EZH2-shRNA–mediated upregulation of p21 waf1/cip1 and its transcriptional enhancers with concomitant downmodulation of mutant p53 in pancreatic ductal adenocarcinoma Ramesh B. Batchu, PhD, a,b Aamer M. Qazi, PhD, a,b Oksana V. Gruzdyn, a,b Assaad Semaan, MD, a,c Shelly M. Seward, MD, a,c Sreedhar Chamala, MD, a,c Vasu B. Dhulipala, MD, a,b David L. Bouwman, MD, a Donald W. Weaver, MD, a and Scott A. Gruber, MD, PhD, MBA, a,b Detroit, MI Purpose. Enhancer of zeste homologue 2 (EZH2), a component of the chromatin modification protein complex, is upregulated in pancreatic ductal adenocarcinoma (PDAC), whereas loss of p53 and its downstream target, p21 waf1/cip1 , is also observed frequently. We sought to investigate the role of the p53-p21 waf1/cip1 pathway in relation to EZH2-mediated inhibition of PDAC. Methods. The PANC-1 cell line was utilized in chromatin immunoprecipitation, gene profiling, Western blot, cell invasion, cell proliferation, and tumor xenograft assays. Results. Western blot analysis with antibodies that recognize both wild-type and mutant p53 did not show any alterations in band intensity; however, antibody that detects only mutant p53 showed a band of significantly lesser intensity with EZH2 knockdown. Western blot analysis further revealed a significant upregulation of p21 waf1/cip1 . Gene expression profile analysis indicated significantly enhanced transcripts of transcriptional inducers of p21 waf1/cip1 , with downregulation of mutant p53 transcript, corroborating the Western blot analysis. PANC-1 cells expressing EZH2-short hairpin RNA displayed markedly attenuated growth in SCID mice. Conclusion. Downregulation of mutant p53 with concomitant enhanced expression of p21 waf1/cip1 and its transcriptional trans-activators may contribute toward EZH2-mediated suppression of PDAC. (Surgery 2013;154:739-47.) From the Laboratory of Surgical Oncology & Developmental Therapeutics, a Department of Surgery, Wayne State University, the John D. Dingell VA Medical Center, b and the Department of Obstetrics and Gynecology, c Wayne State University, Detroit, MI ENHANCER OF ZESTE HOMOLOGUE 2 (EZH2), a methyl- transferase that trimethylates histone H3 at lysine residue 27 (H3K27me3), results in dense packing of chromatin at the site of tumor suppressor genes, leading to their silencing. 1 EZH2 has been associ- ated with aggressive forms of prostate, breast, hepatocellular, and gastric cancers, as well as mela- noma and transitional cell carcinoma of the bladder, making it a robust, solid tumor marker. 2 RNA interference (RNAi)–mediated suppression of EZH2 expression suppresses tumor growth and angiogenesis in multiple cancer types. 3 Recent studies indicate a direct role for EZH2 in the progression of pancreatic cancer. 4,5 Indeed, high levels of EZH2 have been detected in biopsies of pancreatic cancer tissue 6,7 and stem cells. 5 Further, its expression has been correlated with poor prognosis and its depletion resulted in inhibi- tion of cell proliferation. 8 We reported recently an inverse relationship between the translational regulator microRNA-101 (miR-101) and EZH2 in pancreatic cancer, 9 which has been corroborated subsequently by others. 10 Depletion of EZH2 in pancreatic cancer increases expression of the G1 phase checkpoint cell-cycle inhibitors, tumor sup- pressor gene p27 kip1 (cyclin-dependent kinase [CDK]N1B), 6 and p57 kip2 (CDKN1C). 11 While studying the role of miR-101 in ovarian cancer, we observed an inverse relationship between Accepted for publication June 25, 2013. Reprint requests: Ramesh B. Batchu, PhD, Assistant Professor, Department of Surgery, Wayne State University, John D. Dingell VA Medical Center, Lab 4242, 4646 John R Street, Detroit, MI 48201. E-mail: [email protected]. 0039-6060/$ - see front matter Ó 2013 Mosby, Inc. All rights reserved. http://dx.doi.org/10.1016/j.surg.2013.06.041 SURGERY 739
Transcript
Accepte
ReprintDepartmVA MedMI 4820
0039-60
� 2013
http://d
EZH2-shRNA–mediated upregulationof p21waf1/cip1 and its transcriptionalenhancers with concomitantdownmodulation of mutant p53 inpancreatic ductal adenocarcinomaRamesh B. Batchu, PhD,a,b Aamer M. Qazi, PhD,a,b Oksana V. Gruzdyn,a,b Assaad Semaan, MD,a,c
Shelly M. Seward, MD,a,c Sreedhar Chamala, MD,a,c Vasu B. Dhulipala, MD,a,b
David L. Bouwman, MD,a Donald W. Weaver, MD,a and Scott A. Gruber, MD, PhD, MBA,a,b Detroit, MI
Purpose. Enhancer of zeste homologue 2 (EZH2), a component of the chromatin modification proteincomplex, is upregulated in pancreatic ductal adenocarcinoma (PDAC), whereas loss of p53 and itsdownstream target, p21waf1/cip1, is also observed frequently. We sought to investigate the role of thep53-p21waf1/cip1 pathway in relation to EZH2-mediated inhibition of PDAC.Methods. The PANC-1 cell line was utilized in chromatin immunoprecipitation, gene profiling, Westernblot, cell invasion, cell proliferation, and tumor xenograft assays.Results. Western blot analysis with antibodies that recognize both wild-type and mutant p53 did notshow any alterations in band intensity; however, antibody that detects only mutant p53 showed a bandof significantly lesser intensity with EZH2 knockdown. Western blot analysis further revealed asignificant upregulation of p21waf1/cip1. Gene expression profile analysis indicated significantlyenhanced transcripts of transcriptional inducers of p21waf1/cip1, with downregulation of mutant p53transcript, corroborating the Western blot analysis. PANC-1 cells expressing EZH2-short hairpin RNAdisplayed markedly attenuated growth in SCID mice.Conclusion. Downregulation of mutant p53 with concomitant enhanced expression of p21waf1/cip1 andits transcriptional trans-activators may contribute toward EZH2-mediated suppression of PDAC.(Surgery 2013;154:739-47.)
From the Laboratory of Surgical Oncology & Developmental Therapeutics,a Department of Surgery, WayneState University, the John D. Dingell VA Medical Center,b and the Department of Obstetrics and Gynecology,c
Wayne State University, Detroit, MI
ENHANCER OF ZESTE HOMOLOGUE 2 (EZH2), a methyl-transferase that trimethylates histone H3 at lysineresidue 27 (H3K27me3), results in dense packingof chromatin at the site of tumor suppressor genes,leading to their silencing.1 EZH2 has been associ-ated with aggressive forms of prostate, breast,hepatocellular, and gastric cancers, as well as mela-noma and transitional cell carcinoma of thebladder, making it a robust, solid tumor marker.2
RNA interference (RNAi)–mediated suppression
d for publication June 25, 2013.
requests: Ramesh B. Batchu, PhD, Assistant Professor,ent of Surgery, Wayne State University, John D. Dingellical Center, Lab 4242, 4646 John R Street, Detroit,1. E-mail: [email protected].
60/$ - see front matter
Mosby, Inc. All rights reserved.
x.doi.org/10.1016/j.surg.2013.06.041
of EZH2 expression suppresses tumor growthand angiogenesis in multiple cancer types.3
Recent studies indicate a direct role for EZH2 inthe progression of pancreatic cancer.4,5 Indeed,high levels of EZH2 have been detected in biopsiesof pancreatic cancer tissue6,7 and stem cells.5
Further, its expression has been correlated withpoor prognosis and its depletion resulted in inhibi-tion of cell proliferation.8 We reported recently aninverse relationship between the translationalregulator microRNA-101 (miR-101) and EZH2 inpancreatic cancer,9 which has been corroboratedsubsequently by others.10 Depletion of EZH2 inpancreatic cancer increases expression of the G1phase checkpoint cell-cycle inhibitors, tumor sup-pressor gene p27kip1 (cyclin-dependent kinase[CDK]N1B),6 and p57kip2 (CDKN1C).11 Whilestudying the role of miR-101 in ovarian cancer,we observed an inverse relationship between
Fig 1. Cell growth, colony formation, wound healing, and invasion assays. A, Cell growth assays performed after trans-fection with EZH2-shRNA or control vector. B, Colony formation assays were performed after EZH2-shRNA transfectionand incubation for 2 weeks in 6-well plates. C, Wound healing assay was performed by making a uniform scratch fol-lowed by the measurement of cell migration in both control and EZH2-shRNA–transfected PANC-1 cells. The closureof the ‘‘wound’’ was monitored with phase-contrast microscopy and representative pictures taken at 0 and 20 hours aredepicted. D, EZH2-shRNA– and control vector–transfected cells were trypsinized and seeded in Boyden transwell cham-bers. Migrated cells at the lower surface of the transwell filter were stained with 5 mmol/L calcein and counted in 3random fields under a fluorescent microscope. (Color version of figure is available online.)
SurgeryOctober 2013
740 Batchu et al
p21waf1/cip1 and EZH2,12 the key downstreamtarget of p53.13 Loss of both p21waf1/cip1
(CDKN1A) and p53 is also observed frequently inpancreatic cancers.14 Therefore, the purpose ofthis work was to examine the effects of specific in-hibition of EZH2 on the p53-p21waf1/cip1 pathway,which may offer new opportunities to furtherunravel and target therapeutically the intracellularsignaling mechanisms in pancreatic ductal adeno-carcinoma (PDAC).
MATERIALS AND METHODS
Antibodies and reagents. Poly (ADP-ribose)polymerase (PARP), CDK 4, bcl-2, bak, H3k27me3,p21waf1/cip1, and b-actin antibodies were purchasedfrom Santa Cruz Biotechnology (Santa Cruz, CA).
Antibodies to 3 different epitopes of p53 (Ab2recognizing amino acids 36–45, Ab6 recognizingamino acids 21–25, and Ab3 recognizing aminoacids 213–217) were obtained from OncogeneResearch Products (Cambridge, MA). The first 2detect both wild-type and mutant p53; the third de-tects only mutant p53 under nondenaturing condi-tions. Control shRNA and EZH2-shRNA pLKO.1vectors were obtained from Sigma-Aldrich (St.Louis, MO). The control vector sequence is: 59CCG GCA ACA AGA TGA AGA GCA CCA ACTCGA GTT GGT GCT CTT CAT CTT GTT GTTTTT 39. The EZH2-shRNA vector sequence is: 59CCG GCC CAA CAT AGA TGG ACC AAA TCTCGA GAT TTG GTC CAT CTA TGT TGG GTTTTT G 39.
Fig 2. Flow cytometric analysis of apoptosis and its cellular markers after EZH2-shRNA transfection of PANC-1 cells. A,Control and EZH2-shRNA–transfected PANC-1 cells (1 3 105) were fixed, stained both with Annexin V fluorescein iso-thiocyanate and propidium iodide (20 mg/mL), and processed for flow cytometric analysis. i, Pre-apoptotic cells. ii, Post-apoptotic cells. B, Time-dependent changes in the expression levels of the apoptotic molecular markers PARP, bak, andbcl-2. b-actin levels were used to calibrate the loading of the samples. RI, Relative intensity (calculated using ImageJ soft-ware, a Java-based image processing program obtained from a public domain developed at the National Institutes ofHealth).
SurgeryVolume 154, Number 4
Batchu et al 741
Cell culture. The human pancreatic tumor celllines PANC-1, BxPc-3, and MIA PaCa-2 wereobtained from American Type Culture Collection(Manassas, VA) and grown in subconfluent mono-layer cultures as described previously.9 Cells werecultured in a humidified atmosphere with 5%CO2 at 378C. Standard prototype growth curvesand number of viable cells were determined foreach cell line (treated and control groups) in trip-licate according to the CCK-8 Kit (Dojindo, Gai-thersburg, MD) manufacturer’s instructions.
Colony formation assays. These assays wereperformed as described previously.9,12 In brief,control and transfected cells were incubated for14 days. Cells were then washed and fixed in abso-lute methanol for 15 minutes at �208C and stainedwith 0.1% crystal violet in 25% methanol. Colonieswere counted using the ImageJ program andphotographed.
Wound healing assay. EZH2-shRNA– and con-trol vector–transfected monolayer cells were
disrupted mechanically with a pipette tip togenerate a wound ridge free of cells. Cell migra-tion was measured by the ability to close theartificially created gap and was documented undera light microscope.
Cell invasion assay. Assays were performed withEZH2-shRNA– and control vector–transfectedPANC-1 cells using a modified Boyden chambermethod with a pore size of 8 mm according to ourearlier protocol.9
Apoptosis assay. EZH2-shRNA–transfectedPANC-1 cells were serum starved for 24 hoursfollowed by incubation with complete growth me-dium. After another 48 hours, cells were assessedfor cell-cycle phase. Apoptotic cells were analyzedusing the Annexin V FITC apoptosis detection kit(Calbiochem, Gibbstown, NJ) according to themanufacturer’s instructions. The DNA content ofthe cells was analyzed by flow cytometry and thesub G1 population was considered to representapoptotic cells. Propidium iodide was added to
Fig 3. Chromatin immunoprecipitation (ChIP) assay showingmodulation of the interaction between p21waf1/cip1 (cyclin-dependent kinase [CDK]N1A) and EZH2 and effects of EZH2 knockdown on global methylation of H3k27. A, i, ChIPassay using anti-EZH2 antibody and p21waf1/cip1 primers performed with vector-transfected cells (control, lane 1) andEZH2-shRNA–transfected cells (lane 2) showing interaction of EZH2 with p21waf1/cip1 promoter region. A 255-bp bandis shown targeting the –93 region of p21waf1/cip1 promoter after performing standard polymerase chain reaction. Theinput (b-actin) lanes indicate equal loading of the ChIP samples. A, ii, Analysis of the expression of p21waf1/cip1 byWesternblot for control and EZH2-shRNA–treated samples at 48 hours. RI, Relative intensity. B, Western blot analysis for theexpression of H3k27me3 at indicated time intervals after transfection with EZH2-shRNA in PANC-1 cells.
SurgeryOctober 2013
742 Batchu et al
discriminate early apoptotic from late apoptotic ornecrotic cells.
DNA microarray analysis. This was performedusing the Human Whole Genome OneArray v5(Phalanx Biotech, Palo Alto, CA) according to ourearlier protocol.12
Western blotting. Proteins were separated using4–15% SDS Tris-Glycine gels and transfer ofproteins to nitrocellulose membranes was conduct-ed with the iBlot dry blotting device (InVitrogenCorp, Carlsbad, CA) as previously described.9
Chromatin immunoprecipitation assay. Chro-matin immunoprecipitation (ChIP) was per-formed using the ChIP Assay kit (EZ-Magna ChIPA, Upstate, Temecula, CA) according to manufac-turer’s instructions. Briefly, 1 3 107 control andEZH2-shRNA–transfected PANC-1 cells weregrown for 48 hours and protein:nucleic acid com-plexes were cross-linked with 1% formaldehydefor 10 minutes at 258C. Cells were harvested, lysed,and sonicated (83 for 15 seconds with 50-secondcooling). Soluble chromatin was immunoprecipi-tated with anti-EZH2 antibody (Millipore, Billerica,MA). DNA:protein immune complexes wereeluted, reverse cross-linked, and DNA wasextracted using a spin filter column. Standardendpoint polymerase chain reaction was per-formed using JumpStart REDTaq polymerase
chain reaction mix (Sigma). The p21waf1/cip1 pro-moter domain in immunoprecipitated DNA wasidentified using the forward primer, 59-GGT GTCTAG GTG CTC CAG GT-39, and reverse primer,59-GCA CTC TCC AGG AGG ACA CA-39, withpreviously described reaction conditions.12
Mice xenograft studies. Six-week-old CB17/Cr-SCID mice were obtained from Taconic Farms(Germantown, NY) and maintained underpathogen-free conditions. PANC-1 cells (4 3 106)transfected with either with EZH2-shRNA or scram-bled shRNA controls were suspended into 100 mLphosphate-buffered saline. Cells were injected sub-cutaneously into both flanks of mice. Tumor diame-ters were measured every week from 1 to 3 monthspostinjection using digital calipers. Tumor volumein cubic millimeters was calculated using the for-mula, (L3W 2)/2, where L is the maximum lengthand W is the maximum width. All animal experi-ments in this study were conducted in compliancewith Institutional Animal Care and Use Commit-tee/Wayne State University guidelines.
Statistical analysis. Data were analyzed using theStudent’s t-test.
RESULTS
Silencing of EZH2 inhibits cell proliferation,colony formation, cell migration, and invasion.
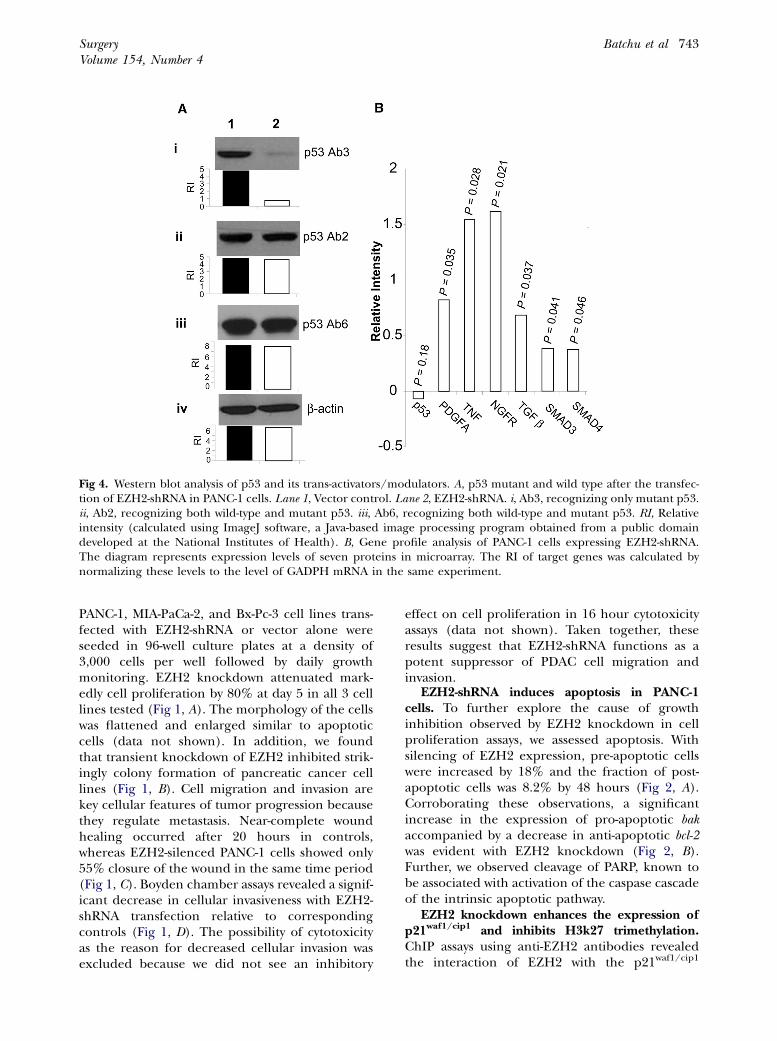
Fig 4. Western blot analysis of p53 and its trans-activators/modulators. A, p53 mutant and wild type after the transfec-tion of EZH2-shRNA in PANC-1 cells. Lane 1, Vector control. Lane 2, EZH2-shRNA. i, Ab3, recognizing only mutant p53.ii, Ab2, recognizing both wild-type and mutant p53. iii, Ab6, recognizing both wild-type and mutant p53. RI, Relativeintensity (calculated using ImageJ software, a Java-based image processing program obtained from a public domaindeveloped at the National Institutes of Health). B, Gene profile analysis of PANC-1 cells expressing EZH2-shRNA.The diagram represents expression levels of seven proteins in microarray. The RI of target genes was calculated bynormalizing these levels to the level of GADPH mRNA in the same experiment.
SurgeryVolume 154, Number 4
Batchu et al 743
PANC-1, MIA-PaCa-2, and Bx-Pc-3 cell lines trans-fected with EZH2-shRNA or vector alone wereseeded in 96-well culture plates at a density of3,000 cells per well followed by daily growthmonitoring. EZH2 knockdown attenuated mark-edly cell proliferation by 80% at day 5 in all 3 celllines tested (Fig 1, A). The morphology of the cellswas flattened and enlarged similar to apoptoticcells (data not shown). In addition, we foundthat transient knockdown of EZH2 inhibited strik-ingly colony formation of pancreatic cancer celllines (Fig 1, B). Cell migration and invasion arekey cellular features of tumor progression becausethey regulate metastasis. Near-complete woundhealing occurred after 20 hours in controls,whereas EZH2-silenced PANC-1 cells showed only55% closure of the wound in the same time period(Fig 1, C). Boyden chamber assays revealed a signif-icant decrease in cellular invasiveness with EZH2-shRNA transfection relative to correspondingcontrols (Fig 1, D). The possibility of cytotoxicityas the reason for decreased cellular invasion wasexcluded because we did not see an inhibitory
effect on cell proliferation in 16 hour cytotoxicityassays (data not shown). Taken together, theseresults suggest that EZH2-shRNA functions as apotent suppressor of PDAC cell migration andinvasion.
EZH2-shRNA induces apoptosis in PANC-1cells. To further explore the cause of growthinhibition observed by EZH2 knockdown in cellproliferation assays, we assessed apoptosis. Withsilencing of EZH2 expression, pre-apoptotic cellswere increased by 18% and the fraction of post-apoptotic cells was 8.2% by 48 hours (Fig 2, A).Corroborating these observations, a significantincrease in the expression of pro-apoptotic bakaccompanied by a decrease in anti-apoptotic bcl-2was evident with EZH2 knockdown (Fig 2, B).Further, we observed cleavage of PARP, known tobe associated with activation of the caspase cascadeof the intrinsic apoptotic pathway.
EZH2 knockdown enhances the expression ofp21waf1/cip1 and inhibits H3k27 trimethylation.ChIP assays using anti-EZH2 antibodies revealedthe interaction of EZH2 with the p21waf1/cip1
Fig 5. Effects of EZH2-shRNA transfection on in vivo growth of PANC-1 cells. A, Xenograft tumor growth of PANC-1cells expressing EZH2-shRNA in CB17/Cr-SCID mice. Top panel, In contrast with control vector-transfected cells,EZH2-shRNA–transfected cells have decreased ability to form tumors in vivo. Bottom panel, Resected xenograft tumorsof control (left) and EZH2-shRNA–transfected (right) PANC-1 cells. B, Box plot depiction of tumor volume. Plot repre-sents mean tumor volume, where the horizontal line in the box represents the median, the upper and lower lines of thebox the 75th and 25th percentiles, respectively, and the uppermost and lowermost lines the extreme values. (Colorversion of figure is available online.)
SurgeryOctober 2013
744 Batchu et al
promoter. We observed that this interaction wasdecreased by half in cells transfected with EZH2-shRNA (Fig 3, Ai). Western blot analysis confirmedenhanced expression of p21waf1/cip1 protein inresponse to EZH2 knockdown (Fig 3, Aii). Tran-scriptional repression mediated by EZH2 involvesconvergence of both histone deacetylation andmethylation.15 Corroborating ChIP assay results,Western blot analysis showed a global downregula-tion ofH3k27me3with EZH2 knockdown (Fig 3,B).
Downmodulation of mutant p53 with concomi-tant increased expression of p21waf1/cip1 transcrip-tional activators by EZH2-shRNA. Western blotanalysis with mutant-specific p53 antibody (Fig 4,Ai) showed a band of lesser intensity with EZH2knockdown compared with p53 antibody that rec-ognizes both wild type and mutants (Fig 4, Aii,Aiii). Further, gene expression profiling showeddownregulation of mutant p53 with concomitantupregulation of the expression of transcriptionalinducers of p21waf1/cip1, such as platelet-derivedgrowth factor A, tumor necrosis factor, nervegrowth factor receptor, transforming growth fac-tor-b, and SMAD proteins 3 and 4 (Fig 4, B).
Inhibition of in vivo tumor growth by EZH2-shRNA. To understand the in vivo effect of EZH2-shRNA, tumor xenograft studies were conductedusing PANC-1 cells. Control vector– and EZH2-shRNA–transfected PANC-1 cells were injectedsubcutaneously into the left and right posteriorflanks of CB17/Cr-SCID mice, respectively. Tumorxenografts formed by cells transfected with EZH2-
shRNA were significantly smaller than controltumors throughout the study period of 60 days(Fig 5, A). The mean total tumor volume was1,565 ± 18 mm3 in the control group comparedwith 590 ± 8 mm3 in the EZH2-shRNA–transfectedgroup (P < .05; Fig 5, B).
DISCUSSION
A growing body of recent work has correlatedupregulation of EZH2 with the progression ofpancreatic and many other cancers, but themolecular mechanism behind this phenomenonremains largely unknown. In this report, we haveshown that knockdown of EZH2 using RNAiexhibits anti-tumor activity against PDAC cellsboth in vitro by inhibiting cell proliferation, colonyformation, and cell migration and invasion, andin vivo by attenuating tumor growth at least inpart by activation of the p53-p21waf1/cip1 signalingpathway. Activation of pro-apoptotic bak and inhi-bition of anti-apoptotic bcl-2 indicate that theapoptosis observed due to EZH2 knockdown isvia the intrinsic pathway.
ChIP analyses revealed that knockdown of EZH2results in chromatin relaxation via downmodulationof H3k27 methylation, leading to reexpression andhigh levels of the tumor suppressor gene p21waf1/cip1. Given that p21waf1/cip1 is a downstream targetof p5313 and p53 is inactivated in the majority ofpancreatic tumors,14 we hypothesized that modula-tion of the p53–p21 pathway is important in PDACgrowth inhibition. EZH2-shRNA–mediated
SurgeryVolume 154, Number 4
Batchu et al 745
downmodulationofmutant p53, whichnormally en-hances metastatic potential and tissue invasive-ness,16 lends credence to this hypothesis. In fact,EZH2-mediated repression has been shown forp27kip1 (CDK1B)6 and p57kip2 (CDKN1C)11 tumorsuppressor genes. Whether EZH2 simultaneouslyacts at multiple levels to inhibit p21waf1/cip1 andenhance mutant p53, or whether a commonsignaling mechanism brings about modulation ofboth of these key check points of cell proliferation,is unclear and needs further investigation. Hence,identification of an ideal RNAi target that works atmultiple levels to treat PDAC may be beneficial.Because EZH2-shRNA promotes reexpression of anumber of tumor suppressor genes along with sup-pression of mutant p53, it may prove to be an effec-tive therapeutic intervention in PDAC. In thisregard, our observation that EZH2-shRNA–trans-fected cells displayed markedly attenuated growthpotential in SCID mice lends further support tothis idea.
By virtue of its trimethylation of histone H3k27,EZH2 is responsible in part for control of tran-scriptional memory and maintains long-termsilencing of specific sets of genes resulting inchromatin compaction.1 In transitional cell carci-noma of the bladder, EZH2 knockdown was shownto repress H3K27me3.17 Corroborating this obser-vation, we found a global decrease in the levelsof H3K27 in response to EZH2 knockdown inPDAC. In fact, epigenetic silencing by histoneH3K27 methylation has been well-characterizedfor various genes.18
Transforming growth factor-b enhances the for-mation of heterodimeric complexes of SMAD3 andSMAD4, leading to the translocation of thesecomplexes into the nucleus. SMAD4 then activatesintra-nuclear transcription of p21waf1/cip1.19 Ourdata from gene profile analysis showing high levelsof p21waf1/cip1 trans-activating genes is indicative ofEZH2-shRNA action at multiple levels to bringabout tumor suppression.
Although the mechanism by which EZH2becomes overexpressed in a variety of cancers isnot entirely clear, aberrant regulation by miRNAshas been proposed by both our laboratory andothers. We demonstrated recently that miR-101inhibits pancreatic9 and ovarian cancer12 via nega-tive regulation of EZH2 expression, corroboratingresults of others in lung cancer.20
In conclusion, we have provided evidence thatRNAi-mediated knockdown of EZH2 inhibits chro-matin condensation contributing to enhancedexpression of p21waf1/cip1 and its transcriptionaltransactivators. This observation, together with
downregulation of mutant p53 and H3k27me3,provides a possible molecular mechanism forEZH2-shRNA–mediated inhibition of PDAC.
REFERENCES
1. Chen YH, Hung MC, Li LY. EZH2: a pivotal regulator incontrolling cell differentiation. Am J Transl Res 2012;4:364.
2. Chang C, Hung M. The role of EZH2 in tumour progres-sion. Br J Cancer 2011;106:243-7.
3. Crea F, Fornaro L, Bocci G, Sun L, Farrar WL, Falcone A,et al. EZH2 inhibition: targeting the crossroad of tumor in-vasion and angiogenesis. Cancer Metast Rev 2012:1-9.
4. Bao B, Ali S, Banerjee S, Wang Z, Logna F, Azmi AS, et al.Curcumin analogue CDF inhibits pancreatic tumor growthby switching on suppressor microRNAs and attenuatingEZH2 expression. Cancer Res 2012;72:335-45.
5. van Vlerken LE, Kiefer CM, Morehouse C, Li Y, Groves C,Wilson SD, et al. EZH2 is required for breast and pancreaticcancer stem cell maintenance and can be used as a func-tional cancer stem cell reporter. Stem Cells Transl Med2013;2:43-52.
6. Ougolkov A, Bilim V, Billadeau D. Regulation of pancreatictumor cell proliferation and chemoresistance by the his-tone methyltransferase enhancer of zeste homologue 2.Clin Cancer Res 2008;14:6790.
7. Wei Y, Xia W, Zhang Z, Liu J, Wang H, Adsay NV, et al. Lossof trimethylation at lysine 27 of histone H3 is a predictor ofpoor outcome in breast, ovarian, and pancreatic cancers.Mol Carcinog 2008;47:701-6.
8. Matsubara T, Sato Y, Sasaki M, Harada K, Nomoto K, Tsu-neyamaK,et al. Immunohistochemical characteristics andma-lignant progression of hepatic cystic neoplasms in comparisonwith pancreatic counterparts. Hum Pathol 2012;43:2177-86.
9. Qazi AM, Gruzdyn O, Semaan A, Seward S, Chamala S, Dhu-lipala V, et al. Restoration of E-cadherin expression inpancreatic ductal adenocarcinoma treated with micro-RNA-101. Surgery 2012;152:704-11.
10. Nakahara O, Takamori H, Iwatsuki M, Baba Y, Sakamoto Y,Tanaka H, et al. Carcinogenesis of intraductal papillarymucinous neoplasm of the pancreas: loss of microRNA-101 promotes overexpression of histone methyltransferaseEZH2. Ann Surg Oncol 2012;19:565-71.
11. Guo J, Cai J, Yu L, Tang H, Chen C, Wang Z. EZH2 regulatesexpression of p57 and contributes to progression of ovariancancer in vitro and in vivo. Cancer Sci 2011;102:530-9.
12. Semaan A, Qazi AM, Seward S, Chamala S, Bryant CS, Ku-mar S, et al. MicroRNA-101 inhibits growth of epithelialovarian cancer by relieving chromatin-mediated transcrip-tional repression of p21 waf1/cip1. Pharm Res 2011;28:3079-90.
13. el-Deiry WS, Harper JW, O’Connor PM, Velculescu VE, Can-man CE, Jackman J, et al. WAF1/CIP1 is induced in p53-mediated G1 arrest and apoptosis. Cancer Res 1994;54:1169-74.
14. Garcea G, Neal C, Pattenden C, Steward W, Berry D. Molec-ular prognostic markers in pancreatic cancer: a systematicreview. Eur J Cancer 2005;41:2213-36.
15. van der Vlag J, Otte A. Transcriptional repression mediatedby the human polycomb-group protein EED involves his-tone deacetylation. Nat Genet 1999;23:474-8.
16. Hsiao M, Low J, Dorn E, Ku D, Pattengale P, Yeargin J, et al.Gain-of-function mutations of the p53 gene induce lympho-hematopoietic metastatic potential and tissue invasiveness.Am J Pathol 1994;145:702.
17. Friedman J, Liang G, Liu C, Wolff E, Tsai Y, Ye W, et al. Theputative tumor suppressor microRNA-101 modulates thecancer epigenome by repressing the polycomb group pro-tein EZH2. Cancer Res 2009;69:2623.
18. Bracken AP, Kleine-Kohlbrecher D, Dietrich N, Pasini D,Gargiulo G, Beekman C, et al. The Polycomb group pro-teins bind throughout the INK4A-ARF locus and are disas-sociated in senescent cells. Genes Dev 2007;21:525-30.
19. Hunt KK, Fleming JB, Abramian A, Zhang L, Evans DB,Chiao PJ. Overexpression of the tumor suppressor geneSmad4/DPC4 induces p21waf1 expression and growth in-hibition in human carcinoma cells. Cancer Res 1998;58:5656.
20. Kota J, Chivukula RR, O’Donnell KA, Wentzel EA, Mont-gomery CL, Hwang HW, et al. Therapeutic microRNA deliv-ery suppresses tumorigenesis in a murine liver cancermodel. Cell 2009;137:1005-17.
DISCUSSION
Dr Margo Shoup (Warrenville, IL): You have actuallydemonstrated your efforts to decipher the mechanismsof proliferation and growth of pancreatic cancer. Weknow that EZH2 is shown to be upregulated in pancre-atic cancer, and that’s been demonstrated previously,but you’re now trying to demonstrate a potentialpathway for when inhibiting EZH2 leads to attenuationof growth of pancreatic cancer. I think your final slideis really the one that wraps this whole study up veryelegantly.
The way I understand it is that you found that byknocking down EZH2, you showed a decrease inexpression of mutant p53. But, really, what we’re lookingfor is p53 as a tumor suppressor gene, so did you find acorrelation with an increase in the wild-type p53 whenyou knocked down EZH2?
Also, you found in several really elegant, clinicalexperiments that the pancreatic cancer cells that weretransfected with EZH2-shRNA are knocking down EZH2and had attenuated growth, both in subculture and denovo in the xenograft model. However, there was stillgrowth of tumor. It was attenuated growth, but there wasstill growth.
So my question is, to make this more clinicallyapplicable, have you tried actually transfecting the actualtumors when you see them? That’s how we really treatpatients in real life. And I think it would be naive for anyof us to think there’s any one magic bullet for any ofthese cancers. Is there any kind of synergistic effect thattransfecting these cells may have with something likegemcitabine or another targeted agent of some type toattenuate cell growth?
Dr Ramesh Batchu (Detroit, MI): We actually areworking on a few of your comments. And coming toyour first question, I want to go back to the slides, if itis possible.
When we tested with the antibody, the antibodyaccurately measures both wild-type and mutant p53. Wefound another antibody that binds to only mutant p53.When we see similar levels of p53 total protein and whenwe see the drastic direction in only the mutant, just bycomparing these results, we would want to think that
wild-type p53 actually increased. But we are actuallylooking for other antibodies available that we could testthe wild-type p53 itself, just to confirm our results. Thankyou for pointing that out. We are actually doing that.
And your second question, yes, there is still growth oftumor in EZH2 knockdown cells in SCID mice, althoughit was 3 times less than the control cells. We do thinkthat there are other ways to inhibit the tumor processand perhaps we have to increase the amount of EZH2 weare using, or we actually look at gemcitabine as acombination therapy with the EZH2 with 1 particularcell line, we have conclusive results. With another cellline, we do not have conclusive results yet. But I do agreewith you that we should go for some kind of combina-tion therapy.
Dr Ronald J. Weigel (Iowa City, IA): I appreciate yourstudies. Very nicely presented. I had 2 questions. Thefirst is, your ChIP data implies that EZH2 is binding tothe p21 promoter. So my first question is, where is thatprobe and where is that binding relative to the bindingsites for p53 and other factors that are known to regulatep21 expression?
The second is, this finding that EZH2 knockdownspecifically inhibits expression of mutant p53. Becausewild-type p53 is either not changing or increasing, itsuggests that EZH2 is functioning through a nontran-scriptional mechanism. So what mechanism do youpropose for EZH2 knockdown to specifically knock outexpression of a specific mutant of p53 and leave wild-type p53 unaffected?
Dr Ramesh Batchu:These are actually wonderful ques-tions. My apologies, I do not have answers for that.
Yes, we see that the p53 mutant is decreased and thenp21 is increased in ChIP assays. And your question isvalid, and how are we going to make sense of these two?And, as of now, we do not have the data to show thatthese two are interlinked or in any way connected, ormaybe EZH2 is acting on different pathways to work onthese two. So, unfortunately, I do not have an answer forthat. We are still working on that.
And then, your second question is actually related tothe first one, of the mutant p53, whether it is workingnot at the level of transcription but at the level of proteintranslation, because only the mutant p53 is changed.Obviously, it is not at the level of transcription, but we donot know which pathway EZH2 is taking to inhibit thep53. We have several ideas on how to go about it. Rightnow, I don’t want to comment on what the pathway is, wewant to check it.
Dr Scott Wilhelm (Cleveland, OH): My comment toyou is I applaud you for not parceling out your data. Somany times we come to meetings and people will takemechanistic data such as yours and they’ll parcel it outandmake 4 abstracts out of it. We get very limited picturesat 4 separate meetings and you don’t get to see the wholeend product. So I compliment youon taking amechanismand giving us both in vitro and in vivo data.
My question to you is that one of the problems withcell culture data is that individual cell lines can producedifferent problems, different mechanisms, and different
gene upregulations. There’s 3 other fairly good pancre-atic cell culture lines that are out there. Have you triedlooking at your data in any other culture lines tointernally validate your study?
Dr Ramesh Batchu: I want to thank you for acknowl-edging our work.
Yes, we worked on several other pancreatic cancer celllines: MIA-PaCa-2, BxPc-3, and then PANC-1. For cell
growth analysis and a few other assays, which are easierto do, we did use all the cell lines. We do have all thedata. But coming to the gene profile analysis and ChIPassays, we choose PANC-1 cells for various reasons, suchas they are easier to grow and so on.
Most of the studies we did with 1 cell line. The mostbasic work we did do with 3 more cell lines, but they arenot presented here.