Page 1
www.elsevier.com/locate/vetimm
Veterinary Immunology and Immunopathology 104 (2005) 117–127
Expression patterns of chicken Toll-like receptor mRNA
in tissues, immune cell subsets and cell lines
Muhammad Iqbal, Victoria J. Philbin, Adrian L. Smith*
Enteric Immunology Group, G2D Stewart Building, Division of Immunology and Pathology, Institute for Animal Health,
Compton Laboratory, Compton, Newbury, Berkshire, RG20 7NN, UK
Received 3 May 2004; received in revised form 13 October 2004; accepted 8 November 2004
Abstract
The Toll-like receptor (TLR) family of cell surface molecules represent a major component of the pattern recognition
system, which enables both vertebrates and invertebrates to detect invading microorganisms and mount an anti-microbial
response. The TLR repertoire of mouse and man has been intensively studied and in this manuscript we report the identification
of ESTs with homology to chTLR5 and chTLR7, and independently confirm the identification of chTLR 1/6/10 and 3 in the EST
databases. We have determined the mRNA expression patterns for seven chicken TLRs (chTLR) in a wide range of chicken
tissues, isolated immune cell types and cultured cells. Some of the chTLR were expressed in most tissues (chTLR1/6/10,
chTLR3, chTLR4 and chTLR5), whereas others exhibited more restricted expression patterns (chTLR2 type 1, type 2 and
chTLR7). Similarly distinct patterns of chTLR expression were seen with innate and adaptive immune cell types isolated from
peripheral blood or spleen and with cultured cells of somatic or immunological origin. An understanding of the TLR repertoire
for different tissues, immune cell subsets and cultured cell types allows more refined interpretation of immune induction in
response to chicken pathogens.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Toll-like receptors; Innate immunity; Chickens
1. Introduction
Toll-like receptors (TLRs) are well recognized as
major components of the pattern recognition receptor
Abbreviations: chTLR, chicken Toll-like receptor; EST,
expressed sequence tag
* Corresponding author. Tel.: +44 1635 577293;
fax: +44 1635 577263.
E-mail address: [email protected] (A.L. Smith).
0165-2427/$ – see front matter # 2004 Elsevier B.V. All rights reserved
doi:10.1016/j.vetimm.2004.11.003
system (Medzhitov and Janeway, 2000; Akira, 2004;
Beutler, 2004) that detects invading pathogens by
virtue of a series of conserved molecular structures
known as pathogen associated molecular patterns
(PAMPs). The importance of TLR function is evident
by their role in the immune system of organisms as
diverse as Drosophila and man. Although studies of
TLR immune function are most advanced in mouse
and man, a series of TLRs have been identified in other
vertebrates including fish (Oshiumi et al., 2003;
.
Page 2
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127118
Hirono et al., 2004; Jault et al., 2004; Meijer et al.,
2004), chickens (Fukui et al., 2001; Leveque et al.,
2003) and cattle (Werling and Jungi, 2003).
Toll-like receptors are type 1 transmembrane
proteins with N-terminal extracytoplasmic leucine-
rich repeat (LRR) domains and an intracytoplasmic
conserved region called the TIR (Toll/IL1-R) domain,
with homology to the mammalian interleukin 1
receptor (IL1-R) and plant disease resistance genes
(Rock et al., 1998). TLRs induce signals through TIR
domains that interact with different adapter proteins
such as MyD88, resulting in the activation of nuclear
factor kB (NF-kB) and the mitogen-activated protein
kinase signaling cascade (Barton and Medzhitov,
2003).
The TLRs have distinct specificity, with particular
PAMPs acting as agonists for different TLRs. These
agonists (and their associated TLR) include lipopo-
lysaccharide (LPS) from gram negative bacteria
(TLR4), lipoprotein and peptidoglycan from gram
positive bacteria (TLR1, 2 and 6), flagellin (TLR5),
double stranded RNA (TLR3), unmethylated CpG
dinucleotide motifs (TLR9), single stranded uridine
rich RNA (TLR7) and the synthetic antiviral
compound R-848 (TLR7 and TLR8). Upon activation
with an appropriate ligand, TLRs induce a range of
responses including cell proliferation or maturation
and the production of various cytokines, chemokines
or effector molecules, including nitric oxide and
reactive oxygen intermediates (e.g., Thoma-Uszynski
et al., 2000; Hemmi et al., 2002; Smith et al., 2003).
Signalling through TLRs leads to widespread immune
induction of the cellular components of the innate and
adaptive immune systems as well as directing the host
response into particular differentiation pathways.
In the chicken, TLR2 and TLR4 have been
identified and characterised at molecular and func-
tional levels (Boyd et al., 2001; Fukui et al., 2001;
Leveque et al., 2003). Interestingly, Fukui et al. (2001)
identified two types of chicken TLR2 and mapped
them both to the same chromosomal location on
chicken chromosome 4, suggesting that they arose by
gene duplication (Fukui et al., 2001). Chicken ESTs
have been identified with highest homology to
mammalian TLR 1, 6 or 10 and TLR3 (Lynn et al.,
2003). We have independently identified these ESTs
and also identified ESTs with high homology to TLR5
and TLR7 (this paper). Moreover, chicken ESTs have
been identified with homology to many components of
the known TLR signaling pathways (Lynn et al., 2003)
suggesting there are considerable similarities between
the mammalian and chicken TLR systems.
The tissue and cell distribution is an important
characteristic of TLR function since it influences the
capacity to detect different microorganisms during
their entry via, and growth in, different tissues. The
distribution of mammalian TLRs has revealed
characteristic patterns of expression for each TLR
(e.g., Sebastiani et al., 2000; Rehli, 2002; Zarember
and Godowski, 2002; Zhang et al., 2004) and in this
manuscript we have determined the distribution of
chTLR mRNA in tissues, immune cell subsets and in a
variety of chicken cell lines. A number of studies have
defined the response of cultured chicken cells after
exposure to a variety of challenges with microorgan-
isms or their products (e.g., Zhang et al., 1995; Kaiser
et al., 2000; Dil and Qureshi, 2002a; Farnell et al.,
2003; Kogut et al., 2003). We used RT-PCR to
determine the TLR repertoire of chicken tissues, ex
vivo sorted cell populations and commonly used
cultured cell types. Understanding the distribution
patterns of chTLR will enable more defined inter-
pretation of immune induction and the host-pathogen
relationships that define infectious disease biology in
the chicken.
2. Materials and methods
2.1. Experimental animals
Specific-pathogen-free inbred Line 72 (White
Leghorn) and Rhode Island Red chickens were
maintained by the Poultry Production Unit of the
Institute for Animal Health (IAH), Compton Labora-
tory. The birds did not receive any vaccination and
were reared in wire cages and given ad libitum access
to water and a vegetable-based protein diet (Special
Diet Services, Witham, UK).
2.2. Database mining
Chicken EST databases were screened with DNA
sequences from human TLR1-10 deposited in EMBL
(http://www.ebi.ac.uk; see Table 1 for the relevant
accession numbers) and by key word searches, in the
Page 3
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127 119
Table 1
Sequence of primers used in RT-PCR
Primer Sequence Accession number
TLR1/6/10 F CGGAAAGCCTATCATTGTCA BQ484541/BU471924
TLR1/6/10 R TTTGTCTGCGTCCACTGC BQ484541/BU471924
TLR2 type1 F TTAAAAGGGTGTGCCAGGAG AB050005
TLR2 type1 R GTCCAAACCCATGAAAGAGC AB050005
TLR2 type2 F AGGCACTTGAGATGGAGCAC AB046533
TLR2 type2 R CCTGTTATGGGCCAGGTTTA AB046533
TLR3 F CCACTCTGGAAGAAAATGAGC BI066273
TLR3 R TCATTCTCACCGCTTTTCAG BI066273
TLR4 F AGTCTGAAATTGCTGAGCTCAAAT AY064697
TLR4 R GCGACGTTAAGCCATGGAAG AY064697
TLR5 F TGCACATGTTTTCTCCTAGGT AJ626848
TLR5 R CCACATCTGACTTCTGCCTTT AJ626848
TLR7 F GCCTCAAGGAAGTCCCCAGA AJ632302
TLR7 R AAGAAACATTGCATGGATTACGG AJ632302
b-Actin F TGCTGTGTTCCCATCTATCG L08165
b-Actin R TTGGTGACAATACCGTGTTCA L08165
period between January 2001 and April 2002. The
databases contained sequences originating from bursal
cell cDNA library (Buerstedde et al., 2002) or from a
range of control and infected chicken tissues
(Tirunagaru et al., 2000). All sequences corresponding
to potential chicken TLRs were re-screened against
mammalian sequence databases to identify or confirm
the mammalian TLR with the highest degree of
sequence similarity.
2.3. Tissue collection, cell isolation and cell cultures
All tissue samples were obtained from inbred 8-
week-old Line 72 (White Leghorn) chickens main-
tained under SPF conditions at the IAH, Compton.
Chicken tissues were immediately placed in RNA later
(Qiagen) and stored temporarily in the fridge before
longer term storage at �20 8C according to manu-
facturer’s instructions.
Splenic B cells, CD4+, CD8+, TCR1, TCR2 and
TCR3 were enriched by sorting with magnetic beads
(Miltenyi) using the AutoMACSTM system, according
to manufacturer’s instructions. Briefly, spleens were
disrupted by passage through a coarse sieve and
lymphocytes isolated using Ficoll-PaqueTM Plus
(Amersham Biosciences). The spleen cell suspension
was incubated with primary antibodies (all obtained
from Southern Biotechnology) against B cells or T cell
subsets as follows: anti-Bu-1 PE, anti-CD4 FITC, anti-
CD8a PE, anti-TCR1 (IgG1; TCRgd) or anti-TCR3
(IgG1; TCRab2) at 200 ml/108 cells or TCR2 PE
(TCRab1) at 500 ml/108 PBS- 0.5% BSA diluent for
15 min on ice. Following washing by centrifugation
and resuspension of the cell pellet in PBS-0.5% BSA,
different cell populations were isolated by positive
selection using anti-fluorochrome or rat anti-mouse
IgG1 conjugated paramagnetic beads (Miltenyi
Biotech) after 15 min incubation on ice using the
AutoMACSTM system (Miltenyi Biotech). The sorted
cell populations were enriched to 90% or greater
purity, except for the CD4+ cell population, which was
71%, as measured by FACS analysis. Peripheral blood
monocyte-derived macrophages were obtained from
Line 72 chickens according to the methodology of
(Wigley et al., 2002). Heterophils were isolated from
peripheral blood as described by (Kogut et al., 1995)
and were in excess of 90% purity by microscopic
analysis of hematoxylin/eosin stained cytospin pre-
parations.
Primary cell cultures of chicken kidney (2-week-old
chickens) and embryonic fibroblasts (10 day embryos)
were prepared by the IAH cell culture service
according to standard techniques. All cells were
cultured in RPMI 1640 medium supplemented with
penicillin/streptomycin, 7% foetal calf serum and 3%
chicken serum (complete medium) at 41 8C, 5% CO2.
The macrophage-like HD11 cell line (Beug et al., 1979)
and the bursal B cell-like lymphoma DT40 (ATCC:-
CRL2111) were grown in 80 cm2 tissue culture flasks
with complete medium at 41 8C, 5% CO2.
Page 4
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127120
Isolated cell subsets or cultured cells were
disrupted directly in RLT buffer (Qiagen) and frozen
at �20 8C until RNA extraction.
2.4. RNA extraction
Chicken tissues were disrupted by homogenisation
using a mini-bead beater (Biospec products). RNA
was extracted, from tissue homogenates or disrupted
cells, in the presence of buffer containing b-
merceptoethanol and guanidine by using RNeasy
Mini Kit (Qiagen) following manufacturer’s instruc-
tions. To avoid contamination with DNA the samples
were exposed to an on-column treatment with RNase-
Free DNase 1 (Qiagen) for 90 min at room tempera-
ture (20–25 8C). The column was washed with RW1
and RWP buffers and the RNA eluted with RNase free
water.
2.5. RT-PCR
The extracted RNA was reverse transcribed using
the Promega Reverse Transcription kit (Promega)
according to manufacturer’s instructions. Briefly,
oligo dT primer (0.5 mg) was used to reverse
transcribe 1 mg of respective RNA in the presence
of dNTP’s (250 mM), reverse transcriptase buffer
(10 mM Tris–HCl, 50 mM KCl, 0.1% TritonR-X-100),
AMV Reverse Transcriptase High Conc. (15 units/mg)
and RNasin Ribonuclease inhibitor (1 unit/ml) at
42 8C for 60 min following inactivation at 99 8C for
5 min.
All RNA preparations were standardised by RT-
PCR for b-Actin (see Table 1 for primer sequences)
and were free from DNA contamination as judged by a
lack of signal from non-reverse transcribed RNA with
all primer sets (data not shown).
Polymerase chain reactions (PCR) were performed
according to standard protocols with primers indicated
in Table 1. Briefly, cDNA (3 ml) was reacted with
200 mM dNTPs, 1 � reaction buffer (Promega),
forward and reverse primers (5 pM) and 0.5 units
Taq polymerase (Promega) in 50 ml final reaction
volume. PCR conditions were as follows, 1 cycle,
95 8C for 2 min followed by 30 cycles of 30 s at 95 8C,
58 8C for 1 min and 72 8C for 2 min followed by 1
cycle at 72 8C for 10 min using a iCycler (BIO RAD).
The amplified products were analysed by agarose gel
(2%) electrophoresis in 0.5� TBE buffer at 50 mA for
1 h and products visualised by staining with ethidium
bromide.
3. Results and discussion
The TLRs represent an evolutionary highly
conserved group of pattern recognition receptors,
involved in the induction of anti-microbial responses
in species as diverse as flies, fish, mouse and man.
Although the TLRs are germ-line encoded and can be
considered receptors of the innate immune system,
TLR-mediated signals are central players in the
induction and differentiation of adaptive immune
responses.
To date, homologues of TLR2 and TLR4 have been
characterised in the best-studied avian genome, the
chicken, Gallus gallus domesticus. Interestingly,
chTLR2 genes have been described which co-localise
to a region on chicken chromosome 4, suggesting that
they arose by gene duplication (Fukui et al., 2001).
Although type 1 chTLR2 and type 2 chTLR2 share
88.5% nucleic acid homology with each other, only
chTLR2 type 2 mediates a strong response to classical
mammalian TLR2 agonists (Fukui et al., 2001). There
is a single TLR2 gene in mammals although two other
sets of mammalian TLRs seem to have arisen by local
gene duplication. TLR1, 6 and 10 lie adjacent to each
other on human chromosome 4 (Takeuchi et al., 1999)
and TLR7 and TLR8 lie adjacent to each other on the
X chromosome (Du et al., 2000). Evidence for similar
duplication events can be found with zebrafish TLR4
and TLR8 genes (Jault et al., 2004; Meijer et al.,
2004). Chicken TLR4 has also been cloned and
mapped to a region associated with susceptibility to
Salmonella infection in some inbred chicken lines
(Leveque et al., 2003). Chicken TLR2 and TLR4 share
52 and 46% overall identity with human TLR2 and
TLR4, respectively at the protein level, with the
greatest degree of homology being seen in the TIR
domain. The homology and mapping data indicate that
TLR2 and TLR4 were distinct entities before the
divergence of birds and mammals over 300 million
years ago.
Analysis of various databases revealed the exis-
tence of chicken ESTs with highest sequence
homology to human TLR1, TLR2, TLR3, TLR4,
Page 5
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127 121
TLR5, TLR6 and TLR7 (Table 1). Lynn et al. (2003)
reported the identification of ESTs with sequence
homology to human TLR1, 6 or 10 and TLR3, our
independent analysis confirmed these findings and
identified further ESTs with sequence similarity to
TLR5 and TLR7 (Table 1). The genomic localisation
for chTLR3 (chromosome 4), chTLR5 (chromosome
3) and chTLR7 (chromosome 1) has been identified
experimentally and these TLRs have been subjected to
detailed functional analysis (manuscript in prepara-
tion). Two non-overlapping EST with similarity to
TLR1, 6 or 10 were identified but RT-PCR revealed
that these two EST represent a single RNA species,
which was confirmed by sequence of the RT-PCR
product (data not shown). Hence, we have only
performed RT-PCR with primers designed to
amplify the longer product and the extra sequence
that we obtained also failed to discriminate between
TLR1, 6 or 10 as the orthologous sequence. Indeed,
molecular evolutionary analysis of TLR1, 6 and 10
TIR sequences suggest that the ancestral TLR first
duplicated in mammals at around the time of
divergence from birds (�300 million years ago)
followed by a later duplication at �130 million
years ago (Beutler and Rehli, 2002). Detailed analysis
of the genomic region containing chTLR7 has
revealed that chTLR8 is disrupted (manuscript in
preparation). We were unable to identify any EST
with sequence homology to human TLR9 or mouse
TLR11.
In order to define the cell and tissue distribution for
the chTLR mRNAs we designed primers using the
available EST sequences, the coding sequence for
chTLR5 and chTLR7 that we have obtained (manu-
scripts in preparation) or previously published coding
sequences to generate chTLR-specific RT-PCR pro-
ducts. Tissue-specific expression of TLRs has been
assessed in man, mouse and fish (e.g., Sebastiani et al.,
2000; Zarember and Godowski, 2002; Jault et al.,
2004; Meijer et al., 2004; Zhang et al., 2004) revealing
distinct distribution patterns for each TLR. Tissue-
specific expression of the chTLR mRNA was assessed
in 18 different tissues obtained from 8-week-old Line
72 chickens. The tissues included those with primary
immunological function (thymus, bursa of Fabricius,
bone marrow and spleen), dominated by epithelial
cells (kidney, liver), neurological (brain), muscle
(skeletal and heart) as well as tissues that interface
with the internal (various regions of the small and
large intestine) and external (skin) milieu.
To allow comparison of the expression of chTLRs
between different tissues the chicken tissue cDNA
pools were normalised by RT-PCR to the house
keeping gene b-actin. All RNA samples used for
cDNA synthesis were demonstrated to be free from
genomic DNA contamination by the absence of
product on non-reverse transcribed samples (data
not shown). Although the analysis is not strictly
quantitative, all RT-PCRs were performed in parallel
with a non-saturating number of cycles therefore,
comparison of the levels of signal give a guide of
relative tissue expression of the respective TLR
mRNA.
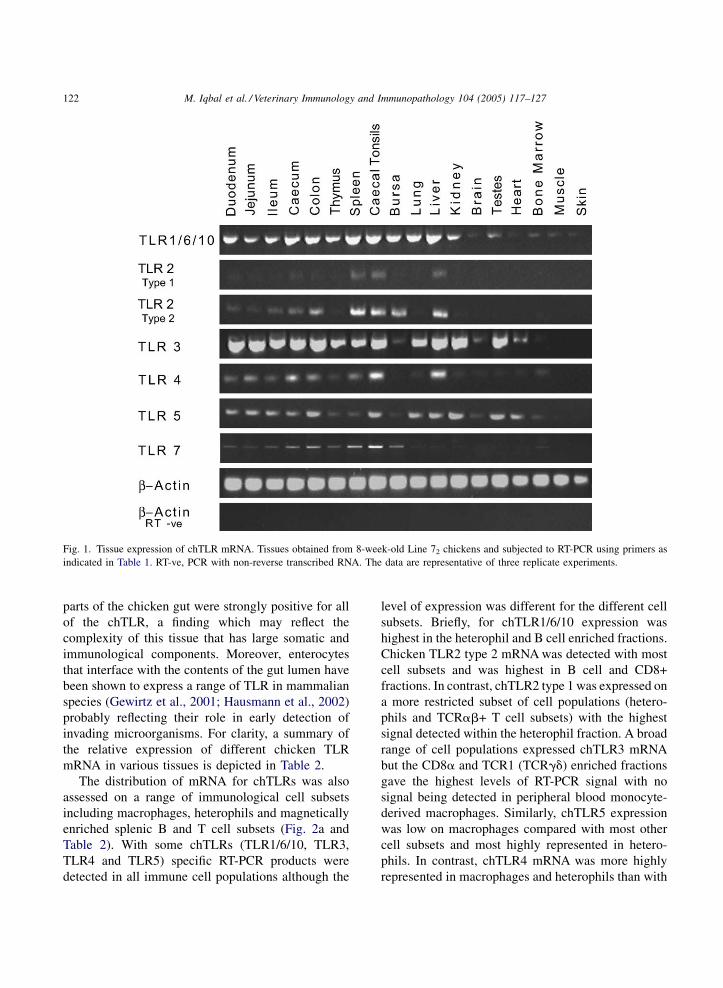
A broad pattern of tissue expression was obtained
for chTLR1/6/10, chTLR3, chTLR4 and chTLR5
mRNA in tissues that comprise either a large or
minimal immunological compartment (Fig. 1). For
example, chTLR1/6/10 mRNA was expressed at
reasonably high levels in the spleen and in the kidney,
the latter a tissue without an organised immune
compartment. The breadth of tissue expression for
chTLR1/6/10 is in contrast with humans in where the
three similar TLRs are mostly expressed in the spleen
and peripheral blood lymphocytes (Zarember and
Godowski, 2002). Similarly, the expression patterns of
chTLR3, 4 and 5 share some aspects of tissue
expression patterns with humans but there are also
significant differences in the levels of expression in
different tissues (for example, compare our data with
that of Rock et al., 1998; Zarember and Godowski,
2002). With chTLR4 the level of mRNA appears to be
highest in tissues that have relatively high numbers of
macrophages or macrophage-like cells, including the
liver which is populated by large numbers of Kupffer
cells.
The expression of chTLR2 type 1, chTLR2 type 2
and TLR7 was more restricted than that seen with the
other chTLRs and highest levels of expression were
found in tissues with large immunological compart-
ments. Chicken TLR2 type 1 had a much more
restricted tissue expression than chTLR2 type 2, with
the former only being expressed at moderate levels in
the spleen, caecal tonsil and liver samples. These data
are in agreement with that of Fukui et al. (2001)
although we have extended analysis to a broader range
of tissues. A range of tissues derived from different
Page 6
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127122
Fig. 1. Tissue expression of chTLR mRNA. Tissues obtained from 8-week-old Line 72 chickens and subjected to RT-PCR using primers as
indicated in Table 1. RT-ve, PCR with non-reverse transcribed RNA. The data are representative of three replicate experiments.
parts of the chicken gut were strongly positive for all
of the chTLR, a finding which may reflect the
complexity of this tissue that has large somatic and
immunological components. Moreover, enterocytes
that interface with the contents of the gut lumen have
been shown to express a range of TLR in mammalian
species (Gewirtz et al., 2001; Hausmann et al., 2002)
probably reflecting their role in early detection of
invading microorganisms. For clarity, a summary of
the relative expression of different chicken TLR
mRNA in various tissues is depicted in Table 2.
The distribution of mRNA for chTLRs was also
assessed on a range of immunological cell subsets
including macrophages, heterophils and magnetically
enriched splenic B and T cell subsets (Fig. 2a and
Table 2). With some chTLRs (TLR1/6/10, TLR3,
TLR4 and TLR5) specific RT-PCR products were
detected in all immune cell populations although the
level of expression was different for the different cell
subsets. Briefly, for chTLR1/6/10 expression was
highest in the heterophil and B cell enriched fractions.
Chicken TLR2 type 2 mRNA was detected with most
cell subsets and was highest in B cell and CD8+
fractions. In contrast, chTLR2 type 1 was expressed on
a more restricted subset of cell populations (hetero-
phils and TCRab+ T cell subsets) with the highest
signal detected within the heterophil fraction. A broad
range of cell populations expressed chTLR3 mRNA
but the CD8a and TCR1 (TCRgd) enriched fractions
gave the highest levels of RT-PCR signal with no
signal being detected in peripheral blood monocyte-
derived macrophages. Similarly, chTLR5 expression
was low on macrophages compared with most other
cell subsets and most highly represented in hetero-
phils. In contrast, chTLR4 mRNA was more highly
represented in macrophages and heterophils than with
Page 7
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127 123
Table 2
Summary of chTLR mRNA expression
TLR1/6/10 TLR2 Type1 TLR2 Type2 TLR3 TLR4 TLR5 TLR7
Tissues
Duodenum +++ + + ++++ ++ ++ +
Jejunum +++ + + ++++ ++ ++ +
Ileum +++ + + ++++ ++ ++ +
Caecum +++ + + ++++ ++ ++ +
Colon +++ + + ++++ ++ ++ +
Thymus +++ + + +++ ++ ++ +
Spleen +++ ++ ++ +++ ++ ++ +
Ceacal tonsil +++ ++ ++ ++++ +++ ++ ++
Bursa +++ + ++ + +/� + +
Lung +++ +/� +/� +++ + ++ +
Liver +++ ++ ++ ++++ ++ ++ +
Kidney + � + ++++ + ++ +
Brain + � + ++ + + �Testes + � + +++ + ++ �Heart + � + ++ + ++ +/�Bone marrow + � + + + + +
Muscle + � +/� +/� � + �Skin + � � � � + �
Cultured cells
CKC � + � + + ++ �CEF + + � ++ + +++ +/�DT-40 ++ ++ +++ + + +/� ++
HD11 +++ ++ + + +++ + ++++
Immune cell subsets
Macrophages + � + � ++++ + +
Heterophils +/� ++ + + ++++ +++ +/�B cells ++ +/� +++ + ++ ++ ++++
CD4+ ++ � + + + + +++
CD8+ ++ + ++ +++ ++ ++ ++
TCR1 (gd) + � + ++ + + +
TCR2 (ab1) + + + + + + +
TCR3 (ab2) + + + + + + +
Relative level of RT-PCR product denoted on a � (undetectable signal) to ++++ (strong signal) scale. Total RNAwas isolated from tissues, blood
monocyte derived macrophages, blood heterophils, magnetically sorted spleen cell subpopulations and cultured cells. These data summarise RT-
PCR presented in Figs. 1 and 2. CKC, chicken kidney cells; CEF, chicken embryo fibroblasts and has been constructed from three independent
experiments for each sample set.
other immune cell populations. As is the case with
mammalian TLR7, chTLR7 was expressed predomi-
nantly within the B cell fraction but was also detected
on various T cell subsets. We also detected a faint
signal within the polymorphonuclear cell fraction
(heterophils), which is similar to that reported with
murine eosinophils and neutrophils (Nagase et al.,
2003).
When chTLR expression is assessed in various cell
fractions it is clear that some cell subsets expressed a
greater repertoire of chTLR than other cell types. Most
notably, peripheral blood monocyte-derived macro-
phages expressed considerable levels of chTLR4 and
chTLR1/6/10, moderate levels of chTLR2 type 2, low
but detectable levels of chTLR5 and chTLR7 and no
chTLR3 or chTLR2 type 1 message. Heterophils
expressed message for all of the known chTLRs with
chTLR1/6/10, chTLR2, chTLR4 and chTLR5 most
prominent, moderate expression for chTLR3 and low
but detectable levels for chTLR7. These patterns of
chTLR mRNA expression in immune cell subsets are
broadly similar to those detected in mammalian
species (Applequist et al., 2002; Hornung et al., 2002;
Zarember and Godowski, 2002) with the broadest
Page 8
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127124
Fig. 2. Expression of chTLR mRNA in immune cell populations and cultured cells. (a) Total RNA was isolated from blood monocyte derived
macrophages, blood heterophils and magnetically sorted spleen cell subpopulations. (b) Total RNA was isolated from cultured cell types. CKC,
chicken kidney cells; CEF, chicken embryo fibroblasts; DT-40 (B cell-like cell line); HD11 (chicken macrophage-like cell line). RT-ve, PCR with
non-reverse transcribed RNA. The data are representative of three replicate experiments.
range of TLR expression on cell subsets representing
the innate immune system.
Two widely used primary cell culture systems were
selected for analysis, representing somatic cells of
predominantly epithelial (chicken kidney; CKC) or
fibroblast (chicken embryo fibroblasts; CEF) origin.
Two established cell lines, both of immunological cell
derivation (DT40 and HD11), were also assessed for
their chTLR repertoire. The DT40 cell line
(ATCC:CRL2111) is derived from an avian leukosis
virus-transformed pre-B-cell, whereas the HD11 cell
line (Beug et al., 1979) is a retrovirally transformed
macrophage-like cell line. These four cell culture
systems have proven valuable tools for studies of the
interaction of cells with pathogens or pathogen-
derived products (e.g., Zhang et al., 1995; Kaiser et al.,
2000; Farnell et al., 2003; Kogut et al., 2003) and
determination of the TLR repertoires would allow
more refined interpretation of such studies in the
future. For example, Kaiser et al. (2000) documented
the differential production of pro-inflammatory
cytokines after exposure of CKC to the Salmonella
enterica serovars Typhimurium, Enteritidis and
Gallinarum. One obvious difference in TLR agonists
is the flagella status of these organisms, S. Gallinarum
is an aflagellate serovar, which obviously has
implications for TLR5 signalling since CKC are
TLR5 positive (see below).
Although most of the chTLRs (chTLR1/6/10,
chTLR2 type 1, chTLR3, 4 and 5) could be detected
in all four cultured cell types chTLR2 type 2 was only
detected in the cultured cell lines of immunological
origin (DT-40 and HD11). Chicken TLR7 mRNA was
principally detected in the immune-derived cell lines
although a very faint RT-PCR product was detected in
the CEF cell cultures. Nonetheless, the relative
Page 9
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127 125
intensity of the chTLR PCR products differed
considerably between the cultured cell types. Briefly,
of the broadly expressed chTLRs, chTLR3 and
chTLR5 were expressed at higher levels in somatic
cell cultures (CKC and CEF), whereas chTLR1/6/10,
chTLR2 type 1 and chTLR4 were expressed in at
higher levels in one, or both (chTLR1/6/10), of the
immune derived cell lines. The RT-PCR product for
chTLR2 type 2 was most intense in the DT40 cells (B
cell-like) and the chTLR4 product was most intense in
the macrophage-like cell line (HD11). A summary of
the expression patterns is depicted in Table 2.
The high expression of chTLR7 mRNA in the
HD11 cell line may be considered surprising (ex vivo,
TLR7 expression is highest in B cells and dendritic
cell subsets) but many murine macrophage-like cell
lines including J774 cells express TLR7 message at
high levels (Applequist et al., 2002). Moreover, we
have determined that the chTLR7 found in HD11 cells
is functional and exposure to imadazoquinoline
agonists stimulates upregulation of cytokine mRNA
(manuscript in preparation). The high levels of chTLR
expression in the primary somatic cell dominated
cultures (CKC and CEF) indicate the importance of
epithelial and fibroblastic cell types in the sensing of
microbial invasion. The role of some epithelial cell
subsets in induction of immunity is becoming more
apparent, particularly with reference to the gut
enterocytes (e.g., Cario and Podolsky, 2000; Gewirtz
et al., 2001).
In this report, we have documented the expression
of chTLR mRNA in a broad range of tissues, isolated
immune cell subsets and cultured cell populations.
While many of the patterns of chTLR expression are
broadly comparable to that found for mammalian
TLRs there do appear to be some significant
differences that are specific to the chicken. It is of
note that there are also substantial differences in the
patterns of TLR expression between mouse and man
(Rehli, 2002) indicating the importance of determin-
ing the distribution of TLR in the target species. More
detailed analysis of the regulation of expression of
TLR in a non-mammalian system, especially one
with well documented patterns of pathogen challenge
(e.g. the chicken), may offer opportunities to
define evolutionarily important components in the
regulation of TLR expression. We recognise that the
expression profiles documented in this manuscript
only represent a starting point in the analysis of chTLR
since there may be differences associated with host
genetics (e.g., with TLR4 polymorphism (Leveque
et al., 2003); or expression levels, (Dil and Qureshi,
2002a,b) or environmental factors (e.g., the presence
of challenge infections). Nevertheless, the work
presented represents an important contribution to
understanding the induction of avian immune
responses and disease processes and may ultimately
lead to the formulation of rational immunological
intervention strategies.
Acknowledgements
The authors wish to acknowledge the input of Dr.
Yvonne Boyd for discussions and comment on the
manuscript. The work was supported by the BBSRC
Grant numbers 201/S15839 and 02/A1/S/08451, and
the IAH.
References
Akira, S., 2004. Toll receptor families: structure and function.
Semin. Immunol. 16, 1–2.
Applequist, S.E., Wallin, R.P., Ljunggren, H.G., 2002. Variable
expression of Toll-like receptor in murine innate and adaptive
immune cell lines. Int. Immunol. 14, 1065–1074.
Barton, G.M., Medzhitov, R., 2003. Toll-like receptor signaling
pathways. Science 300, 1524–1525.
Beug, H., von Kirchbach, A., Doderlein, G., Conscience, J.F., Graf,
T., 1979. Chicken hematopoietic cells transformed by seven
strains of defective avian leukemia viruses display three distinct
phenotypes of differentiation. Cell 18, 375–390.
Beutler, B., 2004. Innate immunity: an overview. Mol. Immunol. 40,
845–859.
Beutler, B., Rehli, M., 2002. Evolution of the TIR, tolls and TLRs:
functional inferences from computational biology. Curr. Top.
Microbiol. Immunol. 270, 1–21.
Boyd, Y., Goodchild, M., Morroll, S., Bumstead, N., 2001. Mapping
of the chicken and mouse genes for toll-like receptor 2 (TLR2) to
an evolutionarily conserved chromosomal segment. Immunoge-
netics 52, 294–298.
Buerstedde, J.M., Arakawa, H., Watahiki, A., Carninci, P.P., Haya-
shizaki, Y.Y., Korn, B., Plachy, J., 2002. The DT40 web site:
sampling and connecting the genes of a B cell line. Nucleic
Acids Res. 30, 230–231.
Cario, E., Podolsky, D.K., 2000. Differential alteration in intestinal
epithelial cell expression of toll-like receptor 3 (TLR3) and
TLR4 in inflammatory bowel disease. Infect. Immun. 68, 7010–
7017.
Page 10
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127126
Dil, N., Qureshi, M.A., 2002a. Differential expression of inducible
nitric oxide synthase is associated with differential Toll-like
receptor-4 expression in chicken macrophages from different
genetic backgrounds. Vet. Immunol. Immunopathol. 84, 191–
207.
Dil, N., Qureshi, M.A., 2002b. Involvement of lipopolysaccharide
related receptors and nuclear factor kappa B in differential
expression of inducible nitric oxide synthase in chicken macro-
phages from different genetic backgrounds. Vet. Immunol.
Immunopathol. 88, 149–161.
Du, X., Poltorak, A., Wei, Y., Beutler, B., 2000. Three novel
mammalian toll-like receptors: gene structure, expression, and
evolution. Eur. Cytokine Netw. 11, 362–371.
Farnell, M.B., Crippen, T.L., He, H., Swaggerty, C.L., Kogut, M.H.,
2003. Oxidative burst mediated by toll like receptors (TLR) and
CD14 on avian heterophils stimulated with bacterial toll ago-
nists. Dev. Comp. Immunol. 27, 423–429.
Fukui, A., Inoue, N., Matsumoto, M., Nomura, M., Yamada, K.,
Matsuda, Y., Toyoshima, K., Seya, T., 2001. Molecular cloning
and functional characterization of chicken toll-like receptors. A
single chicken toll covers multiple molecular patterns. J. Biol.
Chem. 276, 47143–47149.
Gewirtz, A.T., Navas, T.A., Lyons, S., Godowski, P.J., Madara, J.L.,
2001. Cutting edge: bacterial flagellin activates basolaterally
expressed TLR5 to induce epithelial proinflammatory gene
expression. J. Immunol. 167, 1882–1885.
Hausmann, M., Kiessling, S., Mestermann, S., Webb, G., Spottl, T.,
Andus, T., Scholmerich, J., Herfarth, H., Ray, K., Falk, W.,
Rogler, G., 2002. Toll-like receptors 2 and 4 are up-regulated
during intestinal inflammation. Gastroenterology 122, 1987–
2000.
Hemmi, H., Kaisho, T., Takeuchi, O., Sato, S., Sanjo, H., Hoshino,
K., Horiuchi, T., Tomizawa, H., Takeda, K., Akira, S., 2002.
Small anti-viral compounds activate immune cells via the TLR7
MyD88-dependent signaling pathway. Nat. Immunol. 3, 196–
200.
Hirono, I., Takami, M., Miyata, M., Miyazaki, T., Han, H.J., Takano,
T., Endo, M., Aoki, T., 2004. Characterization of gene structure
and expression of two toll-like receptors from Japanese flounder.
Paralichthys olivaceus. Immunogenetics 56, 38–46.
Hornung, V., Rothenfusser, S., Britsch, S., Krug, A., Jahrsdorfer, B.,
Giese, T., Endres, S., Hartmann, G., 2002. Quantitative expres-
sion of toll-like receptor 1-10 mRNA in cellular subsets of
human peripheral blood mononuclear cells and sensitivity to
CpG oligodeoxynucleotides. J. Immunol. 168, 4531–4537.
Jault, C., Pichon, L., Chluba, J., 2004. Toll-like receptor gene family
and TIR-domain adapters in Danio rerio. Mol. Immunol. 40,
759–771.
Kaiser, P., Rothwell, L., Galyov, E.E., Barrow, P.A., Burnside, J.,
Wigley, P., 2000. Differential cytokine expression in avian cells
in response to invasion by Salmonella Typhimurium, Salmonella
Enteritidis and Salmonella Gallinarum. Microbiology 146 (Pt
12), 3217–3226.
Kogut, M.H., McGruder, E.D., Hargis, B.M., Corrier, D.E.,
DeLoach, J.R., 1995. In vivo activation of heterophil function
in chickens following injection with Salmonella enteritidis-
immune lymphokines. J. Leukoc. Biol. 57, 56–62.
Kogut, M.H., Rothwell, L., Kaiser, P., 2003. Differential regulation
of cytokine gene expression by avian heterophils during recep-
tor-mediated phagocytosis of opsonized and nonopsonized Sal-
monella enteritidis. J. Interferon Cytokine Res. 23, 319–327.
Leveque, G., Forgetta, V., Morroll, S., Smith, A.L., Bumstead, N.,
Barrow, P., Loredo-Osti, J.C., Morgan, K., Malo, D., 2003.
Allelic variation in TLR4 is linked to susceptibility to Salmo-
nella enterica serovar Typhimurium infection in chickens.
Infect. Immun. 71, 1116–1124.
Lynn, D.J., Lloyd, A.T., O’Farrelly, C., 2003. In silico identification
of components of the Toll-like receptor (TLR) signaling path-
way in clustered chicken expressed sequence tags (ESTs). Vet.
Immunol. Immunopathol. 93, 177–184.
Medzhitov, R., Janeway Jr., C.A., 2000. How does the immune
system distinguish self from nonself? Semin. Immunol. 12,
185–188, discussion 257–344.
Meijer, A.H., Gabby Krens, S.F., Medina Rodriguez, I.A., He, S.,
Bitter, W., Ewa Snaar-Jagalska, B., Spaink, H.P., 2004. Expres-
sion analysis of the Toll-like receptor and TIR domain adaptor
families of zebrafish. Mol. Immunol. 40, 773–783.
Nagase, H., Okugawa, S., Ota, Y., Yamaguchi, M., Tomizawa, H.,
Matsushima, K., Ohta, K., Yamamoto, K., Hirai, K., 2003.
Expression and function of Toll-like receptors in eosinophils:
activation by Toll-like receptor 7 ligand. J. Immunol. 171, 3977–
3982.
Oshiumi, H., Tsujita, T., Shida, K., Matsumoto, M., Ikeo, K., Seya,
T., 2003. Prediction of the prototype of the human Toll-like
receptor gene family from the pufferfish, Fugu rubripes, gen-
ome. Immunogenetics 54, 791–800.
Rehli, M., 2002. Of mice and men: species variations of Toll-like
receptor expression. Trends Immunol. 23, 375–378.
Rock, F.L., Hardiman, G., Timans, J.C., Kastelein, R.A., Bazan, J.F.,
1998. A family of human receptors structurally related to
Drosophila Toll. Proc. Natl. Acad. Sci. USA 95, 588–593.
Sebastiani, G., Leveque, G., Lariviere, L., Laroche, L., Skamene, E.,
Gros, P., Malo, D., 2000. Cloning and characterization of the
murine toll-like receptor 5 (Tlr5) gene: sequence and mRNA
expression studies in Salmonella-susceptible MOLF/Ei mice.
Genomics 64, 230–240.
Smith Jr., M.F., Mitchell, A., Li, G., Ding, S., Fitzmaurice, A.M.,
Ryan, K., Crowe, S., Goldberg, J.B., 2003. Toll-like receptor
(TLR) 2 and TLR5, but not TLR4, are required for Helicobacter
pylori-induced NF-kappa B activation and chemokine expres-
sion by epithelial cells. J. Biol. Chem. 278, 32552–32560.
Takeuchi, O., Kawai, T., Sanjo, H., Copeland, N.G., Gilbert, D.J.,
Jenkins, N.A., Takeda, K., Akira, S., 1999. TLR6: A novel
member of an expanding toll-like receptor family. Gene 231, 59–
65.
Thoma-Uszynski, S., Kiertscher, S.M., Ochoa, M.T., Bouis, D.A.,
Norgard, M.V., Miyake, K., Godowski, P.J., Roth, M.D., Modlin,
R.L., 2000. Activation of toll-like receptor 2 on human dendritic
cells triggers induction of IL-12, but not IL-10. J. Immunol. 165,
3804–3810.
Tirunagaru, V.G., Sofer, L., Cui, J., Burnside, J., 2000. An expressed
sequence tag database of T-cell-enriched activated chicken
splenocytes: sequence analysis of 5251 clones. Genomics 66,
144–151.
Page 11
M. Iqbal et al. / Veterinary Immunology and Immunopathology 104 (2005) 117–127 127
Werling, D., Jungi, T.W., 2003. TOLL-like receptors linking innate
and adaptive immune response. Vet. Immunol. Immunopathol.
91, 1–12.
Wigley, P., Hulme, S.D., Bumstead, N., Barrow, P.A., 2002. In vivo
and in vitro studies of genetic resistance to systemic salmonel-
losis in the chicken encoded by the SAL1 locus. Microbes Infect.
4, 1111–1120.
Zarember, K.A., Godowski, P.J., 2002. Tissue expression of human
Toll-like receptors and differential regulation of Toll-like recep-
tor mRNAs in leukocytes in response to microbes, their pro-
ducts, and cytokines. J. Immunol. 168, 554–561.
Zhang, D., Zhang, G., Hayden, M.S., Greenblatt, M.B., Bussey, C.,
Flavell, R.A., Ghosh, S., 2004. A toll-like receptor that prevents
infection by uropathogenic bacteria. Science 303, 1522–1526.
Zhang, S., Lillehoj, H.S., Ruff, M.D., 1995. Chicken tumor necrosis-
like factor. I. In vitro production by macrophages stimulated
with Eimeria tenella or bacterial lipopolysaccharide. Poult. Sci.
74, 1304–1310.