1993;53:2919-2926.Cancer Res Gene G. Finley, Raymond A. Koski, Mona F. Melhem, et al. and Colorectal AdenocarcinomaExpression of the Gastrin Gene in the Normal Human Colon
Updated Version http://cancerres.aacrjournals.org/content/53/12/2919
Access the most recent version of this article at:
Expression of the Gastrin Gene in the Normal Human Colon and ColorectalAdenocarcinoma1
Gene G. Finley, Raymond A. Koski, Mona F. Melhem, James M. Pipas, and Arnold I. Meisler2
Departments of Medicine ¡C.G. F., A. I. M.] and Pathology ¡M.F. M.¡.University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania 15261; Department of BiologicalSciences ¡J.M. P.I, University of Pittsburgh, Pittsburgh, Pennsylvania 15260: Molecular ami Cellular Biology Department ¡R.A. KJ, Amgen, Inc., Thousand Oaks. California
9I32Ü
ABSTRACT
Gastrin, produced in the G-cells of the gastric antrum and regulating
acid secretion in the stomach, also acts as a trophic factor in the gastrointestinal tract. Because of its possible role in colon cell proliferationand differentiation, evidence for its presence in normal colorectal mucosaand adenocarcinoma was sought. Utilizing tumors and matched normalmucosa from 26 patients, mature gastrin and progastrin were studied byimmunohistochemistry. In normal colonie mucosa! crypts, occasional cellsstained concordantly for gastrin, progastrin, and chromogranin A, suggesting that they are of neuroendocrine origin. Adenomatous polypsstained neither for gastrin nor chromogranin A. In 22 of 23 adenocarci-
nomas, more than 50% of tumor cells stained for gastrin and progastrin.The expected gastrin transcript was demonstrable by polymerase chainreaction and RNase protection in tumors and by polymerase chain reaction in normal mucosa. Its identity was confirmed by sequencing thepolymerase chain reaction product. A larger transcript containing IntronII was present in both cancers and normal mucosa but was barely discernible in the gastric antrum. Aberrant expression of gastrin may contribute to deregulated proliferation of many colorectal carcinomas.
INTRODUCTION
The treatment of disseminated colorectal cancer is unsatisfactoryand patients with this disease almost invariably succumb. Over theyears a wide variety of antineoplastic chemotherapeutic agents, surgical procedures, and radiation therapy has been utilized therapeuti-
cally with only modest success in stabilizing the disease, without clearevidence that life has been consistently prolonged. These modalitieshave been reviewed recently (1). While investigative work in theseareas continues, the advances in cell and molecular biology presentopportunities for developing alternative therapies. An array of proliferation-promoting peptides appears to play a crucial role in the growth
and differentiation of the normal colon. Selective inhibition of one ormore of these may provide a novel and effective mode of treatment.
Growth factors mediating cell proliferation within particular organsmay be identified in tumors removed from these organs. Tumorsobtained at surgery are readily available, often in relatively largeamounts for direct study or for establishing cell lines which canfacilitate identification of growth factors. Thus, from colorectal cancers and cell lines derived from them, mRNAs encoding a variety ofgrowth promoting peptides have been identified, including TGF-a3
and -ß,EOF, gastrin, insulin-like growth factors I and II, PDGF, andfibroblast growth factor (2-4). Moreover, receptors such as c-erbB-\,c-erbB-2, the PDGF receptor, and the insulin-like growth factor I
receptor have been shown to be present, suggesting the existence ofautocrine loops (5-8). Paracrine stimulation may also promote growth
Received 8/18/92: accepted 4/12/93.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by a Merit Review Grant from the Veterans Administration and by
Amgen. Inc.- To whom requests for reprints should be addressed at VA Medical Center, University
Drive C. Pittsburgh. PA 15240.*The abbreviations used are: TGF. transforming growth factor; cDNA. complementary
in colorectal cancers and normal mucosa. For example, proliferationfactors may be produced by the sparse but ubiquitous neuroendocrinecells dispersed throughout the gastrointestinal tract (9-11). Pericryptal
fibroblasts located in the lamina propria adjacent to the crypt (12) oreven lymphoid cells diffusely present throughout the intestine (13)may be sources of growth and differentiation factors.
Gastrin, a peptide hormone produced by the G-cells of the gastric
antrum, was initially thought to control only acid secretion by thestomach. The structure of the peptide and its mRNA and genomicsequence have been established (14, 15). In recent years it has beensuggested that gastrin might also function as a trophic factor for theentire gastrointestinal mucosa. This role is clearly separable from itseffect on acid secretion and is summarized in recent reviews (3, 16,17).
Gastrin expression has been detected in colon cancer cell lines (18,19). Most of the gastrin studies on colon carcinoma cell lines testedthe effects of adding gastrin to medium and assessing cell proliferation(18, 20-23). The results have varied. In a recent study, four different
human colon cancer cell lines were tested for growth stimulation byadded pentagastrin and only one showed growth stimulation (24).Such experiments are difficult to interpret. Since gastrin or an analogue is usually added to the medium, lack of a proliferative responsemay indicate that the peptide does not act as an effector for proliferation, that the receptor for gastrin is absent, or that the cells are alreadyproducing gastrin. Quantitative PCR measurements detected gastrinmRNA in several colon cancer cell lines (25).
Whether gastrin is involved in the pathogenesis and progression ofcolon cancer remains unclear. Watson et al. (26), using flow cytomet-ric detection with an anti-gastrin 17 antibody, demonstrated the pres
ence of intracellular gastrin in 6 of 28 human colorectal tumors. Inthese tumors, more than 20% of the cells stained positively. Thegastrin peptide and gastrin-like intermediates have also been found in
neural tumors (27, 28), pituitary tumors (29), gastrinomas (30), lungtumors (31), and certain endocrine tumors (32, 33). Conflicting datahave been published as to whether blood levels of gastrin are elevatedin colorectal cancer (34. 35).
Limited data are available to support a role for gastrin as a positivegrowth stimulus for colorectal carcinoma in vivo and these have beenreviewed (17). In this report, we provide additional evidence thatgastrin may be an effector for proliferation, and perhaps differentiation, in both normal colon and colorectal tumors. We demonstrate thatprogastrin and gastrin are present in neuroendocrine cells of normalcolorectal mucosa and that colorectal tumors comprise cells that commonly express gastrin.
MATERIALS AND METHODS
Immunohistochemistry. Human colorectal surgical specimens were fixedin formalin, embedded in paraffin, and cut into 4-/xm sections. Representative
samples of normal mucosa and tumors were evaluated by routine pathologicalstudies and these samples were also stained with an indirect immunoperoxidasemethod (36) using Vectastain Elite ABC Kits (Vector Laboratories. Burlin-game. CA) with 3,3'-diaminobenzidine substrate and Mayer's hematoxylin
counterstain. Polyclonal rabbit anti-human progastrin antibody (CambridgeResearch Biochemicals, Wilmington. DE) directed against the peptide Ser-Ala-
Glu-Asp-Glu-Asn (amino acids 96-101 of preprogastrin) was used at a 1:2000
dilution. This peptide is cleaved off progastrin during normal processing and isnot present in biologically active G-34 and G-17 gastrins. Polyclonal rabbitanti-human gastrin-17 antibody and polyclonal normal rabbit IgG were ob
tained from DAKO (Carpenteria, CA) and monoclonal antibody against chro-
mogranin A was obtained from Boehringer Mannheim Biochemicals (Indianapolis, IN). These were used as recommended by the manufacturers. Sectionsof paraffin-embedded gastric antrum were obtained from DAKO and used as
positive controls.RNA Preparation and Northern Analysis. Tumors and normal tissues
were obtained from patients undergoing surgery for colorectal carcinoma. Themucosa of normal colon was stripped from the underlying muscular layer byblunt dissection. Nonnecrotic specimens of tumor were used in their entirety.Admixture with mononuclear cells was minimal for normal mucosa but variable for tumors, certainly less than one-third of the cells in tumor specimens.
Tissue was rapidly frozen in liquid nitrogen for later use. Whole cellular RNAwas prepared by centrifugaron of guanidine isothiocyanate homogenatesthrough a CsCI cushion according to methods described by Chirgwin el al.(37). RNA was enriched for polyadenylated message by oligodeoxythymidylicacid-cellulose column chromatography.
Detailed descriptions of denaturing agarose gel electrophoresis, transfer toGene Screen membranes (Dupont NEN, Boston, MA), fixation by UV cross-
linking, prehybridization, probe labeling, hybridization, stringency washes,and autoradiography have been reported (38). The probe used for these studies.pHG529, a full length gastrin cDNA clone, was kindly provided by Dr. JensVuust (University Aarhus, Denmark) (14). The entire plasmid was radiolabeledby nick translation to a specific activity of IO8-IO9 cpm//j,g.
Polymerase Chain Reaction. First strand cDNA was prepared from oligodeoxythymidylic acid-primed whole cellular RNA from tumor and normal
tissues using Moloney Murine Leukemia Virus reverse transcriptase (GibcoBRL, Gaithersburg, MD). This cDNA was used as a template in PCR. GastrincDNA was amplified from base 4 to 288 with gastrin specific primers (forwardprimer: G[GAATTC]CAGCGACTATGTGTGTATGTG; reverse primer: C-|GGATCC]ACTGCGGCGGCCGAAGTCCAT). Brackets indicate EcoRl-(GAATTC) and flc/mHHGGATCC) restriction sites inserted into the 5' ends of
the primers to facilitate cloning. Reaction conditions were as follows: 5-10 ngof reverse transcribed cDNA prepared from whole cellular RNA or 5-20 pg oflinearized plasmid DNA; 50 mm KC1; 1-2 m.MMgCI2; 10 mm Tris-HCl, pH 8.3;0.02% gelatin; 200 JHMeach deoxynucleotide triphosphate; 1 ^M each deoxy-
nucleotide triphosphate; I JJ.Meach primer; 2.5 units Taq polymerase (PerkinElmer Cetus, Norwalk, CT). The thermal cycler was programmed as follows:denaturation, 94°Cfor 1 min; reannealing, 50°Cfor 1 min; extension, 72°Cfor
1 min. The reaction were carried out for 40 cycles and the products weredirectly analyzed on \% agarose, 3% Nu-Sieve (FMC Bioproducts, Rockland,ME), in 1 x TAE buffer (40 mm Tris-acetate-1 mm EDTA) and stained with
ethidium bromide. Because of the sensitivity inherent in the PCR reaction,great care was taken to avoid contamination of the colon specimens with
gastrin PCR products (39).Analysis of PCR Products. For Southern hybridization, PCR products
were separated in agarose gels, denatured, and blotted onto Gene Screen Plusfilters. Hybridization conditions and stringency washes were as previouslyreported (38). Gastrin cDNA plasmid pHG529 was used as a hybridizationprobe. Autoradiograms were typically exposed overnight at -70°C, using Du
pont intensifying screens and Kodak X-OMAT AR film.
The major PCR products from colon tumor and normal mucosa were gel
purified and cloned into M13 mp 8 and 9. Recombinant plaques detected bycolor selection were isolated. DNA sequencing of the cloned PCR productswas carried out using the dideoxy method described by Sanger (40) withSequenase Version II (U.S. Biochemicals, Cleveland, OH).
Kihonuclea.se Protection Analysis. The probe used for these experimentswas derived from the 299 bp amplified gastrin fragment generated by PCR.This PCR product was purified, restricted with ¿.VuRIand H<im\\\, and clonedinto pOKM 3 and 4 vectors, producing a source of both control gastrin senseRNA and a continuously labeled antisense RNA probe which were used in thenuclease protection assay. The assay was performed by a modification of themethod of Berk and Sharp (41).
Up to 10 fig of polyadenylated RNA or 1-10 ng of control gastrin senseRNA was mixed with 5 X K)4cpm of the continuously labeled antisense probe.
Following hybridization overnight, the mixture was subjected to digestion with
RNase A and Tl. The resultant products were analyzed on a denaturingpolyacrylamide/urea gel.
RESULTS
Immunohistochemical Demonstration of Gastrin and Progastrin in Colorectal Adenocarcinoma and Normal Mucosa. The human preprogastrin peptide comprises 101 amino acids including: a 19amino acid NH2-terminal signal peptide sequence; a 37 amino acid
spacer region; a sequence containing the major bioactive forms ofgastrin (gastrin 34 and gastrin 17); and a COOH-terminal flanking
peptide (Fig. 1). The molecule undergoes extensive posttranslationalprocessing (42) to yield: (a) progastrins extended beyond glycine atCOOH-terminus; (b) glycine-extended progastrins, the immediate
precursors of mature gastrins; and (c) amidated, biologically activegastrins in which the COOH-terminal phenylalanine residue is ami-
dated using glycine as the amide donor. In gastrins and progastrins,tyrosine 87 has variable degrees of sulfation (43). In the gastricantrum and duodenum about 50% of the gastrin molecules are sul-fated. The antibodies used in this study include rabbit anti-humangastrin-17 and anti-progastrin antibodies directed against different
regions of the gastrin primary translation product (Fig. 1).Previous gastrointestinal immunohistochemical studies localized
gastrin and progastrin to gastric antrum G-cells and duodenal neu-
roendocrine cells (44, 45). Consistent with these reports, we findstrong immunohistochemical staining of gastric antrum G-cells withboth anti-gastrin and anti-progastrin antibodies (Fig. 2). This staining
is absent when a negative control rabbit IgG antibody is used (data notshown).
In normal colonie mucosa, serial 4-ju.m sections demonstrate that
occasional crypt cells stain concordantly for gastrin (Fig. 3a) andprogastrin (data not shown). Staining is mostly cytoplasmic and wasabsent when nonimmune rabbit IgG was used as a negative controlantibody. In Fig. 3, b and c, successive sections were stained forgastrin and human chromogranin A, a neuroendocrine marker, suggesting that the cells containing gastrin are of neuroendocrine origin.All cells which stained for gastrin also stained for chromogranin A.but the converse is not true. All normal mucosa studied were frompatients who had a neoplasm somewhere in the colon or rectum. Thespecimens were at least 5 cm from the border of the tumor. We havenot yet had an opportunity to study normal colon from a normalpatient.
In contrast to the infrequent staining of normal colorectal mucosalcells, we found that in 22 of 23 colorectal adenocarcinomas, more than80% of the tumor cells stained for gastrin (Table 1). Of these 23tumors, 20 were also stained for progastrin and all of these werepositive. As in the case of normal mucosa, there was concordantcellular staining for gastrin and progastrin in the same cells. Fig. 4shows a typical example of a colonie tumor stained for gastrin andprogastrin. Staining is predominantly cytoplasmic. Virtually everytumor cell stains, but the intensity of staining may vary betweentumors and within a given tumor. In Fig. 4«,the staining is diffuse anduniform throughout as is true for the progastrin staining shown in Fig.4b. This was not always the case, and in some tumors the pattern ofstaining was more heterogeneous. In these tumors some cells did notstain at all and variation in staining intensity was seen from one sectorof the tumor to another (data not shown). Nevertheless, in no casewere less than 50% of the cells stained. A negative control utili/ingrabbit IgG showed no staining (Fig. 4c). Interestingly, although notalways the case, the majority of these tumors which stained positivelyfor gastrin showed no staining for chromogranin A, including thematched tumor of the chromogranin A positive normal mucosa shownin Fig. 3/>.
poly AFig. l. Human gastrin cDNA sequence. Oligonucleotide primers are shown with brackets indicating restriction enzyme recognition sites added to facilitate cloning, ¿VijRl-
IGAATTC]. ßamHI[GGATCC]. Hindììlrestriction sites in the native sequence are indicated, ßfur.v.dibasic processing sites for trypsin-like proteases which cleave the preprogastrin.The circlet!amino acids in the gastrin-17 sequence are homologous to the region surrounding tyrosine 3I5 in polyoma virus middle T-antigen. The tyrosine residue in gastrin-17 {doublecircle) K the site of sull'ation and in \-iiro phosphorylation.
Because of a lack of chromogranin A staining in tumors, we examined staining for chromogranin A and gastrin in six adenomatouspolyps. These polyps varied in size from less than I cm to more than2 cm. No evidence of positive staining was detected (data not shown).Normal mucosa stains for chromogranin A and gastrin. polyps stainfor neither, and the tumors we examined stained positively for gastrinand negative for chromogranin A. As is evident from Table I. gastrinstaining correlated neither with the degree of tumor differentiation norDukes stage. Normal mucosa directly adjacent to tumors showed noevidence of increased staining (data not shown).
Polymerase Chain Reaction of Gastrin mRNA Transcripts inAdenocarcinoma and Normal Mucosa. Ten tumors and theirmatched normal mucosa were assayed for gastrin mRNA by PCR. Thepredicted gastrin cDNA PCR product based on the primers used was299 base pairs (Fig. I ). In fact, PCR amplified a 299-base pair product, as well as a larger 428-base pair product from all 20 colorectal
specimens (Fig. 5). The larger product could barely be detected incDNA reverse transcribed from gastric antrum and not at all in clonedgastrin cDNA, nor from RNA samples that were not reverse transcribed (Fig. 5; data not shown). The yield of the 299-base pair PCR
product is consistently higher from tumor tissue than from matchednormal mucosa (Figs. 5 and 6). To confirm that the PCR reactionproducts from the colon were indeed gastrin related, they were subjected to Southern blot analysis shown in Fig. 6. These results indicatethat gastrin-related mRNAs are present in both normal and neoplastic
colonie tissue.
To further characterize the colonie gastrin PCR products, the 299-and 428-base pair DNAs from colonie mucosa and from a colon
adenocarcinoma were gel purified, cloned into M13 mp 8 and 9, andsequenced. This revealed the 299-base pair fragment to be identical in
sequence to the published gastrin cDNA sequence (14). The longercDNA was also encoded by the human gastrin gene but contained the129-base pair Intron II sequence indicated in Fig. 1. This larger
product apparently is amplified from unspliced gastrin mRNA presentin colonie mucosa and tumors but largely absent from gastric antrum.It is unlikely to be due to genomic DNA contamination of the colonieRNA used to prepare cDNA for PCR since the RNA used as a targetin PCR failed to produce any visible product without reverse transcription.
Direct Detection of Gastrin mRNA in Colon Tumors by Ribo-
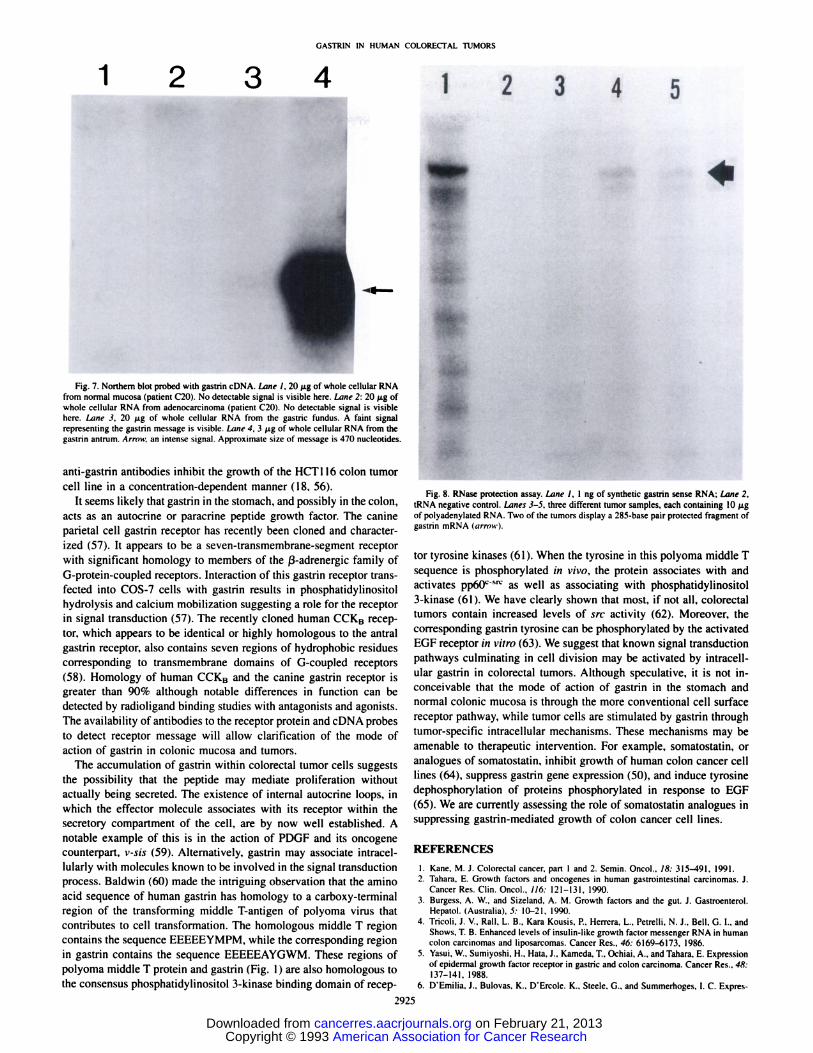
nuclease Protection. Despite numerous attempts. Northern blot analysis of colorectal carcinomas and normal mucosal tissue utilizing upto 20 ju,gof whole cellular RNA fai led to reveal the presence of gastrinmRNA although the transcript was easily demonstrable in RNA prepared from the gastric antrum (Fig. 7).
Ribonuclease protection assays provide considerably more sensitivity in detecting mRNA than Northern hybridization analysis (41)and are more quantitative than PCR amplification. Four colorectaltumors were assayed for the presence of gastrin mRNA by this method. Ten /ig of polyadenylated mRNA were used from each specimen.For two of these tumors, sufficient corresponding normal mucosa wasavailable to provide RNA for the assay. Neither of the normal mucosal
Fig. 2. Immunohistochemical staining of the normal gastric antral mucosa ( X400). a, staining with anti-gastrin antibody, b, same tissue stained with anii-progastrin antibody showingidentical staining pattern to a.
samples showed a protected band, but two of the tumors containeddetectable message (Fig. 8).
DISCUSSION
Our results support the conclusion that gastrin is present in normalcolonie mucosa and in colorectal tumors. The concordant staining forgastrin and progastrin in the same cells allows us to infer gastrinsynthesis in situ. Moreover, the presence of the progastrin hexapeptideSAEDEN (amino acids 96-101) serves to distinguish gastrin from the
physiologically active cholecystokinin octapeptide, also produced inthe gastrointestinal tract. This is of concern since the gastrin antibodywe have utilized cross reacts with cholecystokinin (46).
The coincident staining for gastrin and chromogranin A in normalcolonie mucosa strongly suggests that gastrin expression in coloniecrypts is restricted to neuroendocrine cells (28). In the common co-
Ionic neoplasms, adenomatous polyps and adenocarcinomas. theformer stain neither for gastrin nor chromogranin A while the latterstain for gastrin but frequently not for chromogranin A. This suggeststhat increased gastrin expression occurs at a late stage in malignant
Fig. 3. Immunohistochemical staining of normalcolonie mucosa ( X 4(X)). a, colonie mucosa stainedwith anti-gastrin antibody showing occasional cellstaining positively, b and c. consecutive sectionsstained with anti-gastrin and anti-chromogranin Aantibodies, respectively. The coincident stainingpattern suggests that the gastrin-containing cells are
" WDA. well differentiated adenocarcinoma: MWDA. moderately well differentiated
udenocarcinoma: PDA. poorly differentiated adenocarcinoma.h Intensity of stained cells, in different areas of the tumor section on a scale of 0
(negative) to 4+. ND. not done.' D. dead; A&W, alive and well; R. relapse; P. perioperative death within 6 weeks of
surgery; M. months.
transformation of colonie epithelial cells. This observation furthersuggests that the origin of the adenomatous polyp is not from neu-roendocrine cells. Finally, PCR-detection of gastrin mRNA in normal
mucosa and colorectal adenocarcinomas, much more abundant in thelatter than in the former, and sequencing of the PCR products providesadditional evidence for gastrin expression in these tissues. The ribo-
nuclease protection assay confirms this conclusion.Gastrin gene expression is regulated by multiple mechanisms. The
gastrin promoter contains negative and positive elements to whichfra/u-acting protein factors bind (47, 48). TGF-a, EGF (9) and gas
trointestinal peptides such as somatostatin (49, 50) and bombesin/gastrin releasing peptide (51) appear to be mediators of gastrin transcription and release in the gastric antrum. TGF-a. which is expressed
in most colorectal carcinomas (52) may promote gastrin gene expression in these tumor cells. It is also clear that gastrin mRNA processingis altered in the colon and colonie tumors since the large PCR productfrom unspliced gastrin mRNA precursors is barely detectable in theantrum. Colorectal tumor cells, although the majority contain gastrin,have low levels of gastrin mRNA. In contrast, the gastric antrum,while comprising about 30% G-cells (53), produces enough gastrin
mRNA to be seen on Northern analysis. The accumulation of gastrinin tumor cells suggests more efficient gastrin mRNA translationand/or less efficient secretion of gastrin than in G-cells.
The precise role of gastrin in colorectal cancer remains unclear.Table 1 shows that most adenocarcinomas contain gastrin immunore-
active protein irrespective of Dukes stage, morphology, or clinicaloutcome. Further, gastrin staining does not correlate with patient age,tumor size, or location. It is unlikely that an intestinal hormone asphysiologically active as gastrin occurs in 22 of 23 colorectal tumorsby chance alone and exerts no effect on tumor cell growth. Gastrinmay act in a permissive capacity, necessary but insufficient for tumordevelopment and progression. In the normal colon it seems likely thatthe neuroendocrine cells, acting in a paracrine manner, release gastrinand other peptides that play a role in the proliferation and differentiation of adjacent crypt cells. Studies using exogenously administered
B
Fig. 4. Immunohistochemical staining of adenocarcinomas (X 400). a, moderately welldifferentiated adenocarcinoma of the colon (patient C13) stained with the anti-gastrinantibody showing cytoplasmic homogeneous staining in all tumor cells. A rare interstitialfibroblast is also positive, b, consecutive section of the same tumor stained with anti-
progastrin. Note the identical staining pattern within the same tumor, c. negative control,a consecutive section to that seen in a and b. Only rare nonspecific positively is seen ininterstitial inflammatory cell with normal rabbit IgG antibody.
gastrin strongly suggest a role for gastrin in the growth of normalcolonie mucosa (16, 17) which can be abrogated using a gastrinantagonist, proglumide (54. 55).
Colon tumor cell studies support an autocrine growth mediatingrole for gastrin. Hoosein. et al. (18) showed that the HCT116 coloncancer cell line contained mRNA that hybridized to a gastrin-specific
probe, although the transcripts were much longer than those anticipated for gastrin mRNA. In another study. Baldwin, et al. (19) convincingly demonstrated gastrin mRNA in the human Okajima andHCT116 cell lines. Recently, they have demonstrated gastrin mRNAin seven additional cell lines (25). Moreover, both proglumide and
Fig. 5. Gastrin PCR products from colorectal tissues, a. Lane I, positive control; PCR product from the linearized plasmid pHG529 which contains the full length human gastrincDNA. This lane demonstrates the predicted 299-base pair hand (middle arrow). Lanes 2 and 3, duplicate lanes demonstrating the amplification products from u human colon carcinomacDNA. There is a prominent hand at 299 hase pairs (middle arrow) and a weaker hand about 428 hase pairs in length (top arrow]. The larger product was not seen in the positive control.Lane 4, negative control; no added template DNA. h, PCR products from normal colonie mucosa. IMHC¡,positive control; PCR product from the lineari/ed plasmid pHG529 whichcontains the full length human gastrin cDNA. This lane demonstrates the predicted 299-base pair band. Lane 2, negative control; no added template DNA. l¿mes3 and 4. duplicate
lanes demonstrating the amplification products from normal colonie mucosal cDNA. There are products of 299 and about 428 base pairs in length. The larger product was not seenin the positive control (¡MueI ). In both a and h a primer dimer band is visible at the bottom of the gel (bottom arrow).
1 8Fig. 6. Southern blot analysis of gastrin PCR products from three adenocarcinomas and matched normal mucosa. Lane I, gastric mucosal cDNA, approximately 1 ng utilized as a
target in the PCR reaction. Note predominate signal at 299 base pairs (bottom arrow) and faint signal at 428 base pairs (lop arrow). ¡Mne2, positive PCR control utili/ing of 20 pgof linearized gastrin cDNA plasmid (pHG529), demonstrating the single 299-base pair product. IMHC3, normal mucosa showing predominately the 428-base pair product. The 299-hasepair fragment is poorly seen; iMne 4, adcnocareinoma from patient whose normal mucosa is shown in Lane 3. The signals from both the 299- and 428-base pair fragment are easilyseen. I¿me5 and 6, normal mucosa and tumor, respectively, from a patient. Signals from both products are easily seen in both tissues. Lane 7 and fi, normal mucosa and tumor,respectively from a patient. The 299-base pair product is poorly seen in the normal mucosa and the 428-base pair product is poorly seen in the tumor Lanes 3-fi utilized approximately5 ng of cDNA for each amplification reaction. Ten % of the amplified product was run on the gel in each instance (Lanes 1-8).
Fig. 7. Northern blot probed with gastrin cDNA. Lane I, 20 /¿gof whole cellular RNAfrom normal mucosa (patient C20). No detectable signal is visible here. iMne 2: 20 /Agofwhole cellular RNA from adenocarcinoma (patient C20). No detectable signal is visiblehere. bine 3, 20 ng of whole cellular RNA from the gastric tundus. A faint signalrepresenting the gastrin message is visible. Lane 4. 3 (ig of whole cellular RNA from thegastrin antrum. Arnt\\: an intense signal. Approximate size of message is 470 nucleotides.
anti-gastrin antibodies inhibit the growth of the HCT116 colon tumorcell line in a concentration-dependent manner (18. 56).
It seems likely that gastrin in the stomach, and possibly in the colon,acts as an autocrine or paracrine peptide growth factor. The canineparietal cell gastrin receptor has recently been cloned and characterized (57). It appears to be a seven-transmembrane-segment receptorwith significant homology to members of the ß-adrenergicfamily ofG-protein-coupled receptors. Interaction of this gastrin receptor trans-fected into COS-7 cells with gastrin results in phosphatidylinositol
hydrolysis and calcium mobilization suggesting a role for the receptorin signal transduction (57). The recently cloned human CCKB receptor, which appears to be identical or highly homologous to the antralgastrin receptor, also contains seven regions of hydrophobic residuescorresponding to transmembrane domains of G-coupled receptors
(58). Homology of human CCKB and the canine gastrin receptor isgreater than 90</{-although notable differences in function can be
detected by radioligand binding studies with antagonists and agonists.The availability of antibodies to the receptor protein and cDNA probesto detect receptor message will allow clarification of the mode ofaction of gastrin in colonie mucosa and tumors.
The accumulation of gastrin within colorectal tumor cells suggeststhe possibility that the peptide may mediate proliferation withoutactually being secreted. The existence of internal autocrine loops, inwhich the effector molecule associates with its receptor within thesecretory compartment of the cell, are by now well established. Anotable example of this is in the action of PDGF and its oncogenecounterpart, v-sis (59). Alternatively, gastrin may associate intracel-
lularly with molecules known to be involved in the signal transductionprocess. Baldwin (60) made the intriguing observation that the aminoacid sequence of human gastrin has homology to a carboxy-terminalregion of the transforming middle T-antigen of polyoma virus that
contributes to cell transformation. The homologous middle T regioncontains the sequence EEEEEYMPM, while the corresponding regionin gastrin contains the sequence EEEEEAYGWM. These regions ofpolyoma middle T protein and gastrin (Fig. I ) are also homologous tothe consensus phosphatidylinositol 3-kinase binding domain of recep-
Fig. 8. RNase protection assay. Lane 1, 1 ng of synthetic gastrin sense RNA; Lane 2.tRNA negative control. Lanes .ÃŽ-5.three different tumor samples, each containing 10 u.gof polyadenylated RNA. Two of the tumors display a 285-base pair protected fragment ofgastrin mRNA (tirnnv).
tor tyrosine kinases (61 ). When the tyrosine in this polyoma middle Tsequence is phosphorylated in vivo, the protein associates with andactivates pp60l SIVas well as associating with phosphatidylinositol
3-kinase (61 ). We have clearly shown that most, if not all. colorectal
tumors contain increased levels of src activity (62). Moreover, thecorresponding gastrin tyrosine can be phosphorylated by the activatedEGF receptor in vitro (63). We suggest that known signal transductionpathways culminating in cell division may be activated by intracell-
ular gastrin in colorectal tumors. Although speculative, it is not inconceivable that the mode of action of gastrin in the stomach andnormal colonie mucosa is through the more conventional cell surfacereceptor pathway, while tumor cells are stimulated by gastrin throughtumor-specific intracellular mechanisms. These mechanisms may be
amenable to therapeutic intervention. For example, somatostatin, oranalogues of somatostatin, inhibit growth of human colon cancer celllines (64). suppress gastrin gene expression (50). and induce tyrosinedephosphorylation of proteins phosphorylated in response to EGF(65). We are currently assessing the role of somatostatin analogues insuppressing gastrin-mediated growth of colon cancer cell lines.
REFERENCES
1. Kane. M. J. Coloreclal cancer, part I and 2. Semin. Oncol.. 18: 3I5^»9I, 1991.2. Tahara. E. Growth factors and oncogenes in human gastrointestinal carcinomas. J.
Cancer Res. Clin. Oncol.. 116: 121-131, 1990.3. Burgess. A. W., and Si/eland. A. M. Growth factors and the gut. J. Gastroenterol.
Hepatol. (Australia). 5: 10-21. 1990.
4. Tricoli. J. V.. Rail. L. B.. Kara Kousis. P., Herrera. L., Petrelli. N. J.. Bell. G. I., andShows. T. B. Enhanced levels of insulin-like growth factor messenger RNA in humancolon carcinomas and liposarcomas. Cancer Res., 46: 6169-6173, 1986.
5. Yasui. W.. Sumiyoshi. H.. Hata. J.. Kameda. T.. Ochiai. A., and Tahara. E. Expressionof epidermal growth factor receptor in gastric and colon carcinoma. Cancer Res.. 4fi:137-141. 1988.
6. D'Emilia. J.. Bulovas. K.. D'Ercole. K.. Steele. G.. and Summerhoges. I. C. Expres-
7. Ito. M., Yoshidu. K.. Kyo, E.. Ayhan. A., Nakayama. H.. Yasui. W.. Ito. H., and Tahara.E. Expression of several growth factors and their receptor genes in human coloncarcinomas. Virchow Archiv. B. Cell Pathol.. 5V: 173-178. 1990. 37.
8. Guo, Y. S.. Naragon, S.. Yallampalli. C.. and Singh. P.Characterization of insulin-likegrowth factor I receptor in human colon cancer. Gastroenterology. 102: 1101-1108.1992. 38.
9. Godley. J. M.. and Brand. S. J. Regulation of the gastrin promoter by epidermalgrowth factor and neuropeptides. Proc. Nati. Acad. Sci. USA. 86: 3036-3040. 1989.
10. Chejfec, G.. Capella. C.. Solcia, E.. Wellington. J.. and Gould. V. E. Amphicrine cells. 39.dysplasias and neoplasias. Cancer (Phila.). 56: 2683-2690. 1985.
11. Jansson,D.. Gould. V. E-, G(x»ch,G. T.. et al. Immunohistochemical analysis of coloncarcinomas applying exocrine and neuroendocrine markers. Acta Pathol. Microhiol.Immunol. Scand.. 96: 1129-39. 1988. 40
12. Akashi. M.. Yamato. K., and Koeffler. H. P.Hematopoietic growth factors: regulationof production. In: R. Mertelsmann and F. Herrmann (eds.). Hematopoietic Growth 41.Factors in Clinical Applications, pp. 41-61. New York: Marcel Dekker. Inc.. 1990.
13. Riecken. E. O., Stallmach. A.. Zcitz. M.. Schulzke. J. D.. Merge, H., and Gregor, M. 42.Growth and transformation of small intestinal mucosa: importance of connectivetissue gut associated lymphoid tissue and gastrointestinal regulatory peptides. Gut. 43.30: 1630-1640. 1989.
14. Boel. E., Vuust. J.. Norris. F.. et al. Molecular cloning of human gastrin cDNA: 44.evidence of evolution of gastrin by gene duplication. Proc. Nati. Acad. Sci. USA. 80:2866-2869. 1983. 45.
15. Wiborg. O., Berglund, L.. Boel. E., Norris. F.. Norri. K., Rehfeld. J. F.. Marcker. K.A., and Vuust. J. Structure of a human gastrin gene. Proc. Nati. Acad. Sci. USA. 8l:1067-1069. 1984. 46.
16. Johnson. L. R. Regulation of gastrointestinal mucosa! growth. Physiol. Rev.. 68: 47.456-501, 1988.
17. Mulholland, M. W., and Debas. H. T. Physiology and pathophysiology of gastrin: areview. Surgery (St. Louis). 10): 135-147. 1988. 48.
18. Hoosein. N. M.. Kiener. P.A.. Curry. R. C.. and Brattain. M. G. Evidence for autocrinestimulation of cultured colon tumor cells by a gastrin-cholecystokinin-like peptide.Exp. Cell Res.. 186: 15-21. 1990. 49.
19. Baldwin. G. S.. Casey. A.. Matanadiotis, T.. McBride, K.. Sizeland. A. M.. andThumwood. C. M. PCR cloning and sequenceof gastrin mRNA from carcinoma cell 50.lines. Biochem. Biophys. Res. Commun., 170: 691-697, 1990.
20. Sirinek, K. R., Levine. B. A., and Mayer, M. P.Pentagastrin stimulates in \-itn>growth 51.of normal and malignant human colon epithelial cells. Am. J. Surg.. 149: 35-39. 1985.
21. Kusyk. C. J., McNiel. N. O.. and Johnson, L. R. Stimulation of growth of a coloncancer cell line by gastrin. Am. J. Physiol.. 251: G597-G601. 1986. 52.
23. Smith, J. P.. and Solomon. T. E. Effects of gastrin. proglumide and somatosialin on 53.growth of human colon cancer. Gastroenterology. 95: 1541-1548. 1989. 54.
24. Eggstein. S.. Imdahl. A.. Kohler. M.. Waibel. M., and Farthmann. E. H. Influence ofgastrin, gastrin receptor blockcrs. epidermal growth factor, and difluoromethylomi- 55.
thine on the growth and the activity of ornithine decarboxylase of colon carcinomacells. J. Cancer Res. Clin. Oncol.. 'll7: 37-42, 1991.
25. Baldwin. G. S.. and Zhang. Q. X. Measurement of gastrin and transforming growth 56.factor-a messenger RNA levels in colonie carcinoma cell lines by quantilalive poly-merase chain reaction. Cancer Res., 52: 2261-2267, 1992.
26. Watson. S. A.. Durant. L. G.. Wencyk. P. M.. Watson. A. L.. and Morris. D. L. 57.Intracellular gastrin in human gastrointestinal tumor cells. J. Nail. Cancer. Inst.. 83:866-871, 1991.
27. Rehfeld. J. F., vanSolinge. W. W.. Tos. M., and Thomson. J. Gastrin. cholecystokinin 58.and their precursors in acoustic neuromas. Brain Res.. 530: 235-238. 1990.
28. Bevan. J. S.. Asa. S. L.. Rossi. M. L.. Esiri. M. M.. Adams. C. B., and Burke. C. W.Incasellar gangliocytoma containing gastrin and growih hormone-releasing hormone 59.associatedwith a growth hormone secreting piluilary adenoma. Clin. Endocrino!.. 30:213-224, 1989. 60.
29. Bardram. L., Lindholm. J.. and Rehfeld. J. F. GaMrin in pituitary tumors. AciaEndocrino!., 115: 419-422, 1987. 6l.
30. Wolfe. M. M., and Jensen. R. T. Zollinger-Ellison syndrome. N. Engl. J. Med., 317:1200-1209. 1987. 62.
31. Rehfeld. J. F., Bardram. L.. and Hilsied. L. Gastrin in human bronchogenic carcinomas: constant expression but variable pnx'essing of gastrin. Cancer Res.. 49: 2840- 63.
2843. 1989.32. Bardram. L.. Hilsted, L., and Rehfeld. J. F. Cholecyslokinin. gastrin and precursors in
pheochromocytomas. Acia Endocrino!., 120: 479-484, 1989. 64.33. Fabri. R. J., Gower. W. R.. Weber. C.. and Tuttle. S. Hypcrparathyroid glands contain
G-17 and G-34 gastrin. J. Surg. Res.. 41: 333-337. 1986. 65.34. Smith, J. P..Wood. J. G.. and Solomon, T. E. Elevated gastrin levels in patients with
colon cancer or adenomatous polyps. Dig. Dis. Sci.. 34: 171-174. 1989.35. Suzuki. H.. Malsumoto. K.. and Terashima. H. Serum levels of gastrin in patients with
colorectal neoplasia. Dis. Colon Recium. 31: 716-717. 1988.Hsu. S. M.. Raine, L., and Fanger. H. Use of avidin-biolin-peroxidase complex (ABC)in immunoperoxidase techniques: acomparison betweenABC and unlabeled antibody(PAP) procedures. J. Hislochem. Cylochem., 29: 577-580. 1981.Chirgwin, J. M.. Przybyla. A. E.. MacDonald. R. J.. and Rutter. W. J. Isolation ofbiologically active ribonucleic acid from sourcesenriched in ribonuclease. Biochemistry, 18: 5294-5299, 1979.Finley. G. G.. Schulz. N. T., Hill. S. A.. Geiser, J. R.. Pipas. J. M., and Meisler. A. I.Expresssion of ihe mvc gene family in differenl stages of human colorectal cancer.Oncogene. 4: 963-971. 1989.Ausubel. F. M.. Breni. R.. Kingston. R E.. Moore, D. D.. Seidman, J. G.. Smith. J.A., and Strahl. K. (eds.) Chapter 15. The polymerase chain reaction. In: CurrentProtocols in Molecular Biology. Suppl. 9. pp. Õ5.3.6-15.3.7.New York: John Wileyand Sons. 1989.Sanger. F.. Nicklen, S.. and Coulson. A. R. DNA sequencing with chain terminatinginhibitors. Proc. Nail. Acad. Sci. USA. 74: 5463-5467. 1977.Berk. A. J.. and Sharp. P.A. Sizing and mapping of early adenovirus mRNAs by gelelectrophoresis of SI endonuclease-digesied hybrids. Cell. 12: 721-732. 1977.Bardram. L.. Hilsted. L.. and Rehfeld. J. F. Progastrin expression in mammalianpancreas. Proc. Nail. Acad. Sci. USA. 87: 298-302. 1990.Anderson, B. Measurement of occurrence of sulfated gastrins. Scand. J. Clin. Lab.Invest. Suppl., 168: 5-24, 1984.Sjolund. K.. Sanden. G.. Hakanson. R.. and Sundler. F. Endocrine cells in humaninteslines: an immunocytochemical study. Gastroenterology. 85: 1120-1123. 1983.Hará.M., Varndell. I. M.. Bishop. A. E., et al. Expression of the C-lerminal flankingpeptide of human progastrin in human gastroduodenal mucosa.G-cell hyperplasia andislet cell lumors producing gasirin. Mol. Cell Probes. /: 95-108, 1987.Dakopatts Specification Sheet. Rabbit Anti-Human Gastrin.Wang. T. C.. and Brand. S. J. Islet cell-specific regulatory domains in Ihe gastrinpromoter contains adjacent positive and negative DNA elements. J. Biol. Chem.. 26:8908-8914. 1990.Merchant. J. L.. Demediek. B.. and Brand. S. J. A GC-rich element confers epidermalgrowih faclor responsiveness lo transcription from the gastrin promoter. Mol. Cell.Biol.. //: 2686-2696. 1991.Wolfe, M. M.. Reel. G. M.. and McGuigan, J. E. Inhibition of gastrin release bysomatostatin in cultured rat antral mucosa. J. Clin. Invest.. 72: 1586-1593. 1983.Karnik. P.S., Monahan. S. J.. and Wolfe. M. M. Inhibition of gastrin gene expressionby somatosialin. J. Clin. Invest., 83: 367-372, 1989.Varner. A. A.. Modlin. I. M.. and Walsh, J. H. High potency of bombesin forstimulation of human gastrin release and gastric acid secretion. Regul. Pepi. /:289-296. 1981.Ciardiello. F.. Kim. N.. Saeki. T.. Dono, R.. Persico, M. G., Plowman. G. D.. Garrigues. J.. Radke. S.. Todaro. G. J., and Salomon. D. S. Differential expression ofepidermal growth factor-related proteins in human colorectal tumors. Proc. Nati.Acad. Sci. USA. 88: 7792-7796. 1991.Owen. D. Normal histology of the stomach. Am. J. Surg. Pathol.. 10: 48-61. 1986.Johnson. L. R.. and Gulhrie. P. D. Proglumide inhibilion of Ihe trophic action ofpentagaslrin. Am. J. Physiol.. 246: G62-G66. 1984.Beauchamp. R. D.. Townsend. C. M.. Singh. P.. Glass. E. J.. and Thompson, J. C.Proglumide. a gasirin receptor antagonist, inhibits growth of colon cancer and enhances survival in mice. Ann. Surg., 202: 303-309, 1985.Hoosein. N. M.. Kiener. P. A.. Curry. R. C.. Rovali. L. C.. McGilbra. D. K.. andBratlain. M. G. Anliproliferalive effects of gastrin receptor antagonists and antibodiesto gastrin on human colon cancer cell lines. Cancer Res., 48: 7179-7183. 1988.Kopin, A. S.. Lee. Y. M.. McBride. E. W.. Miller. L. J.. Lu. M., Lin, H-Y.. Kolakowski.L. F.. and Beinborn, M. Expression cloning and characterizaiion of the canine parietalcell gastrin receptor. Proe. Nati. Acad. Sei. USA. 89: 3605-3609. 1992.Pisegna. J. R.. de Weerth, A.. Huppi. K.. and Wank, S. A. Molecular cloning of thehuman brain and gastric Cholecyslokinin receptor: structure, functional expression andchromosomal localization. Bioehem. Biophys. Res. Commun.. I8V: 296-303. 1992.Browder. T. M.. Dunbar. C. E., and Nienhuis. A. W. Privale and public auUKrine loopsin neoplastic cells. Cancer Cells. /: 9-17. 1989.Baldwin. G. S. Gastrin and the transforming protein of polyoma virus have evolvedfrom a common ancestor. FEBS Leu.. 137: 1-5. 1982.Caniley. L. C.. Auger. K. R.. Carpenter. C., et al. Oncogenes and signal transduction.Cell. 64: 281-302. 1991.Cartwrighi. C. A., Kamps. M. P.. Meisler. A. !.. Pipas. J.. and Eckharl. W. pp60l"-"
activation in human colon carcinoma. J. Clin. Invest., 83: 2025-2033. 1989.Baldwin. G. S.. Knesel. J.. and Monckton. J. M. Phosphorylation of gaslrin-17 byepidermal growth faclor-slimulated tyrosinc kinase. Nature (Lond.l. 301: 435—137.1983.Dy, D. Y. Whitehead. R. H.. and Morris. D. L. SMS20I.995 inhibits in \tini and inVMYÕgrowth of human colon cancer. Cancer Res.. 52: 917-923. 1992.Lee, M. T., Liebow. C.. Kamer, A. R.. and Schally, A. V. Effects of epidermal growthfactor and analogues of luteinizing hormone-releasing hormone and somatostatin onphosphorylalion and dephosphorylalion of lyrosine residues of specific protein sub-slrales in various tumors. Proc. Nati. Acad. Sci. USA. XX: 1656-1660. 1991.