Page 1
Expression of rabies virus coat protein in plants
and its efficacy as anti-rabies antigens
THESIS
SUBMITTED TO THE
UNIVERSITY OF LUCKNOW
FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
IN
BOTANY
BY
ANKIT SINGH M.Sc. BIOCHEMISTRY
PLANT MOLECULAR BIOLOGY & GENETIC ENGINEERING DIVISION
NATIONAL BOTANICAL RESEARCH INSTITUTE
LUCKNOW (INDIA)
&
DEPARTMENT OF BOTANY
FACULTY OF SCIENCE
UNIVERSITY OF LUCKNOW, LUCKNOW (INDIA)
(2013)
Page 2
Dedicated to my Nation, Parents and all those bright sources of knowledge
who enlighten my life….
Page 4
Contents
Acknowledgements i-ii
Abbreviation iii-v List of figures vi-xi List if tables xii
Chapter 1 Introduction 1-6
Chapter 2 Review of literature 7-38
Chapter 3 Materials and methods 39-50
Chapter 4 Observations 51-58
Chapter 5 Discussion 59-66
Chapter 6 Summary and conclusions 67-71
Bibliography 72-118
List of publications 119
Page 5
ACKNOWLEDGEMENTS
i
ACKNOWLEDGEMENTS
I thank Almighty for blessing me with strong will power, patience and confidence, which
helped me in completing the present work. I also very gratefully acknowledge my colleagues for helping
me in various ways to accomplish my thesis work. Without their support and inspiration my endeavors
would not have come to fruition.
First, I thank Dr. Rakesh Tuli, ExDirector, National Botanical Research Institute, for giving
me an opportunity to join his lab and also for excellent guidance, critical suggestions, long scientific
discussions and constant encouragement. His devotion to the work and untiring efforts has left deep
impression in my heart.
I am equally grateful to Dr. Gauri Saxena, Department of Botany, University of Lucknow for
accepting me as a Ph.D. student, technical discussions, critical reading of the thesis and
encouragement.
With genuine perception of moral obligation, I acknowledge my reverence and gratitude to
Dr. P. K. Singh for his constant guidance in planning and completion of this work, useful advices,
tender affection and encouragement which will be always memorable to me.
I acknowledge C.S.I.R with thanks for providing the financial support.
I gratefully acknowledge Dr. Praveen C. Verma for teaching me about hairy root system and
related molecular biology techniques.
I gratefully acknowledge the help and advice given by my seniors Dr. Sribash Roy, Dr. Dinesh
Yadav, Mr. Siddharth Tiwari, Dr. Raju Madanala, Dr. Chandrashekhar and Dr. Hemant Yadav. Very
special thanks to Mr. Rajesh Srivastava for his cooperation.
Page 7
Abbreviations
iii
Abbreviations
A - Adenine
APC - Antigen presenting cell
APS - Ammonium persulphate
ARE - At rich element
BAP - 6- Benzylaminopurine
ME - -mercaptoethanol
BCIP - 5-Bromo-4-chloro-3-indolyl phosphate
Bisacrylamide - N,N’-methylene bisacrylamide
bp/Kb - Base pair/kilo base pair
BSA - Bovine serum albumin
C - Cytosine
CaMV - Cauliflower mosaic virus
cal-s - Calreticulin signal sequence
CIAP - Calf intestinal alkaline phosphatase
cm - Centimeter
CNS - Central nervous system
CPM - Count per minute
cpm - counts per minute
CTAB - Hexadecyl trimethyl-ammonium
bromide
CTB - Cholera toxin B chain protein
ctxB - Cholera toxin B chain gene
CTL - Cytotoxic T Lymhocytes
CTP - Cytosine triphosphate
CVS - Challenge Virus standard
DEPC - Diethyl pyrocarbonate
DFA - Direct fluorescent antibody
Deica - Diethyl ammonium dithiocarbamate
DMTr - Dimethyl trityl
DNA - Deoxyribonucleic acid
DNase-I - Deoxyribonuclease-I
dNTP - Deoxy nucleoside triphosphate
DSE - Downstream element
DTT - Dithiothreitol
EDTA - Ethylene diamine tetra acetic acid
ER - Endoplasmic reticulum
ERA - Evelyn – Rokitnicki – Abelseth
ERIG - Equine rabies immunoglobulin
FAM - 6-carboxy fluorescein
FUE - Far upstream element
G - Guanine
g - Gram
GLP - Good laboratory practices
GMP - Good manufacturing practices
G protein - Glycoprotein
Page 8
Abbreviations
iv
(gp)2
-
Glycine-proline hinge
GTE - Glucose- Tris – EDTA
G-Vacc - Recombinant vaccinea virus
expressing rabies glycoprotein
h - Hour
HEP - High Egg Passage
HEPES - N-2-hydroxy ethyl piperazine-N’-
2ethane sulfonic acid
hptII - Hygromycin phosphotransferase II
IPTG - Isopropyl -D thiogalactopyranoside
IU - International Unit
kDa - Kilo Dalton
L - Rabies polymerase
LA - Luria agar
LB - Luria broth
LMP - Low melting point
M - Matrix protein
MES - (2- [n-Morpholino] ethanesulfonic
acid)
Mg - Milligram
MHC - Major histocompatibity complex
Min - Minute
µg - Microgram
µl - Microliter
ml - Milliliter
MOPS - N-Morpholinopropanesulfonic acid
MS - Murashige and Skoog
N - Nucleoprotein
NAA - -Naphthalene acetic acid
N-Bac - Recombinant baculovirus expressing
rabies nucleoprotein
NBT - Nitro blue tetrazolium
NFQ - Non-fluorescent quencher dye
Ng - Nanogram
NK - Natural killer cells
nptII - Neomycin phosphotransferase II
NS - Non-structural protein
NUE - Near- upstream element
N-Vacc - Recombinant pox virus expressing
rabies nucleoprotein
O.D. - Optical density
P - Phosphoprotein
PAGE - Polyacrylamide gel electrophoresis
PBS - Phosphate buffer saline
PCEC - Purified chicken embryo cell vaccine
PCR - Polymerase chain reaction
PDEV - Purified duck embryo cell vaccine
Page 9
Abbreviations
v
PEG
-
Polyethylene glycol
PFU - Plague forming units
pI - Isoelectric point
PM - Pitman Moore
PMSF - Phenylmethyl sulfonyl fluoride
PPIC - Plant protease inhibitor cocktail
pr-s - Pathogen responsive signal sequence
PTGS - Post transcriptional gene silencing
PV - Pasteur Virus
PVDF - Polyvinylidenedifluoride
PVP - Polyvinyl pyrrolidone
RCGM - Review committee on genetic
manipulation
RGP - Rabies glycoprotein
rgp - Rabies glycoprotein gene
RNA - Ribonucleic acid
RNase-A - Ribonuclease-A
RNP - Ribonucleoprotein
RTB - Ricin toxin B chain protein
rtxB - Ricin toxin B chain gene
SDS - Sodium dodecyl sulphate
Sec - Second
SSC - Saline sodium citrate
STE - Saline Tris-EDTA
T - Thymine
TAE - Tris-acetate-EDTA
TBE - Tris-borate-EDTA
TCA - Trichloro acetic acid
TE - Tris-EDTA
TEMED - N,N,N’,N’-tetramethyl-ethyl-ethlene
diamine
TFA - Trifluoro acetic acid
TGS - Transcriptional gene silencing
TMV - Tobacco Mosaic Virus
Tnos - Nos terminator
Tris - Tris (hydroxymethyl) aminomethane
U - Unit
U - Unit
UV - Ultra-violet
V - Volume
VNA - Virus neutralizing antibodies
VSV - Vesicular stomatitis virus
W - Weight
WHO - World Health Organisation
X-gal - 5-Bromo-4-chloro-3-indolyl--D-
galactoside
Page 10
List of figures
vi
List of figures
Chapter 1
Figure 1.1: Worldwide distribution of rabies.
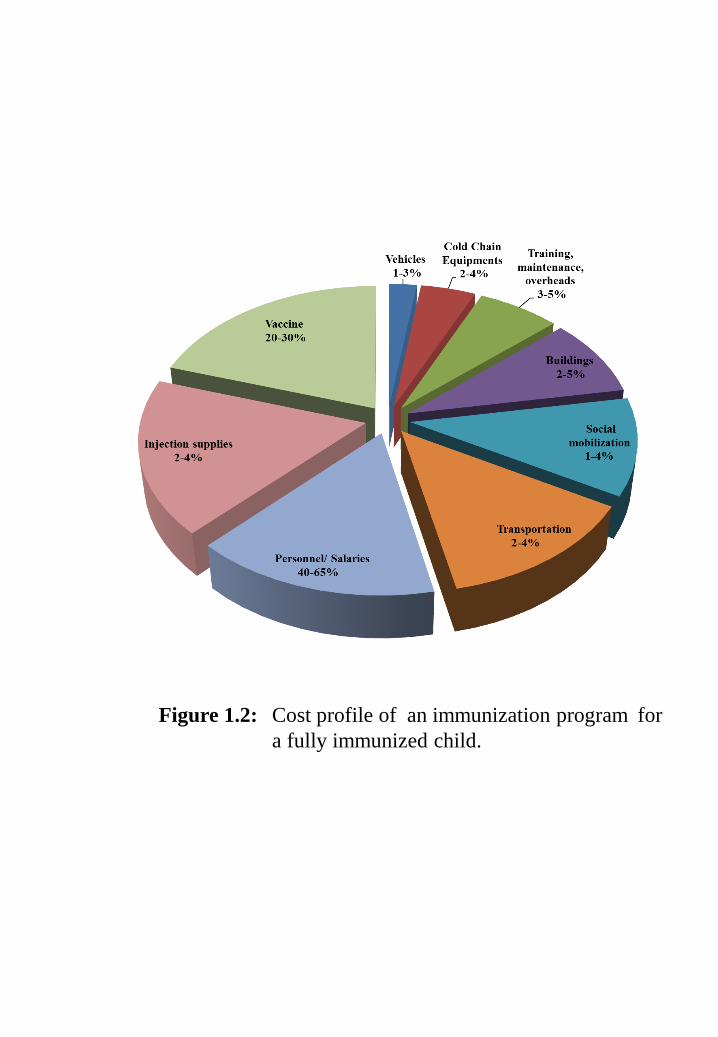
Figure 1.2: Cost profile of an immunization program for a fully immunized child.
Chapter 2
Figure 2.1: Structure of rabies virus. Rabies virions are bullet shaped with 10 nm
spike like glycoprotein peplomers covering the surface. The ribo-
nucleoprotein is composed of RNA encased in nucleoprotein,
phosphoprotein and polymerase.
(A) Photograph is adapted from Centers for Disease Control and
Prevention;
(B) Negatively stained rabies virus seen by transmission electron
microscopy from the Wadsworth Center of the New York State,
Department of Health;
(C) Diagrammatic representation of the rabies virion deduced from
electron microscopy and protein analysis by Vernon et al. 1972.
Chapter 3
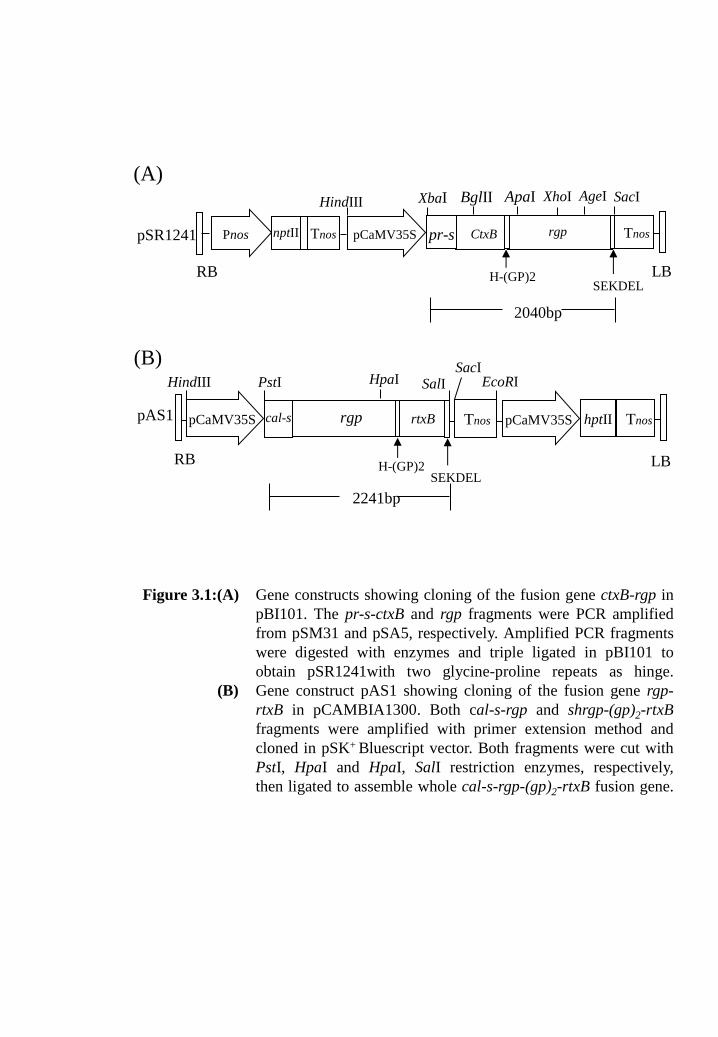
Figure 3.1: (A) Gene constructs showing cloning of the fusion gene ctxB-rgp in
pBI101. The pr-s-ctxB and rgp fragments were PCR amplified
from pSM31 and pSA5, respectively. Amplified PCR fragments
were digested with enzymes and triple ligated in pBI101 to
obtain pSR1241with two glycine-proline repeats as hinge.
(B) Gene construct pAS1 showing cloning of the fusion gene rgp-
rtxB in pCAMBIA1300. Both cal-s-rgp and shrgp-(gp)2-rtxB
fragments were amplified with primer extension method and
cloned in pSK+
Bluescript vector. Both fragments were cut with
PstI, HpaI and HpaI, SalI restriction enzymes, respectively, then
ligated to assemble whole cal-s-rgp-(gp)2-rtxB fusion gene.
Page 11
List of figures
vii
Chapter 4
Figure 4.1: A. tumefaciens strain LBA4404 containing pSR1241and pAS1
plasmid was used for transformation of tobacco leave discs.
(A) Transgenic shoot induction from leave discs.
(B) Transgenic shoots elongation and their respective selection on
Kanamycin and Hygromycin containing media.
(C) Acclimatized mature transgenic tobacco plants in glass house.
Figure 4.2: (A) PCR detection for stable integration of ctxB-rgp fusion gene in
genomic DNA of the transgenic tobacco lines. M, λ DNA
marker, PC, positive control (plasmid DNA); lane 1-5, transgenic
tobacco lines and NT is non-transgenic tobacco.
(B) PCR amplification of rtxB from stable integration of rgp-rtxB
fusion gene in genomic DNA of the transgenic tobacco lines. M,
PC and NT are showing 100bp marker, plasmid DNA and non-
transgenic tobacco lines, respectively.
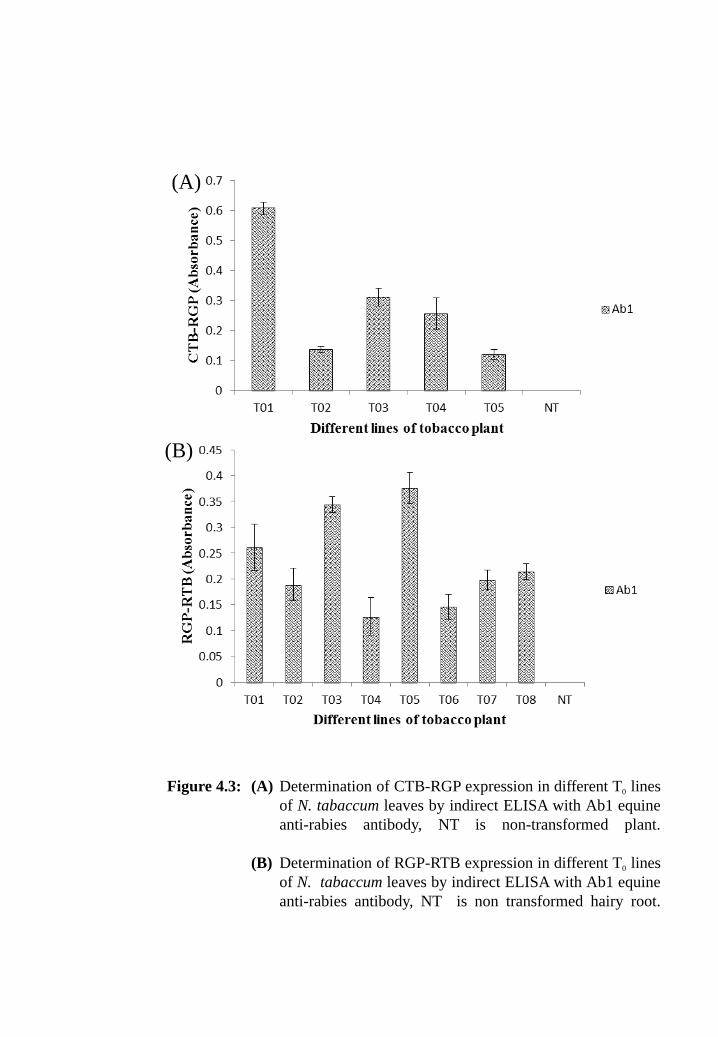
Figure 4.3: (A) Determination of CTB-RGP expression in different T0 lines of N.
tabaccum leaves by indirect ELISA with Ab1 equine anti-rabies
antibody, NT is non-transformed plant.
(B) Determination of RGP-RTB expression in different T0 lines of N.
tabaccum leaves by indirect ELISA with Ab1 equine anti-rabies
antibody, NT is non-transformed hairy root.
Figure 4.4: (A) Plasmid pSA1 containing, Agrobacterium rhizogens strain A4
mediated induction of hairy root culture from the leaves of
Solanum lycopersicum grown in ¼ MSP solid media.
(B) Transformed in vitro grown roots of Solanum lycopersicum after
grown in 28th
days in ¼ MSL in liquid media.
(C) Scale-up process of selected hairy root line (H03) in 5L air lift
bioreactor for large production and isolation of candidate protein.
Page 12
List of figures
viii
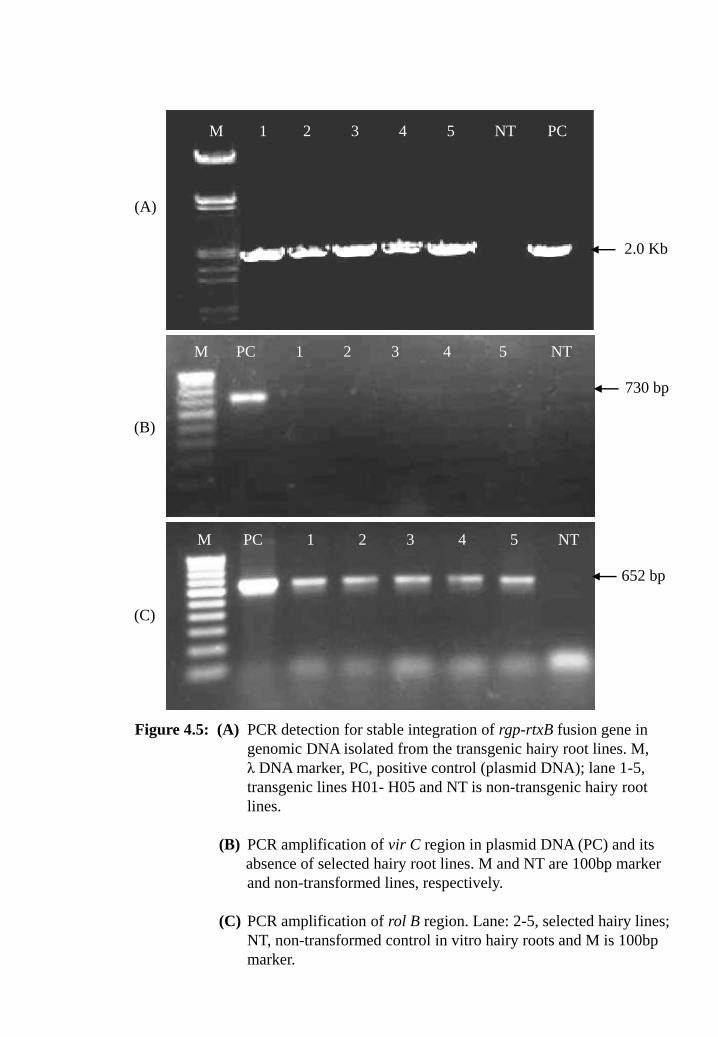
Figure 4.5: (A) PCR detection for stable integration of rgp-rtxB fusion gene in
genomic DNA isolated from the transgenic hairy root lines. M, λ
DNA marker, PC, positive control (plasmid DNA); lane 1-5,
transgenic lines H01-H05 and NT are non-transgenic hairy root
lines.
(B) PCR amplification of vir C region in plasmid DNA (PC) and its
absence of selected hairy root lines. M and NT are 100bp marker
and non-transformed lines, respectively.
(C) PCR amplification of rol B region. Lane: 2-5, selected hairy
lines; NT, non-transformed control in vitro hairy roots and M is
100bp marker.
Figure 4.6: Determination of RGP-RTB expression in different lines of Solanum
lycopersicum hairy roots by indirect ELISA with Ab1 equine anti-
rabies antibody, NT is non-transformed hairy root.
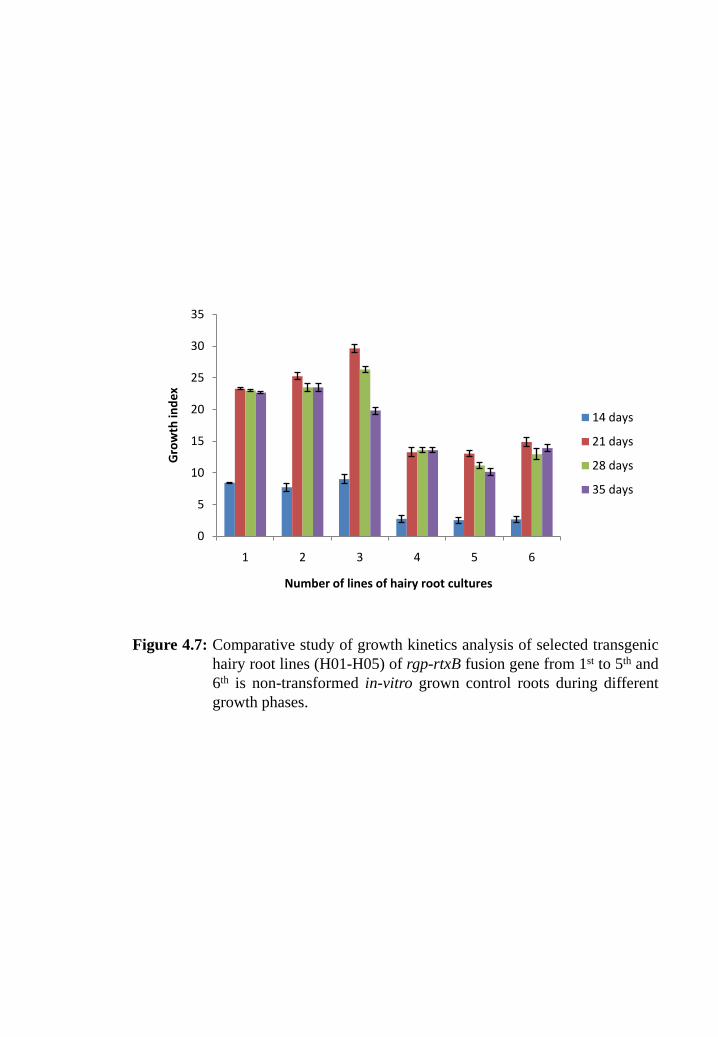
Figure 4.7: Comparative study of growth kinetics analysis of selected transgenic
hairy root lines (H01-H05) of rgp-rtxB fusion gene from 1st to 5
th and
6th
is non-transformed in-vitro grown control roots during different
growth phases.
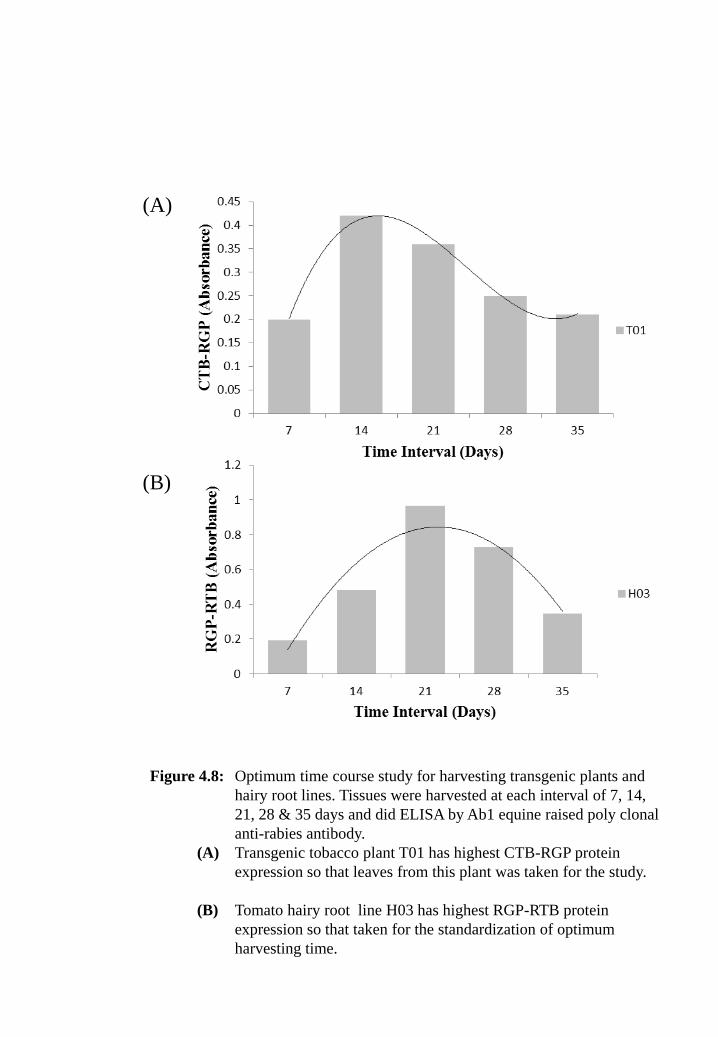
Figure 4.8: Optimum time course study for harvesting transgenic plants and hairy
root lines. Tissues were harvested at each interval of 7, 14, 21, 28 &
35 days and did ELISA by Ab1 equine raised poly clonal anti-rabies
antibody.
(A) Transgenic tobacco plant T01 has highest CTB-RGP protein
expression so that leaves from this plant was taken for the study.
(B) Tomato hairy root line H03 has highest RGP-RTB protein
expression so that taken for the standardization of optimum
harvesting time.
Page 13
List of figures
ix
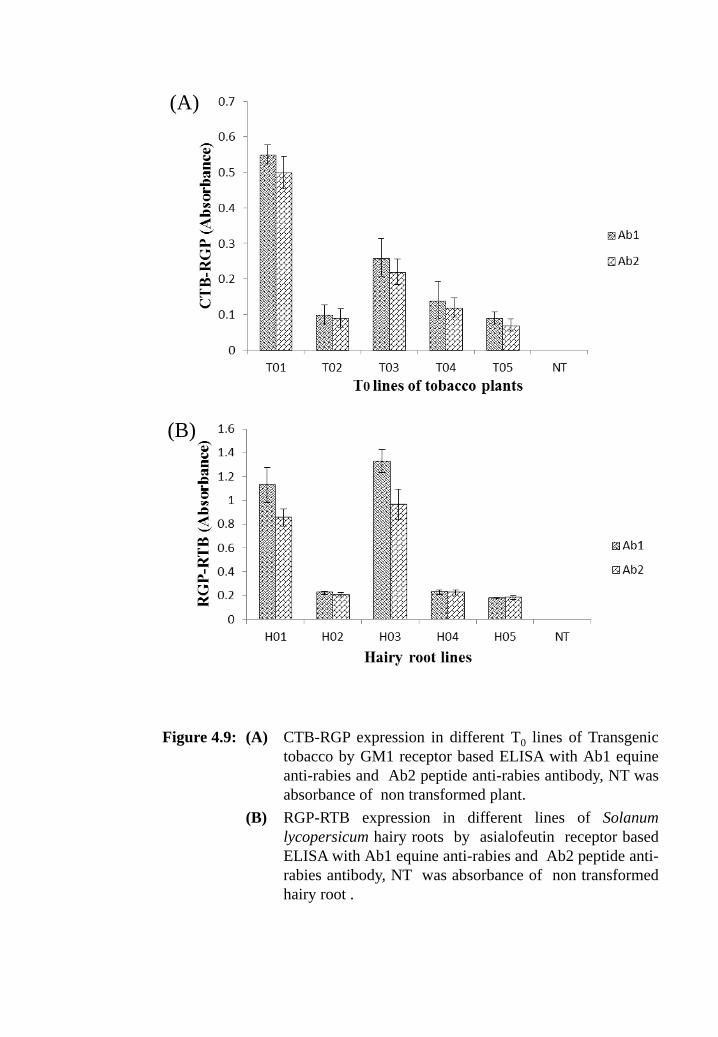
Figure 4.9: (A) CTB-RGP expression in different T0 lines of transgenic tobacco
by GM1 receptor based ELISA with Ab1, equine anti-rabies and
Ab2, peptide anti-rabies antibody; NT was absorbance of non-
transformed plant.
(B) RGP-RTB expression in different lines of Solanum lycopersicum
hairy roots by asialofeutin receptor based ELISA with Ab1,
equine anti-rabies and Ab2, peptide anti-rabies antibody; NT was
absorbance of non-transformed hairy root.
Figure 4.10: Quantitative expression of CTB-RGP (A) and RGP-RTB (B) fusion
proteins in their respective high expression transgenic lines by
receptor mediated ELISA with peptidal anti-rabies antibody (Ab2).
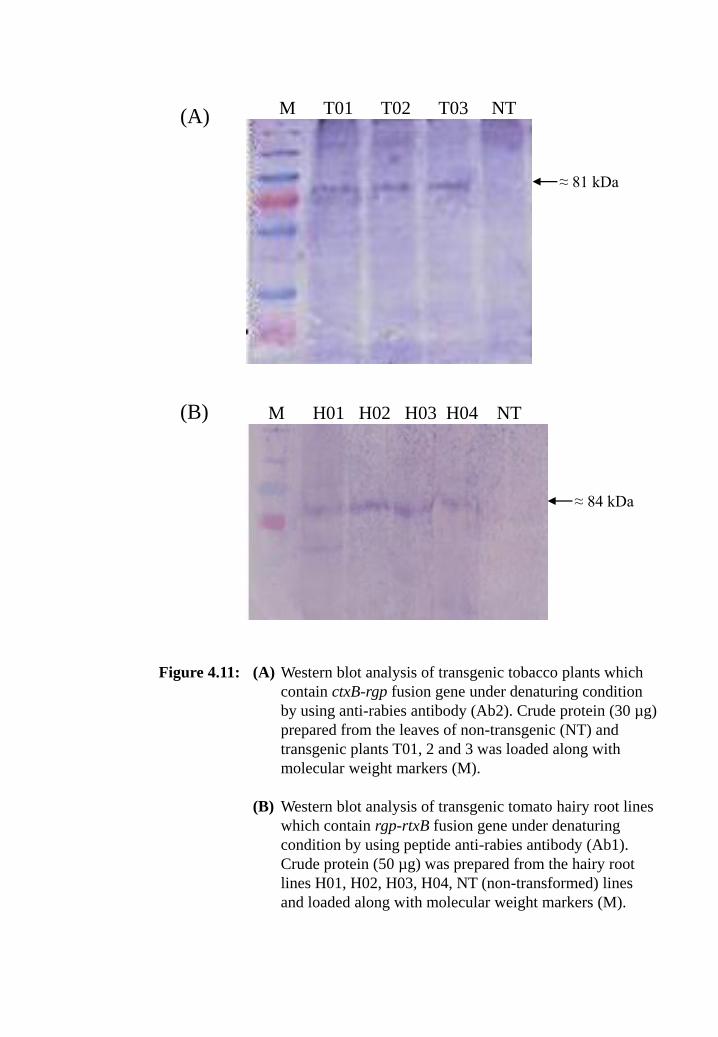
Figure 4.11: (A) Western blot analysis of transgenic tobacco plants which contain
ctxB-rgp fusion gene under denaturing condition by using anti-
rabies antibody (Ab2). Crude protein (30 µg) prepared from the
leaves of non-transgenic (NT) and transgenic plants T01, 2 and 3
was loaded along with molecular weight markers (M).
(B) Western blot analysis of transgenic tomato hairy root lines which
contain rgp-rtxB fusion gene under denaturing condition by using
peptide anti-rabies antibody (Ab1). Crude protein (50 µg) was
prepared from the hairy root lines H01, H02, H03, H04, NT (non-
transformed) lines and loaded along with molecular weight
markers (M).
Figure 4.12: Western blot analysis of transgenic lines under non-denaturing
condition by using polyclonal anti-rabies antibody (Ab2).
(A) Crude protein (30 µg) was prepared from the tobacco line of ctxB-
rgp fusion gene (T01), loaded at lane 2; M, marker and NT is non-
transformed line.
Page 14
List of figures
x
(B) Crude protein (50 µg) was prepared from the tomato hairy root
line of rgp-rtxB fusion gene (H03), loaded at lane 3; M, marker
and NT is non-transformed line.
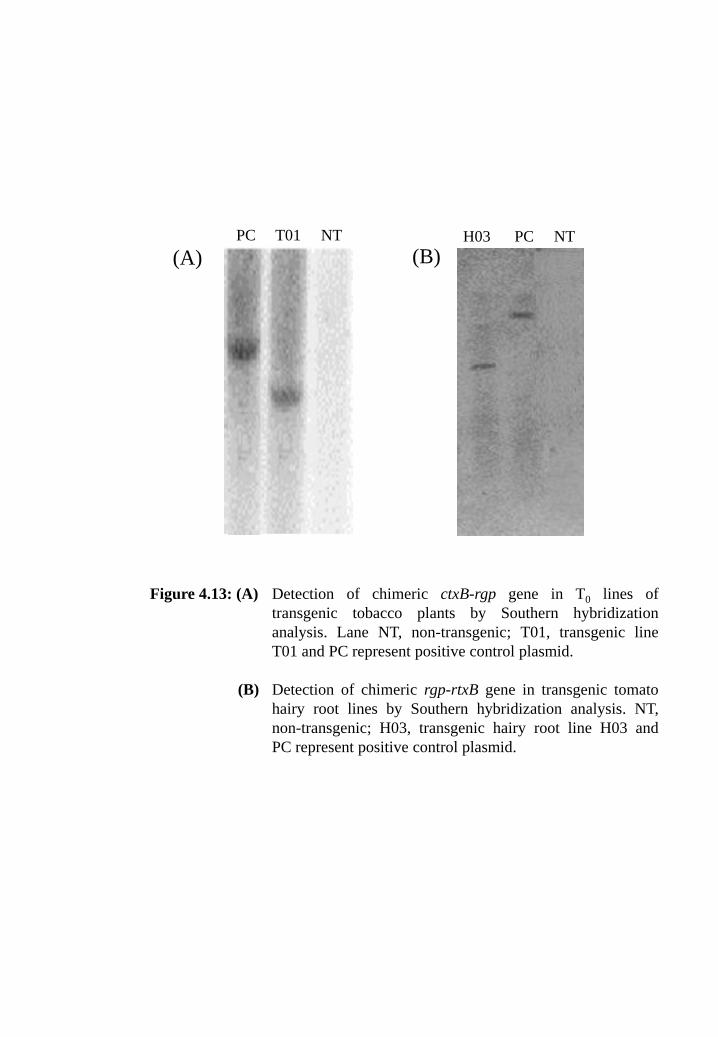
Figure 4.13: (A) Detection of chimeric ctxB-rgp gene in T0 lines of transgenic
tobacco plants by Southern hybridization analysis. Lane NT, non-
transgenic; T01, transgenic line T01 and PC represent positive
control plasmid.
(B) Detection of chimeric rgp-rtxB gene in transgenic tomato hairy
root lines by Southern hybridization analysis. NT is non-
transgenic; H03, transgenic hairy root line of H03 and PC
represent positive control.
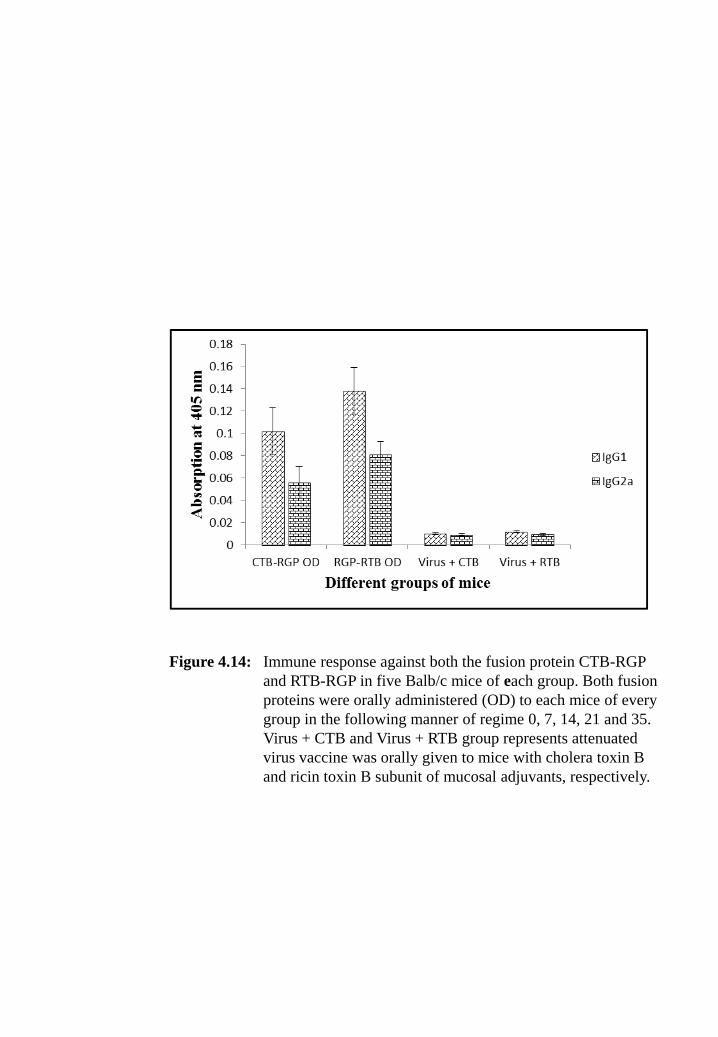
Figure 4.14: Immune response against both the fusion protein CTB-RGP and RTB-
RGP in five Balb/c mice of each group. Both fusion proteins were
orally administered (OD) to each mice of every group in the
following manner of regime 0, 7, 14, 21 and 35. Virus + CTB and
Virus + RTB group represents attenuated virus vaccine was orally
given to mice with cholera toxin B and ricin toxin B subunit of
mucosal adjuvants, respectively.
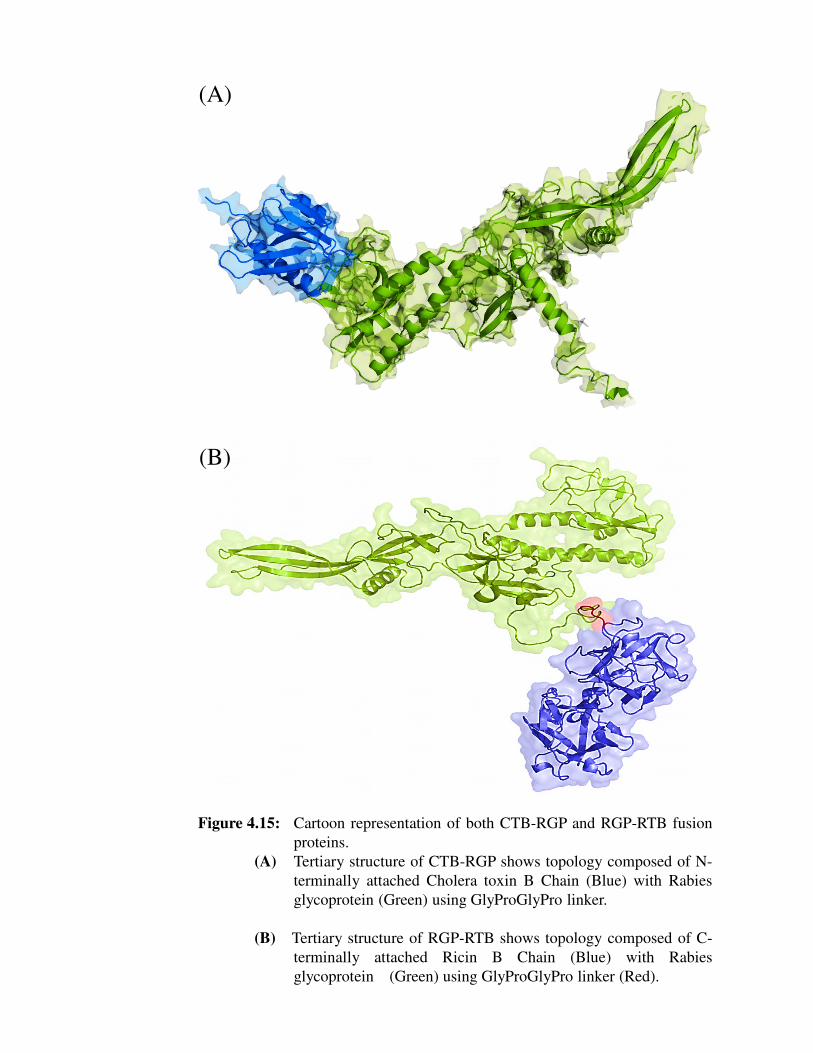
Figure 4.15: Cartoon representation of both CTB-RGP and RGP-RTB fusion
proteins.
(A) Tertiary structure of CTB-RGP shows topology composed of N-
terminally attached Cholera toxin B Chain (Blue) with Rabies
glycoprotein (Green) using GlyProGlyPro linker.
(B) Tertiary structure of RGP-RTB shows topology composed of C-
terminally attached Ricin B Chain (Blue) with Rabies
glycoprotein (Green) using GlyProGlyPro linker (Red).
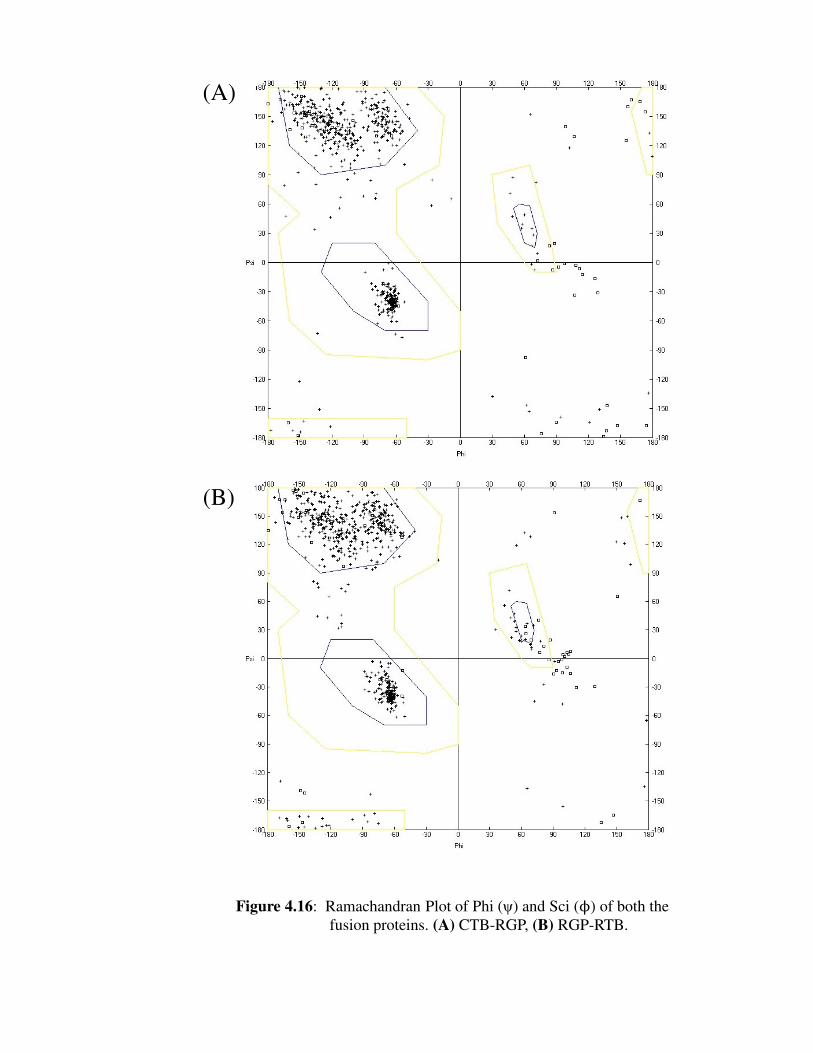
Figure 4.16: Ramachandran Plot of Phi (ψ) and Sci (ϕ) of both the fusion proteins
(A) CTB-RGP; (B) RGP-RTB.
Page 15
List of figures
xi
Figure 4.17: Superimposition studies of overall cartoon structure of both the fusion
proteins.
(A) Tertiary structure of Cholera toxin B Chain and Rabies
glycoprotein shown in Blue and Green in fusion protein CTB-
RGP while superimposed CTB and RGP of PDB database shown
in Sky Blue and pink colours, respectively.
(B) Tertiary structure of Ricin toxin B Chain and Rabies
glycoprotein shown in Orange and Green in fusion protein
RGP-RTB while superimposed RTB and RGP of PDB database
shown in Sky Blue and Blue colours, respectively.
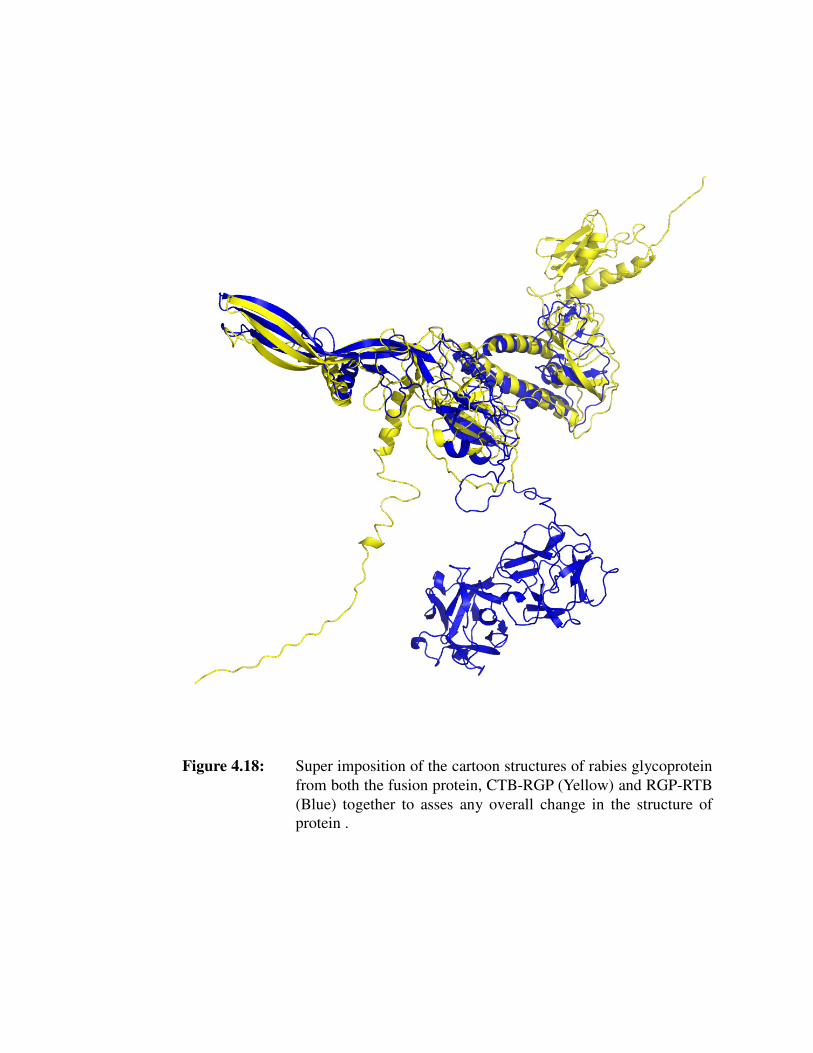
Figure 4.18: Super imposition of the cartoon structures of rabies glycoprotein from
both the fusion protein, CTB-RGP (Yellow) and RGP-RTB (Blue)
together to asses any overall change in the structure of both the
protein.
Figure 4.19: Surface representation of antigenic sites of RGP on both CTB-RGP
(A) and RGP-RTB (B) fusion proteins. Conformationally important
sites K202 and R336 of RGP shows in Hot Pink (A) / Yellow color
(B) and Brown (A) / Orange (B) color, respectively. Whereas
important antigenic sites A (34-42aa), site B (198-200aa), site C (208-
216aa) and D (286-306aa) represents as Red, Blue, Raspberry Red
and Cyan color, respectively.
Figure 4.20: Hopp-Wood (HW) and Surface Exposure (SE) plots for CTB-RGP
and RGP-RTB fusion proteins. A (32-42), B (198-200), C (208-216)
and D (286-306) represent potent antigenic sites as described by other
research groups.
Page 16
List of tables
xii
List of tables
Chapter 2
Table 2.1: Predominant global rabies reservoirs
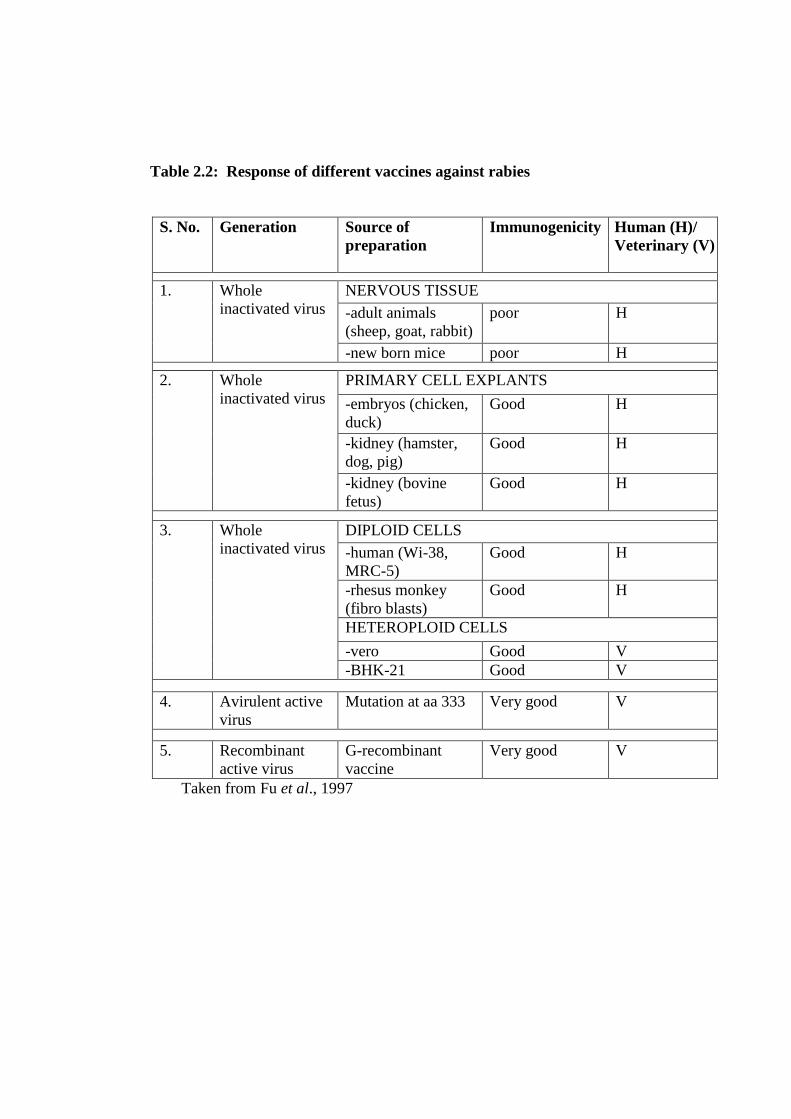
Table 2.2: Response of different vaccines against rabies
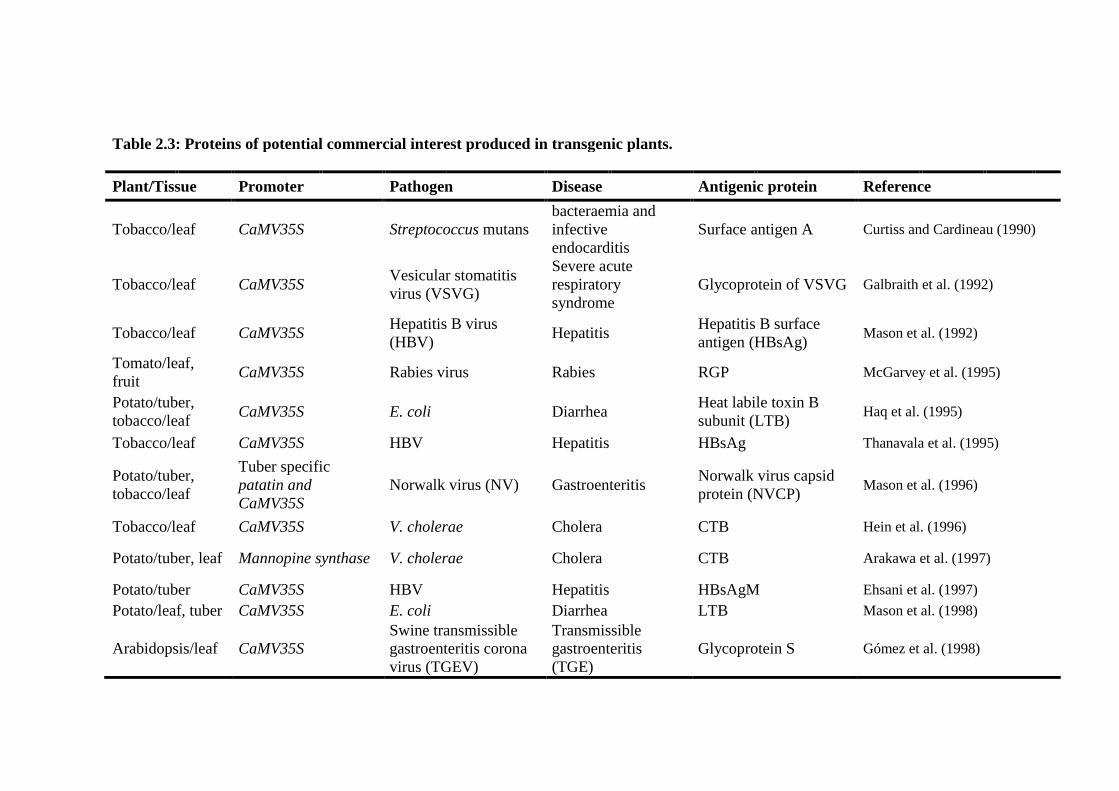
Table 2.3: Proteins of potential commercial interest produced in transgenic
plants
Chapter 3
Table 3.1: List of primers used in rgp-rtxB fusion gene construction
Chapter 4
Table 4.1: Comparative analysis of growth performance and protein expression
Table 4.2: Comparative analysis of CTB-RGP and RGP-RTB fusion proteins by
ProtParam software
Page 17
Chapter 1
Introduction
Page 18
Introduction
1
1.0 INTRODUCTION
The plants are considered as the most renewable production resources for a
variety of proteins, fats, essential amino acids, dyes, drugs, chemicals etc. The advent of
various plant biotechnology techniques, such as, modern breeding methods, cell and
tissue culture, somaclonal variation, clonal propagation, protoplast culture, somatic
hybridization and genetic transformation have played a vital role in establishing plant-
based industries for the production of above mentioned high value added compounds.
The first pharmaceutically relevant protein made in plants was human growth hormone
expressed in transgenic tobacco in 1986. Since then transgenic plants are rapidly
emerging as smart bioreactors for the production of protein drugs, enzymes and
biopolymers for medical and industrial applications. The first plant derived technical
protein has already reached to the market (Hood et al., 1997, Witcher et al., 1998).
Detailed economic evaluations of these proteins have demonstrated their
competitiveness against corresponding market sectors (Kusnadi et al., 1997, 1998).
Several other plant-derived bio-pharmaceutical proteins are on the terminal end of
pipeline for commercial production. These products include antibodies, vaccines,
human blood products, hormones and growth regulators (Fischer et al., 1999; Gidding
et al., 2001). Plants that express foreign proteins with industrial or pharmaceutical value
represent an economical alternative to fermentation-based production systems.
Transgenic plants are raised and cultivated on a large scale for manufacturing
vaccines against both viral and bacterial pathogenic agents of human and poultry
animals. Many proteins including viral (Norwalk virus capsid protein), bacterial
(Pseudomonas aeruginosa outer membrane protein), enterotoxin (E. coli heat-labile
enterotoxin) and non-enteric antigens (Hepatitis B surface antigen) as well as
autoimmune antigens have been produced in an increasingly diverse range of crops. A
few research groups (Hiatt et al., 1989; 1993, During et al., 1993) have also identified
plants as manufacturing units for producing properly assembled heavy and light chains
of certain antibodies. These approaches would prove extremely beneficial for the
developing and the underdeveloped countries.
Vaccination involves the stimulation of the immune system to prepare it for the
event of an invasion from a particular pathogen for which the immune system has been
primed (Masson et al., 1995; 1998). This will generate and prepare pathogen specific T
Page 19
Introduction
2
and B cells for rapid proliferation and differentiation when natural pathogens will be
encountered by immune system. Construction of vaccine in several cases has been
hampered because of varying strains of the pathogen, antigen drift, antigenic shift and
other unrevealed mechanisms that make it hard to determine a suitable peptide sequence
for the priming of immune system.
Although, live attenuated and killed pathogens were used frequently to prime
the immune system but that has resulted in acquiring the same disease by few people
after vaccination. Recombinant subunit vaccines are desirable as an alternative with
potentially fewer side effects than delivering the whole organism. Recombinant subunit
vaccines do not contain whole infectious agent, and thus are safer to administer and
prepare, and doses are more uniform. Advances in molecular biology of diseases have
identified many candidate proteins or peptides that may function as effective subunit
vaccines. DNA was also used as subunit vaccine to prevent and slow down the spread
of disease, known as polynucleotide immunization or DNA vaccine. DNA that is
injected into the subject undergoes the transcription and translation which yield protein,
making the specific T and B cells to differentiate and proliferate. Therefore, the
immune system is ready to combat invading pathogen very quickly, before they get the
chance to spread throughout the body while causing discomfort to the host. Though
most of the above mentioned methods of vaccination have been effective against
diseases for which vaccines can be produced, the introduction of oral vaccines was
made to ease the discomfort associated with the mode of introduction of conventional
vaccines into the body.
Plant-derived edible vaccines have an advantage of being used orally. They have
been observed to stimulate production of mucosal antibodies more effectively than the
injected vaccines. The mucosal immune system is a part of the body’s first line of
defense against many disease-organisms. A mucosal tissue comprises all lymphoid cells
in epithelia and cells lying below the body’s mucosal surfaces. The main site of
mucosal lymphoid tissues is associated with the bronchial system and gut. To stimulate
this system in the gut, an oral vaccine has to be protected from degradation at acidic pH
of the intestine (stomach) which ultimately stimulates mucosal immune system and
results in the production of secretory antibody i.e. IgA. The secretory antibodies
comprise 75% of the total antibody synthesized and secreted in the form of tears, saliva
Page 20
Introduction
3
and milk. These properties of the edible vaccine are gifted by the associated plant
tissues, which protect vaccines from degradation in the digestive tract.
In the developing and the underdeveloped countries, the cost of vaccines is a
limiting factor. A large majority of people remains unprotected against preventable
disease because they cannot afford. Vaccine production, packaging, delivery,
administration by trained people and refrigeration, which are prerequisite for shipment
or storage increase its cost (Figure 1.1). Similar economic factors apply also to large-
scale vaccination of farm animals. The advantages of biopharmaceutical proteins are
not limited to cost savings. The molecular farming of pharmaceuticals in plants
eliminates the risk of human or animal virus and prion transmission or of contamination
by harmful products of chemical synthesis or process solvents.
The current large-scale vaccines producing industries use mostly bacteria, yeast,
insect and mammalian cells and transgenic animals as production resources. Vaccines
expressed in these wild/genetically-engineered systems are essentially processed to
remove host proteins, toxins as well as other noxious compounds, which contribute to
their high cost. In addition, these production systems are highly prone to contamination
by microbes that sometimes evade detection even in the purified vaccines. Hence, these
systems cannot be considered ideal for the production and delivery of quality and
economically viable vaccines.
Designing plant-derived vaccines therefore becomes an important alternative to
meet the increasing market demand of cheaper, safer and quality vaccines. The
successful development of vaccines against human animal pathogen(s) depends
primarily on establishment of stable transformation procedure for widely and
commonly grown crops, ease in scale up of contaminant free production of antigenic
proteins. Production of the vaccines is based on two strategies: (i) Stable
transformation of plant cells with gene(s) encoding antigenic proteins and (ii) its
transient expression. Transgenic plants, especially of the edible variety, provide an
excellent alternative for the production of desirable vaccines. This system also allows
easy and economic scale-up of production by cultivating more of these plants in the
field. In addition, the continuous refinements in the plant genetic engineering
techniques and an increasing understanding of plant molecular biology have helped
improve the genetic design of plants with greater capacity for production of vaccines.
Page 21
Figure 1.1: Worldwide distribution of rabies.
Page 22
Figure 1.2: Cost profile of an immunization program for
a fully immunized child.
Page 23
Introduction
5
enhanced presentation of the conjugated molecule to the immune system (Holmgren et
al., 1975; Nashar et al., 1996). Roy et al. (2010), successfully demonstrated
immunogenicity of cholera toxin B subunit (CTB) as N-terminal fusion partner with the
rabies glycoprotein. Seed specific promoter was also used to enhance the expression of
CTB-RGP expression in ground nut which is important for edible version of fusion
vaccine (Tiwari et al., 2009b).
Similar to the function of CTB, ricin toxin B subunit (RTB) was also used as the
mucosal adjuvant and carrier to enhance immune responses for rota virus infection
(Yu and Langridge, 2001). RTB is able to serve as C-terminal fusion carrier for delivery
of the virus antigen to the mucosal immune system and may act as a potential immune
modulator to enhance the mucosal immune response of antigens (Medina-Bolivar et al.,
2003). However, before proceeding for next step or animal experiments, we have to
check the immunogenic property of both N- and C- terminal fused rabies glycoprotein
Ricin toxin (RT) is composed of a galactose binding B chain (Mr =32,000) and a
cytotoxic A chain (Mr=30,500). RT toxicity is based on A subunit inhibition of protein
synthesis by ribosome in activation. The ricin A chain is a glycosidase that catalyzes the
removal of a single adenine residue from a highly conserved loop of the 28S ribosomal
RNA (A 4324 in rat 28S RNA) (Endo and Tsurugi, 1988). Interaction between sugar
binding moieties on the ricin B chain and terminal galactosides located on the
enterocyte membrane facilitates ricin holotoxin uptake by endocytosis into intra cellular
vesicles (Lambert et. al., 1991). Monomeric ricin B sub-unit has an advantage that they
do not require assembly in to multimeric structures prior to receptor binding as required
in CTB or LTB (Falnes and Sandvig, 2000). RTB has a broader receptor binding
specificity for membrane receptors than CTB or LTB (Falnes and Sandvig, 2000;
Chazaud et. al., 1995; Holmgren et al., 1975). RTB binds to glycoproteins, which is
found on epidermal cell membranes from about 1×107
to 3×107 molecules per cell
(Falnes and Sandvig, 2000). Both CTB and LTB bind to GM1 receptors at lower
frequencies, about 7.5×104 molecules/mucosal epidermal cell (Holmgren et al., 1975).
In contrast to CTB and LTB, the monomeric RTB subunit does not exert fusion protein
size constraints based on oligomer assembly. RTB has the potential for delivery of more
and larger antigen molecules to gut epithelial cells than either CTB or LTB. RTB has
been cloned and expressed in various organisms, e.g. Escherichia coli, Saccharomyces
Page 24
Introduction
6
cerevisiae, Xenopus laevis oocytes and Nicotiana tabacum (Tagge et. al., 1996; Frankel
et. al., 1996 b; Tonevitsky et. al., 1994; Wales et. al., 1991). C-terminal fusion of RTB
with green fluorescent protein (GFP) has been synthesized in transformed tobacco and
found to generate a humoral immune response showing the presence of a Th2 response
in intranasally immunized mice (Medina-Bolivar et al., 2003). Thus, the membrane
targeting RTB sub unit of plant hetero-dimeric AB toxin from R. communis may have
the capacity to serve as a carrier for subunit vaccines. In this study we used ricin toxin
B subunit (RTB) to exploit their fusion ability as a C-terminal fusion and utilize them as
mucosal carrier and have the ability to bind with the receptors even in a single
monomer condition.
Contamination like plant pigments and low expression are the major limiting
factors for producing pharmaceutical proteins in plants. So, the technology which
provides stable integration lines and consistent with the expression level of proteins is
needed. To overcome this problem we use hairy root culture of Solanum lycopersicum
for producing RGP-RTB recombinant fusion protein. Although, Agrobacterium
rhizogene- induced transgenic hairy root cultures are not much popular (Doran PM,
2000; Gidding et al., 2000), but definitely show great advantages over plant cell
cultures and other tissues in terms of protein production (Sharp and Doran, 1999;
Wongsamuth and Doran 1997). Long term production of foreign proteins through hairy
root culture is possible due to its genetic stability and is preferred over bacteria and
yeast on the basis of glycosylation and folding accuracy (Fischer et al., 1999).
Successful establishment of such transformed root cultures holds great potential and is a
significant promise as a laboratory tool, especially for basic mechanistic studies and
quick screening of plants with superior expression of the gene of interest. Maintenance
of these hairy roots in a bioreactor is easily scalable as per the demand and avoids
further stringent regulatory approval due to in-vivo condition which give it a greater
edge over seed based platform.
This study is mainly focused on to check the efficacy of plant produced rabies
glycoprotein with fused mucosal adjuvants and aims to evaluate any change in
immunogenic property of rabies glycoprotein on the fusion of N- and C-terminal
orientation of mucosal adjuvants, hence, providing relevant information for developing
plant based oral vaccine against rabies.
Page 25
Chapter 2
Review of Literature
Page 26
REVIEW OF LITERATURE
7
2.0 REVIEW OF LITERATURE
2.1 History of rabies
Rabies is one of the oldest known diseases of mankind. The word, “rabies”
comes from the Sanskrit word “rabbahs” which means, “to do violence”. Rabies in
India is known since Vedic period as corroborated in Atherva Veda. Yama, the God of
Death in Hindu mythology, has been shown to be attended by dogs as his constant
companions and as emissaries of death (Ahuja et al., 1983). The first description of the
disease dates from the 23rd
century BC in the Mesopotamian Laws of Eshnuma (Fu,
1997), yet it has been only slightly more than 120 years (1885) since Pasteur developed
the first vaccine for post-exposure treatment. Since this first crude nerve tissue vaccine,
numerous other vaccines for human use have been developed. These are used with
varying degrees of effectiveness and safety but we are still facing an epidemic situation
of rabies today. In spite of numerous efforts 35,000-50,000 human fatal cases per year
are reported (WHO, 1994). Actual numbers may be much higher. This makes rabies as
one of the most mortal infectious diseases. Rabies has been and even today is regarded
in the world as one of the most terrifying diseases, and India is no exception to it.
Despite the early discovery of the rabies vaccine, this disease is still a problem today.
Moreover, the new and effective vaccines are often unaffordable for the average
citizen. In addition to these unsolved problems, emerging rabies-related viruses have
recently been described (Fraser et al., 1996), which emphasise the importance of rabies
research.
2.2 Global rabies reservoirs
Rabies is one of the most important zoonotic diseases. Rabies is a worldwide
public health hazard. The most important reason why rabies is still endemic in the huge
global reservoirs, in both domestic and wildlife animals, all mammals are thought to be
susceptible to infection, but reservoirs important to the maintenance and transmission
of rabies virus are limited to the Carnivora and Chiroptera (Rupprecht et al., 2002).
In most of the developing countries, dogs represent the major rabies reservoir.
Dogs remain the most important reservoirs in Asia, Africa and Latin America, where
most human rabies cases occur. Rabies in the wildlife presents a more challenging
problem. In North America, reservoirs of rabies exist in many diverse animal species
Page 27
REVIEW OF LITERATURE
8
(Meslin et al., 1994; Rupprecht et al., 1995). The most frequently reported rabid
wildlife species in USA are raccoons (50.5%), followed by skunks (24.0%) (Rupprecht
et al., 1995). Outbreaks of rabies infections in these terrestrial animals are found in
broad geographical areas in the United States (Meslin et al., 1994). Fox rabies has been
endemic in Europe and North America for many years, although recent endeavours in
oral vaccination through vaccines in an edible bait has been successful in reducing or
even eliminating rabies in many parts of Europe. Recently, raccoon rabies has spread
from a focal point in Florida in the 1970s to all the eastern States in USA by 1990s.
There are other important reservoirs, including coyotes in Asia, Africa and North
America, Skunks in North America, and mongoose in Asia, Africa and the Caribbean
islands. Rabies is also endemic, the vampire bats from Mexico to Argentina and the
insectivorous bats in North America and Europe. The vampire bats transmit rabies to
cattle and cause tremendous problems for the agricultural industry in South America.
Many human rabies cases in the United States have been caused by rabies virus variants
circulating in the insectivorous bat populations. Insectivorous bats in Europe have been
reported to carry rabies-related viruses (Fu, 1997). Bat- associated rabies viruses cause
sporadic disease in humans and livestock species, and major epidemics in terrestrial
mammals are relatively commonplace (Childs et al. 2000). Humans serve only as
accidental hosts. Overlying the disease in terrestrial animals are multiple independent
reservoirs for rabies in several species of bats. As in terrestrial species, distinct viral
variants can be identified for different bat species. However, geographic boundaries
cannot be well defined (Smith, 1996). The most predominant rabies reservoirs are listed
in table 2.1 (Fu, 1997).
Rabies viruses form following two types of association with their host species
(i) The virus establishes a stable infection cycle within a particular mammalian
species, most notably observed in carnivorous mammals (dogs, foxes, raccoons,
and skunks) as well as in a variety of bat species. Whether the virus always causes
disease in these is not clear; although fatal rabies is common in infected dogs,
foxes and raccoons; the same does not always appear to be true of bats (Baer and
Lentz, 1991).
(ii) The second form of virus-host interaction occurs when the virus jumps species
boundaries to infect new hosts. The example of these “spill-over” infections is
Page 28
Table 2.1: Predominant global rabies reservoirs
Dogs Major vector of rabies throughout the world, particularly Asia, Latin
America and Africa.
Foxes Europe, Arctic and North America.
Raccoons Eastern United States.
Skunks Midwestern United States, Western Canada.
Coyotes Asia, Africa and North America.
Mongooses Yellow mongoose in Asia and Africa; Indian mongoose in the
Caribbean Islands.
Bats Vampire bats from Northern Mexico to Argentina, insectivorous bats
in North America and Europe.
Taken from Fu et al., 1997.
Page 29
Introduction
4
Rabies is acute progressive encephalitis. At least, 60,000 human deaths occur
worldwide annually from rabies (Meslin and Stohr, 1997). It is caused by a
promiscuous neurotropic virus of Lyssavirus genus of family Rhabodoviridae. The
disease spreads through domestic and wild animals. Rabies is a major zoonosis of
significant public health concern in many parts of the world, especially in developing
countries (Figure 1.2) where rabies is endemic among dogs (Meslin and Stohr, 1997).
The first rabies vaccine, consisted of subcutaneous inoculation of spinal cord
suspension, derived from rabid rabbits (Pasteur, 1885). Since then, a continuous effort
is going on for the improvement of the vaccine (Perrin et al., 1990; Plotkin et al., 1993).
The virus genome encodes five major proteins of which the G-protein of rabies virus
has been identified as the major viral antigen that induces protective immunity (Cox et
al., 1977). In recent years, plants are emerging as a promising alternative source for
producing safe and cost effective therapeutic proteins (Curtiss and Cardineau, 1990;
Mason et al., 1992). Recombinant proteins expressed in plants have shown sufficient
promise to warrant human clinical traits (Tiwari et al., 2009). In case of rabies
glycoprotein, stable expression in tomato plants has already been reported (McGarvey
et al., 1995), while complete protection has been shown when mice were injected with
rabies G-proteins expressed and purified from tobacco plants (Ashraf et al., 2005).
Though protein was fairly active immune-protective in nature but its expression was
quite low so that downstream scale up process created an arduous task to purify this
protein in homogeneity from plants.
There are various lectins or lectin-like proteins which have binding ability to
glyco-lipids or glyco-proteins (De Aizpurua and Russell-Jones, 1988). We utilize these
lectins for administration of rabies antigen orally to avoid near homogeneity
purification and to enhance immunogenic property of rabies antigen. Many of these
lectins have already been characterized and used as mucosal adjuvant which stimulates
strong humoral as well as cell-mediated immune responses. Out of these cholera toxin
B subunit is one of the most characterized mucosal adjuvant which provides N-terminal
fusion capability with target antigens. CTB binds to the GM1 receptor and can serve as
a mucosal adjuvant (McKenzie et al., 1984), GM1 receptor being present on most of the
cells in the body including epithelial cells and leukocytes. Efficient binding to GM1
could potentially increase the uptake of antigen across the mucosa and lead to an
Page 30
REVIEW OF LITERATURE
9
human rabies. Occasionally, however, rabies viruses are able to establish
productive infections in new host species (Tordo et al., 1993; Nadin Davis et al.,
1994). An important example of such a successful host switch involved the transfer
of the virus from dogs to the red fox (Vulpes vulpes) in Northeast Europe during
1930s (Bourhy et al., 1999). After the initial cross- species transmission event,
rabies virus was able to spread rapidly westward and southward through European
Red fox populations in the subsequent 60 years (Bourhy et al., 1999).
2.3 Rabies pathogenesis
Rabies is a neurotropic virus, usually transmitted through the bite of a rabid
animal (Charlton, 1994; Dietzschold et al., 1996). Neurotropism is a major feature
associated with rabies virus infection, with viral replication restricted almost
exclusively to neuronal tissue (Murphy et al., 1973), but it causes no cytopathic effect
in cell culture (Crick and King, 1988). Rabies virus is a strict neuropathogen in vivo,
and yet it has a wide host range in vitro, infecting nearly all mammalian and avian cell
types (Wunner et al., 1984). In vivo, nicotinic acetylcholine receptor has been
suggested to be rabies virus receptor (Lentz et al., 1982). Other studies showed that the
problem is probably more complex, suggesting the existence of atleast two receptors at
the surface of nerve cells (Lafay et al., 1991). In vitro, gangliosides via their sialic acid
or phospholipids play the role of receptors (Superti et al., 1984a; 1984b; 1986; Wunner
et al., 1984). After exposure (that is dog bite) there are three critical events (Baer and
Lentz, 1991).
(1) Inoculation of the virus. Unless virus is inoculated there is no risk.
(2) The inoculated virus is adsorbed to, and gains entry into a susceptible cell and
begins to multiply.
(3) The virus multiplies in that region, and then enters the nerve endings,
particularly through acetylcholine receptors.
Although rabies virus has a strong neurotropism, replication in vivo does not
only take place in neuronal cells. Several investigators have shown that rabies virus
replicates in muscle cells prior to its invasion of the peripheral and central nervous
system as well as the salivary glands and other non-nervous tissues (Charlton and
Casey 1979; Fekadu and Schaddock, 1984 and Charlton et al., 1997); while other
Page 31
REVIEW OF LITERATURE
10
studies have demonstrated that primary infection of muscle cells may not be necessary
(Coulon et al., 1989 and Johnson, 1965). Although a rapid spread of virus from the
peripheral site of entry to the CNS with little prior replication has been reported
(Ceccaldi et al., 1989; Shankar et al., 1991).
Rabies virus has also been found to cause fusion of cells and hemolysis of
erythrocytes under acidic conditions (Mifune et al., 1982). In addition, chloroquine and
ammonium chloride have been shown to prevent infection (Superti et al., 1984a;
1984b). It has been proposed that rabies virus enters the cell via the endocytic pathway,
and subsequently fuses with the membrane of the endosome after its acidification.
However, little is known about the behaviour of the rabies virus glycoprotein during
virus internalization.
The pathogenicity of rabies virus strains depends on the presence of antigenic
determinants on the viral glycoprotein. Arginine at 333 positions in glycoprotein is
essential for the integrity of an antigenic determinant and for the ability of rabies
viruses to produce lethal infection in adult mice, so arginine at 333 of glycoprotein is
critical in the pathogenicity of rabies virus (Dietzschold et al., 1983b). Mutation of
Lysine at 330 position of glycoprotein also abolishes the penetration of the virus into
motor and sensory neurons after intramuscular inoculation of the virus (Coulon et al.,
1998).
2.3.1 Virus propagation in vivo
The path of movement of the rabies virus towards the CNS after inoculation by
bite has not been clearly demonstrated (Murphy, 1977; Tsiang 1993; Charlton, 1994).
Rabies virus seems to go through a first cycle of replication in the striated muscle cells,
where the virus is in a state of sequestration during the incubation time (Murphy and
Bauer, 1974) .It has been suggested that the infection of muscle cells is mediated by the
nicotine acetylcholine. The interaction of rabies virus with the acetylcholine receptors
at the neuromascular junction is instrumental in transfer of the virus from the periphery
to the central nervous system. However, the uptake and transport of rabies virus by a
wide variety of neuron types suggest that receptors for the virus are ubiquitous.
Morphological, immuno-cytochemical, biochemical immunological techniques have
been used to describe rabies virus binding to a sub-cellular unit and molecular complex
Page 32
REVIEW OF LITERATURE
11
at the neuromuscular junction (Burrage et al., 1985). For further spread, the virus
accumulates at neuromuscular junctions and enters the nervous system through
unmyelinated sensory and motor terminals.
Once the rabies virus enters the nervous system, it is no longer accessible to
anti- rabies antibodies (Murphy and Bauer, 1974). The ability of selected neurotropic
viruses to move transneuronally in the central nervous system makes them particularly
well suited for use as tracers in experimental neuroanatomy (Roberta and Peter, 2000;
Ugolini et al., 1989; Strack and Loewy, 1990; Hoover and Strick, 1999; Lynch et al.,
1994; Middelton and Strick, 1996; Sun et al., 1996; Jasmin et al., 1997; O’ Donnell et
al., 1997). Rabies virus moves in a time dependent manner through the central nervous
system of infected animals. The virus reaches the spinal cord by centripetal spread
(axoplasmic flow of 12 to 24 mm per day) and subsequently transported within hours to
the brainstem (Murphy et al., 1973). Although, there has been some uncertainty about
the direction of rabies transport in the central nervous system. Some investigators
proposed that rabies moves trans-neuronally in both anterograde and retrograde
directions (Gillet et al., 1986; Jackson and Reimer, 1989), and others concluded that the
virus spreads exclusively in the retrograde direction (Ugolini, 1995; Tang et al., 1999).
Rabies infections are largely confined to neurons, while ganglia infection is
rarely seen (Iwasaki and Clark, 1975; Tsiang et al., 1983). Electron microscopic studies
suggest that rabies virus is transported between neurons primarily at synaptic junctions
(Iwasaki and Clark, 1975; Charlton and Casey, 1979). Rabies virus does not cause cell
lysis. One of the hallmarks of rabies infection is the lack of pathology seen in infected
brains even at terminal stage of the disease (Murphy, 1977).
2.3.2 Virus propagation in vitro
Neuronal tropism is also observed in vitro with street rabies virus isolates
extracted from salivary glands or from the brains of rabid animals. In vitro such isolates
can only infect established cell lines of neuronal origin. However, viruses can be
adapted (Kissling, 1958) and several passages are required for the virus to be adapted
fully to the in vitro multiplication. Additional cycles of multiplication in non- neuronal
cells are necessary for the selection of fixed strains that would multiply in established
Page 33
REVIEW OF LITERATURE
12
cell lines such as BHK21, BSR and Vero cells (Wiktor et al., 1964; Schneider et al.,
1971).
Evelyn Rokitnicky Abelseth (ERA), Pasteur Virus (PV) or Challenge Virus
Standard (CVS) are fixed rabies virus strains that have been selected and are used
around the world for laboratory investigation. All have kept their specific tropism for
neurons in animals and propagate in the nervous system like street viruses. Therefore,
adaptation did not abolish neurotropism but rendered the virus able to grow in non-
neuronal cells. It is postulated but not demonstrated that adaptation is at least partly due
to the capability of fixed strains of rabies virus to use ubiquitous receptors present on
every cell types investigated to date (Seganti et al., 1990). Ubiquitous receptors could
be molecules, such as phospholipids (Superti et al., 1984b), gangliosides (Conti et al.,
1986; Superti et al., 1986), or proteins (Wunner et al., 1984; Broughan and Wunner,
1995; Gastka et al., 1996). The neural cell – adhesion molecule is also shown to be a
receptor for rabies virus laboratory strains (Thoulouze et al., 1998). It has also been
proposed that the nicotinic acetylcholine receptor (nAChR) serves as a receptor for
rabies virus (Lentz et al., 1984; 1986; Hanham et al., 1993). But rabies virus infects
neurons that do not express nAChR (McGehee and Lorca, 1995) which suggests the
existence of other molecules mediating viral entry into neurons.
2.4 Classification and epidemiology
Rabies viruses are group members of the serologically related viruses sharing
common determinants in the RNP group antigen (Schneider et al., 1972). The
lyssavirus genus and the vesiculovirus genus make up the family rhabdoviridae.
Rhabdoviruses are members of the Mononegavirales order, which includes the
Paramyxoviruses and the Filoviruses. The lyssaviruses have been subdivided in four
serotypes on the basis of seroneutralization and monoclonal antibody studies
(Schneider et al., 1973; WHO, 1984; Wiktor and Koprowski, 1980), and into 6
genotypes according to their genomic sequence (Bourhy et al., 1993). Classical rabies
virus strains (serotype 1) and rabies – related viruses: Lagos bat virus (serotype 2),
Mokolo virus (serotype 3), and Durenhage virus (serotype 4). European bat lyssaviruses
(EBL) were proposed initially to constitute serotype 5, but were then subdivided into:
biotypes 1 and 2 (EBL1 and EBL2) respectively to be distinguished finally as two
Page 34
REVIEW OF LITERATURE
13
clearly distinct genotypes (Bourhy et al., 1992). Rabies virus (serotype 1) is present
worldwide except on several protected islands. Rabies – related viruses have a large
geographic distribution in Africa and Europe (King and Crick 1988; Bourhy et al.,
1992). Serotypes and members of the rabies group not only vary from each other on the
basis of serological differences but also show a different lethal potential when injected
into mice.
Epidemiology is the study of the distribution and causes of disease in
populations. Epidemiologists study how many people or animals have a disease, the
outcome of the disease (recovery, death, disability etc.), and the factors that influence
the distribution and outcome of the disease. The epidemiology of rabies addresses
several questions: What animals have rabies and in what regions of the country, how
many people get rabies and in what regions of the country, how many people get rabies
and from what animals, and what are the best strategies for preventing rabies in people
and animals. Epidemiologic information is often presented by statistical data (numbers
or percentages in graphs and on maps) .
In order to successfully combat rabies, a clear understanding of the disease
epidemiology is required. The way in which viruse change and adapt to different host
species and the role, that geographical and host factors play in virus proliferation, are
issues that underlying the understanding of the disease (Nadin–Davis et al., 1993; De
Mattos et al., 1996; Bourhy et al., 1999; Badrane et al., 2001).
In recent years, a useful approach towards elucidating infectious disease
epidemiology has been through molecular sequence analysis of specific parts of the
genomes of virus isolates. For rabies sequences of genes of the nucleoprotein (Smith et
al., 1992; Tordo and Kouknetzoff, 1993), the phosphoprotein (Nadin–Davis et al.,
2000), and the G-L intergenic region (Sacramento et al., 1992) have been useful in
determining molecular epidemiology of the disease in Europe (Bourhy et al., 1999),
America (Smith et al., 1992; Nadin–Davis et al., 1993, 1999), the Middle East (David
et al., 2000) and Africa (Von Teichman et al., 1995 ; Warner et al., 1996 and Sabeta et
al., 2003).
In some developing countries of Africa, Asia and India, canine rabies remains
common. In these areas, dogs account for more than 90% of animal rabies cases, and
these countries account for 95% to 98% of human cases worldwide. Rates may range
Page 35
REVIEW OF LITERATURE
14
from 0.01 to 0.6 deaths per 100,000 people in Latin America to 2 to 18 deaths per
100,000 people in India and Ethiopia. Countries in which canine rabies are controlled,
as in the United States, wildlife accounts for more than 95% of animal rabies cases.
These countries account for less than 5% of human case worldwide. Rates are typically
less than 0.02 deaths per 100,000 (Haupt, 1999).
Australia, previously free of rabies, became an endemic area in 1996, when a
new pteroid lyssavirus was found in flying foxes and other bats, including insectivorous
species (Hanna et al., 2000; Fraser et al., 1996). Rabies is distributed in all countries
except Antartica. The rabies vaccine strains for example, ERA (Evelyn Rokitnicki
Abelseth), HEP (High Egg Passage), CVS (Challenge Virus Standard) and PM
(Pitmann-Moor) belong to the same serotype, share common envelope and
nucleocapsid determinants and, in addition, only share ribonucleoprotein (RNP)
determinants with the Mokola virus (Schneider et al., 1972).
2.5 Structure of virus
2.5.1 Virus particle:
The rhabdoviruses comprise a large group of bullet shaped RNA Viruses.
Rabies-virus particle is a bullet shaped structure of about 75 nm by 200 nm, with a
helical nucleocapsid surrounded by a thin protein-studded membrane. It is classified as
a member of the Rhabdoviridae family by virtue of its ‘bullet-shaped’ structure. The
helical RNP (a complex between the RNA, the viral polymerase, the nucleoprotein and
the phosphoprotein) with 30 to 35 coils is surrounded by a lipoprotein envelope.
Intracellular assembly of the nucleocapsid with the matrix and G proteins are believed
to be critical for envelopment and cellular release of mature virus particles (Mebatsion
et al., 1999).
The virion is surrounded by a lipoprotein membrane (envelope), from which
spike-like projections extend outward except at the flat end of the particle (Tordo and
Poch, 1988; Wunner, 1991). Three monomeric units of the glycoprotein form each
projection (Tordo and Poch, 1988). General structure of rabies virus is shown in Figure
2.1.
Page 36
Figure 2.1: Structure of rabies virus. Rabies virions are bullet shaped
with 10 nm spike like glycoprotein peplomers covering the
surface. The ribo-nucleoprotein is composed of RNA encased in
nucleoprotein, phosphoprotein and polymerase. (A) photograph
are adapted from Centers for Disease Control and Prevention;
(B), Negatively stained rabies virus seen by transmission
electron microscopy from the Wadsworth Center of the New
York State, Department of Health; (C), Diagrammatic
representation of the rabies virion deduced from electron
microscopy and protein analysis by Vernon et al. 1972.
A
B C
Page 37
REVIEW OF LITERATURE
15
2.5.2 Virus genome:
The first complete nucleotide sequence of a rabies virus genome of highly
pathogenic strain PV was published by Tordo et al., (1988). The rabies virus genome
consists of a single-stranded, unsegmented, negative-sense RNA of about 12 kb, which
is transcribed on infection to produce five polyadenylated complementary
monocistronic mRNA species. Each of the virus – specific mRNAs representing a
structural gene of the rabies virus genome (Flamand and Delagneau, 1978) codes for a
virion structural protein (Pennica et al., 1980 and Wunner et al., 1980). The five rabies
virus specific mRNAs for L, G, N, M and P proteins have been characterized by sucrose
gradient (Wunner et al., 1980) and gel analysis (Pennica et al., 1980). These are
contained within a bullet-shaped, bilayered envelope (Tordo and Kouknetzoff, 1993).
All the five proteins are contained within the virion particle. The polymerase (L) and
phosphoprotein (P) complex with the nucleoprotein (N) form an inner capsid of the
virion. The matrix protein (M) and the glycoprotein (G) form the inner and outer layer
of the bilayered envelope (Wunner et al., 1988). The G protein spikes regulate the cell
surface receptors and antibody binding sites and any variation in the gene encoding for
this protein may affect the pathogenic and immunogenic properties of the virus
(Wunner, 1991). The nucleoprotein is highly conserved, essential for viral propagation
and an important target for diagnosis (Dean et al., 1996; Yang et al., 1998).
2.5.3 Virus proteins
2.5.3.1 Nucleoprotein (N)
Nucleoprotein is a phosphorylated (at amino acid residue 389) protein of 450
amino acids (50.5 kDa) which is present in the virion in a stochiometric amount of
approximately 1800. It is tightly associated with the viral RNA protecting the RNA
from ribonucleases, supposedly keeping it in a suitable configuration for transcription
(Sokol and Clark, 1973; Tordo et al., 1986a; Dietzschold et al., 1987a).The N protein is
involved in the regulation of transcription and replication.
2.5.3.2 Phosphoprotein (P)
Phosphoprotein is a phosphorylated protein of 297 amino acids (41 kDa) present
at about 950 molecules per virion. The phosphorylation is probably situated in the
Page 38
REVIEW OF LITERATURE
16
amino terminal half of the protein as in the case of P protein of the vesicular stomatitis
virus -VSV (Weiss and Bennet, 1980). The phosphoprotein of rabies virus is an integral
component of the viral ribonucleoprotein complex. (Chenik et al., 1994) but it does not
participate in the viral structure (Delagneau et al., 1981). It has, therefore, been
denominated “non-structural” protein (NS). P protein is an important cofactor for the
viral polymerase.
2.5.3.3 Matrix protein (M)
About 1500 molecules of the M protein (202 amino acids, 23 kDa) are located
on the inner surface of the viral envelope (Delagneau et al., 1981), where they interact
with both the cytoplasmic domain of the anchored G protein and the virus core, binding
the membrane – associated (M and G) proteins to the RNP, possibly through interaction
with the N protein. M protein also plays a role in virus budding (Weiss and Bennet,
1980).
2.5.3.4 Polymerase (L)
The RNA-dependent RNA polymerase L (for “large”) is the largest (2142
amino acids, 244 kDa) and the less present (25 molecules per virus) rabies virus
protein. It is an enzymatic complex showing various activities: RNA dependent
polymerase, guanylyl transferase and poly (A) synthetase.
2.5.3.5 Glycoprotein (G)
The G protein, which reacts with the host cell receptors, is the main target of the
immune response and it also plays an important role in viral pathogenesis (Dietzschold
et al., 1983a; 1983b). Accordingly, extensive mapping of the functional domains of G
protein has been carried out (Seif et al., 1985; Wunner et al., 1985a; Prehaud et al.,
1988; Tuffereau et al., 1989; Dietzschold et al., 1990a; Benmansour et al., 1991;
Heijden et al., 1993; Ni et al., 1995; Raux et al., 1995). The Glycoprotein is organized
in trimers, which protrude from the viral envelope (Whitt et al., 1991; Gaudin et al.,
1992). It is a type 1 integral transmembrane glycoprotein.
Page 39
REVIEW OF LITERATURE
17
(A) Detail structure of glycoprotein
The glycoprotein gene codes for a membrane associated molecule which forms
spike – like projections on the surface of mature rabies virions. The trimeric G protein
is the only viral protein exposed on the surface of the virion (Whitt et al., 1991). The
consequences during evolution and the adaptation to new hosts, of changes in the
binding of G protein to cellular receptors (Tuffereau et al., 1998a; 1998b; Thoulouze et
al., 1998) are presently unknown. The G protein is the major antigen of the rabies
virus, and an immunological response directed against this protein is able to protect
from rabies virus (Wiktor et al., 1984; Flamand et al., 1993; Bai et al., 1993). It is well
reported that G protein induces virus-neutralizing antibodies (VNA) (Wiktor et al.,
1973) and protection against intracerebral challenge (Perrin et al., 1985a and 1985b).
The G protein also induces the production of cell-mediated immunity (Celis et al.,
1988).
The G protein is a glycosylated protein of 505 amino acid (65 kDa) length. Each
spike consists of a homopolymer of 3 molecules and extends 8.3 nm from the viral
membrane (Delagneau et al., 1981; Gaudin et al., 1992). The G protein accounts for
about 40% of the total mass of viral proteins.
The amino acid sequences of the glycoprotein of the Evelyn Rokitnicki
Abelseth (ERA), Challenge Virus Strandard (CVS), Pasteur Virus (PV) and High Egg
Passage (HEP), and Flury strains have been deduced from the nucleotide sequence
(Anilionis et al., 1981; Yelverton et al., 1983; Tordo et al., 1986b). The absence of the
sequence AAUAAA, present in eukaryotic polyadenylated mRNA (Proudfoot and
Brownlee, 1976), in the 3’ noncoding region of rabies G mRNA and vesicular
stomatitis virus (VSV) mRNAs (Rose, 1980) presumably reflects the role of virus-
associated proteins in polyadenylation of these mRNAs (Herman et al., 1980) in place
of host enzymes.
The G protein carries four potential N-glycosylation sites which are
glycosylated to a different extent according to the virus strain. For example, G protein
of the PV strain is not glycosylated at position 37 but glycosylated at positions 158, 247
and 319 (Wunner et al., 1988). On the G protein of CVS, 2 or 3 of the potential
glycosylation sites are glycosylated, resulting in two different protein sizes (Wunner et
al., 1985b; Whitt et al., 1991). Newly synthesized G protein (524 amino acids) contains
Page 40
REVIEW OF LITERATURE
18
4 domains: (1) the cleavable aminoterminal signal peptide (19 hydrophobic amino
acids) responsible for translocation of the nascent protein across the rough endoplasmic
reticulum (RER) membrane (Vishwanath et al., 1978; Lai and Dietzschold, 1981), (2)
the ectodomain (438 amino acids), as constituent of infection, hemagglutination activity
(Halonen et al., 1968) and immunological responses, (3) the transmembrane domain
(22 hydrophobic amino acids) responsible for the interaction with the membrane lipids,
(4) the cytoplasmic domain (44 amino acids) responsible for the interaction with core
proteins.
(B) Fusion activity
Depending on the pH , G protein can be present in at least three conformational
forms which are in a pH-dependent equilibrium : (1) the “native” ( N ) state detected at
the viral surface above pH 7 , (2) the “activated hydrophobic” state which interacts with
the target membrane as a first step of the fusion process , (3) the “fusion-inactive” ( I )
state which is present at low pH (Perrin and Atanasiu, 1981; Gaudin et al., 1993;
1995a; 1995b; 1996). There is a complex pH-dependent equilibrium between these
states. By using electron microscopy, it has been shown that the “fusion-inactive” (I)
state is 3 nm longer than the “native state” (N), from which it is also antigenically
distinct (Gaudin et al., 1993). It is proposed that G protein is transported through the
golgi apparatus in an I-like conformation to avoid undesirable fusion during its
transport through the acidic golgi vesicles (Gaudin et al., 1995a). Wojczyk et al. (1995)
demonstrated that a soluble form of G protein, constructed by insertion of a stop codon
just before the transmembrane domain, was efficiently expressed and secreted in
transfected CHO cells. The glycoprotein is also responsible for the low pH-induced
fusion of the viral envelope with the endosomal membranes (Gaudin et al., 1993; Whitt
et al., 1991). The pH threshold for fusion is about 6.3 and pre-incubation of the virus
below pH 6.75 in the absence of a target membrane results in inhibition of virus fusion
properties. This inhibition is reversible by re-incubating the virus above pH 7 (Gaudin
et al., 1993). This behaviour, shared with another rhabdovirus, VSV (Clague et al.,
1990; Pak et al., 1997), is different from that observed with other viruses, for which
low pH-induced fusion inactivation is irreversible (Gaudin et al., 1995b). The pH
dependent conformational changes are important for RNP delivery into the cell
Page 41
REVIEW OF LITERATURE
19
cytoplasm. The region of G protein that is crucial for fusion seems to be situated
between amino acid 103 and 179 (Durrer et al., 1995). Mutations at the amino acids
124, 127 or 133 lead to altered fusion activities, though the viral particles remain
infectious (Fredericksen and Whitt, 1996). Cell fusion and formation of syncytia have
also been observed under certain conditions in infected or transfected cells. For
example, glycoprotein transfected neuroblastoma cells but not transfected BHK cells
can form synctia; indicating that some cellular factor is needed for interaction
(Morimoto et al., 1992). Transfected HeLa cells also show fusion activity at low pH
(Whitt et al., 1991).
(C) Synthesis and folding
Many details about rabies G protein synthesis and folding are not yet known,
and one often extrapolates from VSV G protein that shows some similarities (Rose et
al., 1982). Viral glycoprotein shares several functional and structural features with
other members of the group of integral membrane proteins (Doms et al., 1993). They
are important for receptor binding, membrane fusion and penetration, viral
morphogenesis at the budding site and also for stimulation of VNA production. The
majority of these properties are mediated by the ectodomain of glycoprotein.
Glycoprotein is translated by the ribosomes of the rough endoplasmic reticulum
(RER) and is inserted cotranslationally in the ER in an unfolded form. During folding,
the different parts of the proteins are exposed to different conditions: (1) the ecodomain
is situated in the ER lumen and is processed like membrane or secretary proteins, (2)
the transmembrane domain is integrated in the ER membrane, most likely in the form
of an -helix, (3) the cytoplasmic domain is located in the cytosol and is, therefore,
processed like cytoplasmic proteins. In the ER, glycoprotein are glycosylated
cotranslationally by the addition of CA-type (complex-type, high mannose content)
carbohydrates composed of mannose, galactose, fucose, N-acetyl glucosamine and N-
acetyl neuraminic acid (sialic acid). Glycosylation is a multi-step process that involves
addition and removal of sugar residues (Bergmann et al., 1981). N-linked glycosylation
is crucial for appropriate intracellular transport of glycoproteins (Burger et al., 1991;
Wojczyk et al., 1995).
Page 42
REVIEW OF LITERATURE
20
The ER is the cellular compartment that assures folding and oxidation of newly
synthesized proteins. It resembles the extracellular space, but shows a very high Ca++
concentration (Baumann et al., 1991), often important for protein folding as well as a
high oxidation potential for the formation of disulfide bonds (Hwang et al., 1992).
Further important features of the ER are the presence of “Chaperons” and folding
enzymes. Molecular chaperons are ubiquitous proteins that assist polypeptide folding
and assembly by binding of nascent polypeptides. These, therefore prevent both
aggregation and release of incorrect or incomplete folded proteins (Ellis and
Hemmingsen, 1989). After glycosylation the rabies G protein associates with two
chaperons: calnexin and GRP78-Bip (Bip) (Gaudin, 1997). Conformational epitopes
(indicating a correct three-dimensional structure) of G are present only after full
oxidation of the protein. After oligomerization (trimer formation) the protein migrates
to the golgi apparatus (Bergmann et al., 1981), where it acquires resistance to certain
enzyme, such as endo--N acetyl glucosaminidase H (Perrin and Atanasiu, 1980) as a
sign of full maturation. Finally, G protein is transported within acidic vesicles to the
cytoplasmic membrane (presumably in the fusion inactive form; Gaudin et al., 1995b)
and assembled (possibly in interaction with M protein) with the other viral proteins at
the site of budding. Altogether, the folding of rabies G protein shows many similarities
with VSV G protein, even though the folding of rabies G protein is slower than VSV
glycoprotein.
In addition to synthesis of a full length glycoprotein, rabies infected cells secrete
a soluble form of G protein which lacks the 58 carboxy terminal amino acids. Even
though the ectodomain is complete, the soluble G protein does not induce VNA
(Dietzschold et al., 1983a), which reflects incorrect folding of the secreted soluble G
protein.
2.6 Role of viral proteins in the protection against rabies
The most important viral proteins involved in the protection against rabies are G
and N proteins. Antigenic variation of G protein between different genotypes is very
important (Wiktor and Koprowski, 1978; Flamand et al., 1980), whereas N protein
exhibits less variability: isolates that belong to different genotypes present up to 92%
amino acid homology in the N protein (Kissi et al., 1995).
Page 43
REVIEW OF LITERATURE
21
Glycoprotein neutralizing monoclonal antibodies (Mabs) rose in BALB/c mice
immunized with rabies virus were shown to delineate numerous epitopes on G protein
(Flamand et al., 1980; Wiktor and Koprowski, 1980). Their patterns of cross-reactivity
to the antigenic mutants selected with some of these Mabs demonstrated that there were
at least three antigenic sites on the CVS G protein (Lafon et al., 1983) and five on the
ERA G protein (Lafon et al., 1992). It has been reported that most of the Mabs with
virus-neutralizing activity recognized conformational epitopes on G protein (Lafon et
al., 1992; Prehaud et al., 1988).
The antigenic sites of G of the ERA strain have been identified using
monoclonal antibodies (Flamand et al., 1980). Two immunodominant sites are
recognized by the majority of antibodies: site II and site III. Site II is a conformational
epitope, consisting of two parts, IIa (aa 198-200) and Ib (aa 34-42) (Prehaud et al.,
1988), that are joined by disulfide bonds (Dietzschold et al., 1982). Under denaturing
conditions this site is, therefore, not recognized by the corresponding antibody (Lafon
et al., 1985). Site III, is also a conformational site, represented by atleast three epitopes
that consist of amino acids in close relation to each other (330-338 aa). It induces a
fourth more distantly located epitope (aa 357) (Seif et al., 1985; Wunner et al., 1988).
Three minor sites have also been identified: a (aa 342-343), b and c (Prehaud et al.,
1988; Benmansour et al., 1991). Sites b and c contain the former site VI (aa 264)
(Dietzschold et al., 1983b; Bunchoten et al., 1989; Benmansour et al., 1991). The
relative importance of the different epitopes has been estimated by the number of
neutralizing monoclonal antibodies directed against them that have been isolated
(Benmansour et al., 1991).
Glycoprotein is the only rabies protein able to induce VNA (Wiktor et al., 1973;
Cox et al., 1977) and can achieve total protection against an intracerebral challenge
without the presence of other antigenic structures (Cox et al., 1977; Perrin et al., 1985b;
Takita-Sonada et al., 1993). G protein specific monoclonal antibodies that show virus
neutralizing activities in vitro protect against a peripheral challenge in vivo
(Dietzschold et al., 1990b).
The three dimensional structure of G protein and its environment are highly
important for the induction of VNA (Perrin et al., 1988a). Purified G protein forms
aggregates (“rosettes”) that induce much less VNA than correctly folded G protein,
Page 44
REVIEW OF LITERATURE
22
indicating that G protein needs to be anchored on a membrane (viral particle or
liposomes) (Cox et al., 1980; Perrin et al., 1984; Perrin et al., 1985b). Similarly, the
soluble G protein form that lacks the transmembrane region induces 15 times less VNA
than the native G protein (Dietzschold et al., 1983a).
The G protein is recognized both by Th cells (MacFarlan et al., 1984; Xiang et
al., 1995b; Bahloul et al., 1997) and CTL (Wiktor et al., 1984; Celis et al., 1988). The
sequential synthetic peptides deduced from the amino acid sequence of rabies G protein
have been shown to stimulate the Th cell response: aa 1-44, aa 244–323 and 386-452
(MacFarlan et al., 1984). Amino acids 18-44 are also involved in the CTL responses
(MacFarlan et al., 1986). Another CTL epitope is located between aa 130 and aa 178,
containing a single disulfide loop (159-169) (Wunner et al., 1985a; MacFarlan et al.,
1986). Anchored on liposomes, purified G protein induces as much IL-2 production
upon in vitro stimulation of lymphocytes as inactivated virus (Oth et al., 1987; Perrin et
al., 1988a and 1988b).
2.6.1 Antigenicity of glycoprotein
Studies on the antigenicity of the protein have identified two immunodominant
conformational sites named sites II and III (Seif et al., 1985; Prehaud et al., 1988), one
minor site (site a) (Benmansour et al., 1991) and several linear epitopes (Bunschoten et
al., 1989; Raux et al., 1995; Lafay et al., 1996) on the external domain.The G protein is
a major determinant of the viral neurotropism. Mutations in the glycoprotein reduce or
abolish neuroinvasiveness without impairing the ability of the virus to multiply in cell
culture. Replacement of Arg 333, situated in site III of the G Protein, results in the loss
of virulence for adult animals (Dietzschold et al., 1983 b; Seif et al., 1985; Tuffereau et
al., 1989). The mutant virus is still able to infect peripheral neurons but is only
transmitted to a few categories of second order neurons in the CNS (Dietzschold et al.,
1985; Coulon et al., 1989; Lafay et al., 1991).
Antigenic site II has always been considered to be the clearly dominant
antigenic site of the rabies glycoprotein (Benmansour et al., 1991). An antigenic site in
the G gene, most notably site III, appears to be an important determinant of
neuropathogenesis (Dietzschold et al., 1983b; Flamand et al., 1993). Mutations in the G
Page 45
REVIEW OF LITERATURE
23
protein are known to modify the virulence of rabies virus in vitro and in vivo
(Dietzschold et al., 1983b; Seif et al., 1985).
Rabies virus glycoprotein is the only rabies protein able to induce virus
neutralizing antibodies (VNA) (Wilktor et al., 1973) which are crucial element of
protection against rabies (Cox et al., 1977). Almost all antigenic mutations affecting
these epitopes are located on the G protein between amino acid positions 34 and 42,
198 and 204 (Prehaud et al., 1988) or 330 and 340 (Seif et al., 1985). Three linear
epitopes for neutralizing Mabs have been mapped on positions 251 (Lafay et al., 1996),
263 (Ni et al., 1995) and 264 (Dietzschold et al., 1990a). Synthetic peptide around
position 263 or 264 induces the neutralizing antibodies to rabies virus, although some
amino acid substitutions have been observed on this site in certain fixed or street strains
(Conzelmann et al., 1990; Bai et al., 1993; Nadin-Davis et al., 1994; Ni et al., 1995;
Morimoto et al., 1996). Most Mabs against G protein recognize the conformational
epitopes (Lafon et al., 1983; Raux et al., 1995). So far, two linear virus-neutralizing
epitopes have been identified on G protein of CVS (Ni et al., 1995) and on HEP-Flury
strains (Dietzschold et al., 1990a). Although 263-phenylalanine and 264-argnine on G
protein, which have been identified as key amino acids for linear epitopes, are
conserved among numbers of rabies strains, the replacement of 263 by leucine has
been observed in some fixed strains (Bai et al., 1993; Ito et al., 1994; Sakamoto et al.,
1999) and that of 264 by histidine has been observed in some street strains (Bai et al.,
1993 ; Nadin-Davis et al., 1994; Morimoto et al., 1996).
It has been shown that the rabies virus G protein is a major contributor to the
pathogenicity of the virus (Dietzschold et al., 1982; Morimoto et al., 1999; Seif et al.,
1985). Several G protein associated pathogenic mechanisms have been identified, for
example -
(1) G protein must interact effectively with cell surface molecules that can mediate
rapid virus uptake (Dietzschold et al., 1985; Lentz et al., 1987; Thoulouze et al.,
1998; Tuffereau et al., 1998b).
(2) G protein must interact optimally with the RNP-M complex for efficient virus
budding (Mebatsion et al., 1996; 1999; Morimoto et al., 2000).
Page 46
REVIEW OF LITERATURE
24
(3) Expression levels of G protein must be controlled to prevent over – expression,
which causes functional impairment of the infected neuron (Morimoto et al.,
1999).
2.6.2 Antigenicity of nucleoprotein
2.6.2.1 Antigenic structure
Three antigenic sites have been described on the N protein of the ERA strain
(Lafon and Wiktor, 1985). Site I (aa 374-383) and site III (aa 313-337) contain
continuous epitopes, whereas site II is discontinuous (Lafon and Wiktor, 1985;
Dietzschold et al., 1987a). The N protein is also an efficacious Th cell inducer, and a
Th epitope has been identified (aa 404-418) (Ertl et al., 1991).
2.6.2.2 Protective properties and subunit vaccine
Viral RNP and N protein have been shown to induce partial protection against
intramuscular challenge, whereas no protection is observed when the challenge virus is
injected intra-cerebrally. Similar results have been obtained when recombinant N
proteins: purified N protein from recombinant baculovirus (N-Bac) (Fu et al., 1991), or
106-10
8 plaque forming units (PFU) of recombinant pox virus (N-Vacc) (Lodmell et al.,
1991; Sumner et al., 1991; Fekadu et al., 1992) induced partial protection against a
homologous intramuscular challenge. Furthermore, cross – protection after
nucleoprotein injection has been reported: RNP from Mokola and ERA virus can
protect against CVS and Duvenhage challenge respectively (Dietzschold et al., 1987b).
N protein has often been discussed to be an important component in a subunit
vaccine against rabies because of its capacity to induce Th cell and its conservation
between the serotypes. N protein might therefore have a possible role in a vaccine with
a broadened spectrum. However, the RNP complex could not be a part of a subunit
vaccine; it has to be extracted from virus particles or infected cells which is expensive
and carries the risk of a contamination with viral RNA (Schneider et al., 1973).
Combinations of N and G proteins have been tested for their performance in a
subunit vaccine: liposomes containing both G protein and RNP only induced a slightly
higher protection against intracerebral challenge by G protein liposome alone
(Dietzschold et al., 1987a). A tandem peptide carrying a Th epitope of N protein and a
Page 47
REVIEW OF LITERATURE
25
B cell epitope of G protein protected against intramuscular challenge (Dietzschold et
al., 1990a). On the other hand, G-Vacc and N-Vacc injected at different sites did not
raise higher titres of VNA than G-Vacc alone (Fekadu et al., 1992).
Thus it can be concluded that while the importance of G protein in the immune
response against rabies virus infection is clearly established, the role of N protein is
uncertain. The mechanism of the protection induced by the N protein is not yet fully
understood. Though, experiments which show protection by N protein, no VNA have
been detected. The protection must therefore have been achieved by other means, such
as the Th response (Dietzschold et al., 1987b; 1989). N protein also induces non-
neutralizing antibodies that seem to be important for protection: higher anti-RNP
antibody titers are correlated with higher survivor rates in RNP or N protein immunized
mice (Fu et al., 1991). Furthermore, anti- N serum is able to protect under certain
conditions against a weak virus challenge (Lodmell et al., 1993). Protective non-
neutralizing antibodies have as well been reported in the case of other viruses, for
example, VSV (Lefrancois, 1984).
2.7 Classical Rabies vaccines and protection
Since the first rabies vaccine that was developed by Louis Pasteur (Pasteur,
1885), consisting of subcutaneous inoculation of spinal cord suspension derived from
rabid rabbits, different generations of vaccines have been developed which shown in
table 2.2 (Wiktor et al., 1988; Celis et al., 1989; Perrin et al., 1990; Plotkin, 1993).
2.8 Treatment and vaccine
Rabies is one of the most lethal infectious diseases known, being fatal in 100%
of cases if no treatment is administered. According to the World Health Organization
(WHO), an estimated 10 million people receive post–exposure treatment for rabies
every year (WHO, 1992). Despite the existence of effective pre-exposure and post–
exposure treatments, at least 60,000 deaths world-wide occur annually from rabies
(WHO, 1996).
Animals immunized with subunit vaccines based on the rabies virus G protein
are protected against a challenge with rabies virus administered peripherally or directly
into the central nervous system (Wiktor et al., 1984). Neutralizing antibodies are known
Page 48
Table 2.2: Response of different vaccines against rabies
S. No. Generation Source of
preparation
Immunogenicity Human (H)/
Veterinary (V)
1. Whole
inactivated virus
NERVOUS TISSUE
-adult animals
(sheep, goat, rabbit)
poor
H
-new born mice poor H
2. Whole
inactivated virus
PRIMARY CELL EXPLANTS
-embryos (chicken,
duck)
Good
H
-kidney (hamster,
dog, pig)
Good
H
-kidney (bovine
fetus)
Good H
3. Whole
inactivated virus
DIPLOID CELLS
-human (Wi-38,
MRC-5)
Good
H
-rhesus monkey
(fibro blasts)
Good H
HETEROPLOID CELLS
-vero Good V
-BHK-21 Good V
4. Avirulent active
virus
Mutation at aa 333 Very good V
5. Recombinant
active virus
G-recombinant
vaccine
Very good V
Taken from Fu et al., 1997
Page 49
REVIEW OF LITERATURE
26
to prevent the spread of rabies virus into the central nervous system (Shanker et al.,
1991). Rabies glycoprotein is effective as a subunit vaccine (Kieny et al., 1984)
provided that it is glycosylated (Yelverton et al., 1983; Lathe et al., 1984).
2.8.1 Different generations of vaccines
The first generation of rabies vaccines consisted of phenol or heat-inactivated
viruses produced from nervous tissue. In spite of often severe side effects that
contribute to the burden of rabies on public health, these vaccines are still used in
developing countries since they are easy to prepare and less expensive than the more
recent generations of vaccines (Meslin et al., 1994). The following types of vaccines
have been developed in recent years: purified duck embryo cell vaccine (PDEV), dog
kidney cell vaccines, purified chicken embryo cell vaccine (PCEC) and vero cell
vaccine (Sureau, 1992). Within the veterinary field, an additional type of vaccination is
available: the oral vaccination of wildlife (Winkler and Bogel, 1992). The first
vaccination campaigns were carried out using live modified virus vaccines, derived
mainly from the SAD – ERA strain, although they have been shown to maintain certain
pathogenicity against wild rodents (Artois et al., 1992). In some countries these
vaccines were replaced by recombinant vaccinia virus expressing rabies glycoprotein
(G-Vacc) (Brochier et al., 1990). The oral vaccination of wildlife, together with
domestic dogs, has led to a significant reduction of rabies incidence.
2.8.2 Anti –rabies vaccination of humans
Whereas the veterinary use of rabies vaccines is limited to preventive measures,
vaccination of human takes place mainly after exposure to a rabid animal. Different
regimens of vaccination are recommended (WHO, 1996), varying in number and site
(muscle or skin) of injections. In the case of severe exposure, vaccination is often
accompanied by injection of human rabies immunoglobulin (HRIG). However, due to
non-availability and high costs, the HRIG is often replaced by equine immunoglobulins
(ERIG). In the future, monoclonal human antibodies might be an alternative source
(Schumacher, 1989). Serum can only be used in addition to vaccination. On its own, it
does not protect against rabies (Sikes et al., 1971). A major drawback of immune serum
is that it inhibits VNA production after vaccination (Baer et al., 1988). This effect could
Page 50
REVIEW OF LITERATURE
27
be partially overcome, if the number of vaccinations was increased or if the vaccination
took place at least 15 days after the immunoglobulin treatment, or if serums were
administered 12-24 h after vaccination (Wiktor et al., 1971). Patients who had
previously received a complete pre– or post– exposure course, or those who have been
shown at some time in the past to have rabies neutralizing antibodies (>0.5 IU/ml),
need an abbreviated injection schedule without immunoglubulin treatment.
2.8.3 Synthetic peptide vaccines
Synthetic peptide vaccines have several advantages to traditional vaccines: they
are safe, they induce well defined mono functional immune responses and they can be
produced with high reproducibility and exquisite purity in large quantities.
Nevertheless, their immunogenicity is low and the mono specificity of the immune
response allows mutating pathogens to escape from immuno surveillance (Ertl and
Xiang, 1996a). Several approaches of synthetic peptide vaccine have been tested.
1. A peptide carrying an immuno-dominant T cell determinant of the nucleoprotein.
2. A peptide containing a B cell epitope of nucleoprotein.
3. A tandem peptide containing both B-cell and T-cell epitopes of the rabies virus
nucleoprotein (Cruz et al., 2002).
4. A tandem peptide containing a Th cell epitope of N protein and a B cell epitope of
the G protein. Both tandem peptides injected together with adjuvant were able to
induce protection against a peripheral challenge with CVS virus (Dietzschold et al.,
1990a).
2.8.4 Recombinant vaccines
Till date recombinant vaccines against rabies are based either on the injection of
recombinant viruses or of recombinant proteins produced in different expression
systems.
2.8.4.1 Recombinant viruses
Apart from recombinant vaccinia viruses expressing rabies virus G protein
(RGP) that are used for wildlife immunization, recombinant vaccines have so far only
experimental importance. Recombinant vaccinia viruses (pox viruses) have been
Page 51
REVIEW OF LITERATURE
28
successfully used for expression of foreign antigens such as hepatitis B virus surface
antigen (Smith et al., 1983a), herpes simplex virus glycoprotein D (Paoletti et al., 1984)
and influenza hemagglutinin (Smith et al., 1983b). Concerning rabies, all structural
proteins have been expressed in vaccinia viruses (Takita-Sonada et al., 1993).
Particular stress has been laid upon G protein (G-Vacc) (Kieny et al., 1984; Wiktor et
al., 1984; Fujii et al., 1994) and N protein (N-Vacc) (Sumner et al., 1991; Fekadu et al.,
1992).The G-Vacc has been used with much success for wildlife immunization in
Europe.
Other pox viruses have been used for the expression of rabies antigens: racoon
pox virus recombinants expressing N protein (Lodmell et al., 1991) and canarypox
recombinants expressing G protein (Cadoz et al., 1992; Taylor et al., 1995). The
canarypox vector has the considerable advantage not to be replicative in human cells. It
would therefore be more suitable than vaccinia virus that has been forbidden for human
use because of safety risks. The same advantage as with recombinant canarypox is valid
for recombinant adenovirus that expresses rabies G protein (Prevec et al., 1990; Lees et
al., 2002).
2.8.4.2 Recombinant proteins
Rabies G protein (RGP) that produced in Escherichia coli was not immunogenic
(Yelverton, 1983; Lathe et al., 1984) whereas G protein expressed in yeast was able to
protect against an intramuscular but not against an intracerebral virus challenge
(Klepfer, 1993). Other studies showed that G protein molecules were not processed
normally in yeast cell and have abnormal folding and multimer formation. Only a small
fraction was occasionally folded normally into the desired conformational epitopes but
these were mostly deprived of the C-terminal portion (Sakamoto et al., 1999). Other
eukaryotic expression systems are the baculoviruses. Different rabies proteins have
been expressed in the baculovirus expression system, the glycosylation seems to be
different, leading to two glycosylated species of recombinant G protein reported in case
of CVS (Tuchiya et al., 1992; Prehaud et al, 1989) and VSV glycoprotein (Bailey et
al., 1989) or influenza virus heam-agglutinin expressed in insect cells (Kuroda et
al.,1990).
Page 52
REVIEW OF LITERATURE
29
2.8.4.3 DNA based immunization
Recently vaccinations with DNA coding for the G protein have been reported to
trigger protective levels of virus neutralizing antibodies in animal models (Lodmell et
al., 2003; Bahloul et al., 2003). The success of such treatment needs to be
systematically assessed since protective levels of neutralizing antibody in some cases
are not detected for 10-30 days after DNA immunization (Xiang et al., 1995a). By this
time, in most instances, rabies virus would have moved from the bite site, entered
peripheral nerves, invaded the central nervous system and escaped neutralization. DNA
based immunization has been tested for both rabies glycoprotein and nucleoprotein.
Plasmids encoding G induced specific CTL and Th responses as well as VNA
production and protection against intramuscular (Xiang et al., 1994) and intra-cerebral
challenges (Bahloul et al., 1997). The injection of a plasmid coding for N failed to
induce an immune response: the protein is not exported to the cell membrane and might
therefore not be recognized by the immune system of the host (Ertl and Xiang, 1996b).
The advantages of DNA immunization are well known while its risks are still discussed
(WHO, 1992a). Some of the potential risks associated with DNA-based immunization
include the integration of the plasmid DNA-based immunization including integration
of the plasmid DNA into the host genome by homologous recombination. The long
lasting immune response might cause major disadvantage, as it could induce either
tolerance or immuno pathological events. The DNA itself might induce antibodies and
therefore cause auto-immune disease.
2.9 Plants as bioreactors
The development of the first transgenic plants that expressed a mammalian
protein (Lefebvre et al., 1987) and plants with the first vaccine (Curtiss and Cardineau,
1990) that expressed the Streptococcus mutans surface protein antigen A (Spa A) in
tobacco are two major milestones in this area. In the latter study, the transgenic tobacco
tissue incorporated into the diet of mice elicited a mucosal immune response to the Spa
A protein. Although mice were not challenged with the pathogen, the induced
antibodies were biologically active when they reacted with intact S. mutans. Mason et
al. (1992) expressed the hepatitis antigen in tobacco and reported that the transgenic
plants synthesized the hepatitis B coat protein at ≈0.01% of the soluble leaf protein.
Page 53
REVIEW OF LITERATURE
30
Since then, several mammalian genes have been expressed and plants have been
recognized as efficient bioreactors with enormous potential for the production of
proteins valuable to both medicine and industry (Ma and Hein 1995; Goddjin and Pen
1995). The utility of plants as bioreactors was further substantiated when the four
chains of a secretory immunoglobin were properly expressed and assembled in plants,
producing an antibody that was functional (Ma et al., 1995). A bacterial antigen (E. coli
enterotoxin) produced in transgenic plants was shown to effectively immunize mice
when the crude protein extracts from the transgenic plant tissue were administered
orally (Curtiss and Cardineau, 1997; Haq et al., 1995). McGarvey et al., (1995)
expressed the rabies virus glycoprotein in transgenic tomatoes. Plants have also been
used to produce vaccine antigens for human viral diseases such as, Norwalk virus
(Mason et al., 1996). In the late 1980s, plants have been suggested as an emerging
alternative in the vaccine development process. They have been assessed as an
effective, inexpensive, and safe production and delivery system for vaccines. Antigens
produced in plants can be used to generate neutralizing or protective antibodies in
animal models (Mason et al., 1992; 1996; Haq et al., 1995 and McGarvey et al., 1995).
In recent years, several groups have begun to investigate the utility of plants as vehicles
for the expression of vaccine antigens (Hiatt et al., 1989; Ma et al., 1995; Kapusta et
al., 1999; Tackett et al., 1998; 2000).
Recombinant proteins can accumulate in plant tissues. The reported levels of
accumulation range from 0.003% to as high as 14.4% of soluble leaf protein (Kusnadi
et al., 1997). The cost of recombinant protein production in plants has been estimated to
be between 1/10 and 1/50 of that in bacterial fermentation (Kusnadi et al., 1997). Plant
derived vaccines against such animal diseases as mink enteritis virus and rabies were
produced by expressing viral epitopes on the surface of plant viruses, followed by
infection of a susceptible host with the modified virus (Dalsgaard et al., 1997;
Modelska et al., 1998). Arakawa et al. (1997) expressed the cholera toxin B subunit
(CTB) in transgenic potato plants. The maximum amount of CTB protein detected in
auxin induced trangenic potato leaf & tuber tissues was approximately 0.3% of total
soluble plant protein. The first human clinical trials for a transgenic, plant-derived
antigen were planned, approved (US Food and Drug Administration) and performed in
1997 (Tackett et al., 1998). Transgenic potatoes constitutively expressing a synthetic
Page 54
REVIEW OF LITERATURE
31
bacterial diarrhea vaccinogen (the B subunit of E. coli heat labile toxin LT-B) were
orally delivered to human volunteers in phase I/II clinical trials.
Oral administration of disease – specific autoantigens can prevent or delay the
onset of auto immune disease symptoms. Arakawa et al. (1998) generated transgenic
potato plants that synthesized human insulin at levels up to 0.05% of total soluble tuber
protein. The list of plant–derived vaccinogens continues to grow and includes viral,
bacterial, enteric and non-enteric pathogen antigens. In an effort to increase the
expression levels, stability and ease of harvest, strategically designed synthetic genes
have been constructed (Mason et al., 1998). Glycoprotein–B of HCMV (Human
cytomegalovirus), which is a membrane protein requiring complex post translational
processing has been produced in tobacco seeds (Tackaberry et al., 1999).
Recent research has concentrated on meeting the pre-requisites for the
application of plant derived vaccines to the human and animal health industries.
Information on dosage, delivery method and response type has been acquired for
pathogens including Vibrio cholerae (Arakawa et al., 1998), HIV (Durrani et al., 1998),
Pseudomonas aeruginosa (Bernnan et al., 1999), murine hepatitis virus (Koo et al.,
1999) and foot and-mouth disease virus (Wigdorovitz et al., 1999). Further
investigations are targeted at the use of chimeric synthetic genes, targeting of
vaccinogen expression to specific plant tissues, investigation of the induced immune
response and progression to human clinical trials.
In general, however, expression levels of these plant derived bio-
pharmaceuticals need to be increased before commercial production can be
accomplished (Daniell et al., 2001). In the past 12 years, substantial research has shown
that plant-derived vaccines are feasible commodities. Walmsely and Arntzen (2000)
gave a general description of this technology and its early development. The use of
plants to express antigens for application as vaccines has seen continued interest over
the past few years, as witnessed by an increasing number of reports on the expression
of new antigen in transgenic plants. A range of different plant and vector systems for
the expression of antigens have been investigated.
Targeting LT-B to specific sub-cellular locations affects its accumulation. The
LT-B levels are optimal if targeted to the vacuole, and are also high if the antigen is
directed to the cell surface. A signal directing retention of LT-B in the endoplasmic
Page 55
REVIEW OF LITERATURE
32
reticulum favours accumulation of the protein to a greater extent than nuclear or plastid
targets, or in the absence of any target signal resulting in cytoplasmic localization
(Streatfield et al., 2003). In case of the LT-B expressed in tobacco leaves or potato
tubers, the retention in the endoplasmic reticulum promotes accumulation of the protein
three to four-fold over targeting to the cell surface (Haq et al., 1995).
The general eukaryotic protein synthesis pathway is conserved between plants
and animals, so plants can efficiently fold and assemble full-size serum
immunoglobulins as first demonstrated by Hiatt et al. (1989) and secretary IgAs , as
reported by Ma et al .(1995). The post translational modifications carried out by plants
and animals are not identical. There are minor differences in the structure of complex
glycans, such as the presence of the plant specific residues 1, 3- fucose and 1, 2-
xylose. The economic advantages of adapting plants as bioreactors on a larger scale
would reduce the cost of recombinant protein therapy and increase the number of
patients with access to these treatments. The widespread acceptance of plants will be
more likely when key bottlenecks are addressed such as the yield after extraction,
regulatory issues and plant specific glycans. Some of the commercially important
proteins expressed in plants are given in table 2.3.
The development of genetically transformed plant tissue cultures and mainly of
roots transformed by Agrobacterium rhizogenes (hairy roots), is a key step in the use of
in vitro cultures for the production commercially important of secondary metabolites
and proteins. Hairy roots are able to grow fast without phytohormones and produce
much more quantity of metabolite & proteins in comparison with parent plants. The
conditions of transformation (nature and age of the explants, bacterial strain, bacterial
density, and the protocol of infection) deeply influence the frequency of the
transformation events as well as the growth and productivity of the hairy roots. Then
optimization of the culture parameters (medium constituents, elicitation by biotic or
abiotic stress) may enhance the capability of the hairy roots to grow fast and to produce
valuable compounds.
Continuous rhizosecretion of recombinant proteins is another useful and
promising strategy. Root biomass can be significantly increased by hairy root formation
using Agrobacterium rhizogenes. The secreted proteins can be recovered easily from
the hydroponic medium and used as simple source material for protein enrichment and
Page 56
Table 2.3: Proteins of potential commercial interest produced in transgenic plants.
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Tobacco/leaf CaMV35S Streptococcus mutans
bacteraemia and
infective
endocarditis
Surface antigen A Curtiss and Cardineau (1990)
Tobacco/leaf CaMV35S Vesicular stomatitis
virus (VSVG)
Severe acute
respiratory
syndrome
Glycoprotein of VSVG Galbraith et al. (1992)
Tobacco/leaf CaMV35S Hepatitis B virus
(HBV) Hepatitis
Hepatitis B surface
antigen (HBsAg) Mason et al. (1992)
Tomato/leaf,
fruit CaMV35S Rabies virus Rabies RGP McGarvey et al. (1995)
Potato/tuber,
tobacco/leaf CaMV35S E. coli Diarrhea
Heat labile toxin B
subunit (LTB) Haq et al. (1995)
Tobacco/leaf CaMV35S HBV Hepatitis HBsAg Thanavala et al. (1995)
Potato/tuber,
tobacco/leaf
Tuber specific
patatin and
CaMV35S
Norwalk virus (NV) Gastroenteritis Norwalk virus capsid
protein (NVCP) Mason et al. (1996)
Tobacco/leaf CaMV35S V. cholerae Cholera CTB Hein et al. (1996)
Potato/tuber, leaf Mannopine synthase V. cholerae Cholera CTB Arakawa et al. (1997)
Potato/tuber CaMV35S HBV Hepatitis HBsAgM Ehsani et al. (1997)
Potato/leaf, tuber CaMV35S E. coli Diarrhea LTB Mason et al. (1998)
Arabidopsis/leaf CaMV35S
Swine transmissible
gastroenteritis corona
virus (TGEV)
Transmissible
gastroenteritis
(TGE)
Glycoprotein S Gómez et al. (1998)
Page 57
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Potato/tuber, leaf Mannopine synthase V. cholerae Cholera CTB Arakawa et al. (1998a)
Potato/tuber, leaf Mannopine synthase V. cholerae and IDDM Cholera and
Diabetes CTB–INS (Insulin) Arakawa et al. (1998b)
Arabidopsis/leaf CaMV35S Foot and mouth
disease virus (FMDV)
Foot and mouth
disease Structural protein VP1 Carrillo et al. (1998)
Potato/leaf, tuber CaMV35S Rabbit haemorrhagic
disease virus (RHDV)
Rabbit hemorrh-
agic syndrome
Structural protein
VP60 Castañón et al. (1999)
Tobacco/seed Seed specific
glutelin Gt3
Human Cytomegalo
Virus (HCMV)
Central nervous
system disease Glycoprotein B Tackaberry et al. (1999)
Lupin/callus,
lettuce/leaf CaMV35S HBV Hepatitis HBsAg Kapusta et al. (1999)
Tomato/fruit CaMV35S and fruit
specific E8
Respiratory syncytial
virus (RSV)
Serious respiratory
tract disease RSV F protein Sandhu et al. (2000)
Potato/tuber CaMV35S TGEV TGE Glycoprotein S Gómez et al. (2000)
Tobacco/leaf Synthetic super
promoter TGEV TGE Glycoprotein S Tuboly et al. (2000)
Potato/tuber,
potato/leaf
Tuber specific
potatin and
CaMV35S
HBV Hepatitis HBsAg and HBsAg-
VSPαS/VSPαL Richter et al. (2000)
Potato/tuber, leaf CaMV35S FMDV Foot & mouth
disease Structural protein VP1 Carrillo et al. (2001)
Tobacco/leaf
(chloroplast)
Plastid rRNA operon
(Prrn) V. cholerae Cholera CTB Daniell et al. (2001a)
Arabidopsis/leaf CaMV35S Canine parvovirus
(CPV)
Canine parvovirus
disease
VP2 capsid protein of
canine parvovirus Gil et al. (2001)
Page 58
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
White clover/leaf CaMV35S
Mannheimia haemoly-
tica (bovine pneum-
onia pasteurellosis)
Bovine viral
diarrhea Leukotoxin (Lkt) Lee et al. (2001)
Tomato/leaf,
fruit CaMV35S
Organophosphate
poisoning
Severe acute
pancreatitis and
myocardial injury
Human
acetylcholinesterase
(AchE)
Mor et al. (2001)
Potato/ tuber Mannopine synthase V. cholerae, rotavirus
and E.coli
Cholera, diarrhea &
gastroenteritis
CTB–NSP4/CTA2,
CFA/1 Yu and Langridge (2001)
Potato/tuber Tuber-specific
patatin E. coli Diarrhea LTB Lauterslager et al. (2001)
Potato/tuber CaMV35S HBV Hepatitis HBsAg Kong et al. (2001)
Tobacco/leaf CaMV35S Measles virus
(paramyxovirus) Measles
MV-H (Measels virus
hemagglutinin) Huang et al. (2001)
Potato/tuber,
callus Mannopine synthase
V. cholerae and
rotavirus
Cholera and Gast-
roenteritis
CTB–Rotavirus
enterotoxin protein
(NSP4)
Arakawa et al. (2001)
Maize/seed CaMV35S
E.coli and Swine
transmissible gastro-
enteritis corona virus
Diarrhea and Swine
transmissible gast-
roenteritis (TGE)
LTB and TGEV
glycoprotein S
Streatfield et al., 2001;
Lamphear et al., 2002
Tobacco/leaf CaMV35S Measels virus Measles MV-H Webster et al. (2002)
Tomato/fruit,
leaf CaMV35S V. cholerae Cholera CTB Jani et al. (2002)
Alfalfa/leaf CaMV35S FMDV Foot and mouth
disease Structural protein VP1 Dus Santos et al. (2002)
Page 59
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Potato/leaf,
tuber
CaMV35S,
Polyubiquitin,
B33patatin
RHDV Rabbit hemorrhagic
syndrome
Structural protein
VP60 Castañón et al. (2002)
Maize/seed Endosperm specific
gamma zein E. coli Diarrhea LTB Chikwamba et al. (2002)
Tobacco/ NT1,
soybean W82
cell suspension
cultures
CaMV35S HBV Hepatitis HBsAg Smith et al. (2002a; 2002b)
Tobacco/leaf CaMV35 S Enteropathogenic
E.coli (EPEC) Diarrhea
Bundle-forming pilus
structural subunit A
(BfpA)
da Silva et al. (2002)
Potato/ leaf,
tuber CaMV35S
Bovine group A
rotavirus (GAR)
severe viral diarrhea
in humans and
animals
Major capsid protein
VP6 Matsumura et al. (2002)
Tobacco/leaf CaMV35S Bacillus anthracis Anthrax Protective antigen Aziz et al. (2002)
Potato/tuber Mannopine synthase V. cholerae and
rotavirus
Cholera and
gastroenteritis CTB–NSP4 Kim and Langridge (2003)
Tomato/fruit,
leaf CaMV35S E. coli Diarrhea
LTB-Mouse (ZP3)
Zonapellucida 3
epitope
Walmsley et al. (2003)
Potato/tuber CaMV35S Caused by neuro-
degeneration Alzheimer's
Human β-amyloid
(Aβ) Kim et al. (2003)
Tobacco/leaf
(chloroplast)
Plastid rRNA operon
(Prrn) Clostridium tetani Tetanus
Tetanus vaccine
antigen (TetC) Tregoning et al. (2003)
Page 60
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Potato/tuber, leaf CaMV35S Rotavirus Gastroenteritis capsid of rotavirus
glycoprotein VP7 Wu et al. (2003)
Cherry
tomatillo/leaf,
stem, fruit
CaMV35S HBV Hepatitis HBsAg Gao et al. (2003)
Tobacco/NT-1
cell line culture CaMV35S HBV Hepatitis HBsAg-VSPαS Sojikul et al. (2003)
Potato/tuber, leaf CaMV35S
human
papillomaviruses
(HPV)
Cervical cancer HPV11 L1 major
capsid protein Warzecha et al. (2003)
Tobacco/leaf,
potato/tuber, leaf CaMV35S
human
papillomaviruses
(HPV)
Cervical cancer HPV 16 Virus-Like
Particles Biemelt et al. (2003)
Potato/tuber CaMV35S RHDV Rabbit hemorrhagic
syndrome
Structural protein
VP60 Martín-Alonso et zal. (2003)
Carrot/leaf, root CaMV35S Measles virus Measles MV-H Marquet-Blouin et al. (2003)
Peanut/leaf CaMV35S Rinderpest virus
(RPV) Rinderpest
H protein of rinderpest
virus Khandelwal et al. (2003)
Potato/tuber
Tuber specific
patatin and
CaMV35S
HBV Hepatitis HBsAg S and preS2
antigens Joung et al. (2004)
Tobacco/leaf psbA
V. cholerae and
Canine parovirus
(CPV)
Cholera and
Haemo-rrhagic
gastroenteritis and
myocarditis
CTB–2L21 Molina et al. (2004)
Tobacco/Leaf CaMV35S V. cholerae Cholera CTB Jani et al. (2004)
Page 61
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Alfalfa/leaf CaMV35S Bovine rotavirus
(BRV)
Gastroenteritis in
mammals eBRV4 Wigdorovitz et al. (2004)
A.thaliana/ leaf CaMV35S E. coli-M.
tuberculosis, M. Bovis
Diarrhea and
Tubercu-losis
LTB-Early secretory
antigenic
target6(ESAT6)
Rigano et al. (2004)
Tobacco/leaf CaMV35S Measels virus Measels MV-H Webster et al. (2005)
Tobacco/leaf and
tomato/leaf, fruit CaMV35S HBV
Hepatitis and
Gastroenteritis HBsAgM/S and NVCP Huang et al. (2005)
Tobacco/leaf CaMV35S Rabies virus Rabies RGP Ashraf et al. (2005)
Tobacco/Leaf CaMV35S E. coli Diarrhea LTB-SEKDEL Kang et al. (2005)
Alfalfa/leaf CaMV35S Rotavirus Viral gastroenteritis PBsVP6 human group
A rotavirus Dong et al. (2005)
Potato/tuber CaMV35S Porcine epidemic
diarrhea virus (PEDV) Diarrhea
Neutralizing epitope of
PEDV (COE) Kim et al. (2005)
Banana/fruit, leaf ubq3 and EFE HBV Hepatitis HBsAg Kumar et al. (2005a)
Tobacco/cell line
suspension
culture
ubq3 and EFE HBV Hepatitis HBsAg Kumar et al. (2005b)
Collard/leaf,
cauliflower/floret
of mature curd
CaMV35S and
synthetic OCS3MAS.
Vaccinia virus, human
SARS coronavirus
Smallpox and
human SARS
vaccinia virus B5 coat
protein and
coronavirus spike
glycoprotein
Pogrebnyak et al. (2006)
Potato/tuber,
hairy root EFE HBV Hepatitis HBsAg Kumar et al. (2006)
Tomato/fruit CaMV35S HBV Hepatitis HBsAg-ENV,GAG
epitopes of HIV-1 Shchelkunov et al. (2006)
Page 62
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Peanut/callus CaMV35S Bluetongue virus
(BTV) Bluetongue VP2 outer capsid Athmaram et al. (2006)
Potato/tuber, leaf CaMV35S Rotavirus Gastroenteritis capsid of rotavirus
glycoprotein VP7 Li et al. (2006a)
Tobacco/Leaf CaMV35S V. cholerae Cholera CTB–InsB3 Li et al. (2006b)
Tobacco/leaf,
lettuce/leaf CaMV 35S
Severe Acute
Respiratory Syndrome
Coronavirus
Severe Acute
Respiratory
Syndrome
Partial spike (S)
protein of SARS-CoV Li et al. (2006c)
Lettuce/leaf CaMV35S V. cholerae Cholera CTB–SEKDEL Kim et al. (2006)
Tobacco/Leaf CaMV35S V. cholerae Cholera CTB Mishra et al. (2006)
Tobacco/leaf CaMV35S Porcine epidemic
diarrhea virus (PEDV) Diarrhea
Neutralizing epitope of
PEDV (CO-26K) Kang et al. (2006a)
Siberian ginseng/
somatic embryos
CaMV35S and
Ubiquitin E. coli Diarrhea LTB Kang et al. (2006b)
Tomato/fruit CaMV35S Yersinia pestis Pneumonic/bubonic
plague
Antiphagocytic
capsular envelope
glycoprotein (F1) and
low calcium response
virulent antigen (V)
fusion
Alvarez et al. (2006)
Tobacco,
collard/leaf rbcS and CaMV35S Vaccinia virus Smallpox
Vaccinia virus B5 coat
protein Golovkin et al. (2007)
Tomato/fruit Fruit specific E8 V. cholerae Cholera CTB Jiang et al. (2007)
Tomato/fruit Fruit specific E8 Human immuno-
deficiency virus (HIV) AIDS Tat protein of HIV-1 Ramírez et al. (2007)
Page 63
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Papaya/ Embryo-
genic clones
(ETgpC)
CaMV35S Taenia solium Cysticercosis
Synthetic peptides
KETc1, KETc12,
KETc7
Hernández et al. (2007)
Tobacco and
Arabidopsis/leaf CaMV35S
HIV-1 and hepatitis B
virus (HBV)
AIDS and Hepatitis
B
Recombinant HIV-
1/HBV virus-like
particles
Greco et al. (2007)
Rice/seed Endosperm specific
GluB-1 V. cholerae Cholera CTB Nochi et al. (2007)
Soybean/ seed Seed specific
glycinin E. coli Diarrhea LTB Moravec et al. (2007)
Potato/tuber CaMV35S HBV Hepatitis HBsAgM Youma et al. (2007)
Tomato/fruit Fruit specific 2A11 HBV Hepatitis PRS-S1S2S Lou et al. (2007)
Rice/leaves,
seeds
ubiquitin and seed
specific glutelin
Newcastle disease
virus (NDV)
Newcastle disease
(ND)
NDV envelope fusion
(F) glycoprotein Yang et al. (2007)
A. thaliana/leaf,
tobacco/leaf CaMV35S HPV Cervical cancer
HPV11 L1 major
capsid protein Kohl et al. (2007)
Tomato/leaf,
fruit CaMV35S
C. diphteriae, B.
Pertussis and
Clostridium tetani
Diphtheria,
Pertussis and
Tetanus (DPT)
Epitopes of the C.
diphtheriae, B.
pertussis and C. tetani
exotoxins
Soria-Guerra et al. (2007)
Carrot/ leaf, root CaMV35S E. coli Diarrhea LTB Rosales-Mendoza et al. (2008)
Tomato/leaf CaMV35S Caused by neuro-
degeneration Alzheimer's
Human β-amyloid
(Aβ) Youma et al. (2008)
Page 64
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Rice/seeds Seed specific
glutelin A
Caused by
inflammation of joint
cartilage
Arthritis Type II collagen
peptide Hashizume et al. (2008)
Tomato/leaf,
fruit CaMV35S Rabies virus Rabies
Rabies nucleoprotein
(RNP) Arango et al. (2008)
Tobacco/leaf psbA Yersinia pestis plague F1-V fusion protein Arlen et al. (2008)
Peanut/seed Seed specific
legumin V. cholera and Rabies Cholera and Rabies
CTB–Rabies
glycoprotein (RGP) Tiwari (2009)
Tomato/fruit and
tobacco/leaf,
seed
Fruit specific E8 and
CaMV35S
V. cholera and
Hepatitis B virus
(HBV)
Cholera and
Hepatitis B CTB and HBsAg He et al. (2008)
Rice/seed Glub-4 HBV Hepatitis B
SS1(HBsAg)-
hepatocyte receptor-
binding presurface 1
(preS1)
Quian et al.(2008)
Tobacco/leaf CaMV35S HIV-1 AIDS Pr55Gag Meyers et al. (2008)
Tomato/fruit CaMV35S/E2L V. cholerae Cholera CTB-P4/CTB-P6/
TCPA Sharma et al. (2008)
Rice/seed Endosperm specific
Bx17 HMW V. cholerae Cholera CTB Oszvald et al. (2008)
Tobacco/leaf CaMV35S Avian influenza virus
H5/HA1 variant Avian flu
H5/HA1 variant-
HDEL
Spitsin et al. (2009)
Page 65
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Tobacco/
chloroplast
16S rRNA promoter
(Prrn)
Enterotoxigenic E.coli
(ETEC) strains Diarrhea
LTB—Heat stable
toxin (ST) Rosales-Mendoza et al. (2009)
Tobacco/
chloroplast PrbcS/ TrbcS5’-UTR HIV-1 AIDS Pr55(gag) Scotti et al. (2009)
Tobacco/
chloroplast Prrn/ psbA5’-UTR HPV-16 Cervical Cancer PVXCP-E7(16) Morgenfeld et al. (2009)
Lettuce/leaf CaMV35S/NtADH5’-
UTR
E. coli-O138, O139,
and O141 Edema disease
Shiga toxin 2e
(Stx2eB-HDEL) Matsui et al. (2009)
Rice/seed GluB-1 Ascaris suum Gastrointestinal parasite
CTB-AS16 Matsumoto et al. (2009)
Carrot/ root CaMV35S Helicobacter pylori
Chronic gastritis,
peptic ulcer, gastric
cancer and mucosal
lymphoma
UreB (urease B) Zhang et al. (2010)
Tobacco/leaves CaMV35S M. tuberculosis Tuberculosis 85B-ESAT-
6ELPKDEL Floss et al. (2010)
A.thhalian/seed Phaseolin promoter P. falciparum Malaria MSP1 Lau et al. (2010)
Lettuce/leaf CaMV35S Yersinia pestis Bubonic and
pneumonic plague F1-V fusion antigen Rosales-Mendoza et al. (2010)
Arabidopsis/leaf CaMV35S Helicobacter pylori Gastritis, peptic
ulcer, gastric cancer
TonB (HP1341)-
SEKDEL Kalbina et al. (2010)
Tobacco/leaf CaMV35S Rota virus (goupA) Paediatric
gastroenteritis RV VLPs(VP 2/6/7) Yang et al. (2011)
Tobacco/
chloroplast Prm HIV-1 AIDS P24-nef
Gonzalez-Rabade et al.
(2011)
Page 66
Plant/Tissue Promoter Pathogen Disease Antigenic protein Reference
Tobacco/
chloroplast Prm HPV-16 Cervical Cancer HPV-16 L1-LTB Waheed et al. (2011)
N. benthamiana/
leaf Prm-PR1a P. falciparum Malaria Pfs230CMB-KDEL Farrance et al.(2011)
A. thaliana/leaf,
D. carota/root CaMV35S
Chlamydia
trachomatis
Sexually transm-
itted infection (STI) MOMP Kalbina et al. (2011)
Maize/corn CaMV35S A. pleuropneumoniae Porcine
pleuropneumonia (CTB)-ApxIIA Shin et al. (2011)
N. benthamiana/
leaf CaMV35S H1N1, H5N1 Influenza HAC-1, HA1-05 Shoji et al. (2011)
Tobacco/
chloroplast Prm HIV-1 AIDS C4V3 Rubio-Infante et al. (2012)
Tobacco/ leaf CaMV35S Avian influenza A
(H5N1) Influenza Haemagglutinin (H5) Mortimer et al.(2012)
Tobacco/ leaf CaMV35S Bovine viral diarrhea
virus (BVDV) Diarrhea
Glycoprotein E2-
KDEL Nelson et al. (2012)
N. benthamiana/
leaf CaMV35S H1N1 Influenza HAC1 Jul-Larsen et al. (2012)
Tobacco/ leaf CaMV35S GA733 Colorectal
carcinomas GA733-Fc-KDEL Lu et al. (2012)
Page 67
REVIEW OF LITERATURE
33
purification (Komarnytsky et al., 2004). Potato hairy roots were utilized for the
expression of HBsAg (Richter et al., 2000; Kumar et al., 2006). These offer several
advantages for the production of HBsAg, including the availability of efficient genetic
transformation, short regeneration time, availability of tissue specific promoters and
genetic stability. The rhizosecretion has also been exploited recently for heterologous
expression of human alkaline phosphatase (Gaume et al., 2003) and IgG antibodies
(Komarnytsky et al., 2006).
Use of hairy root technology for the production of secondary metabolites such
as gossypol (Verma et al., 2009), isoflavonoid (Udomsuk et al., 2009) and withanolides
(Mirjalili et al., 2009) and important pharmaceuticals recombinant proteins like human
interferon alpha-2b (Luchakivskaia et al. 2012), P450 (Banerjee et al., 2002), mouse IL-
12 (Liu et al., 2009) have successfully established the utility of this system.
2.9.1 Oral Vaccines
Various plant biotechnological techniques, such as, modern breeding methods,
clonal propagation, cell suspension culture, hairy root culture and genetic
transformation can play a vital role in establishing the use of plants as “surrogate
production organisms”. One or more immuno-protective antigens of pathogens can be
produced in plants by the expression of gene(s) encoding the protein(s). In recent years,
plant-based novel production systems aimed at developing edible or oral vaccines have
also been discussed (Ma et al., 2003, 2005; Koprowski, 2005; Lal et al., 2007; Mishra
et al., 2008; Houdebine et al., 2009).
Oral vaccines are more affordable and accessible to the inhabitants of
developing countries, who needlessly die, in the thousands, from diseases, which can
easily be prevented by vaccination. Food vaccines are like subunit preparations in that
they are engineered to contain antigens but bear no genes that would enable whole
pathogens to form. These vaccines basically work in the same way as the injected DNA
vaccine, since a peptide sequence similar to an infectious part of a pathogen is
synthesized, by itself, and is used to prime T and B cells in the body. The big difference
in this case is that the protein sequences are encoded in a plant to form the desired
protein. This protein is then ingested, as the plant or its fruit is eaten. One becomes
immune against the ingested protein, as T and B cells become stimulated to proliferate
Page 68
REVIEW OF LITERATURE
34
and differentiate. Successful expression of antigens in plants was achieved for Rabies
virus G-protein in tomato (McGarvey et al., 1995), Norwalk virus capsid protein in
tobacco and potato (Mason et al., 1996), Hepatitis B virus surface antigen in tobacco
and potato (Thanavala et al., 1995), E. coli heat-labile enterotoxin B subunit (LTB) in
tobacco & potato (Haq et al., 1995), Cholera toxin B subunit (CT-B) in potato
(Arakawa, 1997). Food vaccines are also used to suppress autoimmune disorders like
type-1 diabetes, multiple sclerosis, rheumatoid arthritis etc. Foods under study include
potatoes, bananas, lettuce, rice, wheat, soybean, corn and legumes.
There are various lectins or lectin-like proteins which have binding ability to
glyco-lipids or glycoproteins (De Aizpurua and Russell-Jones, 1988) which mediate the
internalization of therapeutically important proteins through gut epidermis. Many of
these proteins have already characterized and used as mucosal adjuvant which
stimulates strong humoral as well as cell-mediated immune responses. Cholera toxin B
subunit is one which well-studied and provides N- terminal fusion capability only.
While ricin toxin B subunit can provides N-terminal as well as C-terminal fusion
capability with their target antigens. Ricin is the toxic lectin which isolated from the
castor bean plant Ricinus communis, is a 65 kDa glycoprotein containing an A chain
(RTA) and a B chain (RTB) linked by a disulphide bond. Ricin is extremely toxic to
eukaryotic cells. Since Lin et al. reported that ricin showed anti-tumour activity, the
biochemical and biological properties of this protein have been extensively studied by
several investigators. The toxin is composed of two subunits linked together by a single
disulphide bond. The A chain (M, = 30,600) has been shown catalytically to inactivate
the 60S ribosomal subunit such that it has a greatly reduced interaction with the
elongation factors. The B chain (M, = 31,400) is a lectin with a high affinity for
galactose (Vitetta and Yen, 1990; Sphyris et al., 1995; Swimmer et al., 1992; Frankel et
al., 1996; Steeves et al., 1999). It is also known to bind cell surface receptors from
about 1×107
to 3×107 molecules/ cell (Sandvig et al., 1976) presumably via
oligosaccharide recognition, and to thereby initiate into the cell RTB is galactose
specific lectin that mediates binding of the toxin to the surface of mammalian cells.
After binding and internalization of ricin, RTA translocates across intracellular vesicle
membranes to the cytosol, where it catalytically inactivates 60S ribosomal subunits and
thereby inhibits protein synthesis and causes cell death. Mature ricin consists of a
Page 69
REVIEW OF LITERATURE
35
ribosome-inactivating A-chain (RTA) linked by a disulphide bond and non-covalent
interactions to a galactose binding B-chain (RTB). This heterodimer is toxic to
mammalian cells because it can bind via RTB to a variety of galactosylated cell surface
molecules and following retrograde transport to the endoplasmic reticulum (ER). RTB
binds to glycoproteins, which is found on epidermal cell membranes.
The degradation of protein components in the stomach (due to low pH and
gastric enzymes) and gut before eliciting immune response is a main concern of oral
vaccination (Daniell et al., 2001), but the rigid plant cell walls could provide protection
from intestinal degradation (Webster et al., 2002). The degradation can be compensated
by repeating the exposure of the antigen until immunological tolerance is accomplished
(Mason et al., 1995). The M cells lining the small intestine take in the components that
have entered the small intestine (including pathogens) and pass them to other cells of
the immune system, such as antigen presenting cells and macrophages. These cells
chop up their acquisitions and display the resulting protein fragments on the cell
surface. Helper T lymphocytes recognize the displayed fragments as foreign, induce B
lymphocytes to secrete neutralizing antibodies and also help to initiate a broader attack
on the perceived enemy. Mucosal immune responses represent a first line of defence
against most pathogens. Second generation edible vaccines are also called as multi
component vaccines that provide protection against several pathogens. An elegant
approach to achieve this goal, based on epitope fusion to both subunits of the cholera
toxin (CT), was recently demonstrated by Yu and Langridge (2001). CT provides a
scaffold for presentation of protective epitopes of rotavirus and ETEC (Entero-
toxigenic, E. coli), acts as a vaccine candidate by its own right and as a mucosal
adjuvant devoid of toxicity. The trivalent edible vaccine elicited significant humoral
responses, as well as memory B cells and T helper cell responses, important hallmarks
of successful immunization. In the clinical trials described 100 g of raw potato tubers
expressing LTB of E. coli in three doses had to be consumed in order to overcome
digestive losses of the antigen and to elicit a significant immune response (Tackett et
al., 1998).
Traditional production systems that use microbial, insect, mammalian cell
culture and transgenic animals have drawbacks in terms of cost, scalability, product
safety and authenticity. Molecular farming in plants has many practical, economic and
Page 70
REVIEW OF LITERATURE
36
safety advantages as compared to more conventional systems, so the use of plants for
large-scale protein synthesis is gaining wider acceptance. The first pharmaceutically
relevant protein made in plants was human growth hormone expressed in transgenic
tobacco in 1986. Since then many proteins including viral (Norwalk virus capsid
protein), bacterial (Pseudomonas aeruginosa outer membrane protein- Gilleland et al.,
2000), enteric (E. coli heat-labile enterotoxin- Haq et al., 1995) and non-enteric
antigens (Hepatitis B surface antigen- Thanavala et al., 1995) as well as autoimmune
antigens have been produced in an increasingly diverse range of crops. In 1989, the first
antibody was expressed in tobacco, which showed that plants could assemble complex
functional glycoproteins with several subunits. The structural authenticity of plant
derived recombinant protein was confirmed in 1992, when correct assembly of the
Hepatitis B virus surface antigen was reported in tobacco. Further work showed that
plant expressed Hepatitis B particles induced expected immune response in mice.
Generally these recombinant proteins are produced at very low levels in plants,
typically less than 0.1% of total soluble proteins. Besides the factors influencing
transcription and translation efficiency, the accumulation of recombinant protein as
well as their stability strongly depends on the compartments of the plant cell chosen for
expression. All plant production systems can perform a variety of post-translational
modifications, which are similar to those accomplished in mammalian cells. Such
modifications are of special importance for proper function and stability of recombinant
proteins.
The techniques to enhance antigen accumulation in plant tissues are being
explored and include optimization of the coding sequence of bacterial or viral genes for
expression as plant nuclear genes, and defining the sub cellular compartment in which
to accumulate the product for optimal quantity and quality. Despite many recent
advances, low concentrations of heterologous proteins are accumulating in plants.
Several laboratories are also developing alternative expression systems to improve
accumulation. Though, the optimization of expression cassettes is one of the major
ways to make in planta production and purification is economically more feasible. The
regulatory sequence like 5’UTR of the seed storage protein gene arcelin5-I (arc5-I) of
common bean along with β-phaseolin promoter was evaluated in Arabidopsis thaliana.
Nuclear transformations of this construct boosting the heterologous expression up to
Page 71
REVIEW OF LITERATURE
37
36.5% of total soluble protein (TSP) in seeds of homozygous plant (Jaeger et al., 2002).
It indicates the importance of UTR and other signal sequences in expression cassette.
Plants offer a good alternative to develop technologies related to large-scale
manufacture of biologically active proteins. The knowledge of plant using as bioreactor
to produce economically important therapeutics is long standing which successfully
commercially exploited and use to produce several examples. In our laboratory, gene of
rabies glycoprotein was first optimized to clone against CaMV35S in plant expression
vector and transformed it into Nicotiana tabaccum. Glycosylated rabies glycoprotein
not only expressed at the level of 0.38% of TSP but also immuno-protective in the
nature (Ashraf et al., 2005). However, localization of glycoprotein is plays an important
and ER localized protein shows more immuno-protection than non-ER localized protein
(Yadav et al., 2012). But the low level of expression limits the extent of immune
response that leads the development of an effective plant-based oral vaccine. Strategies
like co-administration with an adjuvant (Wang et al., 2005) and fusion of antigens with
an effective carrier molecule, either chemically or genetically, can increase the
immunogenicity of antigens (Kanq et al., 2006; Woffenden et al., 2008). We are not
only successfully employed strategies like N- or C- terminal genetically fusion of
adjuvant to rabies glycoprotein but also scale up to them to perform animal experiment
for fulfillment of commercialization aspect. Compared to traditional vaccines, edible
vaccines offer simplicity of use, lower cost, convenient storage, economic delivery and
mucosal immune response.
This work aims to examine the efficacy of plant based oral vaccine against
rabies which expressed in different part of the plants as well as explore its scalability
option so that it could provide economically viable potions which in turn solve the
problem of needle-free vaccination for animals and humans.
Page 72
REVIEW OF LITERATURE
38
2.10 Objectives of this study
In lieu of the available information, the present study was undertaken with the
following objectives:
1. Construction of N-terminal fusion of CTB to RGP to form CTB-RGP fusion
gene.
2. Sub-cloning of CTB-RGP fusion gene in plant expression vectors.
3. Isolation and cloning of RTB subunit from Ricinus communis.
4. Construction of C-terminal fusion of RTB and RGP to form RGP-RTB fusion
gene.
5. Sub-cloning of RGP-RTB fusion gene in plant expression vectors.
6. Agrobacterium mediated genetic transformation in plant tissues of Nicotiana
tabaccum and Solanum lycopersicum with the above gene constructs.
7. Molecular screening and analysis of transgenic expressing CTB-RGP and RGP-
RTB fusion proteins.
8. Evaluation of antigenic and immunogenic property of CTB- RGP and RGP-
RTB fusion proteins.
Page 73
Chapter 3
Materials and Methods
Page 74
Materials and Methods
39
3.0 MATERIALS AND METHODS
3.1 Construction of fusion gene and their cloning into plant expression cassette
3.1.1 Construction of ctxB & rgp fusion gene and their cloning into plant
expression vector
We have earlier reported designing and cloning of plant codon optimized
synthetic ctxB gene of Vibrio cholerae O139 strain 1854 (Mishra et al, 2006) and rgp
gene of rabies virus glycoprotein (Ashraf et al., 2005) and their expression in tobacco
leaves. In this study, a glycine-proline hinge was used at the point of fusion of
translational frames of the CTB and RGP proteins. The signal sequence PR-S, of the
pathogenesis induced tobacco protein PR-1a was used to facilitate transport of the
fusion protein to endoplasmic reticulum (Sijmons et al., 1990). The pr-s-ctxB was PCR
amplified using a forward primer (5’ACTCTAGAATGAACTTCCTCAAGTCCTT
C3’) with XbaI and a reverse primer (5’AGGCCCGGGACCGTTAGCCATGGAGAT
AG 3’) with SmaI site using pSM31 (Mishra et al, 2006) as the template DNA. The
reverse primer was de-signed to include codons for the glycine-proline hinge at the
3’end of ctxB. The synthetic G protein gene was PCR amplified using pSA5 (Ashraf et
al., 2005) as template DNA and a forward primer (5’GGTCCCGGGCCTAAGTTCCC
TATCTACAC3’) with SmaI site including codons for the glycine–proline of the hinge
region. The reverse primer (5’ACGAGCTCTCATCACAACTCATCCTTCTC3’)
carried a SacI site. Amplified PCR fragments were digested with the respective
enzymes and triply ligated in the vector, pBI101 containing the enhanced CaMV35S
promoter to obtain pSR1241 with two glycine-proline repeats as hinge at the 3’end of
ctxB (Figure 3.1A).
3.1.2 Construction of rgp & rtxB fusion gene and their cloning into plant
expression vector
The rgp gene was amplified with Phusion polymerase (Finzyme, England) from
pSA33 (Ashraf et al., 2005) by using RGP F1 forward and RGP R1, reverse primers to
amplified fragment of ~1.5kb. ER targeting signal of calreticuline of tobacco and
glycine-proline (GP)2 hinge region were added into the amplified rgp fragment as
primer extension method by using Cal F1, Cal F2 , Cal F3 forward and RGP R1
reverse primers. The gene of rtxB was intron-less and directly amplified from the
Page 75
Figure 3.1:(A) Gene constructs showing cloning of the fusion gene ctxB-rgp in
pBI101. The pr-s-ctxB and rgp fragments were PCR amplified
from pSM31 and pSA5, respectively. Amplified PCR fragments
were digested with enzymes and triple ligated in pBI101 to
obtain pSR1241with two glycine-proline repeats as hinge.
(B) Gene construct pAS1 showing cloning of the fusion gene rgp-
rtxB in pCAMBIA1300. Both cal-s-rgp and shrgp-(gp)2-rtxB
fragments were amplified with primer extension method and
cloned in pSK+ Bluescript vector. Both fragments were cut with
PstI, HpaI and HpaI, SalI restriction enzymes, respectively,
then ligated to assemble whole cal-s-rgp-(gp)2-rtxB fusion gene.
pSR1241 pCaMV35S
HindIII SacI BglII
SEKDEL
pr-s rgp
2040bp
CtxB
H-(GP)2
ApaI
RB
Tnos nptII
LB
Pnos
Tnos
XbaI XhoI AgeI
(A)
pAS1
RB
Tnos pCaMV35S
HindIII SacI
SalI PstI
Tnos G-Protein
SEKDEL
cal-s rgp hptII
LB
2241bp
rtxB
H-(GP)2
EcoRI
pCaMV35S
HpaI
(B)
Page 76
Materials and Methods
40
genomic DNA of Ricinus communis. Forward primers Ricin F1 which contains the
sequences of glycine-proline hinge region and reverse primer Ricin R1 and Ricin R2
which contains the SEKDEL sequences were used to extent the rtxB gene at N and C
terminal respectively. The fragment cal-rgp-gp and gp-rtxB-SEKDL was joined by
overlapping assembly PCR method as describe by Shevchuk et al., (2004) and Young
& Dong, (2004). The assembled cal-rgp-gp-rtxB-SEKDL gene was finally amplified
with terminal Cal F1forward and Ricin R2 reverse primer which contains restriction
site PstI and SacI respectively. cycling parameters for amplification of whole gene from
phusion polymerase was used: 98°C for 2 min; 35 cycles of 98°C for 20 sec; 60°C for
15 sec, 72°C for 30 sec; 72°C for 5 min. After amplification of whole gene, PCR
product was cleaned with PCR cleanup kit (Sigma chemicals) and ligated into EcoRV
digested SK+ Bluescript for the sequencing and verification of proper assembly of
whole gene. After sequencing, whole gene was sub-cloned into pCAMBIA1300 at PstI
and SacI restriction sites. CaMV35S double enhancer promoter was amplified from
pCAMBIA1300 with CaMV1F forward and CaMV1R reverse primer containing
HindIII and PstI sites respectively. The amplified product was sub-cloned into same
pCAMBIA1300 which have assembled gene, as describe above into the HindIII and
PstI restriction sites. Finally, Tnos fragment was isolated by digesting with SacI and
EcoRI restriction enzymes from pBI121 and cloned into the same pCAMBIA1300
which contain CaMV35S promoter and whole gene to obtain pAS1(Figure 3.1B). List
of all primers used in above amplifications are given in table 3.1.
3.2 Transformation of constructs
3.2.1 Generation of transgenic tobacco lines
Agrobacterium tumefaciens LBA4404 was transformed with pSR1241 and
pAS1 by electroporation and used for tobacco (Nicotiana tabaccum cv. Petit Havana)
transformation using the leaf disc method (Horsch et al., 1985). A single isolated
colony of A. tumefaciens LBA 4404 (pAL4404) harbouring binary vector pSR1241 and
pAS1 were inoculated in YEP medium containing antibiotics streptomycin, rifampicin,
kanamycin and hygromycine (for pCAMBIA 13000) and grown (200 rpm, overnight,
28oC). Fifty l of the overnight culture was diluted to 100 ml in YEP medium and
grown till OD600 reached to 0.8. Cells were recovered by centrifugation in SS34 rotor
(5,000 rpm, 10 min, 4oC). The pellet was suspended in co-cultivation medium [MS
Page 77
Table 3.1: List of primers used in rgp-rtxB fusion gene construction
S.
No
Primer
Name 5’ to 3’ Sequence
Nt.
Length
1. RGP F1
TCTCTCTGCTCGTCGCTGTCGTCTCCGCTAAGTT
CCCTATCTACACTATC 50
2. Cal F1
ACTGCAGATGGCTACTCAACGAAGGGCAAACC 33
3. Cal F2
CTACTCAACGAAGGGCAAACCCATCTTCTCTTC
ACCTAATTACTG 45
4. Cal F3
CATCTTCTCTTCACCTAATTACTGTATTCTCTCT
GCTCGTCGCTGTC 47
5. RGP R1
TGGCCCTGGCCCCTTACCCCAGTTTGGGAGA 31
6. Ricin F1
GTAAGGGGCCAGGGCCAGCTGATGTTTGTATGG
ATCCT 38
7. Ricin R1
CATCCTTCTCGGAAAATAATGGTAACCATATTT
GGTTTG 39
8. Ricin R2
GCTCTAGATCATCACAACTCATCCTTCTCGGAA
AATAATG 40
9.
CaMV1
F
ATTTACTGAATTCGCGTATTGGCTAGAGCAGCT
TGCCAACATGGTG 46
10.
CaMV1
R
TTCTGCAGAGAGATAGATTTGTAGAGAGAGAC 32
11. Rol B F
ACTATAGCAAACCCCTCCTGC 21
12. Rol B R
TTCAGGTTTACTGCAGCAGGC 21
13. vir C F
ATCATTTGTAGCGACT 16
14. vir C R
AGCTCAAACCTGCTTC 16
Page 78
Materials and Methods
41
salts, 2% glucose, 10 mM MES and 100 mM acetosyringone (3, 5-
Dimethoxyacetophenone) pH 5.6] to OD600 0.6. Tobacco leaf discs were co-cultivated
with Agrobacterium tumefaciens for two days in dark. After co-cultivation, the leaf
discs were transferred to regeneration medium supplemented with cefotaxime (250
g/ml), kanamycin (100 g/ml) and hygromycine (30 mg/ml). The culture was
incubated in light (photoperiod 16/8) for a period of four weeks. After this, the
transgenic shoots were harvested and transferred to rooting medium containing
kanamycin (50 g/ml). After incubation for 2-4 weeks, the putative transgenic plantlets
were transferred to Hoagland’s solution for acclimatisation and then transferred to
vermiculite for hardening. Kanamycin resistant T0 plantlets of pSR1241 and
hygromycine resistant T0 hardened plantlets were transferred to pot which containing
soil and shifted to the green house until these grown up to maturity or seed setting
stage.
3.2.2 Generation of transgenic tomato hairy roots lines
Plasmid (Figure 3.1B) was electroporated into Agrobacterium rhizogens A4 by
electroporation method (Cangelosi et al., 1991). The second and third leaves of in vitro
grown Solanum lycopersicum plants were used to transform according to a method
described previously by Banerjee et al., (2002). The seeds of Solanum lycopersicum
were germinated in vitro on MS media after 2.5 minute of surface sterilization with
0.1% HgCl2 followed by extensive washing with distilled water. Hygromycin-sensitive
control and Hygromycin-resistance transgenic hairy roots were excised from each
wound site and maintained further at 24oC. Transgenic hairy roots were
micropropagated on MS medium (Medina-Bolivar et al., 2003), supplemented with 100
mg/l Kanamycin and 250 mg/l Cefotaxime.
3.3 Molecular screening of putative transgenic tobacco plants and tomato hairy
roots lines
DNA was isolated from transgenic leaves, hairy roots and non-transgenic
control leaves, roots frozen in liquid nitrogen by using DNeasy mini kit (Qiagen,
Valencia, CA). Transgenic plants were screened for the presence of the ctxB-rgp gene
by PCR, using forward 5’ATCGATGTCGACTAACAACTTCCTCAAGTCTT3’ and
Page 79
Materials and Methods
42
reverse 5’AGATCGTCGACT CATCACAACTCATCCCTCTCGGAC3’ primers. with
cycling parameters: 95°C for 3 min; 35 cycles of 95°C for 1min; 60°C for 1 min, 72°C
for 2 min; 72°C for 5 min.
Detection of target gene in putative transgenic hairy root lines was performed
by PCR. Genomic DNA was isolated and subjected to PCR using end RGP F1 and
Ricin R1 primer of RGP-RTB fusion gene with cycling parameters: 95°C for 3 min; 35
cycles of 95°C for 1min; 62°C for 1 min, 72°C for 2 min; 72°C for 5 min. The
polymerase chain reaction was also performed to confirm the presence of the rol B and
absence of vir C genes in the hairy root and control roots. PCR experiments were
performed by using Rol B F forward and Rol B R reverse primer designed for the
amplification of a 652-bp rol B fragment. Forward primer vir C F and reverse primer
vir C R were used to amplify a 730-bp of vir C fragment. Each PCR reaction was
carried out in 50 μl, containing 200 mM of each dNTP, 0.2–0.5 μg of genomic DNA,
1mM of each primer, 1.25U Taq DNA polymerase (Banglore genei, Banglore) and
PCR buffer contained 2mM magnesium chloride. The mixture was amplified in a
thermal cycler (Thermojet, Eurogentec).
Samples were subjected to 35 cycles of 1 min denaturation at 95°C, 1 min
annealing at 55°C for the amplification of the rol B and vir C fragments, and 1 min
extension at 72°C. Amplified DNAs were detected on 1% (w/v) agarose gels.
3.3 Expression analysis of transgenic lines
3.3.1 Extraction of protein
Tomato hairy root and leave tissues were harvested from culture, drained on
paper towels, and stored at -80°C until use. Protein extracts were prepared as
follows: hairy root and leave tissues were ground in liquid nitrogen with mortar and
pestle and transferred to a centrifuge tube containing Tris-Cl extraction buffer [100
mM Tris base, 150 mM NaCl, 2.5% (w/v) PVP-40, 0.1% Tween 20; pH 8.0] at 3:1
buffer to tissue ratio and vortexed vigorously for 1 min. Extracts were centrifuged at
~ 13,000 x g for 10 min at 4°C and supernatants were filtered through four layers of
MiraclothTM
(Calbiochem-EMD Biosciences, San Diego, CA). The total protein
concentration of crude protein extracts was measured using the Advanced Protein
Assay (Cytoskeleton, Denver, CO) in a Bio-Tek EL808 Ultra Microplate
Page 80
Materials and Methods
43
Reader. Total protein determinations were made in reference to bovine albumin
serum (BSA) standards dissolved in Tris-Cl extraction buffer.
3.3.2 Enzyme Linked Immuno Sorbent Assay (ELISA)
3.3.2.1 Indirect ELISA
Total soluble protein was estimated by the Bradford reagent (Bio-Rad). A 96-
well micro-titre plate was coated with sample (100 l) of total soluble protein of each
transformed and non-transformed lines of tobacco leaves and hairy roots which lysed in
20 mM Tris-Cl buffer (pH 8.0). The plates were incubated overnight at 4C or at 37C
for 2h and processed as per ELISA method described by Harlow and Lane, 1988. The
plate was blocked with 1.0% BSA in PBS containing 0.05% Tween-20 (PBS-T buffer).
Between any two incubations, the plates were washed with PBS-T three times with 2
min soak time on Plate washer (PW-40 Bio-Rad). After blocking, the plates were
probed with the peptide antibody against RGP and equine anti-rabies polyclonal
antibody (primary antibodies) at 1:5000 dilutions and incubated for 2h in PBST
containing 0.25% BSA. The plates were further incubated with ALP conjugated anti-
rabbit and anti-horse IgG in 1:20000 dilutions for 2h. The wells were washed 100 μl
pNPP (p-nitrophenyl phosphate disodium salt) esubstrate (Bangalore Genie,
Bangalore) was applied per well. The reaction was stopped after 15 min by the
addition of 50 μl of 2N NaOH. Absorbance (A405nm) was read in a Microplate Reader
(Bio-Rad).
3.3.2.2 Direct ELISA
(A) GM1-binding ELISA
Expression of ctxB-rgp gene in leaves was determined by quantitative mono-
sialoganglioside-dependent enzyme linked immuno-sorbent assay (GM1-ELISA). Total
soluble proteins of different sample were diluted serially. Volume of 100 l was added
into the wells in triplicate and incubated for 2h. Wells were washed three times between
each step using 300 l of phosphate buffered saline with added Tween-20 (PBST;
0.01M Na2HPO4, 0.003M KH2PO4, 0.1M NaCl, pH 7.4, 0.05% Tween-20 v/v). ELISA
was carried out at 370C. The 96 well micro-titre plates (Greiner, Germany) were coated
for 1h with 3.0 g/ml GM1 made in sodium carbonate coating buffer pH 9.6 (15 mM
Page 81
Materials and Methods
44
Na2CO3, 35mM NaHCO3). As control, BSA (3.0 g/ml) in bicarbonate buffer was
coated in some wells. The wells were blocked with 300 l of 1% BSA in PBST for 1h,
followed by washing with PBST. After washing three times with PBST, the peptide
antibody against RGP and equine anti-rabies polyclonal antibody were added at 1:5000
dilutions and incubated for 2h in PBST containing 0.25% BSA. The plates were further
incubated with ALP conjugated anti-rabbit and anti-horse IgG in 1:20000 dilutions for
2h. Plates were developed with p-nitrophenyl phosphate substrate. The reaction was
terminated by addition of 3M NaOH. The plates were read at 405 nm and the RGP
expression level was quantified on a linear standard curve. All estimations were made
on the basis of three independent experiments.
(B) Asialofeutin-binding ELISA
The functionality of recombinant protein in the protein extracts of transgenic
hairy root lines was determined via binding to asialofeutin. Two hundred microliters
of asialofeutin (Sigma; St. Louis, MO) at a concentration of 100 μg/ml in
biocarbonate buffer (pH 9.6) was coated per well of an Immulon 4HBX (Fisher,
Pittsburg, PA) microtiter plate for 2h at room temperature (RT). The coating solution
was discarded, and the wells were blocked with 300 μL of 1% BSA, 0.05% Tween-20
in PBS for 1h at RT. The blocking solution was discarded and 100 μl each of RTB
standards (described below) and sample protein extracts (prepared as above) was
applied in triplicate wells and incubated for 1h at RT. The blocking solution was
discarded, and wells were washed three times with PBS, 0.1% Tween 20. The rabbit
peptide antibody against RGP and equine anti-rabies polyclonal antibody were added at
1:5000 dilutions and incubated for 2h in PBS-T containing 0.25% BSA at RT. The
wells were then washed as before. Alkaline phosphatase-conjugated anti-rabbit and
anti-horse IgG (Sigma; St. Louis, MO) secondary antibodies were applied at a
1:20000 dilution in blocking buffer and incubated for 1h at RT, then discarded. The
wells were washed three times as before, and 100 μl pNPP (p-nitrophenyl phosphate
disodium salt) substrate (Bangalore Genie, Bangalore) was applied per well. The
reaction was stopped after 15 min by the addition of 50 μl of 2N NaOH. Absorbance
(A405nm) was read in a Microplate Reader (Bio-Rad). Hundred µg of total protein
extract from hairy root tissue or 100 ml of media from hairy root cultures were
Page 82
Materials and Methods
45
assessed compared to a standard curve consisting of serially diluted castor E. coli-
derived SUMO-RGP (Singh et al., 2012) in Tris-Cl buffer (above), in concentrations
ranging from 2.5ng to 500ng RTB per well.
3.4 Polyacrylamide gel electrophoresis of plan proteins
3.4.1 SDS-Polyacrylamide gel electrophoresis
Total soluble plant proteins were electrophoresed on denaturing polyacrylamide
gel for western blot analysis. The SDS-polyacrylamide gel was made from stock of
acrylamide and bis-acrylamide solution (30% w/v in a ratio 29:1). Composition of gel
was as follows
Running gel
Acrylamide-bis-acrylamide → 10% (w/v)
Tris- Cl (pH 8.8) → 0.125 M
SDS → 0.1% (w/v)
TEMED → 0.067% (v/v)
Ammonium per sulfate → 0.33% (w/v)
Stacking gel
Acrylamide-bis-acrylamide → 5% (w/v)
Tris-Cl (pH 6.8) → 0.375 M
SDS → 0.1% (w/v)
TEMED → 0.067% (v/v)
Ammonium per sulfate → 0.33% (w/v)
The electrophoresis was carried out in Mini Protean II Dual Slab Cell System
(Bio-Rad). Thirty g protein extract of transgenic and non-transgenic plants along with
molecular weight markers mixed with equal volume of 2X gel loading buffer
containing glycerol 20% (v/v); 0.1 M Tris-Cl, pH 6.8; 4% SDS, 100mM DTT and 0.2%
(w/v) bromophenol blue. Protein samples were loaded directly on the gel with heat
treatment or boiled for 5 min to dissociate oligomer into monomers. The samples were
centrifuged in microfuge (12,000xg, 5 min, 4oC) and loaded on 10% sodium dodecyl
sulfate polyacrylamide gel. The electrophoresis was carried out in buffer (25 mM Tris-
Cl, pH 8.8; 192 mM glycine and 0.1% SDS) at constant current of 16 mA.
Page 83
Materials and Methods
46
3.4.2 Native polyacrylamide gel electrophoresis
The native State of CTB-RGP and RGP-RTB fusion protein were detected by
using 6% SDS-PAGE. Unboiled (non-reduced) samples were loaded without adding
DTT in sample loading buffer (Areas et al., 2004). The gel was run at constant 30 V for
at least 5h and blotted into PVDF membrane by electro-blotting on Hoefer TransphorTM
apparatus with cooling.
3.5 Western blot analysis
Samples were mixed with equal volume of sample loading buffer (25mM Tris-
Cl, pH 6.8, 2% SDS, 200 mM DTT, 20% glycerol and 0.25% Bromophenol Blue),
immediately boiled in a water bath for 5 min and centrifuged at 13,000xg. The
supernatant was electrophoresed on a 10% Tris-acrylamide gels, a discontinuous SDS-
PAGE in Mini-gel apparatus (Bio-Rad, Hercules, CA) and transferred to 0.2 µm
immunoBlot® PVDF membrane (Bio-Rad) in blotting buffer (25mM Tris-base,
192mM glycine and 20% methanol). All washings, blocking and antibody dilutions
were made in TBS-T buffer (100 mM Tris pH 7.5, 150 mM NaCl, 0.1% Tween-20).
The membrane was blocked with 5% non -fat dry milk powder (Bio-Rad) for one hour,
followed by incubation with the designated primary antibody in blocking solution for
further 2 h. Membranes were washed three times for 5 min each with TBS-T buffer and
incubated with secondary antibody for 2h and washed 3 times as above. The primary
and secondary antibody was used at 2,000-fold and 10,000 dilutions respectively and
the blot was developed with AP substrate colour developer kit (Bio-Rad; Hercules,
CA).
3.6 Southern analysis of transgenic plants
3.6.1 Isolation of the genomic DNA from tissue
The genomic DNA was isolated from mature tissue following modified CTAB
method. Cut-micro tips were used to handle genomic DNA. Following steps were
performed for this purpose. One gram of fresh tissue was ground to fine powder in
liquid nitrogen. 12 ml of CTAB extraction buffer (2%, w/v CTAB; 50 mM Tris-base,
20 mM EDTA, 1.4 NaCl and 0.1% v/v, β-ME) was added to powdered tissue and
homogenized. The mixture was kept at 68oC for 2h for lysis. After incubation, the
Page 84
Materials and Methods
47
temperature of lysate was brought down to the room temperature. The lysate was
extracted with 0.7 volume chloroform: isoamyl alcohol. The aqueous layer was
collected in a fresh tube after centrifugation (SS34 rotor, 10,000 rpm, 10 min, 22oC).
The nucleic acids were precipitated with 0.7 volume isopropanol by keeping on ice for
10 min and then recovered by centrifugation. The pellet was washed with 70% ethanol
and dried at room temperature. The nucleic acids containing genomic DNA were
dissolved in 750 l of 10 mM Tris-Cl, pH 7.4 containing DNase free RNase A (50
µg/ml) and transferred to eppendorf tubes. The tubes were kept overnight at 37oC to
carry out RNA digestion. The genomic DNA was extracted once with phenol:
chloroform: isoamyl alcohol (25:24:1) and twice with chloroform: isoamyl alcohol
(24:1). DNA was precipitated with 0.7 volume isopropanol, dried in air, dissolved in
water and quantified on spectrophotometer.
3.6.2 Digestion of the genomic DNA
Twenty g genomic DNA was digested with Xho I restriction enzymes in 300
l reaction mixture, 5 U/g restriction enzyme was used in two steps. The enzyme was
mixed by swirling the solution with microtip and the digestion was carried out for 16h.
Digested genomic DNA was checked on 0.8% agarose gel to confirm complete
digestion. Rest of the digested genomic DNA was extracted with chloroform: iso-amyl
alcohol. The DNA was precipitated and dissolved in 18 l of TE buffer (pH 8.0).
3.6.3 Transfer of the DNA from gel to nylon membrane
After electrophoresis, the agarose gel was washed with sterile water and placed
in 0.25 M HCl for depurination. The treatment was carried out for 20 min. The gel was
washed with water. The DNA of the gel was transferred to positively charged nylon
membrane (Hybond N+ membrane, Amersham Life Science) with 20X SSC as transfer
buffer following capillary blot method as instructed by the manufacturer and discussed
by Sambrook et al. (1989). The transfer was carried out for overnight. After the
transfer, membrane was washed with 2X SSC buffer, wrapped in Saran wrap and stored
at 4oC.
Page 85
Materials and Methods
48
3.6.4 Hybridization of probe and autoradiography for Southern analysis
Stable integration and copy number of the transgene were established by
Southern hybridization as described in (Ashraf et al., 2005). Hybridization was
performed at 65oC for 16 h, using [α P
32] dCTP labeled probe, comprising 570 bp Xho
I–Age I and 519 bp EcoR V– Hpa I fragment at 3’end of ctxB-rgp and rgp-rtxB genes
respectively. The membrane was exposed to Fuji screen for 24 h and scanned on
phosphor imager (Molecular Imager FX, Bio-Rad, and Hercules, USA).
3.7 Immunization of Balb/c mice
BALB/c mice (5 in each group) were orally primed by 50g of each extracted
and partially purified CTB-RGP, RGP-RTB fusion proteins along with phosphate
buffer saline used as a negative control. Then, three booster doses of 25g were given
on the 7th
, 14th
and 28th
day. Serum was collected after 7th
days from the third booster
means 35th
days. SUMO-RGP + CTB and SUMO-RGP + RTB group represents 25µg
of attenuated viral vaccine was orally given to mice with 25µg of each mucosal
adjuvant, cholera and ricin toxin B subunit, respectively. The mice were bled on 35th
day, from the retro-orbital sinus for the estimation of anti-rabies antibody titre in serum.
3.7.1 Titration of antibody response from sera of immunized mice
The ELISA was carried out in a 96 well micro-titre plate (Nunc Maxisorp). The
micro-titre plates were incubated with 100l per well of the commercial virus based
vaccine (Abhayrab, Indian Immunologicals) at the dilution of 1:50 in PBS-Tween buffer
containing 0.25% BSA at 4oC for 2h. The micro-titre plates were again washed with
PBS-Tween buffer. The washed micro-titre plates were further filled by 100l/ well of
serum (1:100) of different groups of mice and incubated for 2h at 4oC. The micro-titre
plates were then washed and incubated for 2 h at 37oC, containing 100l/ well of
horseradish peroxidase conjugated anti-mouse anti-IgG1 (1:1,000) and anti-mouse anti-
IgG2a (1:1,000) in PBS-T containing 0.25 % BSA at 37oC for 2 h. The chromo-genic
reaction was allowed to take place by adding 100µl/well of tetra-methylbenzidine for
horseradish peroxidase conjugates and reaction was allowed for 30 min at 37oC. The
enzymatic reaction was stopped by adding 1N sulphuric acid (50µl/well). Absorbance
was measured at 450 nm.
Page 86
Materials and Methods
49
3.8 In silico analysis of CTB-RGP and RGP-RTB fusion proteins
An alignment of the amino acid sequences of Ricin chain B chain, Rabbies
glycoprotein G and Hybrid was produced with Clustal W program. This alignment
project file was submitted to Swiss‐Model in the expasy server
(http://www.expasy.ch/spdbv) and a preliminary model for Hybrid was retrieved. The
tertiary structure with PDB file of RGP-RTB fusion protein was generated by using
MULTICOM protein tertiary structure prediction server (http://www.molbiol-
tools.ca/protein_tertiary structure.htm) which uses multiple templates for homology
dependent tertiary structure prediction. We used RAMPAGE : Ramachandran
Plot Assessment tool to analyse the quality of the deduced model
(http://mordred.bioc.cam.ac.uk/~rapper/rampage.php). Pymol software was used for
superimposition study. The Hopp–Woods hydrophilicity scale of amino acids is a
method of ranking the amino acids in a protein according to their water solubility in
order to search their surface locations on proteins, especially those locations that tend to
form strong interactions with other macromolecules and antigenically important was
done by using Sci-Ed software. Protparam analysis of CTB-RGP and RGP-RTB fusion
proteins were also perform.
3.9 Up-scaling of the selected hairy root clone in bioreactor
In course of the present study, attempts were also made for up-scaling the
selected hairy root clone of Lycopersicon esculantum expressing RGP-RTB in a 5L
Bench top fermenter (model Bioflo-3000 from M/s New Brunswick Scientific Co. Inc.,
USA) through applying the design modifications, already established for the scaling-up
of hairy root cultures of other plant system (Banerjee et al., 2002) and optimization of
critical parameters.
Design modifications which were employed for the present study included the
following amendments: (i) inclusion of a nylon mesh septum (pore size 200u),
tightened on the lower stainless steel semi-circular ring of the baffle assembly, dividing
the reactor vessel into an upper and a lower chamber, (ii) the marine blade impeller was
positioned in the lower chamber (9 cm down from the nylon septum) and sparger was
placed in lower chamber (12cm down the septum); the agitation speed was kept
Page 87
Materials and Methods
50
constant at 100 rpm and the air flow rate was fixed at 1.3L per minute using an air-
compressor via hydrophobic membrane filter (0.22µ; Whatman, USA).
The culture conditions were maintained as follows:
(i) 2.5 liters of half -strength MS medium with B5 vitamins and 3% sucrose was
used in the bioreactor vessel;
(ii) Initial inoculum of 4.0 g (FW) per liter was used, which was obtained from
fifteen days old hairy root cultures and was inoculated aseptically through the
optimally-flamed inoculation-port of the bioreactor under a sterile laminar-hood
(iii) Culture temperature was maintained constantly at 25±2° C by circulating cold
water from a chiller attachment through the cooling jacket;
(iv) Roots were grown in dark.
(v) The culture period continued till the 21st day of inoculation.
After 21 days of sterile run, the roots were harvested through dismantling the
vessel-assembly, washed under running tap water, blotted dry between filter papers and
weighed on a Mettler pan balance to determine the fresh weights. The harvested root
sample was analysed through ELISA as per the already stated method in-order to
quantify the contents of the two specified proteins, i.e. RGP and RTB.
Page 88
Chapter 4
Observations
Page 89
Observations
51
4.0 OBSERVATIONS
4.1 Construction of ctxB-rgp and rgp-rtxB fusion genes
The amplified coding sequences of ctxB-rgp and rgp-rtxB fusion gene were
cloned in pBI101 and pCAMBIA1300 vector, respectively, in a way that the complete
ORF produces CTB-RGP (Figure 3.1A) and RGP-RTB protein, respectively (Figure
3.1B). The ctxB-rgp and rgp-rtxB fusion gene were containing 63bp of N-terminal ER
targeting pathogen responsive signal sequence (PR-S) sequence and 27bp of ER
targeting Calreticulin (Cal-S) sequence from tobacco, respectively. Both the fusion
protein C-terminally extended for 18bp of SEKDEL, an ER retention signal which were
amplified through primer extension method.
The pr-s-ctxB was PCR amplified using a forward primer with XbaI and a
reverse primer with SmaI site using pSM31 (Mishra et al, 2006) as the template DNA.
The reverse primer was designed to include codons of glycine-proline hinge at the
3’end of ctxB. SmaI containing forward primer and SacI containing reverse primer were
used for PCR amplification of rgp gene from pSA5. Amplified PCR fragments were
digested with the respective enzymes and triply ligated in pBI101 vector containing the
enhanced CaMV35S promoter to obtain pSR1241. The size of ctxB-rgp fusion gene is
2.04 kb along signal sequences and two glycine-proline repeats as hinge.
The rgp-rtxB gene was amplified with forward and reverse end primer which
contains PstI and SacI restriction sites, respectively. PCR amplified product was blunt
end ligated into EcoRV digested pBluescript SK+.
After validating the fusion gene
through sequencing, restriction digestion was perform with PstI and SacI which
produced the band of and rgp-rtxB fusion gene of 2.24 kb which was purified from
agarose gel and finally subcloned into the downstream of CaMV35S double enhancer
containing pCAMBIA1300 plant expression vector. The expression construct was
known as pAS1.
4.2 Transformation, molecular screening of putative transgenic tobacco and
tomato hairy root lines
Plasmid pSR121 and pAS1 were transformed into tobacco (Figure 4.1). Five
lines for ctxB-rgp (Figure 4.2A) and eight lines for rgp-rtxB (Figure 4.2B)
independently transformed kanamycin resistant tobacco plants were verified for the
Page 90
(C)
(A) (B)
Figure 4.1: A. tumefaciens strain LBA4404 containing pSR1241and pAS1
plasmid was used for transformation of tobacco leave discs.
(A) Transgenic shoot induction from leave discs.
(B) Transgenic shoots elongation and their respective selection on
Kanamycin and Hygromycin containing media.
(C) Acclimatized mature transgenic tobacco plants in glass house.
Page 91
Figure 4.2: (A) PCR detection for stable integration of ctxB-rgp fusion gene in
genomic DNA of the transgenic tobacco lines. M, λ DNA marker,
PC, positive control (plasmid DNA); lane 1-5, transgenic tobacco
lines and NT is non transgenic tobacco.
(B) PCR amplification of rtxB from stable integration of rgp-rtxB
fusion gene in genomic DNA of the transgenic tobacco lines.
M, PC and NT are showing 100bp marker, plasmid DNA and
non- transgenic tobacco lines, respectively.
(A)
(B)
M 1 2 3 4 5 NT PC
2.0 Kb
800 bp
M NT 1 2 3 4 5 6 7 8 PC
Page 92
Observations
52
presence of respective genes by PCR amplification of the genomic DNA. The expected
amplification product of ~2.0 kb for ctxB-rgp gene (lanes 1-5) and ~800bp of rtxB gene
from rgp-rtxB gene (lanes 1-8) was observed in all the plants along with respective
positive control plasmid (lane PC) whereas genomic DNA of the non-transgenic plant
gave no amplification (lane NT). The quantitative analysis of ctxB-rgp (Figure 4.3A)
and rgp-rtxB (Figure 4.3B) gene expression carried out by ELISA showed the highest
expression level in plant T01 and T05, respectively. Transgenic tobacco plant T01 of
ctxB-rgp and T05 of rgp-rtxB shows 5-fold and 3-fold higher expression as compared
to that of least expressing plants, respectively.
Plasmid pAS1 was also transformed in tomato hairy root lines (Figure 4.4).
Initially eight hairy root lines were isolated and kept on selection of 15 mg/l
hygromycin for two cycles of 10-10 days with 250 mg/l Cefotaxime. Out of eight hairy
root lines, five hairy root lines remained healthy however three stopped proliferating
and showed necrosis in root tip region. The hairy root lines were screened by PCR
using rgp-rtxB fusion gene spe f e od ed 2.1kb band from isolated
genomic DNA of different transgenic hairy root lines (Figure 4.5A). Lane 1 to 5 of
figure 4.5A shows amplification from transgenic hairy root lines from H01 to H05,
respectively. Lane NT and PC, shows amplification from hairy roots which developed
from non-transformed A4 strain as negative control and pSA1 plasmid as a positive
control, respectively. Primers of vir C and rol B gene were used for validating as
negative and positive control of Agrobacterium contamination in the genomic DNA of
hairy root lines. A band of 730 bp of vir C in lane PC was amplified through isolated
plasmid pAS1 from transformed A. rhizogens (Figure 4.5B). In figure 4.5B except lane
PC which is a positive control, all the lane 1-5 of five hairy root lines from H01-H05
and NT of negative control shows no amplification, respectively. Figure 4.5C, shows
the amplification of rol B gene. Lane PC shows amplification of the positive control
and same as vir C lane 1-5 shows amplifications from genomic DNA isolated from
hairy root lines and NT is negative control. The quantitative analysis of rgp-rtxB gene
expression in hairy root lines carried out by ELISA (Figure 4.6) showed the highest
expression level in plant H03. Transgenic tomato hairy root lines H03 of rgp-rtxB
showed 5-fold higher expression as compared to that of least expressing line H05,
respectively.
Page 93
Figure 4.3: (A) Determination of CTB-RGP expression in different T0 lines
of N. tabaccum leaves by indirect ELISA with Ab1 equine
anti-rabies antibody, NT is non-transformed plant.
(B) Determination of RGP-RTB expression in different T0 lines
of N. tabaccum leaves by indirect ELISA with Ab1 equine
anti-rabies antibody, NT is non transformed hairy root.
(A)
(B)
Page 94
Figure 4.4: (A) Plasmid pSA1 containing, Agrobacterium rhizogens strain A4
mediated induction of hairy root culture from the leaves of
Solanum lycopersicum grown in ¼ MSP solid media.
(B) Transformed in vitro grown roots of Solanum lycopersicum after
grown in 28th days in ¼ MSL in liquid media.
(C) Scale-up process of selected hairy root line (H03) in 5L air lift
bioreactor for large production and isolation of candidate protein.
(B)(A)
(C)
Page 95
Figure 4.5: (A) PCR detection for stable integration of rgp-rtxB fusion gene in
genomic DNA isolated from the transgenic hairy root lines. M,
λ DNA marker, PC, positive control (plasmid DNA); lane 1-5,
transgenic lines H01- H05 and NT is non-transgenic hairy root
lines.
(B) PCR amplification of vir C region in plasmid DNA (PC) and its
absence of selected hairy root lines. M and NT are 100bp marker
and non-transformed lines, respectively.
(C) PCR amplification of rol B region. Lane: 2-5, selected hairy lines;
NT, non-transformed control in vitro hairy roots and M is 100bp
marker.
M PC 1 2 3 4 5 NT
M 1 2 3 4 5 NT PC
M PC 1 2 3 4 5 NT
730 bp
(A)
(B)
(C)
2.0 Kb
652 bp
M 1 2 3 4 5 NT PC
M PC 1 2 3 4 5 NT
M PC 1 2 3 4 5 NT
Page 96
Figure 4.6: Determination of RGP-RTB expression in different lines of
Solanum lycopersicum hairy roots by indirect ELISA with
Ab1 equine anti-rabies antibody, NT is non transformed
hairy root.
Page 97
Observations
53
The quantitative analysis of rgp-rtxB gene expression in both hairy root lines
and tobacco shows RGP-RTB protein expression. However difference in expression of
rgp-rtxB gene between highest expressing hairy root line (H03) and tobacco leaves
(T05) is approximately 4-fold higher. Comparison between expression analysis of
tobacco leaves and hairy root lines suggest that hairy root lines have higher expression
level probably due to the lower background protein concentrations and easy in vitro
scalability than tobacco. Therefore detailed studies of RGP-RTB protein expression and
immunogenicity were conducted further on tomato hairy root lines.
4.3 Growth Kinetics of tomato hairy root lines
Growth kinetics study of hairy root lines was conducted for the optimization of
harvesting time. Out of eight hairy root lines, six were healthy when grown on selection
media. The six selected hairy root clones were further transferred to liquid half strength
MS medium with B5 vitamins containing 3% sucrose to study their growth
characteristic during different growth phases i.e. 14, 21, 28 and 35 days. Although the
morphological appearance of these five clones was quite similar initially, two of these
clones could be distinguished from the rest three by increased lateral branching and
healthy appearance. A gradual increase in fresh weight up to 21 days of culture could
be noted with all the root clones, amongst which two clones H04 and NT (lane 4 and 6)
continued to grow further even up to 35 days of culture while the rest four clones H01,
H02, H03 and H05 (lane 1, 2, 3 and 5) exhibited a decline in their growth after 21 days
of culture (Figure 4.7). The growth increment was highest with the hairy root clone
H03 which exhibited a maximum of 29.6 fold increase in biomass over 21 days of
culture followed by that of the H02 and H01 after the same period of time.
4.4 Optimization of harvesting time for ctxB-rgp and rgp-rtxB genes expression
by indirect ELISA
Tobacco leaves of T01 line was selected to validate the optimum time of
harvesting on the basis of CTB-RGP fusion protein accumulation. The data of maturing
transgenic leaf was collected on different time interval which is shown in figure 4.8A.
The accumulation of the protein was maximum on the 14th
day (2nd
week from
inoculation) and gradually decreased as culture proceeded to 21st, 28
th and 35
th days of
maturity (Figure 4.8A).
Page 98
Figure 4.7: Comparative study of growth kinetics analysis of selected transgenic
hairy root lines (H01-H05) of rgp-rtxB fusion gene from 1st to 5th and
6th is non-transformed in-vitro grown control roots during different
growth phases.
0
5
10
15
20
25
30
35
1 2 3 4 5 6
Gro
wth
ind
ex
Number of lines of hairy root cultures
14 days
21 days
28 days
35 days
Page 99
Figure 4.8: Optimum time course study for harvesting transgenic plants and
hairy root lines. Tissues were harvested at each interval of 7, 14,
21, 28 & 35 days and did ELISA by Ab1 equine raised poly clonal
anti-rabies antibody.
(A) Transgenic tobacco plant T01 has highest CTB-RGP protein
expression so that leaves from this plant was taken for the study.
(B) Tomato hairy root line H03 has highest RGP-RTB protein
expression so that taken for the standardization of optimum
harvesting time.
(A)
(B)
Page 100
Observations
54
H03 line of hairy root was selected to validate the optimum time of harvesting
hairy root culture on the basis of RTB-RGP fusion protein accumulation. Figure 4.8B
shows that the accumulation of the protein was maximum on the 21st day (3
rd week
from inoculation) and gradually decreased as culture proceeded to 28th
and 35th
days of
post inoculation (Figure 4.8B).
4.5 Immunological activities of CTB-RGP and RGP-RTB fusion proteins
Functionality of CTB-RGP fusion protein was correlated with the ability to bind
to GM1-ganglioside. Three microgram per milliliter of GM1 was optimized within the
linear range of binding at variable molar concentrations of CTB-RGP fusion protein.
Immunological activity of the fusion protein CTB-RGP was checked by performing
both direct GM1-ELISA (Figure 4.9A) and indirect ELISA (Figure 4.3A). Fourteen
days mature leaves of all the transgenic tobacco lines were harvested for analyzing the
immunological activity of fusion protein. Peptide antibodies (Ab2) against the rabies
glycoprotein as well as equine anti-rabies antibodies (Ab1) were used as primary
antibodies in direct GM1-ELISA. The binding of the CTB-RGP protein to GM1
receptors in GM1-ELISA established that the fusion protein expressed in tobacco
leaves was in pentameric form and immunologically active against the rabies
antibodies. It is also established that CTB is in their right confirmation so that they
attain pentameric structure and bind to its receptors. According to indirect ELISA,
CTB-RGP fusion protein of T01 transgenic tobacco line has higher expression and
immunological activity. Consistently in T01 to T05, Anti-rabies antibodies (Ab1) show
more absorbance intensity as compared to peptidal anti-rabies antibody (Ab2).
All the hairy root lines were harvested on the 21st day for analyzing the
maximum accumulation of RGP-RTB fusion protein and immunological activity.
Asialofeutin mediated direct ELISA of all the 5 transgenic hairy lines was performed
with two different antibodies, equine anti-rabies (Ab1) and peptide anti-rabies antibody
(Ab2) (Figure 4.9B). Direct ELISA showed that after 21st day of inoculation of hairy
root line, H03 had maximum accumulation of fusion protein followed by line H01
which had second highest accumulation. This result is in agreement with the result of
indirect ELISA (Figure 4.6). It suggests that hairy root expressed RGP-RTB fusion
protein is immunological active and shows higher expression than CTB-RGP from
leaves of tobacco lines.
Page 101
Figure 4.9: (A) CTB-RGP expression in different T0 lines of Transgenic
tobacco by GM1 receptor based ELISA with Ab1 equine
anti-rabies and Ab2 peptide anti-rabies antibody, NT was
absorbance of non transformed plant.
(B) RGP-RTB expression in different lines of Solanum
lycopersicum hairy roots by asialofeutin receptor based
ELISA with Ab1 equine anti-rabies and Ab2 peptide anti-
rabies antibody, NT was absorbance of non transformed
hairy root .
(A)
(B)
Page 102
Observations
55
4.6 Quantitation of CTB-RGP and RGP-RTB fusion proteins in highest
expressing transgenic lines
The level of pentameric form of CTB-RGP protein expressed in transgenic
tobacco lines was determined by monosialoganglioside dependent enzyme linked
immuno-sorbent assay (GM1-ELISA) of total leaf protein. The TSP extracted from the
leaves of T01 plant showed highest level of CTB-RGP expression (0.4%) whereas
second highest expression observed in T03 (0.21%) plant (Figure 4.10A).
Tomato hairy root lines of H01 and H03 were further taken into account for the
estimation of accumulated RGP-RTB fusion protein by asialofeutin mediated ELISA
(Figure 4.10B). As depicted in figure 4.10B, line no. H01 and H03 showed 0.9 and
1.2 % of total soluble protein accumulation, respectively. Hairy root lines H01 and H03
contained 6 and 8µg of RGP-RTB fusion protein in per gram of tissue, respectively.
4.7 Western blot analysis of CTB-RGP and RGP-RTB fusion proteins in native
and denaturing conditions
Western blotting was attempted for qualitative analysis of transgenic protein and
its accumulation in the tissue. The transgenic tobacco plants T01, T02 and T03 which
showing high expression of CTB-RGP fusion protein, were selected for analysis on
denaturing immunoblot assay. Under the denaturing conditions, a band of ~80.6kDa
(~66kDa glycosylated RGP + ~14.6kDa glycosylated CTB polypeptides), representing
the monomeric fusion polypeptide was detected by using equine anti-rabies antibodies
(Ab1) while non-transformed plant (NT) showed no band (Figure 4.11A). As expected
in the non-reduced and un-boiled condition, a pentamer size of (~403 kDa) CTB-RGP
fusion protein was detected by equine anti-rabies antibodies (Figure 4.12A). No band
was detected in the non-transgenic plants. The highest expression of CTB-RGP protein
was noticed in plant T01 in figure 4.11A which is consistent with the ELISA result.
The transgenic hairy root lines H01, H02, H03 and H04 showing high
expression of RGP-RTB fusion protein, were selected for analysis on denaturing
immunoblot assay. Under the denaturing conditions, a band of ~84kDA (~52.5kDa
glycosylated RGP + ~31.5kDa glycosylated RTB polypeptides) was seen in each lane
from H01 to H04 by using equine anti-rabies antibodies (Ab1) while non-transformed
plant (NT) showed no band (Figure 4.11B). In the non-reduced and un-boiled
Page 103
Figure 4.10: Quantitative expression of CTB-RGP (A) and RGP-RTB (B)
fusion proteins in their respective high expression transgenic
lines by receptor mediated ELISA with peptidal anti-rabies
antibody (Ab2).
(A)
(B)
Page 104
Figure 4.11: (A) Western blot analysis of transgenic tobacco plants which
contain ctxB-rgp fusion gene under denaturing condition
by using anti-rabies antibody (Ab2). Crude protein (30 µg)
prepared from the leaves of non-transgenic (NT) and
transgenic plants T01, 2 and 3 was loaded along with
molecular weight markers (M).
(B) Western blot analysis of transgenic tomato hairy root lines
which contain rgp-rtxB fusion gene under denaturing
condition by using peptide anti-rabies antibody (Ab1).
Crude protein (50 µg) was prepared from the hairy root
lines H01, H02, H03, H04, NT (non-transformed) lines
and loaded along with molecular weight markers (M).
M H01 H02 H03 H04 NT
≈ 84 kDa
M T01 T02 T03 NT
≈ 81 kDa
(A)
(B)
Page 105
Figure 4.12: Western blot analysis of transgenic lines under non-denaturing
condition by using polyclonal anti-rabies antibody (Ab2).
(A) Crude protein (30 µg) was prepared from the tobacco line of
ctxB-rgp fusion gene (T01), loaded at lane 2; M, marker and NT
is non-transformed line.
(B) Crude protein (50 µg) was prepared from the tomato hairy root
line of rgp-rtxB fusion gene H03), loaded at lane 3; M, marker
and NT is non-transformed line.
≈ 403 kDa
≈ 168 kDa
(A)
(B)
M T01 NT
M NT H03
Page 106
Observations
56
condition, ~168kDa dimer size of RGP-RTB fusion protein was detected in H03 line of
hairy root line by equine anti-rabies antibodies (Figure 4.12B). As in ELISA result,
line H03 showed highest expression of RGP-RTB protein in figure 4.11B.
4.8 Southern analysis of transgenic tobacco and hairy root lines
The transgenic tobacco plant T01 showing the highest expression of ctxB-rgp
gene was further analyzed by Southern hybridization for determining the copy number
of the transgene insertion and stable integration. The transgenic plant T01 contained
single copy of the transgene inserted into its genome while non-transformed (NT)
showed no band (Figure 4.13A). XhoI digested linear plasmid of pS1241 was used as
positive control.
Similarly, transgenic tomato hairy root line H03 showing the highest expression
of rgp-rtxB gene, was further analyzed by Southern hybridization for determining the
copy number of the transgene insertion and stable integration. The transgenic hairy root
line H03 contained single copy of the transgene inserted into its genome while non-
transformed (NT) showed no band (Figure 4.13B). HpaI digested linear plasmid of
pAS1 was used as positive control and shown a band.
4.9 Immune response of mice against CTB-RGP and RGP-RTB fusion proteins
The results of the mice experiments (Figure 4.14) show that both the CTB-RGP
and RGP-RTB fusion proteins enriched plant fraction elicits an immune response in
mice. However, the ratio of IgG2a and IgG1 indicate that mice have both Th1 and Th2
response and induction of Th2 is more than Th1 response for both fusion proteins.
Although, the intensity of immune response in RGP-RTB fusion proteins is little higher
than CTB-RGP fusion protein, overall immune response is lower for both proteins. It
suggests that quantity of oral dose for both the fusion proteins were lower and need to
optimize further with higher quantity of priming and booster doses. Oral delivery of
attenuated virus vaccine with cholera toxin B and ricin toxin B subunit (Virus + CTB
and Virus + RTB) were not induced significant immune response.
4.10 Scale-up process of tomato hairy roots
The high protein expressing hairy root line (H03) was further set for the up-
scaling through its cultivation in a structurally modified mechanically agitated
Page 107
H03 PC NTPC T01 NT
(A) (B)
Figure 4.13: (A) Detection of chimeric ctxB-rgp gene in T0 lines of
transgenic tobacco plants by Southern hybridization
analysis. Lane NT, non-transgenic; T01, transgenic line
T01 and PC represent positive control plasmid.
(B) Detection of chimeric rgp-rtxB gene in transgenic tomato
hairy root lines by Southern hybridization analysis. NT,
non-transgenic; H03, transgenic hairy root line H03 and
PC represent positive control plasmid.
Page 108
Figure 4.14: Immune response against both the fusion protein CTB-RGP
and RTB-RGP in five Balb/c mice of each group. Both fusion
proteins were orally administered (OD) to each mice of every
group in the following manner of regime 0, 7, 14, 21 and 35.
Virus + CTB and Virus + RTB group represents attenuated
virus vaccine was orally given to mice with cholera toxin B
and ricin toxin B subunit of mucosal adjuvants, respectively.
Page 109
Observations
57
bioreactor of 5l working capacity consisting of earlier mentioned modification for
optimum growth (Figure 4.4C).
With the earlier optimized bioreactor configuration as described by Banerjee et
al., (2002), a 49.6 fold growth enhancement (496.3 g FW) could be obtained after the
same period of 21 days of culture through controlled air-flow rate (1.5 l/min) and
agitation speed of 50 rpm. Although in terms of fold increase, this growth rate was
higher than that noted in case of the shake flask grown roots which exhibited 29.6 fold
growth increases within the same growth period (Figure 4.7), the biomass yield per
liter of the bioreactor was higher than that of the shake flask grown roots (Table 4.1).
Both in terms of protein expression of RGP-RTB and productivities,
comparable results could be obtained between that recorded in the shake-flask (Figure
4.4B) and bioreactor grown roots (Figure 4.4C). However, productivity recorded
during the bioreactor run remained comparatively lower than shake flask which might
be due to callogenesis of the root tissues in bioreactor.
4.11 In-silico analysis of CTB-RGP and RGP-RTB fusion protein
The predicted tertiary structure of CTB-RGP and RGP-RTB fusion protein are
shown in figure 4.15 and their quality assessment was done and cross verified by
corresponding Ramachandran plot (Figure 4.16). However, in comparison to RGP-
RTB, CTB-RGP shows more scattering in Ramachandran plot. But the quality of
tertiary structure model of CTB-RGP and RGP-RTB fusion proteins are in agreement
with the Ramachandran plot.
Superimposition study of native tertiary structure of Rabies glycoprotein and
cholera toxin B subunit (PDB) was performed with CTB-RGP fusion protein (Figure
4.17A). It showed complete superimposition to native cholera toxin B subunit and
achieved its actual tertiary confirmation so that it could bind readily with GM1
receptors. Similarly as above superimposition study of native tertiary structure of
Rabies glycoprotein and Ricin B chain was performed with RGP-RTB fusion protein
(Figure 4.17B) showed complete superimposition to native ricin B chain and achieved
its confirmation so that it could bind readily with their receptors. This study is in total
agreement with the results of GM1 and asialofeutin mediated ELISA for CTB-RGP and
RGP-RTB fusion proteins. However, Superimposition of Rabies glycoprotein (RGP),
Page 110
Table 4.1: Comparative analysis of growth performance and protein expression
Parameters 250 ml flask 5 l bioreactor
Medium Volume 50 ml 2.5 l
Inoculum Weight (g) 0.18 10.0
Final fresh weight (g) 5.32 493.6
Biomass yield (g/l) 102.4 108
RGP RTB (% expression) 7.3 7.54
Comparative analysis of growth performance and protein expression of selected
hairy root clone H03 that cultivated under shake flask and bioreactor conditions for
21 days.
Page 111
(B)
(A)
Figure 4.15: Cartoon representation of both CTB-RGP and RGP-RTB fusion
proteins.
(A) Tertiary structure of CTB-RGP shows topology composed of N-
terminally attached Cholera toxin B Chain (Blue) with Rabies
glycoprotein (Green) using GlyProGlyPro linker.
(B) Tertiary structure of RGP-RTB shows topology composed of C-
terminally attached Ricin B Chain (Blue) with Rabies
glycoprotein (Green) using GlyProGlyPro linker (Red).
Page 112
(A)
(B)
Figure 4.16: Ramachandran Plot of Phi (ψ) and Sci (ϕ) of both the
fusion proteins. (A) CTB-RGP, (B) RGP-RTB.
Page 113
(A)
(B)
Figure 4.17: Superimposition studies of overall cartoon structure of both the
fusion proteins.
(A) Tertiary structure of Cholera toxin B Chain and Rabies
glycoprotein shown in Blue and Green in fusion protein CTB-
RGP while superimposed CTB and RGP of PDB database shown
in Green and pink colours, respectively.
(B) Tertiary structure of Ricin toxin B Chain and Rabies glycoprotein
shown in Orange and Green in fusion protein RGP-RTB while
superimposed RTB and RGP of PDB database shown in Sky Blue
and Blue colours, respectively.
Page 114
Observations
58
between CTB-RGP and RGP-RTB fusion proteins had somewhat similar confirmation
(Figure 4.18) and showed very little variation (RMS 4.89).
Different important antigenic epitopes and active sites which are crucial for
structural confirmation and stability of rabies glycoprotein (RGP) are shown in CTB-
RGP and RGP-RTB fusion proteins (Figure 4.19). Hopp-Wood hydrophilicity and
surface exposure plot of CTB-RGP and RGP-RTB proteins (Figure 4.20) showed great
harmony with overall predicted tertiary structure of fusion proteins. K202 is
participating in conformational stability of rabies glycoprotein. Whereas, sites A (32-
42aa), B (198-200aa), C (208-216) and D (286-306) constitute the antigenic site II
important for the antigenicity of rabies glycoprotein. R333 are syncytium formation site
and form antigenic site III of rabies glycoprotein. These antigenic epitopes are
commonly marked as A (32-42), B (198-200), C (208-216) and D (286-306) sites in
both the fusion proteins in figure 4.19 & 4.20 (Prehaud et al., 1988; Tomar et al., 2011).
Asialofeutin and GM1 mediated direct ELISA with peptidal anti-rabies antibody (Ab2)
which rose against the D epitopic region of rabies glycoprotein complement the result
of in-silico analysis.
Protparam analysis of CTB-RGP and RGP-RTB fusion proteins showed that
isoelectric point (pI) of both the protein was not neutral (7.28 and 6.68, respectively),
they could not precipitate in alkaline solution of pH8.0 or above. However, RGP-RTB
has negatively charged than neutral CTB-RGP fusion, hence immunogenicity of RGP-
RTB was likely to be more which has also seen in the results of animal experiment
(Table 4.2). Half-life of both the fusion protein as predicted by protparam is 30 hours
in human blood (Table 4.2).
Page 115
Figure 4.18: Super imposition of the cartoon structures of rabies glycoprotein
from both the fusion protein, CTB-RGP (Yellow) and RGP-RTB
(Blue) together to asses any overall change in the structure of
protein .
Page 116
R333 K202
(B)
(A)
Site C Site D Site A Site B
Figure 4.19: Surface representation of antigenic sites of RGP on both CTB-RGP
(A) and RGP-RTB (B) fusion proteins. Conformationally important
sites K202 and R333 of RGP shows in Hot Pink (A)/ Yellow color
(B) and Brown (A)/ Orange (B) color, respectively. Whereas
important antigenic sites A (34-42aa), site B (198-200aa), site C
(208-216aa) and D (286-306aa) represents as Red, Blue, Raspberry
Red and Cyan color, respectively.
Page 117
A B C D
A B C D
(A)
(B)
Figure 4.20: Hopp-Wood (HW) and Surface Exposure (SE) plots for CTB-
RGP and RGP-RTB fusion proteins. A (32-42), B (198-200),
C (208-216) and D (286-306) represent potent antigenic sites
as described by other research groups.
Page 118
Table 4.2: Comparative analysis of CTB-RGP and RGP-RTB fusion proteins
by ProtParam software
S.
No. Different Fields
Fusion Protein
CTB-RGP RGP-RTB
1. Number of atoms 9992 11355
2. Formula C3175H4977N867O938S35 C3607H5637N997O1076S38
3. Molecular weight 71424.7 81403.7
4. Number of amino acid 639 731
5. Theoretical pI 7.28 6.68
6.
Number of negatively
charged residues
(Asp + Glu)
70 74
7.
Number of positively
charged residues
(Arg + Lys)
70 71
8. Extinction
coefficient
Assuming all
pairs of Cys
residues form
cystines
87415 130635
Abs. of 0.1%
(=1g/l) 1.224 1.605
9.
Estimated half-life in
mammalian reticulocytes
(in vitro)
30 hours 30 hours
10. Aliphatic index 82.50 84.62
11. Grand average of
hydropathicity (GRAVY) -0.253 -0.247
Page 119
Chapter 5
Discussion
Page 120
Discussion
59
5.0 DISCUSSION
Plant-based biopharming is a powerful tool for mass production of recombinant
and industrial proteins. The molecular farming of pharmaceuticals in plants eliminates
the risk of human or animal virus and prion transmission or contamination by harmful
chemically synthesized products or process solvents. Many of the quality control tests
that require animals also can be eliminated (Joshi and Lopez, 2005). It has already been
shown by various authors that genes encoding antigens of bacterial and viral pathogens
can be expressed in plants in a form at which they retain their native immunogenic
properties. Plants acting as a source of vaccines would offer the advantage of
inexpensive production of the vaccines which could be made easily available to
developing countries (Ma JK-C et al., 2005). Advances in molecular biology of
diseases have identified many candidate proteins or peptides that may function as
effective subunit vaccines however their availability in biologically functional form and
at an affordable cost still remains a challenge.
Rabies virus genome encodes five major proteins- nucleoprotein -N,
phosphoprotein -P, matrix protein -M, glycoprotein -G and RNA-dependent RNA
polymerase -L (Coslett et al., 1980; Conzelmann et al., 1990). The rabies glycoprotein
(G) plays an important role in viral pathogenesis and functions as a protective antigen
(Wiktor et al., 1973). The G protein is a trans-membrane protein that forms the spikes
of the virus, induces virus neutralizing antibodies (VNA) and gives protection against
intra-cerebral challenge. Rabies glycoprotein (G-protein) also induces the production of
T-helper (Th) and cytotoxic T-cells (CTL). The three dimensional structure of G
protein molecule is critical for both virus neutralizing antibody induction and protection
against the rabies.
Several approaches have been suggested for the preparation of rabies subunit
vaccine. The rabies glycoprotein expressed by baculo virus vectors gives protection
against GT1 (CVS G protein), and GT3 (Mok G protein). However, entire cells were
used for injection and this approach would be of limited use for parenteral vaccination
(Prehaud et al., 1989). Other alternate like, recombinant vaccinia virus (Rupprecht et
al., 1986), adenovirus (Prevec et al., 1990) and animal cell lines are also considered as
less desirable for developing human rabies vaccines because of safety concerns.
However, rabies glycoprotein fairly produced in E. coli, shown immunogenicity (Singh
Page 121
Discussion
60
et al., 2012) but lacks immuno-protection activity probably due to the absence of
appropriate post-translational modifications (Yelverton et al., 1983; Lathe et al., 1984).
On the other hand, yeast eukaryotic system was also used for producing the viral
antigen because of its ability to produce the glycosylated form of proteins. But it gives
protection only against intramuscular and not intra-cerebral virus challenge due to
differences in yeast based glycosylation (Klepfer et al., 1993). Glycosylation of rabies
glycoprotein plays important role in their conformation and also their immunogenic
property. The extracellular domain of rabies virus glycoprotein have three important N-
glycosylation sequins at Asn37, Asn247 and Asn319 out of which N-glycans of
Asn319 is mainly responsible for conformationally relevant form of the glycoprotein
(Wojczyk et al., 1998). The glycosylation of Asn319 is critical in stabilizing the local
confirmation of region as it contains syncytium formation sites at Asn333 (Seif et al.,
1985; Tuffereau et al., 1989) and the putative leucine zipper domain at Asn378-Asn399
responsible oligomerization in ERA strain of the rabies glycoprotein of rabies virus
(Conzelmann et al., 1990), thus allowing proper folding of soluble rabies glycoprotein.
Misfolded and nonglycosylated proteins bind to the grp78/BiP chaperon which can
prevent secretion or cell surface expression and lead to the intracellular proteolysis of
rabies glycoprotein (Wojczyk et al.,1998; Hurtley et al.,1989). Glycosylation of the
RGP is required for immuno-protection by the rabies vaccines (Foley et al., 2000). The
least expensive rabies vaccine used in developing countries is prepared from animal
brain which gives undesirable reactions or consequences in some cases and is not
affordable by the poorest. This has hampered the mass immunization program against
rabies. Hence, affordable and safe rabies vaccines are always needed.
Among plant-based systems, the expression of rabies glycoprotein (RGP) has
been reported in transgenic tomato (McGarvey et al., 1995). However, the expression
level was very low and immuno-protective ability was not examined. Rabies virus
surface glycoprotein gene (rgp) had earlier been engineered and transformed in tobacco
plants in our laboratory which gave complete protection to the mice against the live
virus challenge and led the foundation for the development of plant based rabies
vaccine (Ashraf et al., 2005). Since the expression was low and an alternative method
was required to produce functionally active form of protein (glycosylated RGP) in
sufficient quantity from plant. This creates major constraint for using plant as
Page 122
Discussion
61
bioreactor. One of the most important factors governing the yield of recombinant
proteins is sub-cellular targeting, which affects the interlinked processes of folding,
assembly and post-translational modification. The oxidizing environment of the
endoplasmic reticulum, the lack of proteases and the abundance of molecular
chaperones are important factors for correct protein folding and assembly. Also, protein
glycosylation occurs only in the endoplasmic membrane system and this modification is
required for correct function of many proteins. In the absence of further targeting
information, proteins in the endoplasmic membrane system are secreted to the apoplast,
where they might be retained or secreted into the external environment. The protein is
retained in the ER lumen using an H/KDEL C-terminal tetra-peptide tag, as this
compartment has a stabilizing influence. Retention of proteins in endoplasmic
reticulum cause increase in expression and immunogenicity of glycosylated rabies
glycoprotein (Yadav et al., 2013).
Arango et al. (2008) reported CaMV35S promoter regulated rabies N protein
antigen expression in tomato plants and performed mice immuno-protection assay.
Only intra peritoneal immunized mice showed weak protection against virus challenge
while the orally immunized mice were not protected. This suggested that the lack of
mucosal adjuvant in oral administration of antigen as the possible reason for diminished
immune response. Thus, the development of appropriate formulations for rapid
absorption through buccal mucosa is needed for mucosal vaccination. The
term mucosal vaccination has traditionally been used to describe strategies in which a
vaccine is administered via the mucosal route. Unlike parenteral vaccination, mucosal
vaccines do not require the use of needles, thus enabling vaccine compliance and
reducing logistical challenges and the risks of acquiring blood borne infections. The
great success of mucosal vaccines such as the polio vaccine which produce effective
elicitation of immunity against pathogens has popularised the concept.
This study is mainly focused on plant produced rabies glycoprotein which
fused with mucosal adjuvants and evaluates their immunogenic property after the N-
and C-terminal fusion of mucosal adjuvants. There are various lectins or lectin-like
proteins which have binding ability to glycolipids or glycoproteins (De Aizpurua and
Russell-Jones, 1988). Many of these proteins have already characterized and used as
mucosal adjuvant which stimulates strong humoral as well as cell-mediated immune
Page 123
Discussion
62
responses. Out of which cholera toxin B subunit is one of the most characterized
mucosal adjuvant which provides N-terminal fusion capability with target antigens.
CTB binds to the GM1 receptor in its pentamer confirmation and can serve as a
mucosal adjuvant from very long-time (McKenzie et al., 1984). Efficient binding to
GM1 could potentially increase the uptake of antigen across the mucosa and lead to an
enhanced presentation of the conjugated molecule to the immune system (Holmgren et
al., 1975; Nashar et al., 1996). CTB induces major histocompatibility complex (MHC)
class II expression on B cells (Francis et al., 1992), and also enhances antigen
presentation by macrophages in the absence of enhanced MHC II expression (Nashar et
al., 1996). A number of proteins have been genetically fused to the C-terminus of the
CTB and expressed in different plant tissues in earlier studies. For instance, human
insulin expressed in potato tuber and leaf (Arakawa et al. 1998), rotavirus enterotoxin
protein (NSP4) (Arakawa et al.2001; Kim and Langridge 2003) and anthrax lethal
factor protein (LF) (Kim et al. 2004) in potato tuber, B chain of human insulin (InsB3)
in tobacco leaf (Li et al. 2006) and surface protective antigen (SpaA) of Erysipe-lothrix
rhusiopathiae in tobacco hairy root (Ko et al. 2006). In each case, the fusion protein
retained their functional activity with respect to pentamerization and GM1 binding. The
stable pentamer formation of the CTB-RGP fusion protein in tobacco leaves was
confirmed by Western blot analysis and the expected ~403 kDa (pentamer of ~14.6 kDa
glycosylated CTB + ~66.0 kDa glycosylated RGP) band (figure 4.12A) was observed.
The expression of CTB-RGP protein in different transgenic plants showed wide
variation, i.e. 0.002–0.4% of TSP in leaf (figure 4.10A). This is consistent with the
direct GM1 ELISA result which done by both the antibodies Ab1 and Ab2 (figure
4.9A). The variation in expression is probably due to position effect in which
integration of the transgene occur at different positions in the genome of independent
transgenic lines (Peach and Velten, 1991). The ability of CTB-RGP fusion protein to
induce immune system through mucosal route is validated by immunization experiment
(figure 4.14).
Similarly, ricin B chain was also used here for the C- terminal fusion partner for
RGP which could facilitate its purification and its application as an oral vaccine. RTB
works as mucosal adjuvant and binds to lactose receptors that are present on mucosal
lining of gut epithelium (Sandvig et. al., 1976). It has been previously demonstrated
Page 124
Discussion
63
that intranasal administration of a recombinant fusion protein of ricin B chain (RTB)
with NSP4 rotavirus antigen purified from E. coli was highly effective in stimulating
mucosal antibody response in mice (Choi et al., 2006). Interaction between sugar-
binding moieties on the ricin B chain and terminal galactosides located on the
enterocyte membrane facilitates ricin holotoxin uptake by endocytosis into intracellular
vesicles (Lambert et al., 1991). The sugar-binding activity of the Ricin B chain is
largely dependent on one of three lactose-binding sites (Vitetta and Yen, 1990; Sphyris
et al., 1995; Swimmer et al., 1992; Frankel et al., 1996; Steeves et al., 1999). N-
terminus of RTB is located far from the membrane receptor-binding sites than the C-
terminus and that may permit C-terminal fusion with larger antigen molecules without
generating any problems of steric hindrance (Medina-Bolivar et al., 2003). Woffenden
et al. (2008) demonstrated that RTB fused with F1 and V antigens of Yersinia pestis
expressed in tobacco hairy roots has shown potential for protection of animals and
humans against bubonic and pneumonic plague. The small amount of protein required
for internalization and administration of fusion protein also improves the cost efficiency
of the product due to higher number of receptor per cell than GM1.
Asialofeutin and GM1 mediated direct ELISA with two different antibodies,
one is equine derived anti-rabies polyclonal and other is rabbit derived peptidal
antibody for specific immuno protective epitope region of rabies glycoprotein showed
equal competence. Variation in recognition of both the antibodies for CTB-RGP and
RGP-RTB fusion protein can be easily understood by different coverage of epitope
regions which rationalize the deviation and explain the functional validation of this
plant expressed protein to animal immune system.
Fusions of both the mucosal adjuvants with rabies glycoprotein were separated
by two repeats of glycine-proline hinge region which reduced steric hindrance and
helped in independent folding of each subunit of both the fusion proteins (Arakawa et
al., 1998). RTB monomers have the ability to bind its receptors from any of the three
available receptor binding sites which gives more opportunity of internalization through
mucosal lining, subsequently their presentation to immune system being 103
times
higher than conventionally used CTB pentamer fusion systems. However, ~168 kDa
dimerization of RGP-RTB fusion protein exhibits a structural integrity and stability of
RTB (figure 4.12B) which also complements the result of asialofeutin mediated direct
Page 125
Discussion
64
ELISA (figure 4.9B). RTB presents each molecule of fusion protein to antigen
presenting cell (APC) more readily which also enhances the immune response for
fusion protein than CTB that reflects in the antibody titre assay of the animal
experiment in which IgG1 and IgG2a response is higher in RGP-RTB than CTB-RGP
fusion proteins, however low immune response indicates the need of further
optimization of oral doses with higher concentration (figure 4.14). It could only be
possible by developing technologies like hairy root which can scalable the plant tissues
and produce large quantities of these proteins.
Superimposition of RGP (Figure 4.18) indicated that Gly-Pro hinge plays an
important role in confirmation of both the fusion proteins so that domains of mucosal
adjuvants did not get involved with tertiary folding of rabies glycoproteins. The
important antigenic epitopes and active sites which are crucial for structural
confirmation and stability of rabies glycoprotein (RGP) are shown in CTB-RGP and
RGP-RTB fusion proteins during in-silico analysis (Figure 4.19). Hopp-Wood
hydrophilicity and surface exposure plot of CTB-RGP and RGP-RTB proteins (Figure
4.20) complements the prediction of tertiary structure for both the fusion proteins.
However, Asialofeutin and GM1– receptor mediated direct ELISA with peptidal anti-
rabies antibody (Ab2) which rose against the D epitopic region provides a direct
evidence for in-silico analysis.
For RGP-RTB fusion protein, hairy root culture system was opted over the
transgenic plant leaves due to its ability of expression and accumulation of higher
amount of fusion proteins (figure 4.3B, 4.6), which in turn fulfil our aim to validate
proof of concept and obtaining a continuous biomass through bioreactor for further
experiments. Use of Solanum lycopersicum for production of foreign proteins through
hairy roots cultures in vitro has an advantage that it does not produce any hazardous
alkaloid, unlike tobacco. The possibility of any such alkaloid contamination in oral
application with this chimeric fusion protein is extremely remote. These features may
be helpful in the context of acquiring regulatory approval for proteins produced through
hairy roots of Solanum lycopersicum in future.
A variety of approaches may be used for long term maintenance of transgenic
hairy roots involved in foreign protein production. Maintenance of transgenic plants in
a bio-contained green house or as axenic plantlets in vitro may also represent a feasible,
Page 126
Discussion
65
cheap and technologically simple way to maintain germplasm for species which readily
regenerate plants from hairy roots (Christey, 2001). Further work has also shown that
hairy roots can also be established from offspring of these transgenic plants (De
Guzman et al., 2011) allowing the possibility of long term storage of therapeutic protein
producing hairy root germplasm in the form of seeds. Use of bioreactor for the
production of secondary metabolites such as gossypol (Verma et al., 2009),
isoflavonoid (Udomsuk et al., 2009) and withanolid (Mirjalili et al., 2009) and
important pharmaceutical proteins like human interferon alpha-2b (Luchakivskaia et al.
2012), P450 (Banerjee et al., 2002), mouse IL-12 (Liu et al., 2009) have successfully
established the utility of this system.
The principle cost of most commercial vaccines is in the form of production,
packaging and delivery. Injectable vaccines incur further expenses related to the use
and disposal of needles and syringes, trained personnel to administer injections, and
refrigeration required during shipping and storage. These factors prevent widespread
vaccination of livestock, poultry and swine against preventable diseases. Producing this
fusion protein in hairy root has shown equal competence to overcome all the above
problems. The RGP-RTB fusion protein expresses up to 1.1 % of TSP (figure 4.10) in
hairy roots which accounts nearly 8 µg of total protein in per gram of tissue,
optimization of air lift bioreactor to produce nearly 108 times of initial biomass of hairy
roots as achieved in the present study is sufficient to furbish future oral immunization
goal against rabies.
A critical review of the published reports indicates that the use of mechanically
agitated reactors with immobilization structures is not useful (Verma et al., 2003). In
this study the immobilization structure had been used to isolate the roots from the
stirrer mechanism in order to avoid shear damage of the roots by direct contact with the
impeller. The use of isolated impeller reactors with the nylon mesh had proved
effective for hairy root clone H03 for better growth, immobilization and uniform
distribution of the root tissues through-out the entire bioreactor vessel. The velocity of
the bulk fluid in the reactor vessel and the penetration of convection currents into the
root mass determine the rate of oxygen and nutrient uptake by hairy roots (Flores and
Curtis, 1992). Optimization of the air flow rate and agitation speed were found critical
for efficiently directing the flow of the media through the compact root tissues due to
Page 127
Discussion
66
its powerful axially directed volumetric flow and resulted in comparable biomass as in
the shake flask condition. This study once again reiterates the fact that for optimization
of bioreactor performance, the morphological and physiological properties of the
specific hairy root clone of any particular plant system need to be taken into
consideration as these characters strongly influence the final productivities of the
concerned root clone, both in terms of biomass as well as the recombinant protein of
interest.
Page 128
Chapter 6
Summary and Conclusions
Page 129
Summary and Conclusions
67
6.0 SUMMARY AND CONCLUSIONS
6.1 Summary
Recombinant subunit vaccines are desirable as an alternative with potentially
fewer side effects than delivering the whole organism. Because recombinant subunit
vaccines do not contain an infectious agent, they are safer to administer, prepare and
doses are more uniform. Recombinant vaccines have a potential for being highly
effective in preventing disease, both in humans and animals, but are rather costly to
produce, and therefore are in limited use worldwide. However, the use of plants as
bioreactors is a solution to the current manufacturing bottleneck that is faced by the
pharmaceutical and biotechnology industries. Plants are used as the production units
(i.e. bioreactor, farms or greenhouses) and can be economically expanded or reduced
depending on the demand for a specific plant-derived drug. Rabies is an acute
contagious infection of the central nervous system caused by rabies virus.
Approximately 60,000 human deaths occur worldwide annually from rabies (Meslin
and Stohr, 1997). Plant produced recombinant subunit vaccine of rabies glycoproteins
has shown remarkable promise in protecting animals against rabies, but has lesser
expression and insufficient for purification and mass immunization task for animals and
humans. Contamination like plant pigments and low expression are the major limiting
factors for producing pharmaceutical proteins in plants. This could be avoided by oral
immunization of plant produced vaccines antigens with mucosal adjuvants.
There are various lectins or lectin-like proteins which have binding ability to
glycolipids or glycoprotein. Many of these lectins have already been characterized and
used as mucosal adjuvant which stimulate strong humoral as well as cell-mediated
immune responses. We utilize these lectins for administration of rabies antigen orally to
avoid near homogeneity purification and to enhance immunogenic property of rabies
antigen. It was previously shown that intranasal administration of an antigen which
fused with the mucosal adjuvant and derived from transgenic plants were found highly
effective in stimulating a mucosal antibody response in mice. We have employed
strategies like N- or C- terminal genetically fused adjuvant to rabies glycoprotein for
the assessment of their role in immunization efficacy and internalization ability.
The pentameric B subunit of cholera toxin (CTB) is an efficient N-terminal
mucosal adjuvant for vaccines antigen. We report the expression of a chimeric protein
Page 130
Summary and Conclusions
68
comprising the synthetic cholera toxin B subunit fused at its C-terminal with rabies
surface glycoprotein (G protein) in tobacco plants. The ~80.3 kDa fusion polypeptide
expressed at 0.4% of the total soluble protein in leaves of the selected transgenic lines.
The fusion protein formed a ~403 kDa pentameric protein which was functionally
active in binding to GM1 receptor. The plant-made protein had a higher affinity for
GM1 receptor than the native bacterial CTB. The pentameric fusion protein was
recognized by the anti-rabies antibodies.
Similarly CTB, RTB act as C-terminal mucosal adjuvant and more efficiently
binds to its asialofeutin receptor. However, transgenic tobacco and hairy roots of
Solanum lycopersicum are generated to express a recombinant protein containing a
fusion of rabies glycoprotein and ricin toxin B chain (rgp-rtxB) antigen under the
control of constitutive CaMV35S promoter. Expression of RGP-RTB fusion protein in
transgenic tomato hairy roots is more than the leaves of transgenic tobacco for the same
construct. Hairy root culture shows higher expression and accumulation of RGP-RTB
fusion proteins. Asialofetuin mediated direct ELISA of transgenic hairy root extracts is
performed by using polyclonal anti-rabies glycoprotein (ab1) antibodies and epitope
specific peptidal anti- rabies glycoprotein (ab2) antibodies confirmed expression of
functionally viable RGP-RTB fusion protein. ELISA based on asialofeutin binding
activity was used to screen crude hairy root protein extracts from five transgenic lines.
Expressions of RGP-RTB fusion protein in different tomato hairy root lines vary
between 1.4 to 8µg in per gram of tissue. Immuno blotting assay from hairy root lines
detected ~84kDa protein on denaturing gel which is as per the predicted size of RGP-
RTB fusion protein monomer and ~168kDa protein dimer was also detected on a native
gel. Out of five, one line H03 showed highest level of RGP-RTB protein expression.
Regeneration of tomato hairy roots line H03 is readily achieved and used further in
bench top bioreactor for further optimization and scale up process to produce large
quantities of recombinant proteins. Bioreactor grown hairy roots showed similar growth
pattern and total protein content as in the shake flask grown hairy root. Large quantities
of fusion proteins will be needed to initialize for intranasal immunization of animal
experiment and further evaluation of this recombinant protein for use as oral vaccine.
The important antigenic epitopes and active sites which are crucial for structural
confirmation and stability of rabies glycoprotein (RGP) were studied in CTB-RGP and
Page 131
Summary and Conclusions
69
RGP-RTB fusion proteins during in-silico analysis which showed equal competence for
stability and immunogenicity of predicted tertiary structure for both the fusion proteins.
Primary immunization experiment induces less immune response probably due to the
quantitatively less oral doses but both the fusion protein remarkably induce Th1 and
Th2 responses, even in the lower administered doses. So detailed immunization
experiment should be performed after scaling up our system which may lead to the
foundation of commercialization and mass immunization aspect. Hence, this study is a
first step towards an improved oral vaccine and provides proof of concept for oral
immunization against rabies.
We have not only successfully employed strategies like N- or C- terminal
genetically fusion of adjuvants to rabies glycoprotein but have also scale them up to
perform animal experiment for fulfillment of commercialization aspect. Compared to
traditional vaccines, oral vaccines offer simplicity of use, lower cost, convenient
storage, economic delivery and mucosal immune response. This study can solve the
challenge of rabies prevention by establishment of low-cost alternatives for effective
oral vaccine that provides lasting protection after immunization and is conveniently or
readily available for frequent administration to regions where rabies is endemic.
Page 132
Summary and Conclusions
70
6.2 Conclusions
1. Oral vaccines expressed in plant can offer simplicity of use, lower cost,
convenient to store and give systemic immune response.
2. It can also provide needle-free vaccination and economical delivery system
against rabies.
3. RGP maintained their conformational epitopes and are immunogenically active
in both the fusion proteins.
4. Mucosal adjuvants in both CTB-RGP and RGP-RTB fusion protein attain their
native structure and stably bind to its receptors.
5. Fusion of mucosal adjuvant to rabies glycoprotein enhanced the stability of
expressed vaccine antigens in plant tissues which can be of advantage in
development of formulations for improve immunogenicity and rapid absorption
through mucosal route.
6. RGP-RTB fusion protein induces more immunogenic response than CTB-RGP,
since its monomer has three receptor binding sites and can readily bind to its
receptors.
7. Standardization of dosage for oral vaccination against rabies is needed as plant
produced fusion proteins shows immunogenicity but lower Th responses.
8. The selection of suitable plant tissue for expression and isolation of fusion
protein, ER targeting signal sequence, ER retention sequence, plant preferred
codon usage, protein stabilization, post translational modification etc. will lead
to increase in the accumulation of fusion proteins in plant tissues.
9. Human pathogenic contamination free preparations can be made and used
through oral route. These strategies are more conveniently amenable to
development of GLP and GMP protocols.
10. Expression of RGP-RTB fusion protein in transgenic tomato hairy roots is more
as compared to the leaves of transgenic tobacco for the same construct, probably
due to lower background proteins, pigment concentration and higher growth
rate.
11. Hairy root cultures are genetically stable and show clonal fidelity which in turn
provides stable long term product profile or similar expression.
Page 133
Summary and Conclusions
71
12. Hairy root cultures have greater autonomy for auxin biosynthesis which in turn
gives faster growth rate in comparison to the whole plant.
13. Hairy root cultures have minimum nutritional requirements which reduce the cost
of in-vitro culture practices and give scalability option in fermenter based
systems for industrial application.
14. Issues relating to the ethical, social, biosafety, environmental impact and
RCGM approval which directly or indirectly effect the deployment of
genetically modified plants are very much addressed by using hairy root system.
15. The plant based oral vaccine technology might be first targeted to wild and
domesticated animals to gain experience which in turn will benefit agricultural
productivity and will provide easy vaccination option for farm animals and
poultry.
In view of above, this study checks the efficacy of plant produced rabies
glycoprotein with N- and C-terminally fused mucosal adjuvants and provides relevant
information for developing plant based oral vaccine against rabies.
Page 135
Bibliography
72
BIBLIOGRAPHY
Ahuja S., Tripathi K.K., Saha S.M. and Saxena S.N. (1983). Epidemiology of
Rabies in India. Proceedings of an international conference on rabies control in
the tropics. Tunis. Oct 3-6, 1983.
Alvarez M.L., Pinyerd H.L., Crisantes J.D., Rigano M.M., Pinkhasov J. and
Walmsley A.M. (2006). Plant-made subunit vaccine against pneumonic and
bubonic plague is orally immunogenic in mice. Vaccine 24: 2477–90.
Anilionis A., Wunner W.H. and Crutis P.J. (1981). Structure of the glycoprotein
gene in rabies virus. Nature 294: 275-278.
Arakawa T., Chong D., Merritt J. and Langridge W. (1997). Expression of cholera
toxin B subunit oligomers in transgenic potato plants. Trans. Res. 6: 403-413.
Arakawa T., Yu J., Chong D.K., Hough J., Engen P.C. and Langridge W.H.
(1998). A plant-based cholera toxin B subunit-insulin fusion protein protects
against the development of autoimmune diabetes. Nat. Biotechnol. 16: 934–938.
Arakawa T., Chong D., Langridge W. (1998a). Efficacy of a food plant based oral
cholera toxin B subunit vaccine. Nat. Biotechnol. 16: 292-297.
Arakawa T., Yu J. and Langridge W.H. (2001). Synthesis of a cholera toxin B
subunit-rotavirus NSP4 fusion protein in potato. Plant Cell Rep. 20: 343–348
Arango I.P., Rubio E.L., Anaya E.R., Flores T.O., de la Vara L.G. and Lim
M.A.G. (2008). Expression of the rabies virus nucleoprotein in plants at high-
levels and evaluation of immune responses in mice. Plant Cell Rep. 27: 677–685
Areas A.P.M., Oliveira M.L.S., Miyaji E.N., Leite L.C.C., Aires K.A., Dias W.O.
and Ho P.L. (2004). Expression and characterization of cholera toxin B-
pneumococcal surface adhesion A fusion protein in Escherichia coli: ability of
CTB-PsA to induce humoral immune response in ice. Biochem. Biophys. Res.
Commun. 321: 192–196
Arlen P.A., Singleton M., Adamovicz J.J., Ding Y., Davoodi-Semiromi A. and
Daniell H. (2008). Effective plague vaccination via oral delivery of plant cells
expressing F1-V antigens in chloroplasts. Infect. Immun. 76: 3640–50.
Page 136
Bibliography
73
Artois M., Guittre I., Thomas I., Leblois H., Brochier B. and Barrat J. (1992).
Potential pathogenicity for rodents of vaccines intended for oral vaccination
against rabies: a comparison. Vaccine 10: 524-528.
Ashraf S., Singh P.K., Yadav D.K., Shahnawaz M., Mishra S., Sawant S.V. and
Tuli R. (2005). High level expression of surface glycoprotein in tobacco and its
immuno-protective activity in mice. J. Biotechnol. 119: 1-14.
Athmaram T.N., Bali G. and Devaiah K.M. (2006). Integration and expression of
Bluetongue VP2 gene in somatic embryos of peanut through particle
bombardment method. Vaccine 24: 2994–00.
Aziz M.A., Singh S., Kumar P.A., Bhatnagar R. (2002). Expression of protective
antigen in transgenic plants: a step towards edible vaccine against anthrax.
Biochem. Biophys. Res. Commun. 299: 345–51.
Badrane H., Bahloul C., Perrin P. and Tordo N. (2001). Evidence of two Lyssavirus
phylogroups with distinct pathogenicity and immunogenicity. J. Virology 75:
3268-3276.
Baer G., Shaddock J., Levy H. and Johnston P. (1988). Interferon in post-exposure
prophylaxis. In “Progress in rabies control” (Traenhart O., Koprowski H., Bogel
K. and Sureau, P. Eds), Wells Medical: 245-250.
Baer G.M. and Lentz T.L. (1991). Rabies Pathogenesis to the central nervous system.
In: The Natural History of Rabies Ed: Baer G.M., Boca Raton F.L CRC Press:
105-120.
Bahloul C., Ahmed S.B.H., B’ chir B.I., Kharmachi H., Hayouni E.A. and Dellagi
K. (2003). Post exposure therapy in mice against experimental rabies: a single
injection of DNA vaccine is as effective as five injections of cell culture-derived
vaccine. Vaccine 22: 177-184.
Bahloul C., Jacob Y., Tordo N. and Perrin P. (1997). DNA-based immunization for
exploring the enlargement of immunological cross-reactivity against the
lyssaviruses. Vaccine 16: 417-425.
Page 137
Bibliography
74
Bai X., Warner C.K. and Fekadu M. (1993). Comparisons of nucleotide and deduced
amino acid sequences of the glycoprotein genes of a Chinese street strain
(CGX89-J) and a Chinese vaccine strain (3a G) of rabies virus. Virus Res. 27:
101-112.
Bailey M.J., Mcleod D.A., Kang C.-Y., Bishop D.H.L. (1989). Glycosylation is not
required for the fusion activity of the G protein of vesicular stomatitis virus in
insect cells. Virology 169: 323-331.
Banerjee S., Shang T.Q., Wilson A.M., Moore A.L., Strand S.E., Gordon M.P. and
Doty S.L. (2002). Expression of functional mammalian P450 2E1 in hairy root
cultures. Biotechnol. Bioengin. 77: 462–466.
Baumann O., Walz B., Somlyo A.V. and Somlyo A.P. (1991). Electron probe
microanalysis of calcium release and magnesium uptake by endoplasmic
reticulum in bee photorceptors. Proc. Natl. Acad. Sci. U.S.A. 88: 741-744.
Benmansour A., Leblois H., Coulon P., Tuffereau C., Gaudin Y., Flamand A. and
Lafay F. (1991). Antigenicity of rabies virus glycoprotein. J. Virol. 65: 4198-
4203.
Bergmann J.E., Tokuyasu K.T. and Singer S.J. (1981). Passage of an integral
membrane protein, the vesicular stomatitis virus glycoprotein, through the Golgi
apparatus en route to the plasma membrane. Proc. Natl. Acad. Sci., USA. 78:
1746-1750.
Bernnan F.R., Jones T.D., Gilleland L.B., Bellaby T., Xu F., North P.C.,
Thompson A., Staczek J. and Lin T. Johnson J.E. (1999). Pseudomonas
aeroginosa outer–membrane protein F epitopes are highly immunogenic in mice
when expressed on a plant virus. Microbiol. 145: 211-220.
Biemelt S., Sonnewald U., Galmbacher P., Willmitzer L., Müller M. (2003).
Production of human papillomavirus type 16 virus-like particles in transgenic
plants. J. Virol. 77: 9211–20.
Bourhy H., Kissi B. and Tordo N. (1993). Molecular diversity of the Lyssavirus
genus. Virology 194: 70-81.
Page 138
Bibliography
75
Bourhy H., Kissi B., Audry L. and Smreczak M. (1999). Ecology and evolution of
rabies virus in Europe. Journal of General Virology 80: 2545-2557.
Bourhy H., Kissi B., Lafon M., Sacramento D. and Tordo N. (1992). Antigenic and
molecular characterization of bat rabies virus in Europe. J. Clin. Microbiol. 30:
2419-2426.
Broughan J.H. and Wunner W.H. (1995). Characterization of protein involvement in
rabies virus binding to BHK-21 cells. Arch. Virol. 140: 75-93.
Bunschoten H., Gor M., Claassen I.J.T.M., Uytedehaag F.G.C.M., Dietzschold B.,
Wunner W.H. and Osterhaus A.D.M.E. (1989). Characterization of a new
virus-neutralizing epitope that denotes a sequential determinant on the rabies
virus glycoprotein. Journal of General Virology 70: 291-298.
Burger S.R., Remaley A.T., Danley J.M., Moore J., Muschel R.J., Wunner W.H.
and Spitalnik S.L. (1991). Stable expression of rabies virus glycoprotein in
Chinese hamster ovary cells. Journal of General Virology 72: 359-367.
Burrage T.G., Tiqnor G.H. and Smith A.L. (1985). Rabies virus binding at
neuromuscular junctions. Virus Res. 2: 273-289.
Cadoz M., Strady A., Meignier B., Taylor J., Tartaglia J., Paoletti E. and Plotkin
S. (1992). Immunisation with canarypox virus expressing rabies glycoprotein.
The Lancet 339: 1429-1432.
Cangelosi G.A., Best E.A., Martinetti G. and Nester E.W. (1991). Genetic analysis
of Agrobacterium. Methods Enzymol. 204: 384-97.
Carrillo C., Wigdorovitz A., Oliveros J.C., Zamorano P.I., Sadir A.M. and Gomez
N. (1998). Protective immune response to foot-and-mouth disease virus with VP1
expressed in transgenic plants. J. Virol. 72: 1688–90.
Carrillo C.,Wigdorovitz A., Trono K., Dus Santos M.J., Castañón S. and Sadir
A.M. (2001). Induction of a virus-specific antibody response to foot and mouth
disease virus using the structural protein VP1 expressed in transgenic potato
plants. Viral Immunol. 14:49–57.
Page 139
Bibliography
76
Castañón S., Marín M.S., Martín-Alonso J.M., Boga J.A., Casais R., Humara J.M.
and Ordás R.J. (1999). Immunization with potato plants expressing VP60
protein protects against rabbit hemorrhagic disease virus. J. Virol. 73: 4452–5.
Castañón S., Martín-Alonso J.M., Marín M.S., Boga J.A., Alonso P. and Parra F.
(2002). The effect of the promoter on expression of VP60 gene from rabbit
hemorrhagic disease virus in potato plants. Plant Sci. 162:87–95.
Ceccaldi P.E., Gillet J.P. and Tsiang H. (1989). Inhibition of the transport of rabies
virus in the central nervous system. J. Neuropathol. Exp. Neurol. 48: 620-630.
Celis E., Ou D.W., Dietzschold B. and Koprowski H. (1988). Recognition of rabies
and rabies-related viruses by T cells derived from human vaccine recipients. J.
Virol. 62: 3128-3134.
Celis E., Ou D., Dietzschold B., Otvos L. and Koprowski H. (1989). Rabies virus
specific T cell hybridomas: identification of class II MHC-Restricted T-cell
epitopes using synthetic peptides. Hybridoma 8: 263-275.
Charlton K.M. (1994). The pathogenesis of rabies and other lyssaviral infections:
recent studies. In: “Lyssaviruses”, Ruprecht C.E., Dietzschold B. and Koprowski
H.( Eds), Springer- Verlag, Berlin, Heidelberg, New York. 95-120.
Charlton K.M. and Casey G.A. (1979). Experimental rabies in skunks. Immuno-
fluorescence light and electron microscopic studies. Lab. Inves. 41: 36-44.
Charlton K.M., Nadin-Davis S., Casey G.A. and Wandeler A.I. (1997). The long
incubation period in rabies: Delayed progression of infection in muscle at the site
of exposure. Acta. Neuropathol. 94: 73-77.
Chazaud B., Muriel M.P., Wantyghem J., Aubery M. and Decastel M. (1995).
Ricin toxicity and intracellular routing in tumoral HT-29 cells. Exp. Cell. Res.
221: 214–220.
Chenik M., Chebli K., Gaudin Y. and Blondel D. (1994). In vivo interaction of
rabies virus phosphoprotein (P) and nucleoprotein (N): existence of two N-
binding sites on P protein. Journal of General Virology. 75: 2889-2896.
Page 140
Bibliography
77
Chikwamba R., Cunnick J., Hathaway D., McMurray J., Mason H. and Wang
K.A. (2002). Functional antigen in a practical crop: LT-B producing maize
protects mice against Escherichia coli heat labile enterotoxin (LT) and cholera
toxin (CT). Transgenic Res. 11: 479 93.
Childs J.E., Cums A.T. and Dey M.E. (2000). Predicting the local dynamics of
epizootic rabies among raccoons in the United States. Proc. Natl. Acad. Sci.,
U.S.A. 97: 13666-13671.
Choi N.W., Estes M.K. and Langridge W.H. (2006). Mucosal immunization with a
ricin toxin B subunit-rotavirus NSP4 fusion protein stimulates a Th1 lymphocyte
response. J. Biotechnol. 121: 272-83.
Christey M.C. (2001). Transgenic crop plants using Agrobacterium rhizogenes
mediated transformation. In: Doran P.M. (Ed.), Hairy roots: culture and
application. Amsterdam: Harwood Academic. 99-111.
Clague M.J., Schoch C., Zech L. and Bluementhal R. (1990). Gating kinetics of pH-
activated membrane fusion of vesicular stomatitis virus with cells: stopped-flow
measurements by sequencing of octadecylrhodamine fluorescence. Biochem. 29:
1303-1308.
Conti C., Superti F. and Tsiang H. (1986). Membrane carbohydrate requirement for
rabies virus binding to chicken embryo related cells. Intervirol. 26: 164-168.
Conzelmann K.K., Cox J.H., Schneider L.G. and Thiel H.I. (1990). Molecular
cloning and complete nucleotide sequence of attenuated rabies virus SAD B19.
Virology 75: 485-499.
Coslett D.G., Hollow B.P., Obijeski J.K. (1980). The structural proteins of rabies
virus and evidence for their synthesis from separate monocistronic RNA species.
J. Gen. Virol. 49: 161–180.
Coulon P., Derbin C., Kucera P., Lafay F., Prehaud C. and Flamand A. (1989).
Invasion of the peripheral nervous systems of adult mice by the CVS strain of
rabies virus and its avirulent derivative Av 01. J. Virol. 63: 3550-3554.
Page 141
Bibliography
78
Coulon P., Ternaux J.P., Flamand A. and Tuffereau C. (1998). An avirulent mutant
of rabies virus is unable to infect motoneurons in vivo and in vitro. J. Virol. 72:
273-278.
Cox J., Dietschold B. and Schneider L. (1977). Rabies virus glycoprotein: Biological
and serological characterization. Infect. Immun. 16: 754 -759.
Cox J., Dietzschold B., Weiland F. and Scheider L. (1980). Preparation and
characterization of rabies virus hemagglutinin. Infect. Immun. 30: 572-577.
Cox J.H., Dietzschold B. and Schneider L.G. (1977). Rabies virus glycoprotein II:
Biological and serological characterization. Infect. Immun. 16: 754–759.
Crick J. and King A. (1988). Culture of rabies virus in vitro. In: Rabies. Eds:
Campbell J.B. and Charlton K.M., Boston, Kluwer Academic. 47-66.
Cruz F.W., McBride A.J.A., Conceicao F.R., Dale J.W., McFadden J. and
Dellagostin O.A. (2002). Expression of the B-cell and T-cell epitopes of the
rabies virus nucleoprotein in Mycobacterium bovis BCG and induction of a
humoral response in mice. Vaccine 20: 731-736.
Curtiss R. and Cardineau G.A. (1990). Oral immunization by transgenic plants. World
Patent Application. WO 90/02484.
Curtiss R. and Cardineau G.A. (1997). Oral immunization by transgenic plants. US
Patent. 5: 686-079.
Da Silva J.V., Garcia A.B., Flores V.M.Q., de Macedo Z.S. and Medina-Acosta E.
(2002). Phytosecretion of enteropathogenic Escherichia coli pilin subunit A in
transgenic tobacco and its suitability for early life vaccinology. Vaccine 20:
2091–101.
Dalsgaard K., Uttenthal A., Jones T.D., Xu F., Merryweather A., Hamilton W.D.,
Langeveld J.P., Boshwizen R.S., Kamstrup S., Lomonossoff G.P., Porta C.,
Vela C., Casal J.L., Meloen R.H. and Rodgers P.B. (1997). Plant derived
vaccine protects target animals against a viral disease. Nat. Biotech. 15: 248-252.
Page 142
Bibliography
79
Daniell H., Streatfield S.J. and Wycoft K. (2001). Medical molecular farming
production of antibodies, biopharmaceuticals and edible vaccines in plants.
Trends plant Sci. 6: 219-226.
David D., Yakobson B., Smith J.S. and Stram Y. (2000). Molecular epidemiology of
rabies virus isolates from Israel and other middle and near-eastern countries. J.
Clinical Microbiol. 38: 755-762.
De Guzman G., Walmsley A.M., Webster D.E. and Hamill J.D. (2011). Hairy roots
cultures from different Solanaceous species have varying capacities to produce E.
coli B-subunit heat-labile toxin antigen. Biotechnol. Lett. 33: 2495-502.
De Mattos C.A., De Mattos C.C., Smith J.S., Miller E.T., Papo S., Utrera A. and
Osburn B.I. (1996). Genetic characterization of rabies field isolates from
Venezuela. J. Clinical. Microbiol. 34: 1553-1558.
De Aizpurua H.J. and Russell-Jones G.J. (1988). Oral vaccination: Identification of
classes of proteins that provoke an immune response upon oral feeding. J. Exp.
Med. 167: 440-51.
Dean D.J., Ablesetg M.K. and Atanasiu P. (1996). The fluorescent antibody test. In:
Laboratory Techniques in Rabies. Meslin F.X., Kaplan M.M. and Koprowski H.
(Eds.) Geneva, WHO. 88-95.
Delagneau J.F., Perrin P. and Atanasiu P. (1981). Structure of the rabies virus:
spatial relationships of the proteins G, M1, M2 and N. Ann. Virol. Inst. Pasteur
132: 473-493.
Dietzschold B., Wiktor T.J., Macfarlan R. and Varrichio A. (1982). Antigenic
structure of rabies virus glycoprotein: ordering and immunological
characterisation of large CNBr peptide fragments. J. Virol. 44: 595-602.
Dietzschold B., Wiktor T. J., Wunner W. and Varrichio A. (1983a). Chemical and
immunological analysis of the rabies soluble glycoprotein. Virology 124: 330-
337.
Dietzschold B., Wunner W., Wiktor T. J., Lopes D., Lafon M., Smith C. and
Koprowski H. (1983b). Characterization of an antigenic determinant of the
Page 143
Bibliography
80
glycoprotein that correlates with pathogenicity of rabies virus. Proc. Natl. Acad.
Sci., U.S.A. 80: 70-74.
Dietzschold B., Wiktor T.J., Trojanowski J.Q., Macfarlan R., Wunner W.H.,
Torres–Anjel M.J. and Koprowski H. (1985). Differences in cell-to-cell spread
of pathogenic and a pathogenic rabies virus in vivo and in vitro. J. Virol. 56: 12-
18.
Dietzschold B., Lafon M., Wang H., Otvos L., Celis E., Wunner W. and
Koprowski H. (1987a). Localization and immunological characterization of
antigenic domains of the rabies virus internal N and Ns proteins. Virus Research
8: 103-125.
Dietzschold B., Wang H., Rupprecht C.E., Celis E., Tollis M., Ertl H., Heber Katz
E. and Koprowski H. (1987b). Induction of protective immunity against rabies
by immunisation with rabies virus ribonucleoprotein. Proc. Natl. Acad. Sci.,
U.S.A. 84: 9165-9169.
Dietzschold B., Gore M., Ertl. H., Celis E., Otvos L. and Koprowski H. (1989).
Analysis of protective immune mechanisms induced by rabies nucleoprotein.
“Genetics and Pathogenicity of negative strand viruses”, In: Mahy B.W.J. and
Kolakofski D. (Eds.), Elsevier Science, New York. 295-309.
Dietzschold B., Gore M., Marchadier D., Niu H.-S., Bunschoten H.M., Otvos L.J.,
Wunner W.H., Ertl H.C.J., Osterhaus A.D.M.E. and Koprowski H. (1990a).
Structural and immunological characterization of a linear virus-neutralizing
epitope of the rabies virus glycoprotein and its possible use in synthetic vaccine.
J. Virol. 64: 3804-3809.
Dietzschold B., Gore M., Casali P., Ueki Y., Rupprecht C., Notkins A. and
Koprowski H. (1990b). Biological characterization of human monoclonal
antibodies to rabies virus. J. Virol. 64: 3087-3090.
Dietzschold B., Rupprecht C.E., Fu Z.F. and Koprowski H. (1996). Rhabdoviruses.
In Fields B.N., Knipe D.M. and Howley P.M. (eds.), Fields Virology III
Lippincott –Raven Press, Philadelphia, PA. 1137-1159.
Page 144
Bibliography
81
Doms R.W., Lamb R.A., Rose J.K. and Helenius A. (1993). Folding and assembly of
viral membranes proteins. Virology. 193:545-562.
Dong J.L., Liang B.G., Jin Y.S., Zhang W.J., Wang T. (2005). Oral immunization
with pBsVP6-transgenic alfalfa protects mice against rotavirus infection.
Virology 339: 153–63.
Doran P.M. (2000). Foreign protein production in plant tissue cultures. Curr. Opin.
Biotech. 11: 199-204.
During K., Porsch P., Fladung M., Lorz H. (1993). Transgenic potato plants resistant
to the phytopathogenic bacterium Erwinia carotovora. Plant J. 3: 587-598.
Durrani Z., Mclnerney T.L., McLain L., Jones T., Bellaby T., Bernnan F.R. and
Dimmock N.J. (1998). Intranasal immunization with a plant virus expressing a
peptide from HIV-1 gp41 stimulates better musoal and systemic HIV-1 specific
1gA and 1gG than oral immunization. J. Immunol. Method. 220: 93-103.
Durrer P., Gaudin Y., Ruigrok R.W.H., Graf R. and Brunner J. (1995).
Photolabeling identifies a putative fusion domain in the envelope glycoprotein of
rabies and vesicular stomatitis virus. J. Biol. Chem. 270: 17575-17581.
Dus Santos M.J., Wigdorovitz A., Trono K., Rios R.D., Franzone P.M., Gil F.
(2002). A novel methodology to develop a foot and mouth disease virus (FMDV)
peptide-based vaccine in transgenic plants. Vaccine 20: 1141–7.
Ehsani P., Khabiri A. and Domansky N.N. (1997). Polypeptides of hepatitis B
surface antigen produced in transgenic potato. Gene 190: 107–11.
Ellis R.J. and Hemmingsen S.M. (1989). Molecular chaperons: proteins essential for
the biogenesis of some macromolecular structures. Trends Biochem. Sci. 14: 339-
342.
Endo Y. and Tsurugi K. (1988). The RNA N-glycosidase activity of ricin A-chain. J
Biol. Chem. 263: 8735–8739.
Ertl H., Dietzschold B. and Otvos L. (1991). T helper epitope of rabies virus
nucleoprotein defined by tri and tetrapeptides. Eur. J. Immunol. 21:1-10.
Page 145
Bibliography
106
Rupprecht C.E., Wiktor T.J., Johnston D.M., Hamir A.N., Dietzschold B.,
Wunner W.H., Glickman L.T. and Koprowski H. (1986). Oral immunization
and protection of raccoons (Procyon lotor) with a vaccinia-rabies glycoprotein
recombinant virus vaccine. Proc. Natl. Acad. Sci., U.S.A. 83: 7947-7950.
Sabeta C.T., Bingham J. and Nel L.H. (2003). Molecular epidemology of canid
rabies in Zimbabwe and South Africa. Virus Research 91: 203-211.
Sacramento D., Badrane H., Bourhy H. and Tordo N. (1992). Molecular
epidemiology of rabies virus in France: comparison with vaccine strains. Journal
of General Virology 73: 1149-1158.
Sakamoto S., Ide T., Tokiyoshi S., Nakao J., Hamada F., Yamamoto M., Grosby
J.A., Ni Y. and Kawai A. (1999). Studies on the structures and antigenic
properties of rabies virus glycoprotein analogues produced in yeast cells. Vaccine
17: 205-218.
Sambrook J., Fritsch E.F. and Maniatis T. (1989). Molecular Cloning: A Laboratory
Mannual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Schneider L.G., Horzinek M. and Matheka H.D. (1971). Purification of rabies virus
from tissue culture. Arch. Gesamte. Virusforsch. 34: 351-359.
Sandhu J.S., Krasnyanski S.F., Domier L.L., Korban S.S., Osadjan M.D. and
Buetow D.E. (2000). Oral immunization of mice with transgenic tomato fruit
expressing respiratory syncytial virus- F protein induces a systemic immune
response. Transgenic Res. 9: 127–35.
Sandvig K., Olsnes S. and Pihl A. (1976). Kinetics of binding of the toxic abrin and
ricin to surface receptors of human cells. J. Biol. Chem. 251, 3977–3984
Schneider L.G., Dietzschold B., Dierks R.E., Matthaeus W., Enzmann P.J. and
Strohmaier K. (1973). Rabies group- specific ribonucleoprotein antigen and a
test system for grouping and typing of rhabdoviruses. J. Virol. 11: 748-755.
Schneider L.G. and Schoop U. (1972). Pathogenesis of rabies and rabies like viruses.
Ann. Institute Pasteur (Paris). 123: 469-476.
Page 146
Bibliography
118
Yadav D.K., Ashraf S., Singh P.K. and Tuli R. (2012). Localization of rabies virus
glycoprotein into the endoplasmic reticulum produces immunoprotective antigen.
Protein J. 31: 447-56.
Yang J., Hooper D.C., Wunner W.H., Koprowski H., Dietzschold B. and Fu Z.F.
(1998). The specificity of rabies virus RNA encapsidation by nucleoprotein.
Virology 242: 107-117.
Yang Z.Q., Liu Q.Q., Pan Z.M., Yu H.X. and Jiao X.A. (2007). Expression of the
fusion glycoprotein of Newcastle disease virus in transgenic rice and its
immunogenicity in mice. Vaccine 25: 591-8.
Yang Y., Li X., Yang H., Qian Y., Z hang Y., Fang R. and Chen X. (2011).
Immunogenicity and virus-like particle formation of rotavirus capsid proteins
produced in transgenic plants. Sci. China Life Sci. 54: 82-9.
Yelverton E., Norton S., Obijeski J.F. and Goeddel D.V. (1983). Rabies virus
glycoprotein analogs: biosynthesis in Escherichia coli. Science 219: 614-620.
Youma J.W., Won Y.S., Jeon J.H., Ryu C.J., Choi Y.K. and Kim H.C. (2007). Oral
immunogenicity of potato-derived HBsAg middle protein in BALB/c mice.
Vaccine 25: 577–84.
Youma J.W., Jeon J.H., Kim H., Kim Y.O., Ko K. and Joung H. (2008). Transgenic
tomatoes expressinghuman beta-amyloid for use as a vaccine against Alzheimer's
disease. Biotechnol. Lett. 30: 1839–45.
Young L. and Dong Q. (2004). Two-step total gene synthesis method. Nucleic Acids
Research 32: e59.
Yu J. and Langridge W.H.R. (2001). A plant-based multicomponent vaccine protects
mice from enteric diseases. Nat. Biotechnol. 19: 548–552.
Zhang H., Liu M., Li Y., Zhao Y., He H., Yang G. and Zheng C. (2010). Oral
immunogenicity and protective efficacy in mice of a carrot-derived vaccine
candidate expressing UreB subunit against Helicobacter pylori. Protein Expr.
Purif. 69:127-31.
Page 147
Bibliography
82
Ertl H. and Xiang Z. (1996a). Novel vaccine approaches. J. Immunol. 156: 3579-
3582.
Ertl H.C.J. and Xiang A.Q. (1996b). Genetic Immunization. Viral Immunology 9:1-9.
Falnes P. and Sandvig K. (2000). Penetration of protein toxins into cells. Curr. Opin.
Cell Biol. 12: 407–413.
Farrance C.E., Rhee A., Jones R.M., Musiychuk K., Shamloul M., Sharma S.,
Mett V., Chichester J.A., Streatfield S.J., Roeffen W., van de Vegte-Bolmer
M., Sauerwein R.W., Tsuboi T., Muratova O.V., Wu Y. and Yusibov V.
(2011). A plant-produced Pfs230 vaccine candidate blocks transmission of
Plasmodium falciparum.Clin Vaccine Immunol. 18: 1351-7.
Fekadu M. and Schaddock J.H. (1984). Peripheral distribution of virus in dogs
inoculated with two strains of rabies virus. An. J. Vet. Res. 45: 724-729.
Fekadu M., Sumner J., Shaddock J., Sanderlin D. and Baer G. (1992). Sickness
and recovery of dogs challenged with street rabies virus after vaccination with
vaccinia virus recombinant expressing rabies virus N protein. J. Virol. 66: 2601-
2604.
Fischer R., Drossard J., Commandeur U., Schillberg S. and Emans N. (1999).
Towards molecular farming in the future: moving from diagnostic protein and
antibody production in microbes to plants. Biotechnol. Appl. Biochem. 30: 101-
108.
Flamand A. and Delagneau J.F. (1978). Transcriptional mapping of rabies virus in
vivo. J. Virol. 28: 518-523.
Flamand A., Wiktor T.J. and Koprowski H. (1980). Use of hybridoma monoclonal
antibodies in the detection of antigenic differences between rabies and rabies-
related virus proteins. I. The nucleocapsid protein. J. Gen. Virol. 48: 97-104.
Flamand A., Blancou J., Coulon P.O., Lafay F., Leblois H., Prehaud C. and
Tuffereau C. (1989). The antigenic structure of the rabies glycoproteins.
Application of basic research to oral vaccination of foxes. “New Development in
Rabies control.” WHO, Paris.
Page 148
Bibliography
83
Flamand A., Raux H., Gaudin Y. and Ruigrok R.W.H. (1993). Mechanisms of
rabies virus neutalization. Virology 194: 302-313.
Flores H.E. and Curtis W.R. (1992). Approaches to understanding and manipulating
the biosynthetic potential of plant roots. In: Pederson H, Mutharsan R, Di Biasio
D, editors. Biochemical Engineering VII: Cellular and Reaction Engineering.
Ann. N.Y. Acad. of Sci. 665: 188-209.
Floss D.M., Mockey M., Zanello G., Brosson D., Diogon M., Frutos R., Bruel T.,
Rodrigues V., Garzon E., Chevaleyre C., Berri M., Salmon H., Conrad U.
and Dedieu L. (2010). Expression and immunogenicity of the mycobacterial
Ag85B/ESAT-6 antigens produced in transgenic plants by elastin-like peptide
fusion strategy. J. Biomed. Biotechnol. 2010: 274346.
Foley H.D., McGettigan J.P., Siler C.A., Dietzschold B., Schnell M.J. (2000). A
recombinant rabies virus expressing vesicular stomatitis virus glycoprotein fails
to protect againstrabies virus infection. Proc. Natl. Acad. Sci., U. S. A. 97: 14680-
5.
Frankel A.E., Tagge E., Chandler J., Burbage C., Hancock G., Vessel J. and
Willingham M. (1996). Characterization of single site ricin toxin B chain
mutants. Bioconjug. Chem. 7: 30–37.
Frankel A.E., Burbage C., Fu T., Tagge E., Chandler J. and Willingham M.
(1996). Characterization of a ricin fusion toxin targeted to the interleukin-2
receptor. Protein Eng. 9: 913–919.
Fraser G.C., Hooper P.T. and Lunt R.A., Gould A.R., Gleeson L.J., Hyatt A.D.,
Russell G.M., Kattenbelt J.A. (1996). Encephalitis caused by a Lyssavirus in
Fruit bats in Australia. Emorg. Infect. Dis. 2: 327-331.
Fredericksen B. L. and Whitt M.A. (1996). Mutations at two conserved acidic amino
acids in the glycoprotein of vesicular stomatitis virus affect pH-dependent
conformational changes and reduce the pH threshold for membrane fusion.
Virology 217: 49-57.
Page 149
Bibliography
84
Fu Z. F. (1997). Rabies and rabies research: past, present and future. Vaccine 15: 520-
524.
Fu Z.F., Dietzschold B., Schumacher C.L., Wunner W.H., Ertl H.C.J. and
Koprowski H. (1991). Rabies virus nucleoprotein expressed in and purified from
insect cells is efficacious as a vaccine. Proc. Natl. Acad. Sci., U.S.A. 88: 2001-
2005.
Fujii H., Takita-Sonoda Y., Mifune K., Hirai K., Nishizono A. and Mannen K.
(1994). Protective efficacy in mice of post-exposure vaccination with vaccinia
virus recombinant expressing either rabies virus glycoprotein or nucleoprotein.
Journal of General Virology 75: 1339-1344.
Galbraith D.W., Zeiher C.A., Harkins K.R. and Afonso C.L. (1992). Biosynthesis,
processing and targeting of the G-protein of vesicular stomatitis virus in tobacco
protoplasts. Planta 186:324–36.
Gao Y., Ma Y., Li M., Cheng T., Li S.W. and Zhang J. (2003). Oral immunization of
animals with transgenic cherry tomatillo expressing HBsAg. World J.
Gastroenterol. 9: 996-1002.
Gastka M., Horvath J. and Lentz T.L. (1996). Rabies virus binding to the nicotinic
acetylcholine receptor -subunit demonstrated by the virus overlay protein
binding assay. Journal of General Virology 77: 2437-2440.
Gaudin Y., Ruigrok R.W., Tuffereau C., Knossow M. and Flamand A. (1992).
Rabies virus glycoprotein is a trimer. Virology 187: 627-632.
Gaudin Y., Ruigrok R.W.H., Knossow M. and Flamand A. (1993). Low-pH
conformational changes of rabies virus glycoprotein and their role in membrane
fusion. J. Virol. 67: 1365-1372.
Gaudin Y., Ruigrok R.W.H. and Brunner J. (1995a). Low-pH induced
conformational changes in viral fusion proteins: implications for the fusion
mechanism. Journal of General Virology 76: 1541-1556.
Gaudin Y., Tuffereau C., Durrer P., Flamand A. and Ruigrock R. (1995b).
Biological function of the low-pH, fusion-inactive conformation of rabies virus
Page 150
Bibliography
85
glycoprotein (G): G is transported in a fusion-inactive state-like conformation. J.
Virol. 69: 5528-5533.
Gaudin Y., Raux H., Flamand A. and Ruigrok R. (1996). Identification of amino
acids controlling the low-pH-induced conformational change of rabies virus
glycoprotein. J. Virol. 70: 7371-7378.
Gaudin Y. (1997). Folding of rabies virus glucoprotein: epitope acquisition and
interaction with endoplasmic reticulum chaperons. J. Virol. 71: 3742-3750.
Gaume A., Komarnytsky S., Borisjuk N. and Raskin I. (2003). Rhizosecretion of
recombinant proteins from plant hairy roots. Plant Cell Rep. 21: 1188–93.
Giddings G., Allison G., Brooks D. and Carter A. (2000). Transgenic plants as
factories for biopharma-ceuticals. Nat. Biotechnol. 18: 1151-1155.
Gil F., Brun A., Wigdorovitz A., Catalá R., Martínez-Torrecuadrada J.L. and
Casal I. (2001). High-yield expression of a viral peptide vaccine in transgenic
plants. FEBS Lett. 488: 13–7.
Gilleland Jr. H.E., Gilleland L.B., Staczek J., Harty R.N., Garcia-Sastre A. and
Palese P. (2000). Chimeric animal and plant viruses expressing epitopes of outer
membrane protein F as a combined vaccine against Pseudomonas aeruginosa lung
infection. FEMS Immunol. Med. Microbiol. 27: 291–7.
Gillet J.P., Derer P. and Tsiang H. (1986). Axonal transport of rabies virus in the
central nervous system of the rat. J. Neuropathol. Exp. Neurol. 45: 619-634.
Goddjin O.J.M. and Pen J. (1995). Plants as bioreactors. TIBTECH. 13: 379-387.
Golovkin M., Spitsin S., Andrianov V., Smirnov Y., Xiao Y. and Pogrebnyak N.
(2007). Smallpox subunit vaccine produced in planta confers protection in mice.
Proc. Natl. Acad. Sci., U.S.A. 104: 6864–9.
Gómez N., Carrillo C., Parra F, Salinas J., Borca M.V., Escribano M. (1998).
Expression of immunogenic glycoprotein S polypeptides from transmissible
gastroenteritis coronavirus in transgenic plants. Virology 249: 352–8.
Page 151
Bibliography
86
Gómez N, Wigdorovitz A, Castañón S, Gil F, Ordás R. and Borca M.V. (2000).
Oral immunogenicity of the plant derived spike protein from swine-transmissible
gastroenteritis coronavirus. Arch. Virol. 145: 1725–32.
Gonzalez-Rabade N., McGowan E.G., Zhou F., McCabe M.S., Bock R., Dix P.J.,
Gray J.C. and Ma J.K. (2011). Immunogenicity of chloroplast-derived HIV-1
p24 and a p24-Nef fusion protein following subcutaneous and oral administration
in mice. Plant Biotechnol. J. 9: 629-38.
Greco R., Michel M., Guetard D., Cervantes-Gonzalez M., Pelucchi N. and Wain-
Hobson S. (2007). Production of recombinant HIV-1/HBV virus-like particles in
Nicotiana tabacum and Arabidopsis thalianaplants for a bivalent plant-based
vaccine. Vaccine 25: 8228–40.
Halonen P.E., Murphy F.A., Fields B.N. and Reese D.R. (1968). Hemagglutination
of rabies and some other bullet-shaped viruses. Proc. Soc. Exp. Biol. Med. 127:
1037-1042.
Hanham C.A., Zhao F. and Tignor G.H. (1993). Evidence from the anti-idiotypic
network that the acetylcholine receptor is a rabies virus receptor. J. Virol. 67:
530-542.
Hanna J.N., Carney I.K., Smith G.S., Tannenberg A.E., Deverill J.E., Bothat J.A.,
Serafin I.L., Harrower H.J., Fitzipatrick P.F. and Searle J.W. (2000).
Australian bat lyssavirus infection: a second Human case, with a long incubation
period. Medical Journal of Australia 172: 597-599.
Haq T.A., Mason H.S., Clements J.D. and Arntzen C.J. (1995). Oral immunization
with a recombinant bacterial antigen produced in transgenic plants. Science 268:
714-716.
Harlow E. and Lane D. (1988). Antibodies: A Laboratory Manual. Cold Spring
Harbor, NY: Cold Spring Harbor Laboratory Press.
Hashizume F., Hino S., Kakehashi M., Okajima T., Nadano D. and Aoki N. (2008).
Development and evaluation of transgenic rice seeds accumulating a type II-
collagen tolerogenic peptide. Transgenic Res. 17: 1117–29.
Page 152
Bibliography
87
Haupt W. (1999). Rabies: risk of exposure and current trends in prevention of human
cases. Vaccine 17: 1742-1749.
He Z.M., Jiang X.L., Qi Y. and Luo D.Q. (2008). Assessment of the utility of the
tomato fruit-specific E8 promoter for driving vaccine antigen expression.
Genetica 133: 207–14.
Heijden R.W.J., Langedijk J.P.M., Goren J., Ujtdehaaad F.G.C.M., Meloen R.H.
and Osterhaus A.D.M.E. (1993). Structural and functional studies on a unique
linear neutralizing antigenic site (G5) of the rabies virus glycoprotein. Journal of
General Virology 49: 1539-1545.
Hein M.B., Yeo T., Wang F. and Sturtevant A. (1996). Expression of cholera toxin
subunits in plants. Ann. N.Y. Acad. Sci. 792: 50–6.
Herman R.C., Schubert M., Keene J.D. and Lazzarini R.A. (1980). Polycistronic
vesicular stomatitis virus RNA transcripts. Proc. Natl. Acad. Sci., U.S.A. 77:
4662-4665.
Hernández M., Cabrera-Ponce J.L., Fragoso G., López-Casillas F., Guevara-
García A. and Rosas G. (2007). A new highly effective anti-cysticercosis
vaccine expressed in transgenic papaya. Vaccine 25: 4252–60.
Hiatt A., Cafferkey R. and Bowdish K. (1989). Production of antibodies in transgenic
plants. Nature 342: 76-78.
Hiatt A.C. and Ma J.K.C. (1993). Characterization and applications of antibodies
produced in plants. Int. Rev. Immunol. 10: 139-152.
Holmgren J., Lonnroth I., Mansson J.-E. and Svennerholm L. (1975). Interaction of
cholera toxin and membrane GM1 ganglioside of small intestine. Proc. Natl.
Acad. Sci, U.S.A. 72: 2520–2524.
Hood E.E., Witcher D., Maddock S., Meyer T., Baszezynski C., Bailey M., Flyn P.,
Register J., Marshall L., Bond D., Kulisek E., Kusnadi A., Evangelista R.,
Nikolov Z., Wooge C., Mehigh R., Hennan R., Heman R., Kappel W.,
Ritland D., Li C-P. and Howard J. (1997). Commercial production of avidin
Page 153
Bibliography
88
from transgenic maize: characterization of transformant, production, processing,
extracting and purification. Mol. Breed. 3: 291-306.
Hoover J.E. and Strick P.L. (1999). The organization of cerebellar and basal ganglia
outputs to primary motor cortex as revealed by retrogade trans neuronal transport
of herpes simplex virus type 1. J. Neuro. Sci. 19: 1446-1468.
Horsch R.B., Fry J.E., Hoffmann N.L., Eicholtz D., Rogers S.G. and Fraley R.T.
(1985). A simple and general method for transferring genes into plants. Science
227: 1229-1231.
Houdebine L.M. (2009). Production of pharmaceutical proteins by transgenic animals.
Comp. Immunol. Microbiol. Infect. Dis. 32: 107–21.
Huang Z., Dry I., Webster D., Strugnell R. and Wesselingh S. (2001). Plant-derived
measles virus hemagglutinin protein induces neutralizing antibodies in mice.
Vaccine 19: 2163–71.
Huang Z., Elkin G., Maloney B.J., Beuhner N., Arntzen C.J. and Thanavala Y.
(2005). Virus-like particle expression and assembly in plants: hepatitis B and
Norwalk viruses. Vaccine 23: 1851–8.
Hurtley S.M., Bole D.G., Hoover L.H., Helenius A. and Copeland C.S. (1989).
Interactions of misfolded influenza virus hemagglutinin with binding protein
(BiP). J. Cell Biol. 108: 2117-26.
Hwang C., Sinskey A.H. and Lodish H.F. (1992). Oxidised redox state of glutathione
in the endoplasmic reticulum. Science 257: 1496-1502.
Ito H., Minamoto N., Watanabe T., Goto H., Luo T.R., Sugiyama M., Kinjo T.,
Mannen K., Mifune K., Konobe T., Yoshida I. and Takamizawa A. (1994). A
unique mutation of glycoprotein gene of the attenuated RC-HL strain of rabies
virus, a seed virus used for production of animal vaccine in Japan. Microbiol.
Immunol. 38: 479-482.
Iwasaki Y. and Clark H.F. (1975). Cell to cell transmission of virus in the central
nervous system II. Experimental rabies in mouse. Lab. Inves. 33: 391-399.
Page 154
Bibliography
89
Jackson A.C. and Reimer D.L. (1989). Pathogenesis of experimental rabies in mice:
An immunohistochemical study. Acta. Neuropathol. 78: 159-165.
Jaeger G., Scheffer S., Jacobs A., Zambre M., Zobell O., Goossens A., Depicker A.
and Angenon G. (2002). Boosting heterologous protein production in transgenic
dicotyledonous seeds using Phaseolus vulgaris regulatory sequences. Nat.
Biotechnol. 20: 1265–1268.
Jani D., Meena L.S., Rizwan-ul-Haq Q.M., Singh Y., Sharma A.K., Tyagi A.K.
(2002). Expression of cholera toxin B subunit in transgenic tomato plants.
Transgenic Res. 11: 447–54.
Jani D., Singh N.K., Bhattacharya S., Meena L.S., Singh Y. and Upadhyay S.N.
(2004). Studies on the immunogenic potential of plant-expressed cholera toxin B
subunit. Plant Cell Rep. 22: 471–7.
Jasmin L., Burkey A.R., Card J.P. and Basbaum A.I. (1997). Transneuronal
labeling of a nociceptive pathway, the spino–(trigemino-) parabrachio-
amygdaloid, in the rat. J. Neuro. Sci. 17: 3751-3765.
Jiang X.L., He Z.M., Peng Z.Q., Qi Y., Chen Q. and Yu S.Y. (2007). Cholera toxin
B protein in transgenic tomato fruit induces systemic immune response in mice.
Transgenic Res. 16: 169–75.
Johnson R.T. (1965). Experimental rabies studies of cellular vulnerability and
pathogenesis using fluorescent antibody staining. J. Neuropathol. Exp. Neurol.
24: 662-674.
Joshi L. and Lopez L.C. (2005). Bioprospecting in plants for engineered proteins.
Curr. Opin. Plant Biol. 8: 223-6.
Joung Y.H., Youm J.W., Jeon J.H., Lee B.C., Ryu C.J. and Hong H.J. (2004).
Expression of the hepatitis B surface S and preS2 antigens in tubers of Solanum
tuberosum. Plant Cell Rep. 22: 925–30.
Page 155
Bibliography
90
Jul-Larsen A., Madhun A.S., Brokstad K.A., Montomoli E., Yusibov V. and Cox
R.J. (2012). The human potential of a recombinant pandemic influenza vaccine
produced in tobacco plants. Hum. Vaccin. Immunother. 8: 653-61.
Kalbina I., Engstrand L., Andersson S. and Strid A. (2010). Expression of
Helicobacter pylori TonB protein in transgenic Arabidopsis thaliana: toward
production of vaccine antigens in plants. Helicobacter 15:430-7.
Kalbina I., Wallin A., Lindh I., Engström P., Andersson S., Strid K. (2011). A
novel chimeric MOMP antigen expressed in Escherichia coli, Arabidopsis
thaliana, and Daucus carota as a potential Chlamydia trachomatis vaccine
candidate. Protein Expr. Purif. 80: 194-202.
Kang T.J., Han S.C. and Yang M.S. (2005). Expression of the B subunit of E. coli
heat-labile enterotoxin in tobacco using a herbicide resistance gene as a selection
marker. Plant Cell Tissue Organ Cult. 81: 165–74.
Kang T.J., Han S.C., Yang M.S. and Jang Y.S. (2006). Expression of synthetic
neutralizing epitope of porcine epidemic diarrhea virus fused with synthetic B
subunit of Escherichia coli heat-labile enterotoxin in tobacco. Protein Expr.
Purif. 46: 16-22.
Kang T.J., Lee W.S., Choi E.G., Kim J.W., Kim B.G. and Yang M.S. (2006b).
Mass production of somatic embryos expressing Escherichia coli heat-labile
enterotoxin B subunit in Siberian ginseng. J. Biotechnol. 121: 124–33.
Kapusta J., Modelska A., Figlerowicz M., Pniewski T., Letellier M. and Lisowa O.,
(1999). A plant derived edible vaccine against hepatitis B virus. FASEB J.
13: 1796-1799.
Khandelwal A., Sita G.L. and Shaila M.S. (2003). Oral immunization of cattle with
hemagglutinin protein of rinderpest virus expressed in transgenic peanut induces
specific immune responses. Vaccine 21: 3282–9.
Kieny M.P., Lathe R., Drillien R., Spehner D., Skory S., Schmitt D., Wiktor T.,
Koprowski H. and Lecocg J.-P. (1984). Expression of rabies virus glycoprotein
from a recombinant vaccinia virus. Nature 312: 163-166.
Page 156
Bibliography
91
Kim T.G. and Langridge W.H.R. (2003a). Assembly of cholera toxin B subunit full-
length rotavirus NSP4 fusion protein oligomers in transgenic potato. Plant Cell
Rep. 21: 884–90.
Kim H.S., Euym J.W., Kim M.S., Lee B.C., Inhee M.J. and Jeon J.H. (2003b).
Expression of human amyloid peptide in transgenic potato. Plant Sci. 165: 1445–
51.
Kim T.G., Galloway D.R. and Langridge W.H.R. (2004). Synthesis and assembly of
anthrax lethal factor-cholera toxin B-subunit fusion protein in transgenic potato.
Mol. Biotechnol. 28: 175–183
Kim Y.S., Kang T.J., Jang Y.S. and Yang M.S. (2005). Expression of neutralizing
epitope of porcine epidemic diarrhea virus in potato plants. Plant Cell Tissue
Organ Cult. 82: 125–30.
Kim Y.S., Kim B.G., Kim T.G., Kang T.J. and Yang M.S. (2006). Expression of a
cholera toxin B subunit in transgenic lettuce (Lactuca sativa L.) using
Agrobacterium-mediated transformation system. Plant Cell Tissue Organ Cult.
87: 203–10.
King A. and Crick J. (1988). Rabies–related viruses In “Rabies” (J.B. Compbell and
K.M. Charlton, Eds.) Kluwer Academic Publishers, Boston. 177-199.
Kissi B., Tordo N. and Bourhy H. (1995). Genetic polymorphism in the rabies virus
nucleoprotein gene. Virology 209: 526-537.
Kissling R.E. (1958). Growth of rabies virus in non-nervous tissue culture. Proc. Soc.
Trop. Med. Hyg. 98: 223-225.
Klepfer S.R., Uffelman J., Jacobs P., Bollen A. and Jones E.V. (1993).
Characterization of rabies glycoprotein expressed in yeast. Arch. Virol. 128: 269-
286.
Ko S., Liu J.R., Yamakawa T. and Matsumoto Y. (2006). Expression of the antigen
(SpaA) in transgenic hairy roots of tobacco. Plant Mol. Biol. Rep. 24: 251–251.
Page 157
Bibliography
92
Kohl T.O., Hitzeroth I.I., Christensen N.D. and Rybicki E.P. (2007). Expression of
HPV-11 L1 protein in transgenic Arabidopsis thaliana and Nicotiana tabacum.
BMC Biotechnol. 7: 56–69.
Komarnytsky S., Gaume A., Garvey A., Borisjuk N. and Raskin I. (2004). A quick
and efficient system for antibiotic-free expression of heterologous genes in
tobacco roots. Plant Cell Rep. 22: 765–73.
Kong Q., Richter L., Yang Y.F., Arntzen C.J., Mason H.S., Thanavala Y. (2001).
Oral immunization with hepatitis B surface antigen expressed in transgenic
plants. Proc. Natl. Acad. Sci., U.S.A. 98: 11539–44.
Koo M., Bendahmane M., Lettieri G.A., Paoletti A.D., Lane T.E., Fitchen J.H.,
Buchmeier M.J. and Beachy R.N. (1999). Protective immunity against murine
hepatitis virus (MHV) induced by intranasal or subcutaneous administration of
hybrids of tobacco mosaic virus that carries an MHV epitope. Proc. Natl. Acad.
Sci., U.S.A. 96: 7774-7779.
Koprowski H. (2005). Vaccines and sera through plant biotechnology. Vaccine 23:
1757-63.
Kumar G.B.S., Ganapathi T.R., Revathi C.J., Srinivas L., Bapat V.A. (2005a).
Expression of hepatitis B surface antigen in transgenic banana plants. Planta 222:
484–93.
Kumar G.B.S., Ganapathi T.R., Srinivas L., Revathi C.J., Bapat V.A. (2005b).
Secretion of hepatitis B surface antigen in transformed tobacco cell suspension
cultures. Biotechnol. Lett. 27: 927–32.
Kumar G.B.S., Ganapathi T.R., Srinivas L., Revathi C.J., Bapat V.A. (2006).
Expression of hepatitis B surface antigen in potato hairy roots. Plant Sci. 170:
918–25.
Kuroda K., Geyer H., Geyer R., Doerfler W. and Klenk H.D. (1990). The
oligosaccharides of influenza virus hemagglutinin expressed in insect cells by a
baculovirus vector. Virology 174: 418-429.
Page 158
Bibliography
93
Kusandi A.R., Evangelista R.L., Hood E.E., Howard J.A. and Niklov Z.L. (1998).
Processing of transgenic corn seed and its effect on the recovery of the
recombinant ß-glucuronidase. Biotechnol. Bioeng. 60:44-52.
Kusnadi A.R., Nikolov Z.L. and Howard J. (1997). Production of recombinant
proteins in transgenic plants: practical considerations. Biotech. Bioeng. 56: 473-
484.
Lafay F., Benomonsour A., Chebli K. and Flamand A. (1996). Immunodominant
epitopes defined by a yeast – expressed library of random fragments of the rabies
glycoprotein map outside major antigenic sites. Journal of General Virology 77:
339-346.
Lafay F., Coulon P., Astic L., Saucier D., Riche D., Holley A. and Flamand A.
(1991). Spread of the CVS strain of rabies virus and of the avirulent mutant
AvO1 along the olfactory pathways of the mouse after intranasal inoculation.
Virology 183: 320-330.
Lafon M., Wiktor T. and MacFarlan R. (1983). Antigenic sites on the CVS rabies
virus glycoprotein: analysis with monoclonal antibodies. Journal of General
Virology 64: 843-845.
Lafon M. and Wiktor T. (1985). Antigenic sites on the ERA rabies virus
nucleoprotein and non-structural protein. Journal of General Virology 66: 2125-
2133.
Lafon M., Perrin P., Versmisse P. and Sureau P. (1985b). Use of a monoclonal
antibody for quantitation of rabies vaccine glycoprotein by enzyme immunoassay.
J. Biol. Stand. 13: 295-301.
Lafon M., Lafage M., Martinezarends A., Ramirez R., Vuillier F., Charron D.,
Lotteau V. and Scottalgara D. (1992). Evidence for a viral super antigen in
humans. Nature 358: 507-510.
Lai C.Y. and Dietzschold B. (1981). Amino acid composition and terminal sequence
analysis of the rabies virus glycoprotein: identification of the reading frame on
the cDNA sequence. Biochem. Biophys. Res. Comm. 103: 536-542.
Page 159
Bibliography
94
Lal P., Ramachandran V.G., Goyal R., Sharma R. (2007). Edible vaccines: current
status and future. Ind. J. Med. Microbiol. 25: 93-102.
Lambert J.M., Goldmacher V.S., Collision A.R., Nadler L.M., Blattler W.A.
(1991). An immunotoxin prepared with blocked ricin: a natural plant toxin
adapted for therapeutic use. Cancer Res. 51: 6236–6242.
Lamphear B.J., Streatfield S.J., Jilka J.M., Brooks C.A., Barker D.K., Turner
D.D. (2002). Delivery of subunit vaccines in maize seed. J. Control Release
85:169-80.
Lathe R.F., Kieny M.P., Schmitt D., Curtis P. and Lecocq J.P. (1984). M13
bacteriophage vectors for the expression of foreign proteins in Escherichia coli:
the rabies glycoprotein. J. Mol. Appl. Gen. 2: 331-342.
Lau O.S., Ng D.W., Chan W.W., Chang S.P. and Sun S.S. (2010). Production of the
42-kDa fragment of Plasmodium falciparum merozoite surface protein 1, a
leading malaria vaccine antigen, in Arabidopsis thaliana seeds. Plant Biotechnol.
J. 8: 994-1004.
Lauterslager T.G.M., Florack D.E.A., van der Wal T.J., Molthoff J.W., Langeveld
J.P.M. and Bosch D. (2001). Oral immunization of naive and primed animals
with transgenic potato tubers expressing LT-B. Vaccine 19: 2749–55.
Lee R.W.H., Strommer J., Hodgins D., Shewen P.E., Niu Y., Lo R.Y.C. (2001).
Towards development of an edible vaccine against bovine pneumonic
pasteurellosis using transgenic white clover expressing a Mannheimia
haemolytica A1 lkt50 fusion protein. Infect. Immun. 69: 5786–93.
Lees C.Y., Briggs D.J., Wu X., Davis R.D., Moore S.M., Gordon C., Xiang Z., Ertl
H.C.J., Tang de C.C. and Fu Z.F. (2002). Induction of protective immunity by
topic application of a recombinant adenovirus expressing rabies virus
glycoprotein. Veterinary Microbiology 85: 295-303.
Lefebvre D.D., Miki B.L. and Laliberte J.F. (1987). Mammalian metallothionein
functions in plants. BioTechnology 5: 1053-1056.
Page 160
Bibliography
95
Lefrancois L. (1984). Protection against lethal viral infection by neutralizing and
nonneutralizing monoclonal antibodies: distinct mechanisms of action. J. Virol.
51:208-214.
Lentz T., Burrage T.G., Smith A.L., Crick J. and Tignor G.H. (1982). Is the
acetylcholine receptor a rabies virus receptor? Science 215: 182-184.
Lentz T.L., Benson R.J.J., Klimowicz D., Wilson P.T. and Hawrot E. (1986).
Binding of rabies virus to purified Torpedo acetylcholine receptor. Mol. Brain
Res. 1: 211-219.
Lentz T.L., Hawrot E. and Wilson P.T. (1987). Synthetic peptides corresponding to
sequences of snake venom neurotoxins and rabies virus glycoprotein bind to the
nicotinic acetylcholine receptor. Proteins Struct. Funct. Genet. 2: 298-307.
Lentz T.L., Wilson P.T., Howrot E. and Speicher D.W. (1984). Amino acid
sequence similarity between rabies virus glycoprotein and snake curaremimetic
neurotoxins. Science 226: 847-848.
Li D., O’Leary J., Huang Y., Huner N.P.A., Jevnikar A.M. and Ma S. (2006).
Expression of cholera toxin B subunit and the B chain of human insulin as a
fusion protein in transgenic tobacco plants. Plant Cell Rep. 25: 417–424
Li J.T., Fei L., Mou Z.R., Wei J., Tang Y., He H.Y. (2006a). Immunogenicity of a
plant-derived edible rotavirus subunit vaccine transformed over fifty generations.
Virology 356: 171–8.
Li H.Y., Ramalingam S.K. and Chye M.L. (2006b). Accumulation of recombinant
SARS-CoVspike protein in plant cytosol and chloroplasts indicate potential for
development of plant derived oral vaccines. Exp. Biol. Med. 231: 1346–52.
Liu C., Towler M.J., Medrano G., Cramer C.L., Weathers P.J. (2009) Production
of Mouse Interleukin- 12 Is Greater in Tobacco Hairy Roots Grown in a Mist
Reactor Than in an Airlift Reactor. Biotechnol. Bioeng. 102: 1074-1086.
Lodmell D.L., Exposito J. and Ewalt L. (1993). Rabies virus anti-nucleoprotein
antibody protects against rabies virus challenge in vivo and inhibits rabies virus
replication in vitro. J. Virol. 67: 6080-6086.
Page 161
Bibliography
96
Lodmell D.L., Parnell M.J., Weyhrich J.T. and Ewalt L. (2003). Canine rabies DNA
vaccination: a single-dose intradermal injection into ear pinnae elicits elevated
and persistent levels of neuroimaging antibody. Vaccine 21: 3998-4002.
Lodmell D.L., Sumner J., Esposito J., Bellini W. and Ewalt L. (1991). Racoon
poxivirus recombinants expressing the rabies virus nucleoprotein protect mice
against lethal rabies virus infection. J. Virol. 65: 3400-3405.
Lou X.M., Yao Q.H., Zhang Z., Peng R.H., Xiong A.S. and Wang H.K. (2007).
Expression of the human hepatitis B virus large surface antigen gene in transgenic
tomato plants. Clin. Vaccine Immunol. 14: 464–9.
Lu Z., Lee K.J., Shao Y., Lee J.H., So Y., Choo Y.K., Oh D.B., Hwang K.A., Oh
S.H., Han Y.S. and Ko K. (2012). Expression of GA733-Fc fusion protein as a
vaccine candidate for colorectal cancer in transgenic plants. J. Biomed.
Biotechnol. 2012: 364240.
Luchakivskaia Iu.S., Olevinskaia Z.M., Kishchenko E.M., Spivak N.Ia., Kuchuk
N.V. (2012). Obtaining of hairy-root, callus and suspension carrot culture
(Daucus carota L.) able to accumulate human interferon alpha-2b. Tsitol.
Genet. 46:18-26.
Lynch J.C., Hoover J.E. and Strick P.L. (1994). In put to the primate frontal eye field
from the substantia nigra, superior colliculus, and dentate nucleus demonstrated
by transneuronal transport. Exp. Brain Res. 100: 181-186.
Ma J.K.C. and Hein M.B. (1995a). Immunotherapeutic potential of antibodies
produced in plants. TIBTECH. 13: 522-527.
Ma J.K.C., Hiatt A., Hein M., Vine N.D., Wang F., Stabila P., Van Dolleweerd C.,
Mostov K. and Lehner T. (1995b). Generation and assembly of secretory
antibodies in plants. Science 268: 716-719.
Ma J.K.C., Hikmat B., Wycoff K., Vine N., Chargelegue D., Yu L., Hein M. and
Lehner T. (1998). Characterization of a recombinant plant monoclonal secretory
antibody and preventive immunotherapy in human. Nat. Med. 4: 601-606.
Page 162
Bibliography
97
Ma J.K.C., Pascal M.W.D. and Christou P. (2003). The production of recombinant
pharmaceutical proteins in plants. Nature 4: 794-805.
Ma J.K.C., Barros E., Bock R., Christou P., Dale P.J., Dix P.J., Fischer R., Irwin
J., Mahoney R., Pezzotti M. (2005). Molecular farming for new drugs and
vaccines: Current perspectives on the production of pharmaceuticals in transgenic
plants. EMBO Rep. 6: 593–599.
Macfarlan R., Dietzschold B., Wiktor T., Kiel M., Houghten R., Lerner R.,
Sutcliffe G. and Koprowski H. (1984). T cell responses to cleaved rabies virus
glycoprotein and to synthetic peptides. J. Immunol. 133:2748-2752.
Macfarlan R., Dietzschold B. and Koprowski H. (1986). Stimulation of cytotoxic T
lymphocyte response by rabies virus glycoprotein and identification of an
immunodominant domain. Mol. Immunol. 23: 733-741.
Marquet-Blouin E., Bouche F.B., Steinmetz A., Muller C.P. (2003). Neutralizing
immunogenicity of transgenic carrot (Daucus carota L.) derived measles virus
hemagglutinin. Plant Mol. Biol. 51: 459–69.
Martín-Alonso J.M., Castañón S., Alonso P., Parra F., Ordás R. (2003). Oral
immunization using tuber extracts from transgenic potato plants expressing rabbit
hemorrhagic disease virus capsid protein. Transgenic Res. 12: 127–30.
Mason H.S., Lam D.M. and Arntzen C.J. (1992). Expression of hepatitis B surface
antigen in transgenic plants. Proc. Natl. Acad. Sci., U.S.A. 89: 11745-11749.
Mason H.S. and Arntzen C.J. (1995). Transgenic plants as vaccine production
systems. Trends Biotechnol. 13: 388-392.
Mason H.S., Ball J.M., Shi J.J., Jiang X., Estes M.K. and Arntzen C.J. (1996).
Expression of the Norwalk virus capsid protein in transgenic tobacco and its
immunogenicity in plants. Proc. Natl. Acad. Sci., U.S.A. 93: 5335-5340.
Mason H., Haq T., Clements J. and Arntzen C. (1998). Edible vaccine protects mice
against Escherichia coli heat-labile enterotoxin (LT): potatoes expressing a
synthetic LT-B gene. Vaccine 16: 1336-1343.
Page 163
Bibliography
98
Matsui T., Asao H., Ki M., Sawada K. and Kato K. (2009). Transgenic lettuce
producing a candidate protein for vaccine against edema disease. Biosci.
Biotechnol. Biochem. 73: 1628-34.
Matsumoto Y., Suzuki S., Nozoye T., Yamakawa T., Takashima Y., Arakawa T.,
Tsuji N., Takaiwa F. and Hayashi Y. (2009). Oral immunogenicity and
protective efficacy in mice of transgenic rice plants producing a vaccine
candidate antigen (As16) of Ascaris suum fused with cholera toxin B subunit.
Transgenic Res. 18: 185-92.
Matsumura T., Itchoda N. and Tsunemitsu H. (2002). Production of immunogenic
VP6 protein of bovine group A rotavirus in transgenic potato plants. Arch. Virol.
147: 1263–70.
McGarvey P.B., Hammond J., Dienelt M.M., Hooper D.C., Fu Z.F., Dietzschold B.
(1995). Expression of the rabies virus glycoprotein in transgenic tomatoes.
Biotechnology 13: 1484-1487.
McGehee D.S. and Lorca L.W. (1995). Physiological diversity of nicotinic
acetylcholine receptors expressed by vertebrate cells. Annu. Rev. Physiol. 57:
521-546.
McKenzie S.J. and Halsey J.F. (1984). Cholera toxin B subunit as a carrier protein to
stimulate a mucosal immune response. J. Immunol. 133:1818–1824.
Mebatsion R., Konig M. and Conzelmann K.K. (1996). Budding of rabies virus
particles in the absence of the spike glycoprotein. Cell 84: 941-951.
Mebatsion T., Weiland F. and Conzelmann K.K. (1999). Matrix protein of rabies
virus is responsible for the assembly and budding of bullet shaped particles and
interacts with the transmembrane spike glycoprotein G. J. Virol. 73: 242-250.
Medina-Bolivar F., Wright R., Funk V., Sentz D., Barroso L., Wilkins T.D., Petri
W. and Cramer C.L. (2003). A non-toxic lectin for antigen delivery of plant
based mucosal. Vaccines 21: 997–1005.
Meslin F.X., Fishbein D.B. and Matter H.C. (1994). Rationale and prospects for
rabies elimination in developing countries. In: Lyssaviruses, Rupprecht C.E.,
Page 164
Bibliography
99
Dietzschold B., Koprowski H. (Eds.), Berlin: Springer, 1-26. Springer-Verlag,
Berlin, Heidelberg, New York.
Meslin F.X. and Stohr K. (1997). Prospects for immunization against rabies in
developing countries. In: Dodet B. and Meslin F.X. (Eds.), Rabies Control in
Asia. 15-18.
Meyers A., Chakauya E., Shephard E., Tanzer F.L., Maclean J. and Lynch A.
(2008). Expression of HIV-1 antigens in plants as potential subunit vaccines.
BMC Biotechnol. 8: 1-15.
Middelton F.A. and Strick P.L. (1996). The temporal lobe is the target of output from
the basal ganglia. Proc. Natl Acad. Sci., U.S.A. 93: 8683-8687.
Mifune K., Ohuchi M. and Mannen K. (1982). Hemolysis and cell fusion by
rhabdoviruses. FEBS Lett. 137: 293-297.
Mirjalili H, Fakhr-Tabatabaei S, Bonfill M, Alizadeh H, Cusido R, Ghassempour
A, Palazon J. (2009) Morphology and withanolide production of Withania
coagulans hairy root cultures. Eng. Life Sci. 9: 197–204.
Mishra S., Yadav D.K., Tuli R. (2006). Ubiquitin fusion enhances cholera toxin B
subunit expression in transgenic plants and the plant-expressed protein binds
GM1 receptors more efficiently. J. Biotechnol. 127: 95-108.
Mishra N., Gupta P.N., Khatri K., Goyal A.K., Vyas S.P. (2008). Edible vaccines: a
new approach to oral immunization. Ind. J. Biotechnol. 7: 283–94.
Modelska A., Dietzschold B., Sleysh N., Fu Z.F., Steplewski K., Hooper D.C.,
Koprowski H. and Yusibov V. (1998). Immunization against rabies with plant-
derived antigen. Proc. Natl. Acad. Sci. USA. 95: 2481-2485.
Molina A., Hervás-Stubbs S., Daniell H., Mingo-Castel A.M. and Veramendi J.
(2004). High-yield expression of a viral peptide animal vaccine in transgenic
tobacco chloroplasts. Plant Biotechnol. J. 2:141–53.
Mor T.S., Sternfeld M., Arntzen C.J., Soreq H. and Mason H.S. (2001). Expression
of recombinant human acetylcholinesterase in transgenic tomato plants.
Biotechnol. Bioeng. 75: 259–66.
Page 165
Bibliography
100
Moravec T., Schmidt M.A., Herman E.M. and Woodford-Thomas T. (2007).
Production of Escherichia coli heat labile toxin (LT) B subunit in soybean seed
and analysis of its immunogenicity as an oral vaccine. Vaccine 25: 1647–57.
Morgenfeld M., Segretin M.E., Wirth S., Lentz E., Zelada A., Mentaberry A.,
Gissmann L. and Bravo-Almonacid F. (2009). Potato virus X coat protein
fusion to human papillomavirus 16 E7 oncoprotein enhance antigen stability and
accumulation in tobacco chloroplast. Mol. Biotechnol. 43: 243-9.
Morimoto K., Foley H.D., McGettigon J.P., Schnell M.J. and Dietzschold B.
(2000). Reinvestigation of the role of the rabies virus glycoprotein in viral
pathogenesis using a reverse genetics approach. J. Neurol. Virol. 6: 373-381.
Morimoto K., Hooper D.C., Spitsin S., Koprowski H. and Dietzschold B. (1999).
Pathogenicity of different rabies virus variants inversely correlates with apoptosis
and rabies virus glycoprotein expression in infected primary neuron culture. J.
Virol. 73: 510-517.
Morimoto K., Ni Y.J. and Kawai A. (1992). Syncytium formation is induced in the
murine neuroblastoma cell cultures which produce pathogenic type G proteins of
the rabies virus. Virology 189: 203-216.
Morimoto K., Patel M., Corisdeo S., Hooper D.C., Fu Z.F., Rupprecht C.E.,
Koprowski H. and Dietzschold B. (1996). Characterization of a unique variant
of bat rabies virus responsible for newly emerging human cases in North
America. Proc. Natl. Acad. Sci., USA. 93: 5653-5658.
Mortimer E., Maclean J.M., Mbewana S., Buys A., Williamson A.L., Hitzeroth I.I.
and Rybicki E.P. (2012). Setting up a platform for plant-based influenza virus
vaccine production in South Africa. BMC Biotechnol 12: 14.
Murphy F.A., Bauer S.P., Harrison A.K. and Winn W.C. (1973). Comparative
pathogenesis of rabies and rabies-like viruses: viral infection and transit from
inoculation site to the central nervous system. Lab. Invest. 28: 361-376.
Page 166
Bibliography
101
Murphy F.A. and Bauer S.P. (1974). Early street rabies virus infection in striated
muscle and later progression to the central nervous system. Intervirology.
3: 256-268.
Murphy F.A. (1977). Rabies pathogenesis. Arch. Virol. 54: 279-297.
Nadin-Davis S.A., Casey G.A. and Wandeler A. (1993). Identification of regional
variants of the rabies virus within the Canadian province of Ontario. Journal of
General Virology. 74: 829-837.
Nadin-Davis S.A., Casey G.A. and Wandeler A. (1994). A molecular
epidemiological study of rabies virus in central Ontario and Western Quebec.
Journal of General Virology 75: 2575-2583.
Nadin-Davis S.A., Sampath M.I., Casey G.A., TinLine R.R. and Wandeler A.
(1999). Phylogeographic patterns exhibited by Ontario rabies virus variants.
Epidemiology and infection 123:325-336.
Nadin-Davis S.A., Sheen M., Abdul-Malik M., Elmgren L., Armstrong J. and
Wandeler A.I. (2000). A Panel of MAB targeting the phosphoprotein identifies a
highly variable epitope of value for sensitive strain discrimination. J. Clinical
Microbiology 38:1397-1403.
Nashar T.O., Webb H.M., Eaglestone S., Williams N.A., Hirst T.R. (1996). Potent
immunogenicity of the B subunits of Escherichia coli heat-labile enterotoxin:
receptor binding is essential and induces differential modulation of lymphocyte
subsets. Proc. Natl. Acad. Sci., U.S.A. 93: 226–230.
Nelson G., Marconi P., Periolo O., La Torre J. and Alvarez M.A. (2012). Immuno-
competent truncated E2 glycoprotein of bovine viral diarrhea virus (BVDV)
expressed in Nicotiana tabacum plants: a candidate antigen for new generation of
veterinary vaccines. Vaccine 30: 4499-504.
Ni Y., Tominaga Y., Honda Y., Morimoto K., Sakamoto S. and Kawai A. (1995).
Mapping and characterization of a sequential epitope on the rabies virus
glycoprotein which is recognized by a neutralizing monoclonal antibody, Rg 719.
Microbiol. Immunol. 39:693-702.
Page 167
Bibliography
102
Nochi T., Takagi H., Yuki Y., Yang L., Masumura T. and Mejima M. (2007). Rice-
based mucosal vaccine as a global strategy for cold-chain and needle-free
vaccination. Proc. Natl. Acad. Sci., U.S.A.104: 10986–91.
O’Donnell P., Lavin A., Enquist L.W., Grace A.A. and Card J.P. (1997). Inter
connected parallel circuits between rat nucleus accumbens and thalamus revealed
by retrograde transynaptic transport of pseudorabies virus. J. Neurosci. 17: 2143-
2167.
Oszvald M., Kang T.J., Tomoskozi S., Jenes B., Kim T.G. and Cha Y.S. (2008).
Expression of cholera toxin B subunit in transgenic rice endosperm. Mol.
Biotechnol. 40: 261–8.
Oth D., Mercier G., Perrin P., Joffret M.L., Sureau P. and Thibodeau L. (1987).
The association of the rabies glycoprotein with liposome (immunosome) induces
as in vitro specific release of Interleukin-2. Cell. Immunol. 108: 220-226.
Pak C.C., Puri A. and Blumenthal R. (1997). Conformational changes and fusion
activity of vesicular stomatitis virus glycoprotein: [125I] iodonaphthyl azide
photolabeling studies in biological membranes. Biochem. 36: 8890-8896.
Pasteur L. (1885). Methode pour prevenir la rage après morsure. Compt. Rendus.
Acad. Sci., Paris 101: 765-774.
Peach C. and Velten J. (1991). Expression variability (position effect) of CAT and
GUS reporter genes driven by linked divergent T-DNA promoters. Plant Mol.
Biol. 17 49-60.
Pennica D., Holloway B.P., Heyward J.T. and Obijeski J.F. (1980). In vitro
translation of rabies virus messenger RNAs. Virology 103: 517-521.
Perrin P. and Atanasiu P. (1980). Partial structural study of oligosides of rabies virus
glycoprotein. Microbiological 3: 57-74.
Perrin P. and Atanasiu P. (1981). Rabies glycoprotein affinity for lipids. I. Structural
study of different types of spontaneous and induced associations.
Microbiological 4: 231-247.
Page 168
Bibliography
103
Perrin P., Thibodeau L., Dauguet C., Fritsch A. and Sureau, P. (1984).
Amplification expression vectors: the requirements for high level expression of
proteins, including glycoproteins. Journal of General Virology 68: 1233-1250.
Perrin P., Sureau P. and Thibodeau L. (1985a). Rabies immunosomes efficiency in
pre and post-exposure vaccinations: role of liposomes in glycoprotein
conformation and immunogenicity. In “Synthetic immunomodulators and
vaccines”, Zaoral M. (Eds.). 252-271.
Perrin P., Thibodeau L. and Sureau P. (1985b). Rabies immunosomes (subunit
vaccine) structure and immunogenicity. Pre- and post-exposure protection
studies. Vaccine 3: 325-332.
Perrin P., Joffret M.L., Leclerc C., Oth D., Sureau P. and Thibodeau L. (1988a).
Interleukin-2 increases protection against experimental rabies. Immunobiol 177:
199.
Perrin P., Joffret M.-L., Oth D., Leclerc C., Sureau P. and Thibodeau L. (1988b).
Interleukin-2 production in vitro: a new approach to the study of rabies vaccine
immunogenicity as appraised by testing different glycoprotein presentations.
Vaccine 6: 331.
Perrin P., Lafon M. and Sureau P. (1990). Rabies vaccines from Pasteur’s time up to
experimental subunit vaccines today. Adv. Biotechnol. Process 14: 325-345.
Plotkin S. (1993). Vaccination in the 21 st century. J. Infect. Dis. 168: 29-37.
Pogrebnyak N., Markley K., Smirnov Y., Brodzik R., Bandurska K. and
Koprowski H. (2006). Collard and cauliflower as a base for production of
recombinant antigens. Plant Sci. 171: 677–85.
Prehaud C., Coulon P., Lafay F., Thiers C. and Flamand A. (1988). Antigenic site
II of the rabies virus glycoprotein: structure and role in viral virulence. J. Virol.
62: 1-7.
Prehaud C., Takehara K., Flamand A. and Bishop D.H.L. (1989). Immunogenic
and protective properties of rabies virus glycoprotein expressed by baculovirus
vectors. Virology 173: 390-399.
Page 169
Bibliography
104
Prevec L., Campbell J.B., Christie B.S., Belbeck L. and Graham F.L. (1990). A
recombinant human adenovirus vaccine against rabies. J. inf. Dis. 161: 27.
Proudfoot N.J. and Brownlee G.G. (1976). 3’ non-coding region sequences in
eukaryotic messenger RNA. Nature 263: 211-214.
Qian B., Shen H., Liang W., Guo X., Zhang C., Wang Y., Li G., Wu A., Cao K.
and Zhang D. (2008). Immunogenicity of recombinant hepatitis B virus surface
antigen fused with preS1 epitopes expressed in rice seeds. Transgenic Res. 17:
621-31.
Ramírez Y.J.P., Tasciotti E., Gutierrez-Ortega A., Torres A.J.D., Flores M.T.O.
and Giacca M. (2007). Fruit-specific expression of the human immunodeficiency
virus type 1 tat gene intomato plants and its immunogenic potential in mice. Clin.
Vaccine Immunol. 14: 685–92.
Raux H., Coulon P., Lafay F. and Flamand A. (1995). Monoclonal antibodies which
recognize the acidic configuration of the rabies glycoprotein at the surface of the
virion can be neutralizing. J. Virol. 210: 400-408.
Richter L.J., Thanavala Y., Arntzen C.J. and Mason H.S. (2000). Production of
hepatitis B surface antigen in transgenic plants for oral immunization. Nat.
Biotechnol. 18: 1167–71.
Rigano M.M., Alvarez M.L., Pinkhasov J., Jin Y., Sala F. and Arntzen C.J. (2004).
Production of a fusion protein consisting of the enterotoxigenic Escherichia coli
heat-labile toxin B subunit and a tuberculosis antigen in Arabidopsis thaliana.
Plant Cell Rep. 22: 502–8.
Roberta M.K. and Peter L.S. (2000). Rabies as a transneuronal tracer of circuits in the
central nervous system. J. Neuro. Sci. Meth. 103: 63-71.
Rosales-Mendoza S., Soria-Guerra R.E., López-Revilla R., Moreno-Fierros L. and
Alpuche-Solís A.G. (2008). Ingestion of transgenic carrots expressing the
Escherichia coli heat-labile enterotoxin B subunit protects mice against cholera
toxin challenge. Plant Cell Rep. 27: 79–84.
Page 170
Bibliography
105
Rosales-Mendoza S., Alpuche-Solıs A.G., Soria-Guerra R.E., Moreno-Fierros L.,
MartnezGonzalez L. and Herrera-Dıaz A. (2009). Expression of an
Escherichia coli antigenic fusion protein comprising the heat labile toxin B
subunit and the heat stable toxin, and its assembly as a functional oligomer in
transplastomic tobacco plants. Plant J. 57: 45–54.
Rosales-Mendoza S., Soria-Guerra R.E., Moreno-Fierros L., Alpuche-Solís A.G.,
Martínez-González L. and Korban S.S. (2010). Expression of an immunogenic
F1-V fusion protein in lettuce as a plant-based vaccine against plague. Planta 232:
409-16.
Rose J.K. (1980). Complete intergenic and flanking gene sequences from the genome
of vesicular stomatitis virus. Cell 19: 415-421.
Rose J.K., Doolittle R.F., Anilionis A., Curtis P.J. and Wunner W.H. (1982).
Homology between the glycoproteins of vesicular stomatitis virus and rabies
virus. J. Virol. 43: 361-364.
Roy S., Tyagi A., Tiwari S., Singh A., Singh P.K., Sawant S.V. and Tuli R. (2010).
Rabies glycoprotein fused with B subunit of cholera toxin is expressed at high
level in tobacco plants and folds into biologically active pentameric protein.
Protein Expr. Purif. 70: 184-90.
Rubio-Infante N., Govea-Alonso D.O., Alpuche-Solís Á.G., García-Hernández
A.L., Soria-Guerra R.E., Paz-Maldonado L.M., Ilhuicatzi-Alvarado D.,
Varona-Santos J.T., Verdín-Terán L., Korban S.S., Moreno-Fierros L. and
Rosales-Mendoza S. (2012). A chloroplast-derived C4V3 polypeptide from the
human immuno-deficiency virus (HIV) is orally immunogenic in mice. Plant
Mol. Biol. 78: 337-49.
Rupprecht C.E., Hanlon C.A. and Hemachudha T. (2002). Rabies re-examined. The
Lancet Infect. Dis. 2: 327-343.
Rupprecht C.E., Smith J.S., Fekadu M. and Childs J.E. (1995). The ascension of
wildlife rabies: a cause for public health concern or intervention? Emerg. Infect.
Dis. 1: 107-114.
Page 171
Bibliography
107
Scotti N., Alagna F., Ferraiolo E., Formisano G., Sannino L., Buonaguro L., De
Stradis A., Vitale A., Monti L., Grillo S., Buonaguro F.M. and Cardi T.
(2009). High-level expression of the HIV-1 Pr55gag polyprotein in transgenic
tobacco chloroplasts. Planta 229: 1109-22.
Seif I., Coulon P., Rollin P.E. and Flamand A. (1985). Rabies virulence: effect on
pathogenicity and sequence characterization of rabies virus mutations affecting
antigenic site III of the glycoprotein. J. Virol. 53: 926-34.
Sharma M.K., Singh N.K., Jani D., Sisodia R., Thungapathra M. and Gautam J.K.
(2008). Expression of toxin co-regulated pilus subunit A (TCPA) of Vibrio
cholerae and its immunogenic epitopes fused to choleratoxin B subunit in
transgenic tomato (Solanum lycopersicum). Plant Cell Rep. 27: 307-18.
Sharp J.M. and Doran P.M. (1999). Effect of bacitracin on growth and monoclonal
antibody production by tobacco hairy roots and cell suspensions. Biotechnol.
Bioprocess Eng. 4: 253-258.
Shchelkunov S.N., Salyaev R.K., Pozdnyakov S.G., Rekoslavskaya N.I., Nesterov
A.E. and Ryzhova T.S. (2006). Immunogenicity of a novel, bivalent, plant-based
oral vaccine against hepatitis B and human immunodeficiency viruses. Biotechnol
Lett. 28: 959–67.
Shevchuk N.A., Bryksin A.V., Nusinovich Y.A., Cabello F.C., Sutherland M. and
Ladisch S. (2004). Construction of long DNA molecules using long PCR-based
fusion of several fragments simultaneously. Nucleic Acids Res. 32: e19.
Shin M.K., Jung M.H., Lee W.J., Choi P.S., Jang Y.S. and Yoo H.S. (2011).
Generation of transgenic corn-derived Actinobacillus pleuropneumoniae ApxIIA
fused with the cholera toxin B subunit as a vaccine candidate. J. Vet. Sci. 12:
401-3.
Shoji Y., Chichester J.A., Jones M., Manceva S.D., Damon E., Mett V., Musiychuk
K., Bi H., Farrance C., Shamloul M., Kushnir N., Sharma S. and Yusibov V.
(2011). Plant-based rapid production of recombinant subunit hemagglutinin
vaccines targeting H1N1 and H5N1 influenza. Hum. Vaccine 7: 41-50.
Page 172
Bibliography
108
Singh A., Yadav D., Rai K.M., Srivastava M., Verma P.C., Singh P.K. and Tuli R.
(2012). Enhanced expression of rabies virus surface G-protein in Escherichia coli
using SUMO fusion. Protein J. 31: 68-74.
Smith G.L., Mackett M. and Moss B. (1983a). Infectious vaccinia virus recombinants
that express hepatitis B virus surface antigen. Nature 302: 490-495.
Smith G.L., Murphy B.R. and Moss B. (1983b). Construction and characterization of
an infectious vaccinia virus recombinant that expresses the influenza
hemagglutinin gene and induces resistance to influenza virus infection in
hamsters. Proc. Natl. Acad. Sci., U.S.A. 80: 7155-7159.
Smith J.S., Orciari L.A., Yager P.A., Seidel H. D. and Warner C.K. (1992).
Epidemiologic and historic relationships among 87 rabies virus isolates as
determined by limited sequence analysis. J. Infect. Dis. 166: 296-307.
Smith J.S. (1996). New aspects of rabies with emphasis on epidemiology, diagnosis
and prevention of the disease in the United States. Clin. Microbiol. Rev. 9: 166-
176.
Smith M.L., Keegan M.E., Mason H.S. and Shuler M.L. (2002a). Factors important
in the extraction, stability and in vitro assembly of the hepatitis B surface antigen
derived from recombinant plant systems. Biotechnol. Prog. 18: 538–50.
Smith M.L., Mason H.S. and Shuler M.L. (2002b). Hepatitis B surface antigen
(HbsAg) expression in plant cell culture: kinetics of antigen accumulation in
batch culture and its intracellular form. Biotechnol. Bioeng. 80: 812–22.
Sojikul P., Buehner N. and Mason H.S. (2003). A plant signal peptide hepatitis B
surface antigen fusion protein with enhanced stability and immunogenicity
expressed in plant cells. Proc. Natl. Acad. Sci., U.S.A. 100: 2209–14.
Sokol F. and Clark F. (1973). Phosphoproteins, structural components of
rhabdoviruses. Virology 52: 246-263.
Soria-Guerra R.E., Rosales-Mendoza S., Márquez-Mercado C., López-Revilla R.,
Castillo-Collazo R. and Alpuche-Solís A.G. (2007). Transgenic tomatoes
Page 173
Bibliography
109
express an antigenic polypeptide containing epitopes of the diphtheria, pertussis
and tetanus exotoxins, encoded by a synthetic gene. Plant Cell Rep. 26: 961–8.
Sphyris N., Lord J.M., Wales R. and Roberts L.M. (1995). Mutational analysis of
the Ricinus lectin B-chains. J. Biol. Chem. 270: 20292–20297.
Spitsin S., Andrianov V., Pogrebnyak N., Smirnov Y., Borisjuk N. and
Portocarrero C. (2009). Immunological assessment of plant-derived avian flu
H5/HA1 variants. Vaccine 27: 1289–92.
Steeves R.M., Denton M.E., Barnard F.C., Henry A. and Lambert J.M. (1999).
Identification of three oligosaccharide binding sites in ricin. Biochemistry 38:
11677–11685.
Strack A.M. and Loewy A.D. (1990). Pseudorabies virus: A highly specific trans
neuronal cell body marker in the sympathetic nervous system. J. Neurosci. 10:
2139-47.
Streatfield S.J., Jilka J.M., Hood E.E., Turner D.D., Bailey M.R. and Mayor J.M.
(2001). Plant-based vaccines: unique advantages. Vaccine 19: 2742–8.
Streatfield S.J., Lane J.R., Brooks C.A., Barker D.K. Poage M.L., Mayor J.M.,
Lamphear B.J., Drees C.F., Jilka J.M., Hood E.E. and Howard J.A. (2003).
Corn as a production system for human and animal vaccines. Vaccine 21: 812-
815.
Sumner J., Fekadu M., Shaddock J., Esposito. and Bellini W. (1991). Protection of
mice with vaccinia virus recombinants that express the rabies nucleoprotein.
Virology 183: 703-710.
Sun N., Cassell M.D. and Perlman S. (1996). Anterograde, transneuronal transport of
herpes simplex virus type 1 strain HI29 in the murine visual system. J. Virol. 70:
5405-13.
Superti F., Derer M. and Tsiang H. (1984a). Mechanism of rabies virus entry into
CER cell. Journal of General Virology 65: 781-789.
Superti F., Seganti L., Tsiang H. and Orsi N. (1984b). Role of phospholipids in
rhabdovirus attachment to CER cells. Arch. Virol. 81: 321-328.
Page 174
Bibliography
110
Superti F., Hauttecoeur B., Morelec M.J., Goldoni P., Bizzini B. and Tsiang H.
(1986). Involvement of gangliosides in rabies virus infection. Journal of General
Virology 67: 47-56.
Swimmer C., Lehar S.M., McCafferty J., Chiswell D.J., Blättler W.A. and Guild
B.C. (1992). Phage display of ricin B chain and its single binding domains:
system for screening galactose-binding mutants. Proc. Natl. Acad. Sci., U.S.A.
89: 3756–3760.
Tackaberry E.S., Dudani A.K. Prior F., Tocchi M., Sardana R., Altosaar I. and
Ganz P.R. (1999). Developemnt of biopharmaceuticals in plant expression
systems: cloning, expression and immunological reactivity of human
cytomegalovirus glycoprotein B (UL55) in seeds of transgenic tobacco. Vaccine
17: 3020-3029.
Tackett C.O., Mason H.S., Losonsky G., Clements J.D., Levine M.M. and Arntzen
C.J. (1998). Immunogenicity in humans of a recombinant bacterial antigen
delivered in transgenic potato. Natl. Med. 4: 607-609.
Tackett C.O., Mason H.S., Losonsky G., Estes M.K., Levin M.M. and Arntzen C.J.
(2000). Human immune responses to a novel Norwalk virus vaccine delivered in
transgenic potatoes. J. Infect. Dis. 182: 302-305.
Tagge E.P., Chandler J., Harris B., Czako M., Marton L., Willingham M.C.,
Burbage C., Afrin L. and Frankel A.E. (1996). Preproricin expressed in
Nicotiana tabacum cells in vitro is fully processed and biologically active. Protein
Expr. Purif. 8: 109–118.
Takita-Sonada Y., Fujii H., Mifune K., Ito Y., Hiraga M., Nishizono A., Mannen
K. and Minamoto N. (1993). Resistance of mice vaccinated with rabies virus
internal structural proteins to lethal infection. Arch. Virol. 132: 51-65.
Tang Y., Rampin O., Giuliano R. and Ugolini G. (1999). Spinal and brain circuits to
motoneurons of the bulbospongiosus muscle: Retrograde transneuronal tracing
with rabies virus. J. Comp. Neurol. 414: 167-192.
Page 175
Bibliography
111
Taylor J., Meignier B., Tartaglia J., Languet B., VanderHoeven J., Franchini G.,
Trimarchi C. and Paoletti E. (1995). Biological and immunogenic properties of
a canary-rabies recombinant, ALVAC-RG (vCP65) in non-avian species. Vaccine
13: 539-549.
Thanavala Y., Yang Y.F., Lyons P., Mason H.S. and Arntzen C. (1995).
Immunogenicity of transgenic plantderived hepatitis B surface antigen. Proc.
Natl. Acad. Sci., U.S.A. 92: 3358–61.
Thoulouze M. I., Lafage M., Sahachmer M., Hartmann U., Cremer H. and Lafon
M. (1998). The neural cell adhesion molecule is a receptor for rabies virus. J.
Virol. 72: 7181-7190.
Tiwari S., Mishra D.K., Roy S., Singh A., Singh P.K and Tuli R. (2009a). High
level expression of a functionally active cholera toxin B: rabies glycoprotein
fusion protein in tobacco seeds. Plant Cell Rep. 28: 1827-36.
Tiwari S., Verma P.C., Singh P.K. and Tuli R. (2009b). Plants as bioreactors for the
production of vaccine antigens. Biotech. Adv. 27: 449–467.
Tonevitsky A., Toptygin A., Agapov I., Pfueller U. and Frankel A. (1994).
Renatured ricin toxin B chain made in Escherichia coli is soluble, stable and
biologically active. Biochem. Mol. Biol. Int. 32: 1139–1146.
Tordo N., Poch O., Ermine A. and Keith G. (1986a). Primary structure of leader
RNA and nucleoprotein genes of the rabies genome: segmented homology with
VSV. Nucleic Acids Research 14: 2671-2683.
Tordo N., Poch O., Ermine A., Keith G. and Rougeon F. (1986b). Walking along
the rabies genome: Is the large G-L intergenic region a reminant gene? Proc. Natl.
Acad. Sci., U.S.A. 83: 3914-3918.
Tordo N. and Poch O. (1988). Structure of rabies virus. In: Rabies, Campbell J.B. and
Charlton K.M. (Eds.), Boston, Kluwer Academic Press. 25-45.
Tordo N., Poch O., Ermine A., Keith G. and Rougeon F. (1988). Completion of the
rabies virus genome sequence determination: Highly conserved domains among
Page 176
Bibliography
112
the L (Polymerase) proteins of unsegmented negative–strand RNA viruses.
Virology 165: 565-567.
Tordo N., Bardane H., Bourhy H. and Sacramento D. (1993). Molecular
epidemiology of lyssaviruses. Focus on the glycoprotein and pseudogenes.
Onderstepoort Journal of Veterinary Research 60: 315-323.
Tordo N. and Kouknetzoff A. (1993). The rabies virus genome: an overview.
Onderstepoort Journal of Veterinary Research 60: 263-269.
Tregoning J.S., Nixon P., Kuroda H., Svab Z., Clare S. and Bowe F. (2003).
Expression of tetanus toxin Fragment C in tobacco chloroplasts. Nucleic Acids
Res 31: 1174–9.
Tsiang H., Koulakoff A., Bizzini B. and Berwald-Nettter Y. (1983). Neurotropism of
rabies virus.An in vitro study. J. Neuropath. Exp. Neurol. 42: 439-452.
Tsiang H. (1993). Pathophysiology of rabies virus infection of the nervous system.
Adv. Virus Research. 42: 375-412.
Tuboly T., Yu W., Bailey A., Degrandis S., Du S. and Erickson L. (2000).
Immunogenicity of porcine transmissible gastroenteritis virus spike protein
expressed in plants. Vaccine 18: 2023–8.
Tuchiya K., Matsuura Y., Kawai A., Ishihama A. and Ueda S. (1992).
Characterization of rabies virus glycoprotein expressed by recombinant
baculoviruses. Virus Res. 25: 1-13.
Tuffereau C., Leblois H., Benejean J., Coulon P., Lafay F. and Flamand A. (1989).
Arginine or lysine in Position 333 of ERA and CVS glycoprotein is necessary for
rabies virulence in adult mice. Virology 172: 206-212.
Tuffereau C., Benejean J., Blondel D., Kieffer B. and Flammand A. (1998a). Low–
affinity nerve growth factor receptor (P7 SNTR) can serve as a receptor for rabies
virus. EMBO J. 17: 7250-7259.
Tuffereau C., Benejean J., Roque Alfonso A.M., Flamand A. and Fishman M.C.
(1998b). Neronal cell surface molecules mediates specific binding to rabies virus
Page 177
Bibliography
113
glycoprotein expressed by a recombinant baculovirus on the surface of
lepiodopteran cells. J. Virol. 72: 1085-1091.
Udomsuk L., Jarukamjorn K., Tanaka H. and Putalun W. (2009). Isoflavonoid
production in a hairy roots culture of Pueraria candollei. Z. Naturforsch. C. 64:
687-91.
Ugolini G., Kuypers H.G.J.M. and Strick P.L. (1989). Transneuronal transfer of
herpes virus from peripheral nerves to cortex and brainstem. Science 243: 89-91.
Verma P.C. (2003). Biotechnological approaches for in vitro production of picroliv
from Agrobacterium rhizogenes mediated hairy root cultures of Picrorhiza
kurroa Royle ex Benth. PhD Thesis, Lucknow University, Lucknow.
Verma P.C., Trivedi I., Singh H., Shukla A.K., Kumar M., Upadhyay S.K.,
Pandey P., Hans A.L. and Singh P.K. (2009). Efficient production of gossypol
from hairy root cultures of cotton (Gossypium hirsutum L.). Curr. Pharm.
Biotechnol. 10: 691-700.
Vishwanath R.L., Katz F.N., Lodish H.F. and Blobel G. (1978). A signal sequence
for the insertion of a transmembrane glycoprotein. J. Biol. Chem. 253: 8667-
8670.
Vitetta E. and Yen N. (1990). Expression and functional properties of genetically
engineered ricin B chain lacking galactose-binding activity. Biochim. Biophys.
Acta. 1049: 151–157.
Von Teichman B.F., Thomson G.R., Meredith C.D. and Nel L.H. (1995). Molecular
epidemiology of rabies virus in South Africa: evidence for two distinct virus
groups. Journal of General Virology 76: 73-82.
Waheed M.T., Thönes N., Müller M., Hassan S.W., Gottschamel J., Lössl E., Kaul
H.P. and Lössl A.G. (2011). Plastid expression of a double pentameric vaccine
candidate containing human papillomavirus-16 L1 antigen fused with LTB as
adjuvant: transplastomic plants show pleiotropic phenotypes. Plant Biotechnol. J.
9: 651-60.
Page 178
Bibliography
114
Wales R., Richardson P., Roberts L., Woodland H. and Lord J. (1991). Mutational
analysis of the galactose binding activity of recombinant ricin B chain. J. Biol.
Chem. 266: 19172–19179.
Walmsley A.M. and Arntzen C.J. (2000). Plants for delivery of edible vaccines. Curr.
Opin. Biotechno. 11: 126-129.
Walmsley A.M., Alvarez M.L., Jin Y., Kirk D.D., Lee S.M. and Pinkhasov J.
(2003). Expression of the B subunit of Escherichia coli heat-labile enterotoxin as
a fusion protein in transgenic tomato. Plant Cell. Rep. 21: 1020–6.
Wang T., Chen J.P., Li H., Zhi K.Q., Zhang L., Yang C.L. and Tao D.C. (2005).
Co-expression and immunity of Legionella pneumophila mip gene and
immunoadjuvant ctxB gene. Acta. Biochim. Biophys. Sin. (Shanghai). 37: 199-
204.
Warner C.K., Schurr T.G. and Fekadu M. (1996). Molecular characterization of
carrier rabies isolates. Virus Research 41: 133-140.
Warzecha H., Mason H.S., Lane C., Tryggvesson A., Rybicki E. and Williamson
A.L. (2003). Oral immunogenicity of human papillomavirus-like particles
expressed in potato. J. Virol. 77: 8702–11.
Webster D.E., Cooney M.L., Huang Z., Drew D.R., Ramshaw I.A. and Dry I.B.
(2002) Successful boosting of a DNA measles immunization with an oral plant-
derived measles virus vaccine. J. Virol. 76: 7910–2
Webster D.E., Thomasc M.C., Huang Z. and Wesselingh S.L. (2005). The
development of a plant-based vaccine for measles. Vaccine 23: 1859–65.
Weiss R.A. and Bennet P.L.P. (1980). Assembly of membrane glycoproteins studied
by phenotypic mixing between mutants of vesicular stomatitis virus and
retroviruses. Virology 100: 252-274.
Whitt M., Buonocore L., Prehaud C. and Rose J. (1991). Membrane fusion activity,
oligomerization, and assembly of the rabies virus glycoprotein. Virology 185:
681-688.
Page 179
Bibliography
115
Wigdorovitz A.W., Carrillo C., Dus Santos M.J., Trono K., Peralta A., Gomez
M.C., Rios R.D., Franzone P.M., Sadir A.M., Escribano J.M. and Borca
M.V. (1999). Induction of a protective antibody response to foot and mouth
disease virus in mice following oral or parental immunization with alfalfa
transgenic plants expressing the viral structural protein VP 1. Virology 255: 347-
353.
Wigdorovitz A., Mozgovoj M., Dus Santos M.J., Parreño V., Gómez C. and Pérez-
Filgueira D.M. (2004). Protective lactogenic immunity conferred by an edible
peptide vaccine to bovine rotavirus produced in transgenic plants. J. Gen. Virol.
85: 1825–32.
Wiktor T., Fernodes M.V. and Koprowski H. (1964). Cultivation of rabies virus in
human diploid cell strain WI–38. J. Immunol. 93: 353-366.
Wiktor T., Lerner R. and Koprowski H. (1971). Inhibitory effect of passive antibody
on active immunity induced against rabies by vaccination. Bull. Wld. Hlth. Org.
45: 747-753.
Wiktor T., Gyorgy E., Schlumberger H.D., Sokol F. and Koprowski H. (1973).
Antigenic properties of rabies virus components. J. Immunol. 110: 269-276.
Wiktor T. and Koprowski H. (1978). Monoclonal antibodies against rabies virus
produced by somatic cell hybridization: detection of antigenic variants. Proc.
Natl. Acad. Sci., U.S.A. 75: 3938-3942.
Wiktor T. and Koprowski H. (1980). Antigenic variants of rabies virus. J. Exp. Med.
152: 99-112.
Wiktor T., MaCFarlan R.T., Reagan K., Dietzschold B., Curtis P.J., Wunner
W.H., Kieny M.P., Lathe R., Lecocg J.P., Mackett M., Moss B. and
Koprowski H. (1984). Protection from rabies by a vaccinia virus recombinant
containing the rabies virus glycoprotein gene. Proc. Natl. Acad. Sci., U.S.A.
81: 7194-7198.
Page 180
Bibliography
116
Wiktor T., Plotkin S. and Koprowski H. (1988). Rabies vaccine. In “Vaccines”,
Plotkin S. and Mortimer E. (Eds.), W.B. Saunders Company, Philadelphia,
London, Toronto. 474-491.
Witcher D.R., Hood E.E., Peterson D., Bailey M., Marshall L., Bond D., Ritland
D., Kusnadi A., Evangelista R., Nikolov Z., Wooge C., Mehigh R., Kappel B.,
Register J. and Howard J. (1998). Commercial production of -glucuronidase
(GUS): a novel system for the production of proteins in plants. Mol. Breed. 4:
301-312.
Woffenden B.J., Nopo L.H., Cramer C.L., Dolan M.C., Medina-Bolivar F. (2008).
Expression of a ricin B: F1: V fusion protein in tobacco hairy roots: steps toward
a novel pneumonic plague vaccine. Elect. Journ. Integrat. Biosci. 3: 10-19.
Wojczyk B., Shakin-Eshleman S.H., Doms R.W., Zhi-Quan X, Ertl. H.C.J.,
Wunner W.H. and Spitalnik S.L. (1995). Stable secretion of a soluble,
oligomeric form of rabies virus glycoprotein: influence of N-glycan processing on
secretion. Biochemistry 34: 2599-2609.
Wojczyk B.S., Stwora-Wojczyk M., Shakin-Eshleman S., Wunner W.H. and
Spitalnik S.L. (1998). The role of site-specific N-glycosylation in secretion of
soluble forms of rabies virus glycoprotein. Glycobiology 8:121-30.
Wongsamuth R. and Doran P.M. (1997). Hairy roots as an expression system for the
production of antibodies. In: Doran P.M. (Ed.), Hairy roots: culture and
application. Amstardam: Harwood Academic. 89-97.
World Health Organisation (WHO), 1984. Expert Committee on Rabies, 7th
Report.
WHO Tech. Rep. Ser. 709:9.
World Health Organisation (WHO), 1992. Expert Committee on Rabies. VIII
Report, WHO Technical Report. WHO Tech. Rep. Ser. 824.
World Health Organisation (WHO), 1994. Report of a WHO Workshop on
Prevention and Control of Rabies in Baltic Countries. Tallinn, Estonia, February
14-15. WHO/CDS/VPH/94.133. WHO, Geneva. 17.
Page 181
Bibliography
117
World Health Organisation (WHO), 1996. The World Health Report 1996. Fighting
disease Fostering development. WHO, Geneva. 57-58.
Wu Y.Z., Li J.T., Mou Z.R., Fei L., Ni B. and Geng M. (2003). Oral immunization
with rotavirus VP7 expressed in transgenic potatoes induced high titers of
mucosal neutralizing IgA. Virology 313: 337–42.
Wunner W.H., Curtis P.J. and Wiktor T.J. (1980). Rabies mRNA translation in
Xenopus laevis oocytes. J. Virol. 36: 133-142.
Wunner W.H., Reagan K.J. and Koprowski H. (1984). Characterization of saturable
binding sites for rabies virus. J. Virol. 50:691-697.
Wunner W., Dietzschold B., Smith C., Lafon M. and Golub E. (1985a). Antigenic
variants of CVS rabies virus with altered glycosylation sites. Virology 140: 1-12.
Wunner W., Dietzschold B., MaCfarlan R., Smith C., Golub E. and Wiktor T.
(1985b). Localization of immunogenic domains on the rabies virus glycoprotein.
Ann. Inst. Pasteur. 136E: 353-362.
Wunner W., Larson J.K., Dietzschold B. and Smith C. (1988). The molecular
biology of rabies viruses. Rev. Infect. Dis. 10:771-783.
Wunner W. (1991). The chemical composition and structure of rabies viruses. In: The
Natural History of Rabies, Baer G.M. and Boca Raton F.L. (Eds.), CRC Press.
31-67.
Xiang Z., Spitalnik S., Tran M., Wunner W., Chen J. and Ertl H. (1994).
Vaccination with a plasmid vector carrying the rabies virus glycoprotein gene
induces protective immunity against rabies virus. Virology 199: 132-140.
Xiang Z., Kwowles B., McCarrick J. and Ertl H. (1995b). Immune effector
mechanisms required for protection to rabies virus. Virology 214: 398-404.
Xiang Z., Spitalnik S., Cheng J., Erikson J., Wojczyk B. and Ertl H. (1995a).
Immune responses to nucleic acid vaccines to rabies virus. Virology 209: 569-
579.
Page 182
List of Publications
Page 183
Publications
119
List of publications
1. Singh A, Verma PC, Mishra DK, Srivastava S, Chouksey A, Roy
S, Singh
PK,
Saxena G and Tuli R. (2013) Expression of Rabies glycoprotein and Ricin toxin B
chain (RGP-RTB) fusion protein in tomato hairy roots: A step for Oral Vaccination
for rabies. Journal of Biotechnology (communicated)
2. Singh A, Yadav D, Rai KM, Srivastava M, Verma PC, Singh PK and Tuli R. (2012)
Enhanced expression of rabies virus surface G-protein in E. coli using SUMO fusion.
Protein J; 31: 68-64.
3. Roy S, Tyagi A, Tiwari S, Singh A, Singh PK, Sawant SV and Tuli R. (2010) Rabies
glycoprotein fused with B subunit of cholera toxin is expressed at high level in
tobacco plants and folds into biologically active pentameric protein. Protein Expr
Purif; 70(2): 184-90.
4. Tiwari S, Mishra DK, Roy S, Singh A, Singh PK, Tuli R. (2009) High level
expression of a functionally active cholera toxin B: rabies glycoprotein fusion protein
in tobacco seeds. Plant Cell Rep; 28(12):1827-36.