jo ur nal ho me p age: www.elsev ier .com/ locate /neule t
xtracellular matrix components mark the territories ofircumventricular organs
ároly Pócsai ∗, Mihály Kálmánepartment of Anatomy, Histology and Embryology, Semmelweis University, Tuzolto 58, Budapest, H-1094 Hungary
i g h l i g h t s
Extracellular matrix components are abundant in circumventricular organs.Their immunoreactivities generally mark the territories of circumventricular organs.Matrix is suggested to control access of blood-borne molecules to adjacent areas.
r t i c l e i n f o
rticle history:eceived 11 November 2013eceived in revised form 5 February 2014ccepted 10 February 2014
In the central nervous system the extracellular matrix has important roles, e.g. supporting the extra-cellular space, controlling the tissue hydration, binding soluble factors and influencing their diffusion.The distribution of the extracellular matrix components in the brain has been mapped but data on thecircumventricular organs (CVOs) is not available yet. The CVOs lack the blood–brain barrier and haverelatively large perivascular spaces. The present study investigates tenascin-R and the lecticans: aggre-can, brevican, neurocan, and versican in the median eminence, the area postrema, the vascular organof the lamina terminalis, the subfornical organ, the pineal body and the subcommissural organ of therat applying immunohistochemical methods, and lectin histochemistry, using Wisteria floribunda agglu-tinin (WFA). The extracellular matrix components were found intensely expressed in the CVOs with twoexceptions: aggrecan immunoreactivity visualized only neurons in the arcuate nucleus, and the subcom-missural organ was not labeled with either WFA, or lecticans, or tenascin-R. The different labelings usually
overlapped each other. The distribution of the extracellular matrix components marked the territories ofthe CVOs. Considering these we suppose that the extracellular matrix is essential in the maintenance ofCVO functions providing the large extracellular space which is required for diffusion and other processesimportant in their chemosensitive and neurosecretory activities. The decrease of extracellular matrixbeyond the border of the organs may contribute to the control of the diffusion of molecules from theCVOs into the surrounding brain substance.
. Introduction
It has been proved that the central nervous system (CNS) has anxtracellular matrix built up mainly from similar components asn other parts of the body although some components occur onlyn the CNS. The CNS extracellular matrix has an important role inrain development, cell migration, axon growth, synaptogenesis,
rain vascularization, cell adhesion, tissue hydration, the bindingf the soluble factors, and the control of their diffusion [1,2].
In the CNS the main extracellular matrix components are thehyaluronic acid, some glycoproteins (laminin, fibronectin, tenascin)and some proteoglycans consisting of a central core protein withglycosaminoglycan side chains, mainly chondroitin sulphates. Themost abundant brain proteoglycans are the hyaluronan binding‘hyalectans’, also called ‘lecticans’, as the versican, aggrecan, neu-rocan and brevican [3–6].
The CNS extracellular matrix is found dispersed in the neuropil[4], or forming perineuronal ‘nets’ [4,6–8].
The distribution of the extracellular matrix components in thebrain has been mapped (for example see [4,9–11]) but data on the
circumventricular organs (CVOs) are not available yet, except forthe occurrence of laminin and fibronectin in the CVOs in general[12] and that of some components in the neurohypophysis [13,14].
Fig. 1. Scheme on the positions of the CVOs mentioned in the text. Mediansagittalplane, based on the photography of [16]. AP – area postrema, EM – median eminence,OVLT – vascular organ of the lamina terminalis, NH – neurohypophysis, P – pinealbody, SCO – subcommissural organ, SFO – subfornical organ.Oc–
TctrroFt
vtlam(gw
2
wi(
xl0fic
f(1AMBM1tt(ipsw
mO
ther structures to help orientation: 1 – corpus callosum, 2 – fornix, 3 – anteriorommissure, 4 – optic chiasm, 5 – interthalamic adhesion, 6 – cerebral aqueduct, 7
superior colliculus, 8 – central canal.
he CVOs are supposed to have special neurosecretory and/orhemosensitive functions, e.g. in the salt- and water-balance,he circulation, and the gastrointestinal functions. Therefore theyequire a free access to cerebral blood (a lack of the blood–brain bar-ier), and relatively large perivascular spaces. The subcommissuralrgan forms an exception having a complete blood–brain barrier.or recent reviews about the CVOs see [15–18] and for a scheme onheir positions see Fig. 1.
The present study investigates the aggrecan, brevican, neurocan,ersican and tenascin-R (the main tenascin of the mature brain) inhe median eminence, the area postrema, the vascular organ of theamina terminalis (OVLT), the subfornical organ, the pineal bodynd the subcommissural organ of the rat by immunohistochemicalethod. Lectin histochemistry with Wisteria floribunda agglutinin
WFA) which binds N-acetyl-d-galactosamine-bearing glycoconju-ates of e.g. chondroitin sulphate-containing proteoglycans [9,19]as applied as well.
. Materials and methods
Adult albino rats (Wistar) of either sex, of 250–300 g bodyeight, were used. All experimental procedures were performed
n accordance with European Communities Council Directive86/609/EEC) guidelines.
The animals were deeply anesthetized with ketamine andylazine injections (20 and 80 mg/kg, respectively, intramuscu-arly) and perfused transcardially with 4% paraformaldehyde in.1 M phosphate buffer (pH 7.4). Brains were postfixed in the samexative for 1 day at 4 ◦C. Serial sections (50 �m thickness) in theoronal plane were cut by a vibration microtome.
Floating sections were pretreated with 20% normal horse serumor 90 min. Sections were incubated with primary immunoreagentsmouse, monoclonal anti-neurocan and anti-versican, both diluted:100, DSHB, Iowa City, IA, investigators: R. K. Margolis and R.. Asher; goat, polyclonal anti-tenascin-R, 1:100, R&D Systems,inneapolis, MN; mouse, monoclonal, anti-brevican, 1:100, BD
iosciences, Sparks, MD; rabbit, polyclonal, anti-aggrecan, 1:500,erck Millipore, Billerica, MA in PBS containing 0.5% Triton X-
00, and 0.01% sodium azide) for 40 h, then processed according tohe avidin-biotinylated peroxidase method. For a detailed descrip-ion see e.g. Kálmán et al. [20]. In the case of the WFA-binding1:100, biotinylated, Vector Laboratories, Burlingame, CA) the 40 hncubation was followed immediately with the avidin-biotinylatederoxidase complex. Control sections were incubated by the omis-ion of the primary immunoreagents. No structure-bound labeling
as observed in these specimens.
Photomicrographs were taken by a DP50 digital cameraounted on an Olympus BX-51 microscope (both from Olympusptical Co. Ltd., Tokyo, Japan).
e Letters 566 (2014) 36–41 37
3. Results
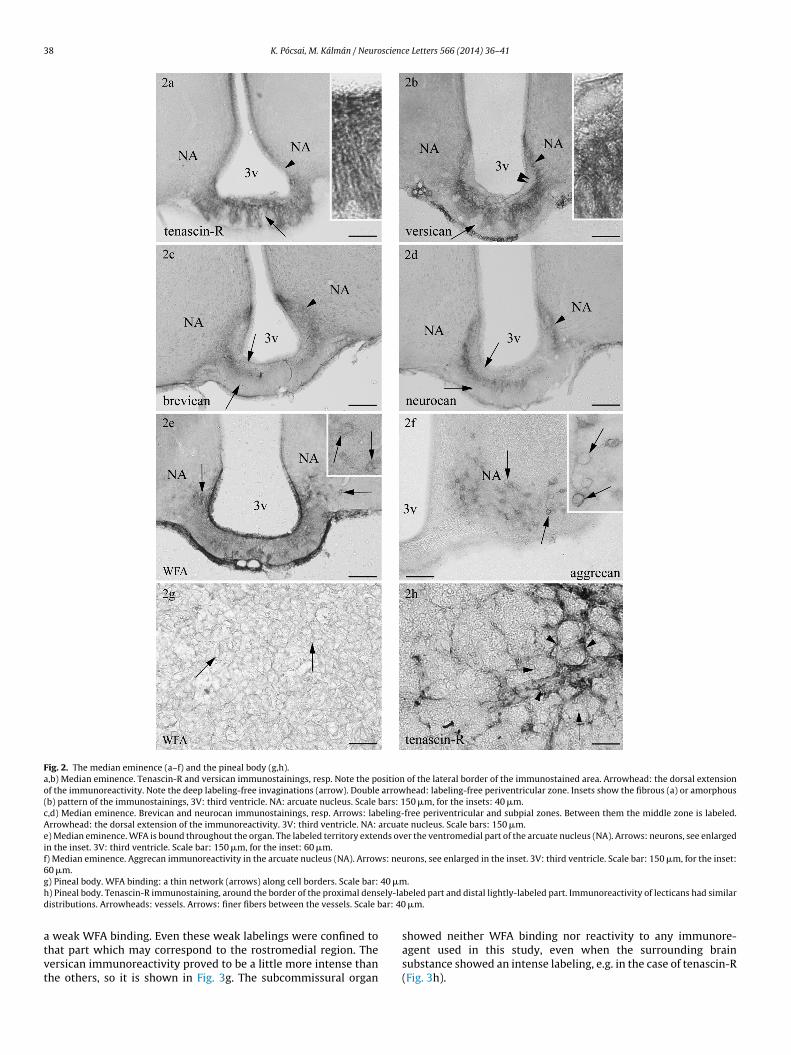
In the median eminence the territories marked by both tenascin-R and versican were very similar (Fig. 2a and b). The imunoreactivityextended from the ependyma to the pial surface where it inter-digitated with deep unstained invaginations. Lateralward theimmunolabeling had a border corresponding to the lateral cornersof the bottom of the ventricle but along the ventricle the reactiv-ity extended dorsally. In the posterior part of the organ this dorsalextension surrounded a narrow periventricular ‘demilune’ (Fig. 2b).The versican immunoreactivity was quite amorphous, whereas thatof tenascin-R had a fibrous pattern perpendicular to the pial surface(see insets).
On the other hand, the distribution of brevican and that of neu-rocan were also similar (Fig. 2c and d). They decorated a middlezone, leaving the periventricular and subpial zones unstained. Thestaining intensity gradually decreased medialward. Laterally theimmunoreactive zone diverged dorsal- and ventralward and closedlaterally the thin-walled bottom of the third ventricle.
The WFA was bound throughout the median eminence (Fig. 2e).The territory of the intense binding extended over the ventromedialpart of the arcuate nucleus at the border of the median eminence,unlike what was seen in the case of versican and tenascin-R. HereWFA binding delineated neurons, otherwise it was amorphous andcontinuous. In several other brain areas mainly in the cortex WFAbinding also visualized neurons. Immunoreactivity of aggrecan alsolabeled neurons in the arcuate nucleus (Fig. 2f) but it was not foundin CVOs.
In the pineal body two different patterns were found. In thecase of WFA, a network was formed as if it delineated the cellborders (Fig. 2g). The vessels were delineated by the immunoreac-tivity of both tenascin-R and versican and neurocan and brevican,but only in the rostral part of the organ. Between the vesselsthin immunopositive filaments were stretched. Of the similarimmunopatterns only that of tenascin-R is shown in figure here(Fig. 2h).
In the subfornical organ the immunoreactivity of versican(Fig. 3a) was intense arranged in a mesh-like pattern throughoutthe organ leaving only a small innermost area unstained. Since thetenascin-R immunostaining resulted in a similar pattern, it is notshown in figure. The neurocan immunoreactivity (Fig. 3b) extendedover the whole organ but it proved to be more intense along theborder of the hippocampal commissure and around the large lat-eral vessels on both sides, as if it formed a ‘shell’ of the organ. Asimilar distribution was found in the case of brevican, therefore itis not shown in figure. The WFA-binding (Fig. 3c) was confined toa ‘shell’ leaving the inner part unlabeled.
In the area postrema tenascin-R (Fig. 3d) had an intenseimmunoreactivity interrupted by ‘holes’. These ‘holes’ consisted ofa transparent ‘core’ and a rim of the very faint tone of unstained tis-sues. A dense labeling occurred at the ventrolateral border zone ofthe organ which seems to be in the position of the funiculus sepa-rans. The immunostaining of versican was less intense and less eventhan that of tenascin-R but delineated a sharp and narrow ventro-lateral border zone (not shown in figure). Brevican (Fig. 3e) and neu-rocan (not shown) had less intense immunoreactivity when com-pared to the previous ones. Actually, their immunoreactivity hardlysurpassed that of the surrounding brain substance but the territoryof the area postrema was marked by an intensely stained borderzone. Within the organ holes similar to those seen with tenascin-R were faintly recognizable. When WFA was applied the territoryof the area postrema displayed intense labeling (Fig. 3f). Ventro-
laterally on both sides there was a border zone which had evenmore intense immunoreactivity than the inner part of the organ.
Of the other organs the OVLT displayed weak immunoreactiv-ity to either versican, or brevican, or neurocan, or tenascin-R and
38 K. Pócsai, M. Kálmán / Neuroscience Letters 566 (2014) 36–41
Fig. 2. The median eminence (a–f) and the pineal body (g,h).a,b) Median eminence. Tenascin-R and versican immunostainings, resp. Note the position of the lateral border of the immunostained area. Arrowhead: the dorsal extensionof the immunoreactivity. Note the deep labeling-free invaginations (arrow). Double arrowhead: labeling-free periventricular zone. Insets show the fibrous (a) or amorphous(b) pattern of the immunostainings, 3V: third ventricle. NA: arcuate nucleus. Scale bars: 150 �m, for the insets: 40 �m.c,d) Median eminence. Brevican and neurocan immunostainings, resp. Arrows: labeling-free periventricular and subpial zones. Between them the middle zone is labeled.Arrowhead: the dorsal extension of the immunoreactivity. 3V: third ventricle. NA: arcuate nucleus. Scale bars: 150 �m.e) Median eminence. WFA is bound throughout the organ. The labeled territory extends over the ventromedial part of the arcuate nucleus (NA). Arrows: neurons, see enlargedin the inset. 3V: third ventricle. Scale bar: 150 �m, for the inset: 60 �m.f) Median eminence. Aggrecan immunoreactivity in the arcuate nucleus (NA). Arrows: neurons, see enlarged in the inset. 3V: third ventricle. Scale bar: 150 �m, for the inset:60 �m.g 40 �mh ely-lad ar: 40
atvt
) Pineal body. WFA binding: a thin network (arrows) along cell borders. Scale bar:) Pineal body. Tenascin-R immunostaining, around the border of the proximal densistributions. Arrowheads: vessels. Arrows: finer fibers between the vessels. Scale b
weak WFA binding. Even these weak labelings were confined tohat part which may correspond to the rostromedial region. Theersican immunoreactivity proved to be a little more intense thanhe others, so it is shown in Fig. 3g. The subcommissural organ
.beled part and distal lightly-labeled part. Immunoreactivity of lecticans had similar
�m.
showed neither WFA binding nor reactivity to any immunore-agent used in this study, even when the surrounding brainsubstance showed an intense labeling, e.g. in the case of tenascin-R(Fig. 3h).
K. Pócsai, M. Kálmán / Neuroscience Letters 566 (2014) 36–41 39
Fig. 3. The subfornical organ (a–c), the area postrema (d–f), the OVLT (g) and the subcommissural organ (h).a) Subfornical organ. The versican immunopositivity delineates a mesh-like pattern. The innermost part is unstained. CH: hippocampal commissure. Arrows: large vessels,c: ‘core’, s: ‘shell’, the border zone. Scale bar: 150 �m.b) Subfornical organ. The neurocan immunoreactivity is more intense in the ‘shell’. i.e. laterally, around the large vessels (arrows), and at the border zone (arrowheads). CH:hippocampal commissure. Scale bar: 150 �m.c) Subfornical organ. The WFA is bound at the ‘shell’ of the organ, i.e. in the border zone (arrowheads) to the hippocampal commissure (CH), and laterally around the largevessels (arrows). Scale bar: 150 �m.d) Area postrema. The tenascin-R displays an intense immunoreactivity with ‘holes’ (arrows). The structure of a hole is shown in the inset: a transparent core and a faint rim,double arrowheads point to the borderline between them. Arrowheads point to a border zone of more intense immunoreactivity which seems to be in the position of thefuniculus separans. ASP: area subpostrema. Scale bar: 150 �m, for the inset: 25 �m.e) Area postrema. The brevican immunoreactivity is less intense than the previous one. Arrows: holes. Arrowheads: the denser border zone. ASP: area subpostrema. CC:central canal. Scale bar: 150 �m.f) Area postrema, WFA binding. Arrowheads: the denser border zone. ASP: Area subpostrema. CC: central canal. Scale bar: 150 �m.g) OVLT, versican. Arrowheads point to the area of the organ. The immunostaining seems to be confined to the rostromedial part of the organ. TO: optic tract. Scale bar:150 �m.h) Subcommissural organ, tenascin-R immunostaining. The organ (arrowheads) is devoid of immunoreactivity. The surrounding brain substance (asterisks) displays labeling.CP: posterior commissure, V: third ventricle. Scale bar: 150 �m.
4 scienc
4
wtilfdonmwt
tst‘re
cb[nhibttt
itzlfba[
liswtps
mtmd
spfmotooeoh
0 K. Pócsai, M. Kálmán / Neuro
. Discussion
Summarizing the results, the extracellular matrix componentsere in general intensely expressed in the CVOs with two excep-
ions: the aggrecan immunoreactivity visualized only neuronsn the arcuate nucleus and the subcommissural organ was notabeled with either WFA, or lecticans, or tenascin-R. The dif-erent labelings usually overlapped each other. In general, theistribution of the extracellular matrix delineated the territoriesf the CVOs. It is to be emphasized that typical perineuronalets, a prevalent form of the arrangement of the extracellularatrix [3,21] were not recognized in the CVOs, although theyere delineated by WFA binding in several places, e.g. in the cor-
ex.In the median eminence the deep unstained invaginations seem
o correspond to spaces containing vessels entering the brain sub-tance here (see e.g. Fig. 5 of the paper cited as [22]). The ‘holes’ inhe area postrema must also be vascular spaces. Their transparentcores’ correspond to the lumina of the vessels, whereas the faintims represent the perivascular space as demonstrated by Shavert al. [23].
Tenascin-R is known to be a ligand of the chondroitin sulphate-ontaining proteoglycans [6]. There are also data on the WFAinding to the chondroitin sulphate-containing proteoglycans7,9]. The distribution patterns of the extracellular matrix compo-ents therefore are expected to be similar [6,9,24]. Discordancesowever, were also observed, e.g. WFA binding was found primar-
ly in the perineuronal net [7] as well as aggrecan [25], whereasrevican was distributed rather in the neuropil [7]. Major part ofhe extracellular matrix is produced by glial cells [8], so cerebral dis-ributions (neurocan: [26]; versican: [8]) were found very similaro that of GFAP.
Comparing the present data to our former studies on the GFAPn the subfornical organ [27,28], the GFAP occurred mainly aroundhe large vessels on the lateral side of the organ and in a borderone (forming a ‘shell’), i.e. on that part where WFA binding and,ess conspicuously, neurocan and brevican immunoreactivity alsoormed a ‘shell’. Note that two regions, a ‘shell’ and a ‘core’ haveeen described in the subfornical organ on the basis of severalspects (e.g. vascularization, nerve connections) by McKinley et al.16].
A ventrolateral border zone of the area postrema similar to thatabeled densely with extracellular matrix markers proved to bentensely immunopositive to GFAP in our former studies [27,29]. Iteems to correspond to the funiculus separans, a glial border zonehich separates the area postrema from the subpostremal area of
he nucleus of the solitary tract [16]. The subdivisions of the areaostrema (lateral, ventral, mantle and the central parts, see [16])howed no considerable difference.
In the median eminence GFAP immunoreactivity occurredainly in a middle zone, like brevican and neurocan immunoreac-
ivity in the present study. The fibrous pattern of tenascin-R in theedian eminence refers to a correlation with the tanycyte system
escribed here (see e.g. [30]).In the OVLT the immunostained area seems to be confined to the
o-called ‘rostromedial vascular region’ [16] reported to be com-osed largely of astrocytic processes. Former studies of our groupound GFAP immunoreactivity in the OVLT [27,28]. In the subcom-
issural organ GFAP did not occur [27,29,31]. The concentrationf the extracellular matrix components in the proximal part ofhe pineal body is also in accordance with the similar distributionf GFAP [27,32]. It is to be emphasized that the subcommissural
rgan is a so-called ‘ependymal’ organ [18], consisting mainly ofpendymocytes in contrast to the other CVOs. The negative resultsbtained in the case of this organ may be attributed to its specialistological structure.
e Letters 566 (2014) 36–41
In general, the extracellular matrix binds various factors (e.g.FGF, TGF-�1, [5,33,34]). It affects their local concentrations andbiological activity. Matrix-bound factors are inactivated slowly buttheir activity is mitigated [35]. The extracellular matrix of the CVOsmay have similar functions.
The extracellular matrix molecules have been suggested to playimportant roles in the formation and function of CNS synapses bye.g. directing the clustering of neurotransmitter receptors and iontransporters [36], or by entrapping the tissue fluid as a “perisynapticbarrier” [37].
Glial process arrangement is to be adapted to the actual require-ments, e.g. to uncover or cover neurohumoral endings releasinghumoral agents. This rearrangement may require an extracellularmatrix-rich environment like in the immature brain [38].
The extracellular matrix has a role in the maintenance of tissuehydration, in which aquaporins (in the CNS mainly aquaporin-4)also play an important role [4]. As it was published formerly [27],CVOs are intensely supplied with aquaporin-4 in a similar distri-bution as it was found for GFAP, including its avoidance in thesubcommissural organ. A common localisation for aquaporin-4 andheparansulphate proteoglycans has also been observed by Costaet al. [4] in the CNS neuropil.
Retarding the cation mobility, the extracellular matrix increasesthe osmotic pressure of the tissues [2] and controls their hydra-tion [1]. Tenascin-R contributes to the maintenance of the optimaldistance between cells [39]. The extracellular matrix may form‘channels’ for the diffusion so it may be facilitated in certain direc-tions. A significant decrease in extracellular space volume and anincrease in diffusion barriers follow the loss of extracellular matrix[39,40]. On the other hand Zamecnik et al. [41] pointed out that theextracellular matrix may form barriers to the diffusion.
Considering these we suppose that the extracellular matrix isessential in CVO functions providing the large extracellular spacewhich is required for the diffusion and other processes importantin their chemosensitive and neurosecretory activities. The decreaseof extracellular matrix beyond the border of the organs may con-tribute to the control of the diffusion of molecules from the CVOsinto the surrounding brain substance.
Acknowledgements
The authors are grateful to Professor Klára Matesz for the gen-erous gift of primary immunoreagents used in the preliminarystudies. The study was supported by the PhD School of SemmelweisUniversity, Budapest, Hungary.
References
[1] M. Aumailley, B. Gayraud, Structure and biological activity of the extracellularmatrix, J. Mol. Med. (Berl.) 76 (1998) 253–265.
[2] S. Carbonetto, The extracellular matrix of the nervous system, Trends Neurosci.10 (1984) 382–387.
[3] M.R. Celio, I. Blumcke, Perineuronal nets–a specialized form of extracellu-lar matrix in the adult nervous system, Brain Res. Brain Res. Rev. 19 (1994)128–145.
[4] C. Costa, R. Tortosa, A. Domènech, E. Vidal, M. Pumarola, A. Bassols, Mappingof aggrecan, hyaluronic acid, heparan sulphate proteoglycans and aquaporin4 in the central nervous system of the mouse, J. Chem. Neuroanat. 33 (2007)111–123.
[5] K.E. Dow, W. Wang, Cell biology of astrocyte proteoglycans, Cell Mol. Life Sci.54 (1998) 567–581.
[6] Y. Yamaguchi, Lecticans: organizers of the brain extracellular matrix, Cell Mol.Life Sci. 57 (2000) 276–289.
[7] J.M. Ajmo, A.K. Eakin, M.G. Hamel, P.E. Gottschall, Discordant localization ofWFA reactivity and brevican/ADAMTS-derived fragment in rodent brain, BMCNeurosci. 9 (2008) 14.
[8] A. Bignami, M. Hosley, D. Dahl, Hyaluronic acid and hyaluronic acid-bindingproteins in brain extracellular matrix, Anat. Embryol. (Berl.) 188 (1993)419–433.
[9] G. Brückner, J. Grosche, M. Hartlage-Rübsamen, S. Schmidt, M. Schachner,Region and lamina-specific distribution of extracellular matrix proteoglycans,
hyaluronan and tenascin-R in the mouse hippocampal formation, J. Chem. Neu-roanat. 26 (2003) 37–50.
10] G. Gáti, M. Morawski, D. Lendvai, C. Jäger, L. Négyessy, T. Arendt, A. Alpár,Distribution and classification of aggrecan-based extracellular matrix in thethalamus of the rat, J. Neurosci. Res. 88 (2010) 3257–3266.
11] D. Lendvai, M. Morawski, L. Négyessy, G. Gáti, C. Jäger, G. Baksa, T. Glasz, J.Attems, H. Tanila, T. Arendt, T. Harkany, A. Alpár, Neurochemical mapping of thehuman hippocampus reveals perisynaptic matrix around functional synapsesin Alzheimerés disease, Acta Neuropathol. 125 (2013) 215–229.
13] Y. Dong, X. Han, Y. Xue, B. Dong, X. Guo, G. Hu, C. Zhu, Y. Lu, Secreted brevicanmRNA is expressed in the adult rat pituitary, Biochem. Biophys. Res. Commun.314 (2004) 745–748.
14] K. Pócsai, Z. Bagyura, M. Kálmán, Components of the basal lamina and dys-trophin dystroglycan complex in the neurointermediate lobe of rat pituitarygland: different localizations of beta-dystroglycan, dystrobrevins, alpha1-syntrophin, and aquaporin-4, J. Histochem. Cytochem. 58 (2010) 463–479.
15] E.E. Benarroch, Circumventricular organs: receptive and homeostatic functionsand clinical implications, Neurology 77 (2011) 1198–1204.
16] M.J. McKinley, R.M. McAllen, P. Davern, M.E. Giles, J. Penschow, N. Sunn, A.Uschakov, B.J. Oldfield, The sensory circumventricular organs of the mam-malian brain, Adv. Anat. Embryol. Cell Biol. 172 (2003) 1–122.
17] S. Sisó, M. Jeffrey, L. González, Sensory circumventricular organs in health anddisease, Acta Neuropathol. 120 (2010) 689–705.
18] B. Vígh, M.J. Manzano e Silva, C.L. Frank, C. David, S.J. Czirok, C. Vincze, G. Racz,A. Lukats, A. Szel, The circumventricular organs of the brain: do they representa cerebrospinal fluid-dependent regulatory system? Med. Hypotheses Res. 1(2004) 77–100.
19] V. Piller, F. Piller, J.P. Cartron, Comparison of the carbohydrate-binding speci-ficities of seven N-acetyl-d-galactosamine-recognizing lectins, Eur. J. Biochem.191 (1990) 461–466.
20] M. Kálmán, H. Somiya, L. Lazarevic, I. Milosevic, C. Ari, K. Majorossy, Absenceof post-lesion reactive gliosis in elasmobranchs and turtles and its bearing onthe evolution of astroglia, J. Exp. Zool. B, Mol. Dev. Evol. 320 (2013) 351–367.
21] M.R. Celio, R. Spreafico, S. De Biasi, L. Vitellaro-Zuccarello, Perineuronal nets:past and present, Trends Neurosci. 21 (1998) 510–515.
22] G. Ambach, M. Palkovits, J. Szentágothai, Blood supply of the rat hypothalamus.IV. Retrochiasmatic area, median eminence, arcuate nucleus, Acta Morphol.Acad. Sci. Hung. 24 (1976) 93–119.
raphy of capillaries in the dorsal vagal complex of rats: I. Morphometricproperties, J. Comp. Neurol. 306 (1991) 73–82.
24] G. Perides, H.P. Erickson, F. Rahemtulla, A. Bignami, Colocalization of tenascinwith versican, a hyaluronate-binding chondroitin sulfate proteoglycan, Anat.Embryol. (Berl.) 188 (1993) 467–479.
[
[
e Letters 566 (2014) 36–41 41
25] M. Morawski, G. Brückner, T. Arendt, R.T. Matthews, Aggrecan: beyond cartilageand into the brain, Int. J. Biochem. Cell. Biol. 44 (2012) 690–693.
26] R.J. McKeon, M.J. Jurynec, C.R. Buck, The chondroitin sulfate proteoglycans neu-rocan and phosphacan are expressed by reactive astrocytes in the chronic CNSglial scar, J. Neurosci. 19 (1999) 10778–107788.
27] O. Goren, I. Adorján, M. Kálmán, Heterogeneous occurrence of aquaporin-4 inthe ependyma and in the circumventricular organs in rat and chicken, Anat.Embryol. (Berl.) 211 (2006) 155–172.
28] M. Kálmán, F. Hajós, Distribution of glial fibrillary acidic protein (GFAP)-immunoreactive astrocytes in the rat brain. I. Forebrain, Exp. Brain Res. 78(1989) 147–163.
29] F. Hajós, M. Kálmán, Distribution of glial fibrillary acidic protein (GFAP)-immunoreactive astrocytes in the rat brain. II. Mesencephalon, rhomben-cephalon and spinal cord, Exp. Brain Res. 78 (1989) 164–173.
30] E.M. Rodríguez, J.L. Blázquez, F.E. Pastor, B. Peláez, P. Pena, B. Peruzzo, P. Amat,Hypothalamic tanycytes: a key component of brain-endocrine interaction, Int.Rev. Cytol. 247 (2005) 89–164.
31] P. Redecker, Immunohistochemical localisation of glial fibrillary acidic protein(GFAP) and vimentin in the subcommissural organ of the Mongolian gerbil(Meriones unguiculatus), Cell Tissue Res. 255 (1989) 595–600.
32] Z. Bagyura, K. Pócsai, M. Kálmán, Distribution of components of basal laminaand dystrophin-dystroglycan complex in the rat pineal gland: differences fromthe brain tissue and between the subdivisions of the gland, Histol. Histopathol.25 (2010) 1–14.
33] P. Bovolenta, I. Fernaud-Espinosa, Nervous system proteoglycans as modulatorsof neurite outgrowth, Prog. Neurobiol. 61 (2000) 113–132.
34] N. Maeda, Structural variation of chondroitin sulfate and its roles in the centralnervous system, Cent. Nerv. Syst. Agents Med. Chem. 10 (2010) 22–31.
35] J.C. Adams, F.M. Watt, Regulation of development and differentiation by theextracellular matrix, Development 117 (1993) 1183–1198.
36] A. Dityatev, C.I. Seidenbecher, M. Schachner, Compartmentalization from theoutside: the extracellular matrix and functional microdomains in the brain,Trends Neurosci. 33 (2010) 503–512.
37] T. Murakami, A. Ohtsuka, Perisynaptic barrier of proteoglycans in the maturebrain and spinal cord, Arch. Histol. Cytol. 66 (2003) 195–207.
38] V. Prevot, B. Dehouck, P. Poulain, J.C. Beauvillain, V. Buée-Scherrer, S. Bouret,Neuronal-glial-endothelial interactions and cell plasticity in the postnatalhypothalamus: implications for the neuroendocrine control of reproduction,Psychoneuroendocrinology 32 (Suppl. 1) (2007) S46–S51.
39] E. Syková, I. Vorísek, T. Mazel, T. Antonova, M. Schachner, Reduced extracellularspace in the brain of tenascin-R- and HNK-1-sulphotransferase deficient mice,Eur. J. Neurosci. 22 (2005) 1873–1880.
40] L. Vargová, E. Syková, Extracellular space diffusion and extrasynaptic transmis-sion, Physiol. Res. 57 (Suppl. 3) (2008) S89–S99.
41] J. Zamecnik, A. Homola, M. Cicanic, K. Kuncova, P. Marusic, P. Krsek, E. Sykova,L. Vargova, The extracellular matrix and diffusion barriers in focal cortical dys-plasias, Eur. J. Neurosci. 36 (2012) 2017–2024.