Vet. Res. 37 (2006) 767–778 767 c INRA, EDP Sciences, 2006 DOI: 10.1051/vetres:2006034 Original article Extensive rearing hinders Maedi-Visna Virus (MVV) infection in sheep Iratxe La , Ramón A. Ja , Jesse Ba , Beatriz Ab , Damián DA´ b , Luis L´ c , Juan Bc , Eduardo Ba , d * a Instituto Vasco de Investigación y Desarrollo Agrario-NEIKER, Derio, Bizkaia, Spain b Instituto de Agrobiotecnología y Recursos Naturales, CSIC-UPNA, Pamplona, Spain c Patología Animal, Universidad de Zaragoza, Zaragoza, Spain d Enfermedades Parasitarias, Facultad de Veterinaria, Universidad de Murcia, 30100 Campus de Espinardo, Murcia, Spain (Received 10 October 2005; accepted 3 May 2006) Abstract – Maedi-Visna Virus (MVV) seroprevalence and its relationship with housing and mode of rearing of replacement ewe-lambs was investigated in 38 non-randomly selected sheep-flocks in Spain. They included extensive lamb-producing Manchega cross-bred flocks raised almost perma- nently at pasture, semi-intensive Latxa dairy flocks housed 2–8 months/year and intensively raised Assaf dairy flocks housed most time and at higher stocking density in less ventilated buildings than other flocks. Most flocks raised replacement lambs naturally with their dams until weaning and as a separate flock thereafter until lambing at one year of age. Seroprevalence (95% confidence inter- vals) was 77%, 25% and 5% (4–6) in intensive, semi-intensive and extensive flocks, respectively and the median (interquartile range) flock-seroprevalence was 82% (66–94) in intensive flocks, 31% (14–31) in semi-intensive flocks and 4% (0–7) in extensive flocks. Seroprevalence was lowest in one year-old sheep and increased to flock levels during the year after introduction into the adult flock in most intensive flocks and more gradually in other flocks. Adult flock seroprevalence was associated with housing time but this relationship was not evident within a particular rearing system, indicat- ing that other unknown factors are critical in horizontal MVV-transmission. Low seroprevalence in extensive flocks further supports previous indications that lactogenic MVV-infection is relatively inefficient and horizontal transmission is necessary to ensure long-term maintenance of MVV and this could explain that MVV has not been reported from countries with mainly extensively reared sheep such as Australia and New Zealand. Moreover, it indicates that MVV-control in extensive and semi-intensive flocks can be simple and inexpensive. Maedi-Visna Virus / sheep / seroprevalence / intensive and extensive rearing / Spain 1. INTRODUCTION Maedi-Visna Virus (MVV) and the closely related Caprine Arthritis En- cephalitis virus (CAEV) are grouped as Small Ruminant Lentiviruses (SRLV) be- * Corresponding author: [email protected]longing to the retroviridae family. Both can infect sheep and goats causing a slow, de- generative and fatal disease of the lungs, central nervous system, mammary gland and joints that can result in very signif- icant production losses and have major welfare implications (recently reviewed by Article available at http://www.edpsciences.org/vetres or http://dx.doi.org/10.1051/vetres:2006034
Extensive rearing hinders Maedi-Visna Virus (MVV)infection in sheep
Iratxe La, Ramón A. Ja, Jesse Ba,Beatriz Ab, Damián D Ab, Luis Lc, Juan Bc,
Eduardo Ba,d*
a Instituto Vasco de Investigación y Desarrollo Agrario-NEIKER, Derio, Bizkaia, Spainb Instituto de Agrobiotecnología y Recursos Naturales, CSIC-UPNA, Pamplona, Spain
c Patología Animal, Universidad de Zaragoza, Zaragoza, Spaind Enfermedades Parasitarias, Facultad de Veterinaria, Universidad de Murcia, 30100 Campus de
Espinardo, Murcia, Spain
(Received 10 October 2005; accepted 3 May 2006)
Abstract – Maedi-Visna Virus (MVV) seroprevalence and its relationship with housing and modeof rearing of replacement ewe-lambs was investigated in 38 non-randomly selected sheep-flocks inSpain. They included extensive lamb-producing Manchega cross-bred flocks raised almost perma-nently at pasture, semi-intensive Latxa dairy flocks housed 2–8 months/year and intensively raisedAssaf dairy flocks housed most time and at higher stocking density in less ventilated buildings thanother flocks. Most flocks raised replacement lambs naturally with their dams until weaning and asa separate flock thereafter until lambing at one year of age. Seroprevalence (95% confidence inter-vals) was 77%, 25% and 5% (4–6) in intensive, semi-intensive and extensive flocks, respectivelyand the median (interquartile range) flock-seroprevalence was 82% (66–94) in intensive flocks, 31%(14–31) in semi-intensive flocks and 4% (0–7) in extensive flocks. Seroprevalence was lowest in oneyear-old sheep and increased to flock levels during the year after introduction into the adult flock inmost intensive flocks and more gradually in other flocks. Adult flock seroprevalence was associatedwith housing time but this relationship was not evident within a particular rearing system, indicat-ing that other unknown factors are critical in horizontal MVV-transmission. Low seroprevalence inextensive flocks further supports previous indications that lactogenic MVV-infection is relativelyinefficient and horizontal transmission is necessary to ensure long-term maintenance of MVV andthis could explain that MVV has not been reported from countries with mainly extensively rearedsheep such as Australia and New Zealand. Moreover, it indicates that MVV-control in extensive andsemi-intensive flocks can be simple and inexpensive.
longing to the retroviridae family. Both caninfect sheep and goats causing a slow, de-generative and fatal disease of the lungs,central nervous system, mammary glandand joints that can result in very signif-icant production losses and have majorwelfare implications (recently reviewed by
Article available at http://www.edpsciences.org/vetres or http://dx.doi.org/10.1051/vetres:2006034
[21, 24]). Viral tissue tropism depends onthe infecting strain and on the host’s ge-netics. Disease is often not apparent untiltwo years or more after initial infection,which occurs from ingesting colostrum andmilk from infected dams and from hor-izontal contact with lung secretions andexcretions from infected sheep. There isevidence that transplacental infection mayoccur, however this route of infection isconsidered of limited epidemiological im-portance and there is now ample epidemi-ological and experimental evidence indi-cating that horizontal infection is the mainform of SRLV-infection in animals raisedwith some degree of confinement [7].For example, in semi-intensively rearedLatxa dairy sheep flocks housed for 2–6 months/year the efficiency of horizontalinfection is such that lactogenic transmis-sion does not significantly increase therisk of MVV-seroconversion [6]. More-over, there is some recent evidence thatSRLV-infection is low or even absent inextensively reared sheep flocks and goatherds in Patagonia [25,26]. The risk of lac-togenic SRLV-infection has only been par-tially quantified and the percentage of nat-urally raised lambs that become infectedfrom suckling colostrum from seropositivedams has been estimated at approximatelyonly 20% [2, 3]. If the risk of lactogenicinfection during the preweaning period isbelow 1 (100%) then it is evident that someother form of infection is necessary toachieve an Re (effective reproduction ratio:average number of secondary infectionsproduced by one infected individual) valueabove one, necessary for infection to lastin the population [4]. It is possible that insome extensive sheep production systems,sheep do not have sufficiently close contactfor horizontal infection to occur, but thishas not been investigated. Moreover, littleis known of the type and extent of contactbetween infected and uninfected animalsnecessary for MVV-infection to occur. Un-infected sheep introduced in an infected
flock may quickly become infected, but insome cases uninfected sheep can be housedwith other infected sheep for several yearswithout becoming infected and the reasonsfor this are unknown [17].
The limited knowledge on the mech-anisms of MVV-transmission betweensheep contrasts with the numerous sero-prevalence studies of Maedi-Visna world-wide [14, 24], indicating that MVV hasbeen reported from most major sheep pro-ducing countries except Australia and NewZealand [12]. Several studies have shownsome evidence of breed associated sus-ceptibility or resistance to MVV-infectionand disease [8, 9, 13, 23, 31]. However,very few have attempted to investigate therelationship between infection and sheepmanagement and transmission and con-trol implications. In a previous study byBerriatua et al. [6], incidence of infec-tion in semi-intensive Latxa dairy flockswas relatively low and in most flocks thenumber of sheep that seroconverted waslower than the number of sheep culled withflock culling percentages being 14–25%.This offers the possibility of graduallyreducing flock seroprevalence by selec-tive culling without increasing the flock’sculling percentage and this represents analternative to the more drastic traditionalMVV-control methods based on flock test-ing and culling of all seropositive animalsand their young progeny (or on developinga separate MVV-free flock) [14] and high-lights the need for similar investigations inother sheep production systems. In orderto investigate this, long-term longitudinalstudies have been initiated in extensiveand intensive sheep production systemsin several regions in Spain. This articlereports MVV-seroprevalence, its relation-ship with housing and mode of rearingpreweaned replacement lambs and inves-tigates MVV in control lamb-producingextensive flocks and intensive and semi-intensive dairy flocks from southern, cen-tral and northern Spain, respectively.
Maedi-visna and sheep management 769
2. MATERIALS AND METHODS
2.1. Study design and population
Thirty-eight flocks were investigated in-cluding 10 intensive Assaf dairy flocks inCastile-León, 5 semi-intensive Latxa dairyflocks in the Basque Country and 23 ex-tensive lamb-producing Manchega crossMerino or Talaverana flocks in Castile-La Mancha. Resources were not availableto investigate more flocks. Moreover, itwas initially predicted that approximately30 flocks of 300 sheep with a similar num-ber of flocks from each production systemcould be investigated. However, differentnumbers of flocks from each system wereselected because average flock sizes var-ied between systems, there was no dataon seroprevalence of MVV-infection in theextensive Manchega cross-bred flocks andit was necessary to accommodate the studydesign to detect MVV in this system. Fur-thermore, given the good knowledge of theepidemiology of MVV-infection in semi-intensive Latxa flocks available [2,3,6,17],it was decided to allocate more resources toinvestigating MVV in other sheep produc-tion systems.
The Basque Country is situated in theAtlantic northern region of Spain wherethe climate is mild and rain ensures greenpastures throughout the year. In contrast,Castile represents the Spanish CentralPlain situated at around 700 m above sealevel, being dominated by a dry continen-tal climate. Castile-León is the northernpart of the Plain and Castile-La Manchais the southern part of the Plain. In 2004,there were 22.7 million sheep in Spain in-cluding 15% dairy sheep. Castile-León had4.3 million sheep and produced 60% of thesheep milk commercialised in Spain andAssaf dairy sheep were present in 86% ofthe dairy flocks in the region. Latxa sheeprepresent 90% of the 0.4 million ovinesin the Basque Country. Manchega crossTalaverana and Merino sheep reared ex-
tensively for lamb production like in thisstudy, represent a minority of the 3.2 mil-lion sheep in Castile-La Mancha. How-ever, most of the 7.8 million sheep inthe neighbouring regions of Extremaduraand Andalucía, represented by the Merinobreed reared for fine wool production in thepast and for lamb production nowadays,are raised similarly and more extensively[10, 18, 27].
Typical intensive Assaf dairy flocks inCastile-León are made of 200–1000 sheep,the animals are reared almost permanentlyindoors, their oestrous is not markedly sea-sonal and they are bred for three lambingsin two years. They have a milking period of7–9 months/year and the average milk pro-duction is 400 kg of milk per ewe (kme).In contrast, Latxa dairy sheep flocks aremostly < 500 sheep, they lamb once a yearin the winter, have a five month milkingperiod with an average milk production of135 kme, and are mostly raised on pas-ture and housed mostly during lactationat night and in rainy weather. Manchegacross-bred sheep are managed in large sizeflocks sometimes including several thou-sand sheep, they are raised extensively inlow yielding “dehesa” pastures, and theyare not seasonally polyoestrus and cycleall year round. In all three systems, mostlambs are weaned at 4–6 weeks of age andsold for slaughter or in Castile, also forfattening in feedlots. The exceptions are re-placement lambs that remain with the damfor 2–3 months and that are thereafter keptas a separate flock until after their firstlambing at 12–14 month-old, when theyjoin the adult flock.
2.2. Selection of flocks and animals
The ten intensive Assaf dairy flockswere selected from a list of 30 flocks un-der the care of a veterinary practitionerconcerned with the high incidence of clini-cal Maedi-Visna (MV) including animals
770 I. Leginagoikoa et al.
Table I. Flock origin, number, size, breed, production system, median weaning age of replacementlambs and housing time and space in an MVV seroprevalence study in sheep in Spain in 2003–2004.
with a typical yet comparatively uncom-mon, Visna nervous form [5]. They werethe only ones in the list registered withthe local Milk-Board. Sheep were born andbred at home, uniquely identified and haddates of birth accurate to the month andyear for three years prior to the start ofthis study. The five selected semi-intensiveLatxa dairy flocks had participated in pre-vious MVV serological and control studiescontinuously since the early 1990s, theyhad no reported MV clinical cases for al-most a decade and the farmers were keen tocontinue testing for MVV-infection. Theywere considered among the most profes-sionalised and intensively raised flocks inthe region and except one flock, nonehad incorporated outside sheep for sev-eral years preceding the study. Sheep wereuniquely identified and had precise dates ofbirth. The 23 Manchega cross-bred flocksselected for the study were from clientsof a veterinarian that had expressed inter-est in investigating MVV although he had
never diagnosed a case of MVV-infection.To maximise resources, selected flocks in-cluded all the flocks under his care with< 500 sheep. Sheep were born and bredat home, uniquely identified and dates ofbirth were accurate to the season and year.Not all sheep in these flocks were sam-pled and tested for MVV-antibodies andthe number of sheep selected in each flockwas calculated to allow detecting at leastone seropositive sheep with a 95% proba-bility, assuming a flock seroprevalence ofat least 5% [32].
2.3. Flock husbandry and housing
Every farm was visited to complete aflock management questionnaire by per-sonally interviewing the farmer and tomeasure sheep housing facilities. Table Idescribes flocks origin, number, size, breedand production system, weaning age ofreplacement lambs and sheep’s housingtime and space.
Maedi-visna and sheep management 771
Housing and management of replace-ment lambs was characteristic of the sheepproduction systems described above. Theywere naturally raised, except in three Latxaflocks where lambs stayed with their damsuntil 24-h-old and were then artificiallyraised with milk replacer until 6 weeks ofage and one Assaf flock where lambs wereraised artificially from birth with bottle-feeding of the dam’s colostrum during thefirst day of life and milk replacer until6-week-old. After being separated fromtheir mothers, the replacement lambs inall flocks were raised as a separate flockuntil a few weeks before lambing at ap-proximately 1 year of age. During thistime, the lambs and adults used separatefields or field partitions and different build-ings except one of the Latxa flocks thatraised lambs artificially preweaning andtwo Assaf flocks that raised lambs withtheir dams preweaning, which used sepa-rate pens within the adult flock buildingwhen housing sheep.
The time sheep were indoors and thecharacteristics of the sheep buildings var-ied between and within systems and foradult and replacement sheep (Tab. I). As-saf adults were housed for longer peri-ods, at somewhat greater stocking densityand in more poorly ventilated buildingsthan adults in other systems. This wasalso the case for replacement lambs exceptthat Latxa and Manchega cross-bred flockshoused lambs for longer than the adults,flooring space for lambs was slightlysmaller in Latxa flocks compared to As-saf flocks and lamb sheds in Manchegacross-bred flocks were smaller than thosefor adults.
2.4. Sampling and MVV-antibodytesting
All sheep ≥ 1 year-old in inten-sive and semi-intensive flocks and 50–65 sheep/flock aged/ ≥ 1 year-old, in ex-tensive flocks were blood sampled from the
jugular vein with vacuum tubes with no an-ticoagulant. Sampling took place betweenNovember 2003 and April 2004 when re-placement lambs were 12–14 months oldand had not yet joined the adult flock. Inthe laboratory, serum was recovered andkept frozen at –20 ◦C until tested for MVV-antibodies using an indirect commercialELISA test (Elitest, Hyphen BIOMED,Neuville sur Oise, France). The assay hasa sensitivity (se) and specificity (sp) and95% confidence intervals compared to theAgar Gel Imunodifussion Test (AGID) andwestern-blotting of 99.4% (98.4–99.8) and99.3% (98.7–99.6), respectively [30].
Single tests were carried out in serumsamples from Castile-León and BasqueCountry and samples were deemed pos-itive for ELISA optical densities (OD)above or equal to the cut-off point andnegative otherwise. The cut-off point wascalculated according to the manufacturer’sinstructions using the following formula:Cut-off = (ODP450−595 – ODN450−595)/4
+ ODN450−595where, OD450−595: optical density at450 nm using 595 nm as reference wave-length, of the positive control (P) andnegative control (N).
Moreover, to increase the precision ofthe estimated low seroprevalence in ex-tensive Castile-La Mancha flocks, a majorfinding of this investigation, samples withOD values of 0.1 units below or abovethe cut-off value, more common in exten-sive than in intensive flocks (Tab. II), weretested once or twice again. The sampleswere tested once again only if the result ofthe second test was similarly above or be-low the cut-off point and accordingly, wereclassified as positive or negative. Instead,if the second test result was different fromthe first result a third assay was performedand the samples were considered positiveif two of the three test results were abovethe cut-off OD values and inconclusiveotherwise. Inconclusive results were notincluded in seroprevalence calculations. To
772 I. Leginagoikoa et al.
Table II. Age-specific distribution of antibody-ELISA ROD values in seropositive sheep from 38extensively raised cross-bred lamb-producing flocks, semi-intensively raised latxa dairy flocks andintensively raised Assaf flocks in Spain.
a Manchega-Merino and Manchega-Talaverana cross-bred ewes.b One sheep only.
standardise the results, relative OD (ROD)were calculated by dividing the sample ODby the OD of the plate’s positive control.
2.5. Statistical analysis
EpiInfo 2002 (CDC, Atlanta, USA)was used for all the statistical analy-sis, including sample size calculations,frequency distributions, chi-square testswith the Yate correction to compare pro-portions, non-parametric Kruskal-Wallisanalysis to compare means and linear re-gression to investigate the relationship be-tween ewe-flock seroprevalence and inde-pendent flock-average variables, includingthe number of days spent indoors per yearand area of floor and open space andvolume of housing per adult sheep avail-able. All variables were incorporated inthe models as continuous variables. Themodels were fitted using the least-squaremethod. Partial F-tests were used to assessthe significance of independent variablestaken at the 5% (p < 0.05) level for adouble-sided test.
3. RESULTS
3.1. MVV-seroprevalence
Seroprevalence was 77% (3046/3974)in intensive Assaf dairy sheep, 25%(391/1570) in semi-intensive Latxa dairysheep and 5% (72/1360) in extensive
Manchega cross-bred sheep (p < 0.001).Seroprevalence (95% CI) in males and fe-males was, respectively, 75% and 77% inintensive flocks (p > 0.05), 19% and 25%in semi-intensive flocks (p < 0.05) and 8%(3–14) and 5% (4–6) in extensive flocks(p > 0.05).
However, seroprevalence varied be-tween flocks within sheep production sys-tems. Table III presents the estimated sero-prevalence in ewes and the distributionof ewe-flock seroprevalence in the threesystems. Ewe seroprevalence was signif-icantly higher in Assaf ewes than in theother ewes and in Latxa ewes comparedto Manchega cross-bred ewes (p < 0.05).Among the latter, 6 flocks had 0% sero-prevalence and the highest flock sero-prevalence was 21%, found in one flock(Tab. III). In contrast, seroprevalence inAssaf flocks was 44–96% and > 80% inhalf the flocks. Seroprevalence in Latxaflocks was intermediate between the othertwo breed groups and ranged between 5%and 47% (Tab. III).
3.2. Age-specific MVV-seroprevalenceand relationship with modeof rearing of replacement sheepand adult flock seroprevalence
Age-specific seroprevalence patterns ininfected flocks differed between and withinsheep production systems depending on
Maedi-visna and sheep management 773
Table III. Ewe and ewe-flock MVV-seroprevalence in 38 extensively raised cross-bred lamb-producing flocks, semi-intensively raised Latxa dairy flocks and intensively raised Assaf flocksin Spain.
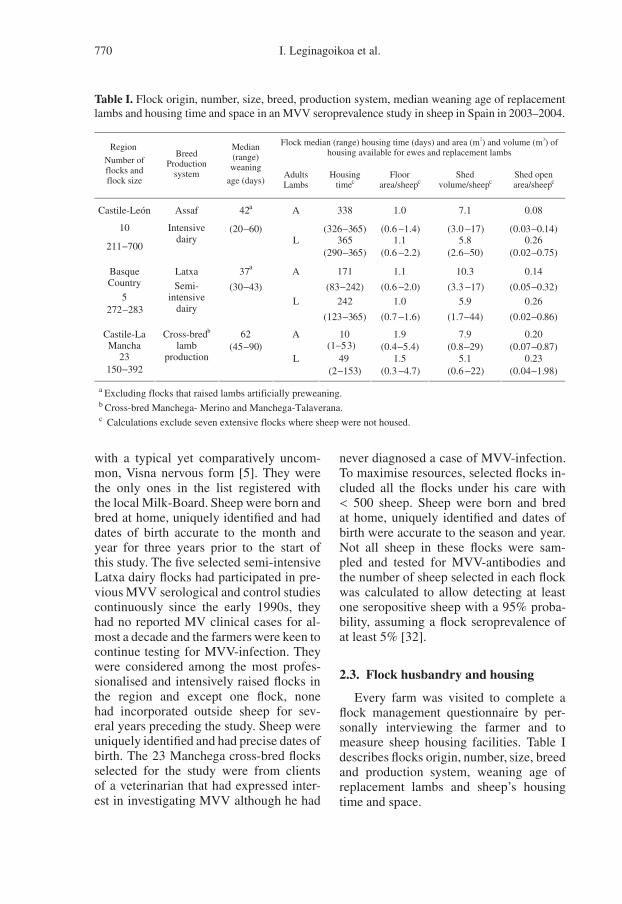
the mode of rearing of replacement sheepduring their first year of life and adult flockseroprevalence. This is graphically shownin Figure 1 where AF, AS, NF and NS re-fer to the mode of rearing of replacementlambs during the first year of life (A: ar-tificially with milk replacer, N: naturallywith the preweaning dam and F: in theadult flock building and S: in a separateshed until 1-year-old). The rate of increasein seroprevalence with age was very slowin extensive flocks and one semi-intensiveflock, moderate in the remaining semi-intensive flocks and one intensive flock andhigh in the other intensive flocks.
In intensive Assaf flocks, seropreva-lence among one-year-old sheep was thelowest (20%) in the flock where lambswere raised artificially preweaning withovine colostrum and milk replacer (AS)and 25–80% elsewhere according to adultseroprevalence and apparently independentof post-weaning place of rearing (Fig. 1).However, at 2 years of age, seropreva-lence was > 60% in all flocks except onewhere seroprevalence was lower and onlyreached 60% among ≥ 4-year-old sheep(Fig. 1). The age-specific pattern in mostsemi-intensive Latxa flocks was close tothat in the latter intensive Assaf flock ex-cept that in the Assaf flock seroprevalencewas greater among one year-old sheep andincreased more slowly thereafter comparedto two Latxa flocks that raised lambs arti-
ficially preweaning (AS and AF) (Fig. 1).Like in Assaf flocks, there was little ev-idence in Latxa flocks of an associationbetween place of rearing post-weaning andseroprevalence at one year of age.
3.3. Relationship between adultflock-seroprevalence andhousing variables
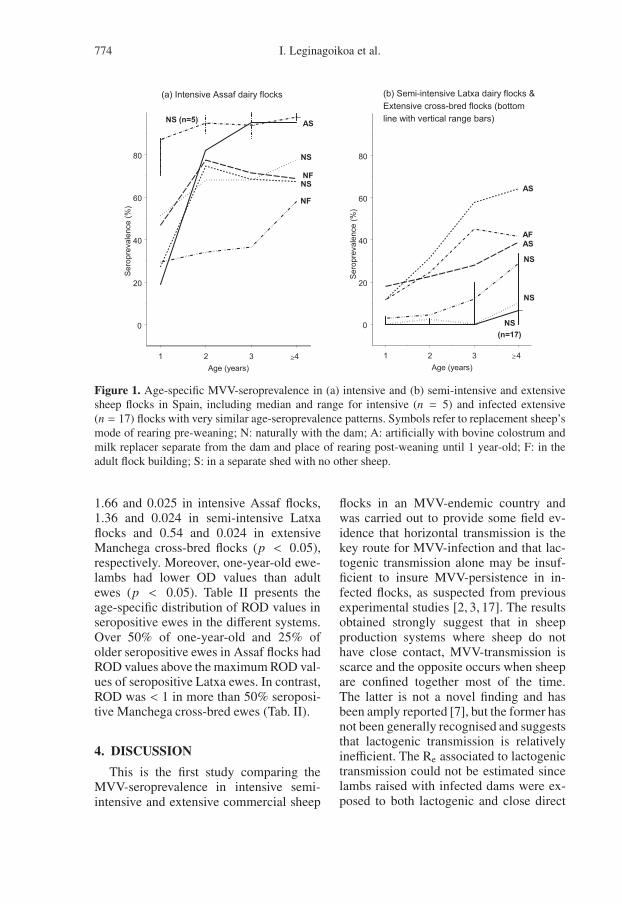
The regression models confirmed thesignificant positive association betweenseroprevalence in the adult flock and thenumber of days sheep are housed per year(p < 0.001). However, these two variableswere not associated within a particularsheep rearing system (results not shown).Moreover, adult flock seroprevalence wasnot associated with open area, flooring areaor volume of shed available per sheep inmodels containing or excluding housingtime. Figure 2 shows the observed un-adjusted relationship between adult flockseroprevalence and housing time per year.The percentage of variation explained bythe bivariate model was 86% (r2 = 0.86)(p < 0.001) and did not change after in-cluding any of the other independent vari-ables relating to stocking density and openarea per sheep available.
3.4. ELISA-antibody optical densities
Mean ELISA-antibody OD values forseropositive and seronegative ewes were,
774 I. Leginagoikoa et al.
Figure 1. Age-specific MVV-seroprevalence in (a) intensive and (b) semi-intensive and extensivesheep flocks in Spain, including median and range for intensive (n = 5) and infected extensive(n = 17) flocks with very similar age-seroprevalence patterns. Symbols refer to replacement sheep’smode of rearing pre-weaning; N: naturally with the dam; A: artificially with bovine colostrum andmilk replacer separate from the dam and place of rearing post-weaning until 1 year-old; F: in theadult flock building; S: in a separate shed with no other sheep.
1.66 and 0.025 in intensive Assaf flocks,1.36 and 0.024 in semi-intensive Latxaflocks and 0.54 and 0.024 in extensiveManchega cross-bred flocks (p < 0.05),respectively. Moreover, one-year-old ewe-lambs had lower OD values than adultewes (p < 0.05). Table II presents theage-specific distribution of ROD values inseropositive ewes in the different systems.Over 50% of one-year-old and 25% ofolder seropositive ewes in Assaf flocks hadROD values above the maximum ROD val-ues of seropositive Latxa ewes. In contrast,ROD was < 1 in more than 50% seroposi-tive Manchega cross-bred ewes (Tab. II).
4. DISCUSSION
This is the first study comparing theMVV-seroprevalence in intensive semi-intensive and extensive commercial sheep
flocks in an MVV-endemic country andwas carried out to provide some field ev-idence that horizontal transmission is thekey route for MVV-infection and that lac-togenic transmission alone may be insuf-ficient to insure MVV-persistence in in-fected flocks, as suspected from previousexperimental studies [2, 3, 17]. The resultsobtained strongly suggest that in sheepproduction systems where sheep do nothave close contact, MVV-transmission isscarce and the opposite occurs when sheepare confined together most of the time.The latter is not a novel finding and hasbeen amply reported [7], but the former hasnot been generally recognised and suggeststhat lactogenic transmission is relativelyinefficient. The Re associated to lactogenictransmission could not be estimated sincelambs raised with infected dams were ex-posed to both lactogenic and close direct
Maedi-visna and sheep management 775
Figure 2. Relationship between flock seroprevalence and housing time in Spanish extensively raisedcross-bred lamb-producing flocks, semi-intensively raised Latxa dairy flocks and intensively raisedAssaf dairy flocks (n = 38).
horizontal infection except in one Assafflock where lambs were raised under iso-lation with ovine colostrum and milk re-placement. Interestingly, seroprevalence inone-year old sheep in this flock was 20%,very similar to the experimentally esti-mated seroprevalence associated to natu-rally suckled colostrum from seropositiveewes [2]. However, the existence of sev-eral extensive flocks with 0% seropreva-lence is compatible with Re associated tocolostrum and milk being below or close toone. Consequently, MVV-control and erad-ication under such circumstances could beeasy and inexpensive. The results in exten-sive flocks may help to explain the appar-ent absence of MVV in geographic areassuch as Australasia, where most sheep areraised extensively. A further finding of thisstudy was that antibody OD of seropositivesheep differed depending on flock sero-prevalence and were greater in aged adultsthan in one year-old sheep. This could berelated to greater virus challenge from pos-sibly a larger variety of infectious strains towhich hosts are exposed.
Given the limited number of flocks in-vestigated and the criteria used to selectflocks it could be argued that the observedseroprevalence is biased and not repre-sentative of these sheep production sys-tems and that MVV-transmission betweensheep readily occurred and sheep did ordid not become infected as a result of forexample, genetic breed-associated suscep-tibility and resistance to MVV-infection.Flock selection within intensive and ex-tensive systems was not random and waslimited to flocks under the care of two vet-erinary practitioners. As indicated before,the veterinarian responsible for the inten-sively raised flocks had expressed concernfor MVV-infection and the contrary wasthe case for the veterinarian looking af-ter the extensive flocks and this could bea source of strong selection bias. How-ever, similarly high SRLV-seroprevalenceis typical for example in intensive dairygoat herds in California [28] and in Ice-landic sheep kept in close confinementin the winter [20] and similar low andnull SRLV-seroprevalence was reported in
776 I. Leginagoikoa et al.
extensive flocks in Patagonia [25, 26]. Fur-ther bias might have been introduced byselecting the smaller flocks in the extensivesystem and those registered with the lo-cal milk-board among intensive flocks, butit is unlikely that either factor contributesgreatly to explaining the enormous differ-ences in MVV-seroprevalence in these twosheep production systems. Moreover, al-though certain alleles have been identifiedthat confer goats and cows some degreeof resistance to lentiviral infections [19,29] and there are indications of inherita-ble susceptibility and resistance to MVV-seroconversion in sheep [6], there is no ev-idence or previous suggestions, that majordifferences in flock MVV-seroprevalenceare attributable to genetic differences be-tween sheep. Breed-associated differencesin susceptibility to infection have beenproposed. However, differences are notconsistent and no reports exist of partic-ular breeds that always remain free orvirtually free of MVV-infection after ex-posure to the virus. Resistance has alsobeen linked to family lines within particu-lar breeds, based on evidence that infectionin flocks is often restricted to the progenyof infected ewes [14]. From these argu-ments it is reasonable to conclude that it isvery unlikely that the observed seropreva-lence in extensive and intensive flocks wasstrongly confounded by selection bias andbreed differences. In contrast, the observedseroprevalence in semi-intensively raisedLatxa dairy flocks (< 30%) is unlikely tobe typical of this sheep production sys-tem because selected flocks had all imple-mented some form of MVV-control andcan be considered low. Indeed, the esti-mated MVV-seroprevalence in the BasqueCountry was 55% in a large survey1 using
1 Lorenzo González, El Maedi o neumonía pro-gresiva en el conjunto de las enfermedadesrespiratorias crónicas del ganado ovino en laComunidad autónoma Vasca, Doctoral thesis,Universidad de Zaragoza, Spain, 1989.
the AGID test, which is less sensitive thanthe ELISA used here.
The estimated seroprevalence in theBasque Country and the similarity inthe age-specific seroprevalence patternsin some Assaf and Latxa flocks in thepresent study, suggest that the epidemiol-ogy of MVV-infection in these two pro-duction systems may not differ greatly.Moreover, seroprevalence differences be-tween and within Assaf and Latxa flockssuggests that MVV-infection depends onadditional factors, other than the degreeof contact between infected and non-infected sheep. This is further supportedby the finding that neither housing timenor any of the other housing variablesinvestigated, explained differences in sero-prevalence between intensive flocks. Otherfactors affecting MVV-infection may in-clude the sheep’s genetic susceptibility,the virulence of MVV-strains associatedwith a cytopathic effect [15, 16], the pres-ence of other diseases such as ovine Pul-monary Adenomatosis that increase therisk of MVV-infection [11] and otherunrecorded management and productionpractices. Specifically, the animal to ani-mal dynamics of SRLV-horizontal infec-tion is not well understood and little isknown of certain husbandry aspects that af-fect the risk of MVV-infection. Althoughnose to nose contact between sheep issuspected to be a major risk factor forMVV-infection, all the intensive flocks inthis study used feeding troughs that al-lowed direct nose to nose contact betweensheep. Furthermore, the lack of an as-sociation between seroprevalence in oneyear-old sheep and place of rearing (flockbuilding or separate shed), suggests that in-direct contact transmission of SRLV maynot be very efficient. This has been inves-tigated, with discrepancies. For example,no CAEV-transmission between infectedand non-infected does penned separatelyin the same building was found in a study[1], whereas in another, indirect CAEV-
Maedi-visna and sheep management 777
transmission between does in differentpens in the same shed was reported andthe risk of infection depended on the de-gree of separation between the animals[22]. Finally, little is known concerningthe effect of production pressure on MVV-infection, however it has been consideredan important risk factor [17] and the re-lationship between milk production andMVV-incidence of seroconversion in theseflocks warrants further investigation.
Control wise, the results of this studyare optimistic for some intensive Assafflocks with moderate seroprevalence sim-ilar to some semi-intensively Latxa flockswhere it was shown that seroprevalencecould be gradually reduced by selectivelyculling seropositives [6] but it is unlikelythat selective culling alone would permitelimination of infection in intensive flockswith high MVV-prevalence. Clearly moreresearch is required to elucidate SRLV-excretion from infected animals and con-tact transmission between animals and theways to avoid it in order to better controlthis lentiviral infection.
ACKNOWLEDGEMENTS
This work was financially supported bygrants AGL2003-08977-c03-03 of the Ministe-rio de Ciencia y Tecnología and SED2003011and SED2005008 of the Departamento de Agri-cultura of the Basque Government. We are alsoindebted for this to study flock owners forallowing us to work on their farms and to veteri-narians Javier Otaola Otxoa, José Angel FuertesMiguélez and Pedro José Mora Fernández fortheir invaluable help in selecting and samplingtheir client flocks.
REFERENCES
[1] Adams D.S., Klevjer-Anderson P.,Carlson J.L., McGuire T.C., GorhamJ.R., Transmission and control of caprinearthritis-encephalitis virus, Am. J. Vet. Res.44 (1983) 1670–1675.
[2] Álvarez V., Arranz J., Daltabuit-Test M.,Leginagoikoa I., Juste R.A., AmorenaB., de Andrés D., Luján L.L., BadiolaJ.J., Berriatua E., Relative contribution ofcolostrum from Maedi-Visna virus (MVV)infected ewes to MVV-seroprevalence inlambs, Res. Vet. Sci. 78 (2005) 237–243.
[3] Álvarez V., Daltabuit-Test M., Arranz J.,Leginagoikoa I., Juste R.A., Amorena B.,de Andrés D., Luján L.L., Badiola J.J.,Berriatua E., PCR detection of colostrum-associated Maedi-Visna virus (MVV) infec-tion and relationship with ELISA-antibodystatus in lambs, Res. Vet. Sci. 80 (2006) 226–234.
[4] Anderson R.M., May R.M., InfectiousDiseases of Humans: dynamics and control,Oxford University Press, Oxford, UnitedKingdom, 1992.
[5] Benavides J., Gómez N., Gelmetti D.,Ferreras M.C., García-Pariente C., FuertesM., García-Marín J.F., Pérez V., Diagnosisof the nervous form of maedi-visna infectionwith a high frequency in sheep in Castilla yLeón, Spain, Vet. Rec.158 (2006) 230–235.
[6] Berriatua E., Ávarez V., Extramiana B.,González L., Daltabuit M., Juste R.,Transmission and control implications ofseroconversion to Maedi-Visna virus inBasque dairy-sheep flocks, Prev. Vet. Med.60 (2003) 265–279.
[7] Blacklaws B.A., Berriatua E., TorsteinsdottirS., Watt N.J., de Andres D., Klein D.,Harkiss G.D., Transmission of small rumi-nant lentiviruses, Vet. Microbiol. 101 (2004)199–208.
[9] Dawson M., Pathogenesis of maedi-visna,Vet. Rec. 120 (1987) 451–454.
[10] Esteban-Muñoz C., Tejon-Tejon D.,Catálogo de razas autóctonas españolas,I. Especies ovina y caprina, Ministerio deAgricultura, Pesca y Alimentación, ISBN:8474790905, 1986.
[11] González L., Juste R.A., Cuervo L.A.,Idigoras I., Saez de Ocariz C., Pathologicaland epidemiological aspects of the coexis-tance of maedi-visna and sheep pulmonaryadenomatosis, Res. Vet. Sci. 54 (1993) 140–146.
[12] Greenwood P.L., North R.N., Kirkland P.D.,Prevalence, spread and control of caprine
778 I. Leginagoikoa et al.
arthritis-encephalitis virus in dairy goatherds in New South Wales, Aust. Vet. J. 72(1995) 341–345.
[13] Houwers D.J., Visscher A.H., Defise P.R.,Importance of ewe lamb relationship andbreed in the epidemiology of maedi-visnavirus infections, Res. Vet. Sci. 46(1989) 5–8.
[14] Houwers D.J., Economic importance, epi-demiology and control, in: Petursson G.,Hoff-Jorgensen R. (Eds.), Maedi-Visna andrelated diseases, Kluwer Academic Press,Massachussets, 1990, pp. 83–117.
[15] Lairmore M.D., Akita G.Y., Russell H.I.,DeMartini J.C., Replication and cytopathiceffects of ovine lentivirus strains in alve-olar macrophages correlate with in vivopathogenicity, J. Virol. 61 (1987) 4038–4042.
[17] Leginagoikoa I., Daltabuit-Test M., ÁlvarezV., Arranz J., Juste R.A., Amorena B.,de Andrés D., Lujan L.L., Badiola J.J.,Berriatua E., Horizontal Maedi-Visna virus(MVV) infection in adult dairy-sheep raisedunder varying MVV-infection pressure in-vestigated by ELISA and PCR, Res. Vet. Sci.80 (2006) 235–241.
[18] MAPA, Encuestas ganaderas 2004,Subdirección general de estadísticasagroalimentarias, Ministerio de Agricultura,Pesca y Alimentación, 2004, pp. 24–25.
[20] Palsson P.A., Maedi and Visna in sheep, in:Kimberlin R.H. (Ed.), Slow virus diseasesof animal and man, New York, AmericanElsevier, 1976, pp. 17–43.
[21] Pépin M., Vitu C., Russo P., Mornex J.F.,Peterhans E., Maedi-visna virus infection insheep: a review, Vet. Res. 29 (1998) 341–367.
[22] Péretz G., Bugnard F., Calavas D., Study ofa prevention programme for caprine arthritis-encephalitis, Vet. Res. 25 (1994) 322–326.
[23] Perk K., Yaniv A., Gazit A., DeMartini J.C.,Evaluation of vaccines for ovine lentivirusinfection, AIDS Res. Hum. Retroviruses 12(1996) 425–426.
[24] Peterhans E., Greenland T., Badiola J.J.,Harkiss G., Bertoni G., Amorena B.,Eliaszewicz M., Juste R.A., Krassnig R.,Lafont J.P., Lenihan P., Petursson G.,Pritchard G., Thorley J., Vitu C., MornexJ.F., Pépin M., Routes of transmission andconsequences of small ruminant lentiviruses(SRLVs) infection and eradication schemes,Vet. Res. 35 (2004) 1–38.
[25] Robles C.A., Lanari M.-R., Pérez-CentenoM., Domingo E., Brucellosis and artritis en-cephalitis survey in cross-bred gotas fromNeuquen province, Patagonia Argentina,Vet. Arg. 16 (1999) 1–6 (in Spanish).
[26] Robles C.A., Layana J.A., Cabrera R.F.,Raffo F., Cutlip R., Retrospective serolog-ical study on Maedi (progressive pneumo-nia) in sheep and arthritis-encephalitis ingoats from Patagonia, Argentina, Rev. Med.Vet. (Buenos Aires) 84 (2003) 96–99 (inSpanish).
[27] Rodríguez L., González M.A., Verberana J.,García J.I., Vitoriano B., Condiciones so-cioeconómicas y perspectivas de futuro delos ganaderos de ovino de Castilla y León, in:El ganado ovino en Castilla y León: Estudiosocio-económico y nuevas tecnologías de re-producción, Instituto Tecnológico Agrario,Junta de Castilla y León, 2005, pp. 27–52.
[28] Rowe J.D., East N.E., Thurmond M.C.,Franti C.E., Pedersen N.C., Cohort study ofnatural transmission and two methods forcontrol of caprine arthritis-encephalitis virusinfection in goats on California dairy, Am. J.Vet. Res. 53 (1992) 2386–2395.
[29] Ruff G., Regli J.G., Lazary S., Occurrenceof caprine leucocyte class I and II antigensain Saanen goats affected by caprine arthritis(CAE), Eur. J. Immunogenet. 20 (1993) 285–288.
[30] Saman E., Van Eynde G., Luján L.,Extramiana A.B., Harkiss G., Tolari F.,González L., Amorena B., Badiola J.J., Anew sensitive serological assay for detectionof lentivirus infections in small ruminants,Clin. Diagn. Lab. Immunol. 6 (1999) 734–740.
[31] Snowder G.D., Gates N.L., Climp H.A.,Gorham J.R., Prevalence and effect of sub-clinical ovine progressive pneumonia virusinfection on ewe wool and lamb production,J. Am. Vet. Med. Assoc. 197 (1990) 475–479.
[32] Thrusfield M., Veterinary Epidemiology,Blackwell Science, Oxford, UnitedKingdom, 1995, pp. 187–188.