturer’s instructions (QIA-GEN, Hilden, Germany). The quality of RNA was assessed by conven-tional gel electrophoresis. One microgram of total RNA from each sam-ple was reverse transcribed in a 20-mL volume reaction using 50 UMoloney murine leukemia virus reverse transcriptase, 20 U ribonucleaseinhibitor (Perkin-Elmer Corp./PE Applied Biosystems, Foster City, CA),1 mmol/L dA/T/C/G (Amersham-Pharmacia Biotech, Uppsala, Swe-den), 5 mmol/L MgCl2, 10 mmol/L Tris-HCl (pH 8.3), 10 mmol/L KCl,and 50 pmol/L of random hexamers (Perkin-Elmer Corp./PE AppliedBiosystems). The complementary DNAs were then diluted 1:20 in nu-clease-free H2O (Promega Corp., Madison, WI).
Oligonucleotide primers and TaqMan (PE Applied Biosystems)probes were designed to be intron spanning, using the computer pro-gram Primer Express (Perkin-Elmer Corp./PE Applied Biosystems).They were purchased from Perkin-Elmer Corp./PE Applied Biosystems,
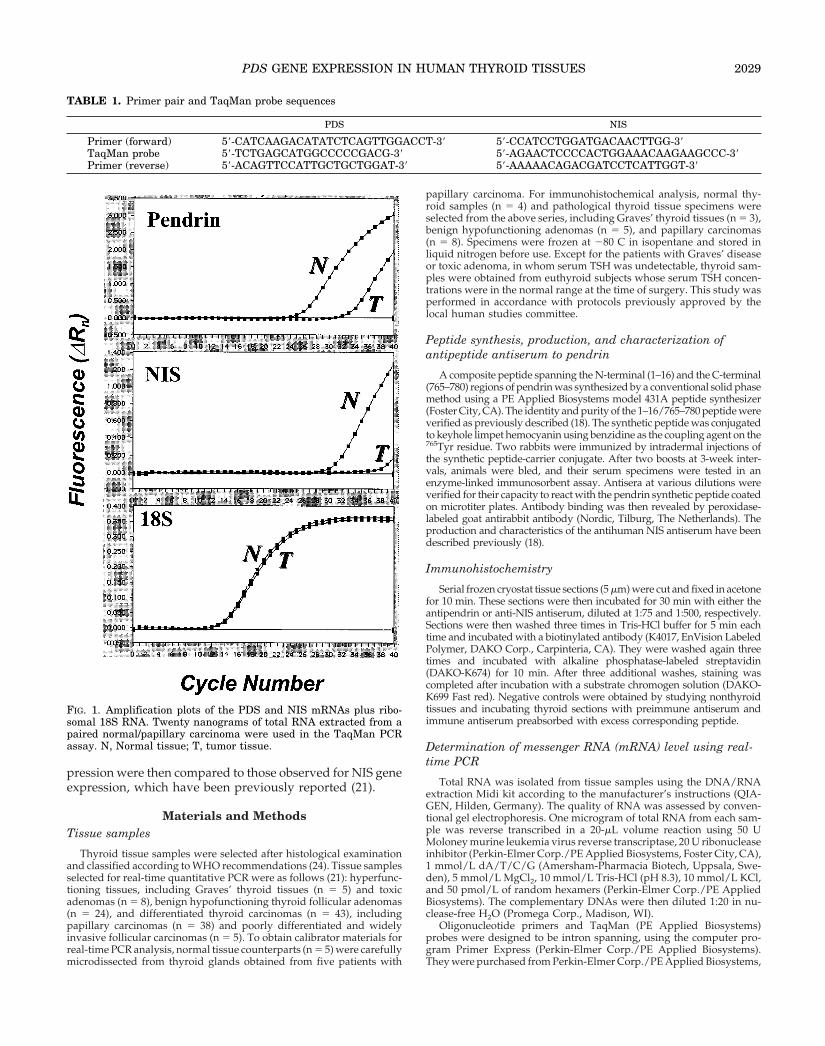
FIG. 1. Amplification plots of the PDS and NIS mRNAs plus ribo-somal 18S RNA. Twenty nanograms of total RNA extracted from apaired normal/papillary carcinoma were used in the TaqMan PCRassay. N, Normal tissue; T, tumor tissue.
TABLE 1. Primer pair and TaqMan probe sequences
PDS NIS
Primer (forward) 59-CATCAAGACATATCTCAGTTGGACCT-39 59-CCATCCTGGATGACAACTTGG-39TaqMan probe 59-TCTGAGCATGGCCCCCGACG-39 59-AGAACTCCCCACTGGAAACAAGAAGCCC-39Primer (reverse) 59-ACAGTTCCATTGCTGCTGGAT-39 59-AAAAACAGACGATCCTCATTGGT-39
PDS GENE EXPRESSION IN HUMAN THYROID TISSUES 2029
and their sequences are presented in Table 1. PCR reaction was carriedout to produce amplicons that were subsequently analyzed by gel elec-trophoresis and sequencing. Real-time quantitative PCR was achievedin 96 sample tubes/assay using a complementary DNA equivalent of 20ng/total RNAz50 mL/tube with the TaqMan PCR core reagent kit ac-cording to the manufacturer’s instructions: 1 3 buffer A, 5 mmol/LMgCl2, 200 mmol/L dA/C/G, 400 mmol/L dU, 1.25 U AmpliTaq GoldDNA polymerase, 2.5 U uracyl N-glycosylase, 100 mmol/L TaqManprobe, and 200 mmol/L of each primer. PCR was developed on the ABIPrism 7700 Sequence Detector (Perkin-Elmer Corp./PE Applied Bio-systems). To normalize for differences in the amount of total RNA addedto the reaction, amplification of 18S ribosomal RNA was performed asan endogenous control. Primers and probes for 18S RNA were pur-chased from Perkin-Elmer Corp./PE Applied Biosystems. The mRNAcontent of each target gene was simultaneously determined in the 85samples, including the 5 paired normal/tumor samples, in a 1-run assay.
Statistical methods
The expression of PDS and NIS genes detected in each histologicaltype was compared with that in normal samples. mRNA expressionpatterns were compared using the t test, and the correlation betweenthese expression profiles was studied using Spearman’s rank ordercorrelation coefficient for each histological type. The level of significancechosen was fixed at 5%.
ResultsProduction and characterization of antipeptideantiserum to pendrin
Rabbit antiserum titers obtained after immunization with thecomposite pendrin (1–16/765–780) synthetic peptide-carrierconjugate were determined by enzyme-linked immunosorbentassay. Both animals produced antisera, and competitive inhi-bition experiments (data not shown) indicated that the affinityof these antisera for the synthetic peptide ranged from 1026 to5 3 1027 mol/L. The characteristics of the antiserum directedagainst human NIS have been described previously (18).
Determination of the mRNA level
To validate the real-time PCR method, standard curves forthe PDS gene and 18S ribosomal RNA were constructed fromPCR products that were serially diluted in nuclease-free H2O.Figure 1 shows the amplification plots for the PDS and NISmRNAs and control 18S RNA. The efficiency of the standardcurve, as determined by its slope, allowed us to quantify thePDS gene expression profile in each thyroid specimen by usingthe comparative threshold cycle (Ct) method according to themanufacturer’s instructions. The characteristics of the assay forNIS gene expression have been reported previously (21). Thecalibrator was constituted from one sample of normal tissue: itwas used as the 1 3 sample, and all other levels were expressedas an n-fold difference relative to the calibrator. The intraassaycoefficient of variation was less than 1%.
PDS gene and pendrin expression in normal thyroid tissues
Among the five specimens of normal thyroid tissue, meanand median PDS gene expression levels were similar (Fig. 2and Table 2). Mean and median NIS gene expression levelswere also similar in normal thyroid specimens (21).
PDS and NIS gene products displayed a different loca-tion pattern, as shown by immunohistochemical analysis;
FIG. 2. Box plots for PDS and NIS markers according to the histological diagnosis (0, normal tissues; 1, hyperfunctioning tissues; 2, hypo-functioning adenomas; 3, carcinomas). The gray box shows the limits of the middle half of the data (the bold line inside the box represents themedian). Whiskers are drawn to the nearest value not exceeding a standard span (1.5 3 interquartile range) from the quartiles. Extreme points(outliers) are highlighted by a cross.
TABLE 2. Distribution of mRNA values according to thehistological group (mean (SD), median [min–max])
Thyroid tissues n PDS NIS
Normal 5 1.1 (0.5) 1.6 (0.9)1.3 [0.3–1.5] 1.1 [1.0–3.0]
Results are expressed as an x-fold difference relative to the cali-brator. n, Number of samples. Hyperfunctioning thyroid tissues in-clude five samples from patients with Graves’ disease and eight toxicadenomas.
a ,0.1.b ,0.01.
2030 BIDART ET AL. JCE & M • 2000Vol 85 • No 5
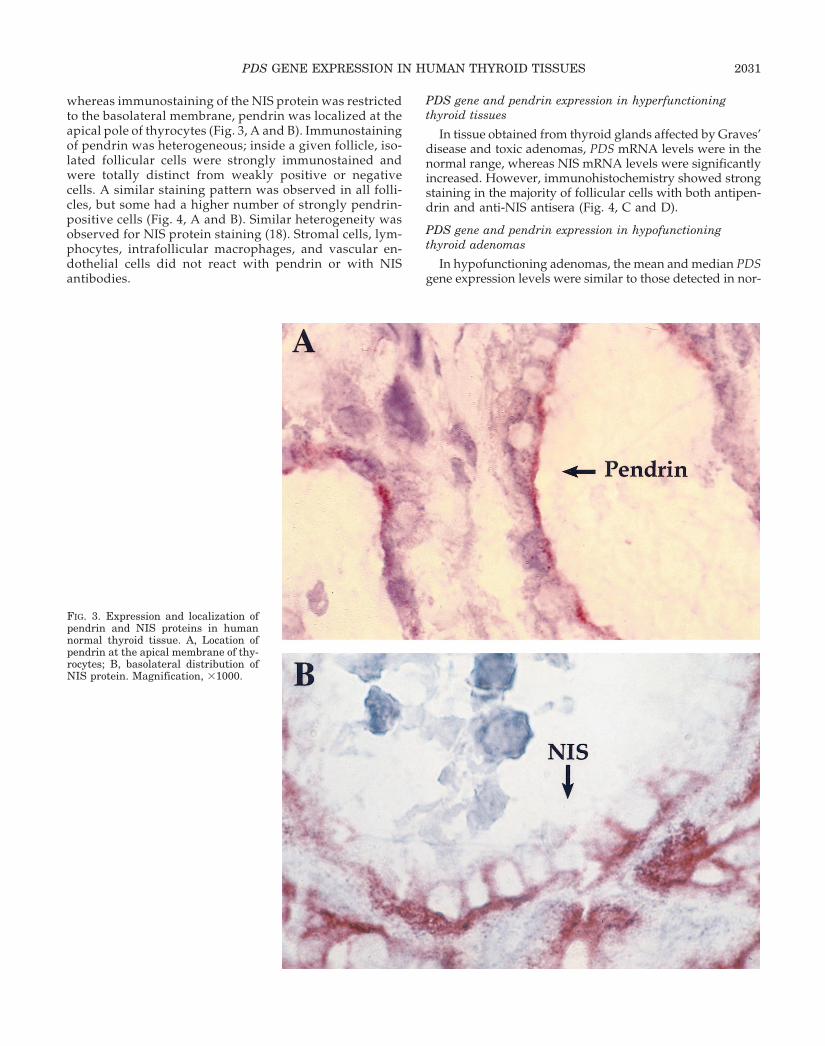
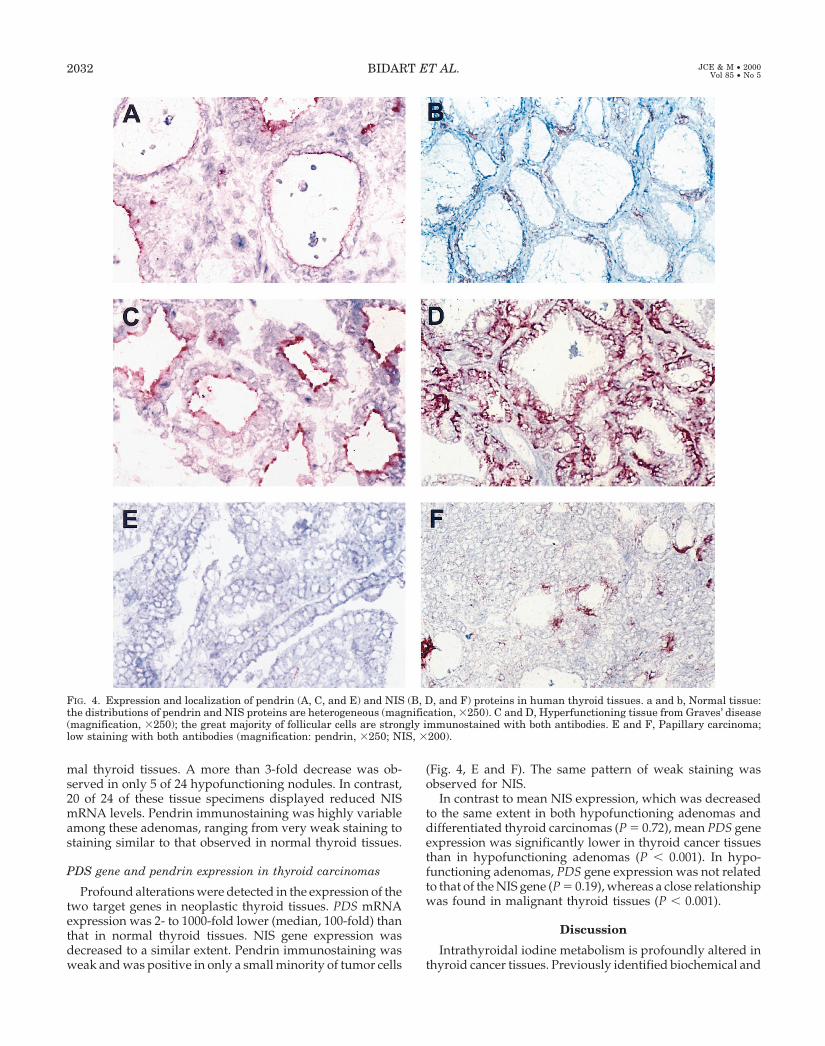
whereas immunostaining of the NIS protein was restrictedto the basolateral membrane, pendrin was localized at theapical pole of thyrocytes (Fig. 3, A and B). Immunostainingof pendrin was heterogeneous; inside a given follicle, iso-lated follicular cells were strongly immunostained andwere totally distinct from weakly positive or negativecells. A similar staining pattern was observed in all folli-cles, but some had a higher number of strongly pendrin-positive cells (Fig. 4, A and B). Similar heterogeneity wasobserved for NIS protein staining (18). Stromal cells, lym-phocytes, intrafollicular macrophages, and vascular en-dothelial cells did not react with pendrin or with NISantibodies.
PDS gene and pendrin expression in hyperfunctioningthyroid tissues
In tissue obtained from thyroid glands affected by Graves’disease and toxic adenomas, PDS mRNA levels were in thenormal range, whereas NIS mRNA levels were significantlyincreased. However, immunohistochemistry showed strongstaining in the majority of follicular cells with both antipen-drin and anti-NIS antisera (Fig. 4, C and D).
PDS gene and pendrin expression in hypofunctioningthyroid adenomas
In hypofunctioning adenomas, the mean and median PDSgene expression levels were similar to those detected in nor-
FIG. 3. Expression and localization ofpendrin and NIS proteins in humannormal thyroid tissue. A, Location ofpendrin at the apical membrane of thy-rocytes; B, basolateral distribution ofNIS protein. Magnification, 31000.
PDS GENE EXPRESSION IN HUMAN THYROID TISSUES 2031
mal thyroid tissues. A more than 3-fold decrease was ob-served in only 5 of 24 hypofunctioning nodules. In contrast,20 of 24 of these tissue specimens displayed reduced NISmRNA levels. Pendrin immunostaining was highly variableamong these adenomas, ranging from very weak staining tostaining similar to that observed in normal thyroid tissues.
PDS gene and pendrin expression in thyroid carcinomas
Profound alterations were detected in the expression of thetwo target genes in neoplastic thyroid tissues. PDS mRNAexpression was 2- to 1000-fold lower (median, 100-fold) thanthat in normal thyroid tissues. NIS gene expression wasdecreased to a similar extent. Pendrin immunostaining wasweak and was positive in only a small minority of tumor cells
(Fig. 4, E and F). The same pattern of weak staining wasobserved for NIS.
In contrast to mean NIS expression, which was decreasedto the same extent in both hypofunctioning adenomas anddifferentiated thyroid carcinomas (P 5 0.72), mean PDS geneexpression was significantly lower in thyroid cancer tissuesthan in hypofunctioning adenomas (P , 0.001). In hypo-functioning adenomas, PDS gene expression was not relatedto that of the NIS gene (P 5 0.19), whereas a close relationshipwas found in malignant thyroid tissues (P , 0.001).
Discussion
Intrathyroidal iodine metabolism is profoundly altered inthyroid cancer tissues. Previously identified biochemical and
FIG. 4. Expression and localization of pendrin (A, C, and E) and NIS (B, D, and F) proteins in human thyroid tissues. a and b, Normal tissue:the distributions of pendrin and NIS proteins are heterogeneous (magnification, 3250). C and D, Hyperfunctioning tissue from Graves’ disease(magnification, 3250); the great majority of follicular cells are strongly immunostained with both antibodies. E and F, Papillary carcinoma;low staining with both antibodies (magnification: pendrin, 3250; NIS, 3200).
2032 BIDART ET AL. JCE & M • 2000Vol 85 • No 5
molecular defects hamper radioiodine concentration in mostneoplastic thyroid tumors and account for its absence inapproximately a third of thyroid cancer metastases (25). Ourstudy demonstrates that the pendrin-PDS gene pathway isalso impaired in thyroid cancer tissues, and this is consistentwith the defective iodine organification found in these tis-sues (26).
Pendrin, the predicted protein, was demonstrated to ac-celerate the transport of chloride and iodide (6). Defectiveiodine organification due to an inactivating mutation of thePDS gene is observed in patients with PDS. This results in agoiter, with various degrees of hypothyroidism; an alterationin iodine organification is attested to by a positive perchlor-ate discharge test and the presence of Tg molecules display-ing a normal monomer size but a low hormone and iodinecontent (27).
More interestingly, our immunohistochemical study,based on an antipeptide antibody, demonstrated that pen-drin is located at the apical pole of thyrocytes. Together withthe above-mentioned data, this observation suggests thatpendrin is the apical iodide transporter that permits theorganification of Tg.
In Graves’ thyroid tissues and in toxic adenomas, PDSgene expression was not significantly increased, in contrastto that of NIS and TPO (21). This suggests that, like Tg mRNAexpression, PDS gene expression is less sensitive to the stim-ulatory effects of the TSH pathway. However, immunohis-tochemistry showed stronger staining in these tissue speci-mens than in normal tissues, indicating that pendrinexpression is regulated at the transcription level, and that thisis increased in these tissues. This finding is in accordancewith the increased rate of thyroid hormone synthesis.
In contrast to NIS mRNA and protein levels, which weredecreased in both benign and malignant hypofunctioningthyroid tumors, PDS gene and pendrin expression levelsappeared to be normal in most hypofunctioning adenomasand were significantly decreased only in differentiated thy-roid carcinomas. A highly significant correlation was foundbetween pendrin and NIS gene expression in differentiatedthyroid carcinomas. Although the mechanism of this alter-ation remains to be investigated, these data suggest that theexpression of these two genes is regulated by different path-ways. These data also suggest that the limited degree ofiodination of Tg found in differentiated thyroid carcinoma isrelated to defects not only in the iodine-trapping ability andin the iodination process through low TPO biochemical ac-tivity, but also to low expression of apical pendrin.
In conclusion, our study demonstrates that pendrin is lo-cated at the apical membrane of thyrocytes and suggests thatit is probably the apical iodide transporter. PDS gene ex-pression is dramatically decreased in DTC, but not in hy-pofunctioning adenomas. It remains to be establishedwhether and to what extent PDS gene expression constitutesa critical alteration in thyroid tumors.
Acknowledgments
We acknowledge Jean-Pierre Levillain for expert technical assistancewith peptide synthesis, and Lorna Saint-Ange for editing themanuscript.
References
1. Billerbeck AE, Cavaliere H, Goldberg AC, Kalil J, Medeiros-Neto G. 1994Clinical and molecular genetics studies in Pendred’s syndrome. Thyroid.4:279–284.
2. Everett LA, Glaser B, Beck JC, et al. 1997 Pendred syndrome is caused bymutations in a putative sulphate transporter gene (PDS). Nat Genet.17:411–422.
3. Coyle B, Reardon W, Herbrick JA, et al. 1998 Molecular analysis of the PDSgene in Pendred syndrome. Hum Mol Genet. 7:1105–1112.
4. Hastbacka J, de la Chapelle A, Mahtani MM, et al. 1994 The diastrophicdysplasia gene encodes a novel sulfate transporter: positional cloning byfine-structure linkage disequilibrium mapping. Cell. 78:1073–1087.
5. Schweinfest CW, Henderson KW, Suster S, Kondoh N, Papas TS. 1993Identification of a colon mucosa gene that is down-regulated in colon adeno-mas and adenocarcinomas. Proc Natl Acad Sci. 90:4166–4170.
6. Scott DA, Wang R, Kreman TM, Sheffield VC, Karnishki LP. 1999 ThePendred syndrome gene encodes a chloride-iodide transport protein. NatGenet. 21:440–443.
7. Kraiem Z, Heinrich R, Sadeh O, Shiloni E, Nassir E, Hazani E, Glaser B. 1999Sulfate transport is not impaired in Pendred syndrome thyrocytes. J ClinEndocrinol Metab. 84:2574–2576.
8. Sheffield VC, Kraiem Z, Beck JC, et al. 1996 Pendred syndrome maps tochromosome 7q21–34 and is caused by an intrinsic defect in thyroid iodineorganification. Nat Genet. 12:424–426.
9. Smanik PA, Liu Q, Furminger L, Ryu K, Xing S, Mazzaferri EL, Jhiang SM.1996 Cloning of the human sodium iodide symporter. Biochem Biophys ResCommun. 226:339–345.
10. Carrasco N. 1993 Iodide transport in the thyroid gland. Biochim Biophys Acta.1154:65–82.
11. Cavalieri RR. 1997 Iodine metabolism and thyroid physiology: current con-cepts. Thyroid. 7:177–181.
12. Brabant G, Maenhaut C, Kohrle J, et al. 1991 Human thyrotropin receptorgene: expression in thyroid tumors and correlation to markers of thyroiddifferentiation and dedifferentiation. Mol Cell Endocrinol. 82:7–12.
13. Hoang-Vu C, Dralle H, Scheumann G, Maenhaut C, Horn R, von zur MuhlenA, Brabant G. 1992 Gene expression of differentiation- and dedifferentiationmarkers in normal and malignant human thyroid tissues. Exp Clin Endocrinol.100:51–56.
14. Schlumberger M, Charbord P, Fragu P, Lumbroso J, Parmentier C, TubianaM. 1980 Circulating thyroglobulin and thyroid hormones in patients withmetastases of differentiated thyroid carcinoma: relationship to serum thyro-tropin levels. J Clin Endocrinol Metab. 51:513–519.
15. Thomas-Morvan C, Nataf B, Tubiana M. 1974 Thyroid proteins and hormonesynthesis in human thyroid cancer. Acta Endocrinol (Copenh). 76:651–669.
16. Trapasso F, Iuliano R, Chiefari E, et al. 1999 Iodide symporter gene expressionin normal and transformed rat thyroid cells. Eur J Endocrinol. 140:447–451.
17. Jhiang SM, Cho JY, Ryu KY, et al. 1998 An immunohistochemical study ofNa1/I2 symporter in human thyroid tissues and salivary gland tissues. En-docrinology. 139:4416–4419.
18. Caillou B, Troalen F, Baudin E, Talbot M, Filetti S, Schlumberger M, BidartJM. 1998 Na1/I2 symporter in human thyroid tissues: an immunohistochem-ical study. J Clin Endocrinol Metab. 83:4102–4106.
19. Arturi F, Russo D, Schlumberger M, et al. 1998 Iodide symporter gene ex-pression in human thyroid tumors. J Clin Endocrinol Metab. 83:2493–2496.
20. Ryu KY, Senokozlieff ME, Smanik PA, et al. 1999 Development of reversetranscription-competitive polymerase chain reaction method to quantitate theexpression levels of human sodium iodide symporter. Thyroid. 4:405–409.
21. Lazar V, Bidart JM, Caillou B, Mahe C, Lacroix L, Filetti S, Schlumberger M.1999 Expression of the Na1/I2 symporter gene in human thyroid tumors: acomparison study with other thyroid-specific genes. J Clin Endocrinol Metab.84:3228–3234.
22. Natali PG, Berlingieri MT, Nicotra MR, Fusco A, Santoro E, Bigotti A,Vecchio G. 1995 Transformation of thyroid epithelium is associated with lossof c-Kit receptor. Cancer Res. 55:1787–1791.
23. Gibson UE, Heid CA, Williams P. 1996 A novel method for real time quan-titative RT-PCR. Genome Res. 6:995–1001.
24. Hedinger C, Williams ED, Sobin LH. 1988 Histological typing of thyroidtumours, 2nd Ed, vol 11. Berlin: Springer-Verlag.
25. Schlumberger M, Challeton C, De Vathaire F, et al. 1996 Radioactive iodinetreatment and external radiotherapy for lung and bone metastases from thy-roid carcinoma. J Nucl Med. 37:598–605.
26. Cooper DS, Axelrod L, DeGroot LJ, Vickery Jr AL, Maloof F. 1981 Congenitalgoiter and the development of metastatic follicular carcinoma with evidencefor a leak of nonhormonal iodide: clinical, pathological, kinetic, and biochem-ical studies and a review of the literature. J Clin Endocrinol Metab. 52:294–306.
27. Mason ME, Dunn AD, Wortsman J, et al. 1995 Thyroids from siblings withPendred’s syndrome contain thyroglobulin messenger ribonucleic acid vari-ants. J Clin Endocrinol Metab. 80:497–503.








![€¦ · GERMAN J. BIDART CAMPOS, MANUAL DE DERECHO CONSTITUCIONAL ARGENTINO [MANUAL OF ARGENTINE CONSTITUTIONAL LAW] 455—60 (1981); Judgment of Oct. 22, 1962 (re Ruggero), Corte](https://static.documents.pub/doc/80x56/5eb2079d140def52c56bd83d/german-j-bidart-campos-manual-de-derecho-constitucional-argentino-manual-of-argentine.jpg)
![arXiv:1909.03402v4 [cs.CV] 1 Apr 2020 · 2020-04-02 · Squeeze-and-Attention Networks for Semantic Segmentation Zilong Zhong1;4, Zhong Qiu Lin 2, Rene Bidart , Xiaodan Hu 2, Ibrahim](https://static.documents.pub/doc/80x56/5f77c476deecde5f0019f49f/arxiv190903402v4-cscv-1-apr-2020-2020-04-02-squeeze-and-attention-networks.jpg)










