Proc. Natl. Acad. Sci. USA Vol. 85, pp. 5556-5560, August 1988 Cell Biology Interleukin 1 prevents loss of corticotropic responsiveness to ,f-adrenergic stimulation in vitro (catecholamine/corticotropin releasing factor) MICHAEL BOYLE*, GAYLE YAMAMOTO, MIA CHEN, JEAN RIVIER, AND WYLIE VALE The Clayton Foundation Laboratories for Peptide Biology, The Salk Institute, 10010 North Torrey Pines Road, La Jolla, CA 92037 Communicated by Julius Axelrod, March 14, 1988 (received for review December 15, 1987) ABSTRACT Corticotropin (ACTH) secretion by the ante- rior pituitary is stimulated by catecholamines in vivo and in vitro. The nature of the response in vivo is controversial but appears to be mediated by .3-adrenergic receptors, whereas the response is dependent on a-adrenergic receptors in cultured anterior pituitary cells. In the present studies, by using a superfusion technique, we demonstrate that catecholamine stimulation of ACTH release from rat anterior pituitaries changes with time from a predominantly P-adrenergic- mediated event to a predominantl a-adrenergic-mediated event. From 0 to 2 hr after initiating the superfusion, release of ACTH from anterior pituitary glands is stimulated up to 2.4-fold by the (1wadrenergic agonist l-isoproterenol. However, the ACTH secretory response to the a-adrenergic agonist l-phenylejhrine is -<5% of that to l-isoproterenol during the same time period. Beginning 2 hr after the start of the superfusion, the responsiveness to the fi-adrenergic agonist declines, and the response to the c-adrenergic agonist increases until, 10 hr after removal, >95% of the catecholamine- inducible ACTH release is mediated by an a-adrenergic pathway. The addition of interleukin 1 alone to the medium from the beginning of the superfusion does not modify basal ACTH secretion rates and does not affect the acquisition of the response to phenylephrine. However, the presence of interleu- kin 1 does allow the maintenance of the full ACTH secretory response to isoproterenol. This effect of interleukin 1 is reversed by an interleukin 1 antagonist. These observations suggest an additional way in which immune regulators might interact with the hypothalamic-pituitary-adrenal axis. The secretion of corticotropin (ACTH) by the anterior pituitary gland is regulated by the interaction of several central and peripheral factors including corticotropin- releasing factor (CRF), vasopressin, catecholamines, and glucocorticoids (1, 2). In vivo studies in rats have suggested that catecholamines secreted from the hypothalamus (3-5) or peripheral tissues (adrenal medulla and sympathetic nerve endings) may contribute to basal or stress-induced stimula- tion of ACTH release from the anterior pituitary gland (6-8). Intravenous administration of low doses of epinephrine stimulates ACTH and corticosterone secretion (7), an effect that was interpreted to be due to direct stimulation of the corticotropic cells in the anterior pituitary gland. This re- sponse can be reproduced by the f8-adrenergic agonist 1- isoproterenol and blocked by the p-adrenergic antagonist propranolol (9, 10). Administration of high doses of a- adrenergic agonists also stimulates ACTH secretion, but this effect may be mediated largely through hypothalamic CRF release (11, 12). The effects of a-adrenergic agonists on ACTH (11, 12) or CRF (13) production are reversed by a1-adrenergic antagonists (11-15). In contrast to the situation observed in vivo, in primary cultures of rat anterior pituitary cells, catecholamines alone or in concert with CRF stimulate ACTH release, but this response is mediated through a1-adrenergic receptors (16, 17). For this reason, we decided to develop an in vitro system that would allow us to study the mechanism and significance of the apparent shift from a 8-adrenergic (in vivo)- to an a-adrenergic (in vitro)-mediated ACTH secretion. In addi- tion, because of the effects of the monokine interleukin 1 (IL-1) on the hypothalamic-pituitary-adrenal axis, we also investigated the effect of this monokine on the adrenergic- stimulated release of ACTH (18-20). METHODS Sprague-Dawley male rats (180-200 g) were decapitated and the pituitary glands were removed immediately. The poste- rior and intermediate lobes were discarded, and the anterior lobes were hemisected and washed in F12 medium supple- mented with Hepes (25 mM), glucose (2 mg/ml), gentamycin (50 mg/ml), 0.1% bovine serum albumin, and ascorbate (1 mg/ml). We also included CRF at a concentration (50 pM) that is found in rat portal blood. This concentration does not release ACTH on its own but augments the effect of cate- cholamines. Six equivalent anterior pituitary glands were placed into a polypropylene/glass column (0.7 x 4 cm; Bio-Rad) containing 0.2 ml of the above medium. Columns were then submerged in a 37°C water bath. Each column was connected to a peristaltic pump by polypropylene tubing (i.d., 0.062 inch; 1 inch = 0.02540 m) and superfused with the above medium at a flow rate of 0.2 ml/min. Five-minute fractions were collected in polypropylene test tubes (12 x 75 mm) and stored at - 20°C until assayed for ACTH by radioimmunoassay (RIA). Pituitaries were challenged for 5 min, at the indicated times, with 1 ml of 1 ,uM epinephrine (in the presence or absence of 10 ,uM prazosin or 10 ,M propranolol), 1 ,M l-isoproterenol, 1 ,uM l-phenylephrine, or 50 nM CRF dissolved in superfusion medium. To avoid complication of results by desensitization, each group of six anterior pituitary glands was exposed to a catecholamine only one time. The typical response to such a challenge would last from 0 to 40 min, depending on the dose and compound used. The increase of ACTH released (as compared to control) was then calculated. Each point reported represents the mean of six experiments. In those assays where the effect of recom- binant human IL-1 (rhIL-1) on ACTH release was tested, 100 pM rhIL-1 was included in the perfusate from the time of removal of the anterior pituitary glands. ACTH Hormone Assay. ACTH was assayed in the column effluent by RIA with porcine ACTH-(1-39) as the standard and iodinated tracer. Ovine antiserum to ACTH-(1-24) was provided by David Orth (Vanderbilt University, Nashville, Abbreviations: ACTH, corticotropin; CRF, corticotropin releasing factor; IL-1, interleukin 1; rhIL-1, recombinant human IL-1. *To whom reprint requests should be addressed. 5556 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript
Proc. Natl. Acad. Sci. USAVol. 85, pp. 5556-5560, August 1988Cell Biology
Interleukin 1 prevents loss of corticotropic responsiveness to,f-adrenergic stimulation in vitro
(catecholamine/corticotropin releasing factor)
MICHAEL BOYLE*, GAYLE YAMAMOTO, MIA CHEN, JEAN RIVIER, AND WYLIE VALEThe Clayton Foundation Laboratories for Peptide Biology, The Salk Institute, 10010 North Torrey Pines Road, La Jolla, CA 92037
Communicated by Julius Axelrod, March 14, 1988 (received for review December 15, 1987)
ABSTRACT Corticotropin (ACTH) secretion by the ante-rior pituitary is stimulated by catecholamines in vivo and invitro. The nature of the response in vivo is controversial butappears to be mediated by .3-adrenergic receptors, whereas theresponse is dependent on a-adrenergic receptors in culturedanterior pituitary cells. In the present studies, by using asuperfusion technique, we demonstrate that catecholaminestimulation of ACTH release from rat anterior pituitarieschanges with time from a predominantly P-adrenergic-mediated event to a predominantl a-adrenergic-mediatedevent. From 0 to 2 hr after initiating the superfusion, releaseof ACTH from anterior pituitary glands is stimulated up to2.4-fold by the (1wadrenergic agonist l-isoproterenol. However,the ACTH secretory response to the a-adrenergic agonistl-phenylejhrine is -<5% of that to l-isoproterenol during thesame time period. Beginning 2 hr after the start of thesuperfusion, the responsiveness to the fi-adrenergic agonistdeclines, and the response to the c-adrenergic agonist increasesuntil, 10 hr after removal, >95% of the catecholamine-inducible ACTH release is mediated by an a-adrenergicpathway. The addition of interleukin 1 alone to the mediumfrom the beginning of the superfusion does not modify basalACTH secretion rates and does not affect the acquisition of theresponse to phenylephrine. However, the presence of interleu-kin 1 does allow the maintenance of the full ACTH secretoryresponse to isoproterenol. This effect of interleukin 1 isreversed by an interleukin 1 antagonist. These observationssuggest an additional way in which immune regulators mightinteract with the hypothalamic-pituitary-adrenal axis.
The secretion of corticotropin (ACTH) by the anteriorpituitary gland is regulated by the interaction of severalcentral and peripheral factors including corticotropin-releasing factor (CRF), vasopressin, catecholamines, andglucocorticoids (1, 2). In vivo studies in rats have suggestedthat catecholamines secreted from the hypothalamus (3-5) orperipheral tissues (adrenal medulla and sympathetic nerveendings) may contribute to basal or stress-induced stimula-tion ofACTH release from the anterior pituitary gland (6-8).Intravenous administration of low doses of epinephrinestimulates ACTH and corticosterone secretion (7), an effectthat was interpreted to be due to direct stimulation of thecorticotropic cells in the anterior pituitary gland. This re-sponse can be reproduced by the f8-adrenergic agonist 1-isoproterenol and blocked by the p-adrenergic antagonistpropranolol (9, 10). Administration of high doses of a-adrenergic agonists also stimulates ACTH secretion, but thiseffect may be mediated largely through hypothalamic CRFrelease (11, 12). The effects of a-adrenergic agonists onACTH (11, 12) or CRF (13) production are reversed bya1-adrenergic antagonists (11-15).
In contrast to the situation observed in vivo, in primarycultures of rat anterior pituitary cells, catecholamines aloneor in concert with CRF stimulate ACTH release, but thisresponse is mediated through a1-adrenergic receptors (16,17). For this reason, we decided to develop an in vitro systemthat would allow us to study the mechanism and significanceof the apparent shift from a 8-adrenergic (in vivo)- to ana-adrenergic (in vitro)-mediated ACTH secretion. In addi-tion, because of the effects of the monokine interleukin 1(IL-1) on the hypothalamic-pituitary-adrenal axis, we alsoinvestigated the effect of this monokine on the adrenergic-stimulated release of ACTH (18-20).
METHODSSprague-Dawley male rats (180-200 g) were decapitated andthe pituitary glands were removed immediately. The poste-rior and intermediate lobes were discarded, and the anteriorlobes were hemisected and washed in F12 medium supple-mented with Hepes (25 mM), glucose (2 mg/ml), gentamycin(50 mg/ml), 0.1% bovine serum albumin, and ascorbate (1mg/ml). We also included CRF at a concentration (50 pM)that is found in rat portal blood. This concentration does notrelease ACTH on its own but augments the effect of cate-cholamines. Six equivalent anterior pituitary glands wereplaced into a polypropylene/glass column (0.7 x 4 cm;Bio-Rad) containing 0.2 ml of the above medium. Columnswere then submerged in a 37°C water bath. Each column wasconnected to a peristaltic pump by polypropylene tubing(i.d., 0.062 inch; 1 inch = 0.02540 m) and superfused with theabove medium at a flow rate of 0.2 ml/min. Five-minutefractions were collected in polypropylene test tubes (12 x 75mm) and stored at - 20°C until assayed for ACTH byradioimmunoassay (RIA). Pituitaries were challenged for 5min, at the indicated times, with 1 ml of 1 ,uM epinephrine (inthe presence or absence of 10 ,uM prazosin or 10 ,Mpropranolol), 1 ,M l-isoproterenol, 1 ,uM l-phenylephrine, or50 nM CRF dissolved in superfusion medium. To avoidcomplication of results by desensitization, each group of sixanterior pituitary glands was exposed to a catecholamine onlyone time. The typical response to such a challenge would lastfrom 0 to 40 min, depending on the dose and compound used.The increase ofACTH released (as compared to control) wasthen calculated. Each point reported represents the mean ofsix experiments. In those assays where the effect of recom-binant human IL-1 (rhIL-1) on ACTH release was tested, 100pM rhIL-1 was included in the perfusate from the time ofremoval of the anterior pituitary glands.ACTH Hormone Assay. ACTH was assayed in the column
effluent by RIA with porcine ACTH-(1-39) as the standardand iodinated tracer. Ovine antiserum to ACTH-(1-24) wasprovided by David Orth (Vanderbilt University, Nashville,
Abbreviations: ACTH, corticotropin; CRF, corticotropin releasingfactor; IL-1, interleukin 1; rhIL-1, recombinant human IL-1.*To whom reprint requests should be addressed.
5556
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 85 (1988) 5557
TN) and the assay was carried out as described (21). Theintra- and interassay coefficients of variation were <10%.
Peptides and Biochemicals. Synthetic ovine CRF and theIL-1 antagonist hIL-1,3-(237-269) were prepared by solid-phase methodology (22). The one letter code sequence ofhIL-lp-(237-269) is as follows:
YISTSQAENMPVFLGGTKGGQDITDFTMQFVSS.
The peptide was characterized by amino acid analysis thatgave the expected integer ratios and by fast atom bombard-ment mass spectrometry (observed MH', 3556.83; calcu-lated, 3556.66); its high purity was demonstrated by HPLCanalysis in two solvent systems with Vydac C18 columns.l-Isoproterenol, l-phenylephrine, propranolol, and epineph-rine were purchased from Sigma. Prazosin hydrochloride wasa generous gift from Pfizer. IL-1 was generously provided byPeter Lomedico (Hoffmann-La Roche) or purchased fromCollaborative Research (Waltham, MA). Each compoundwas dissolved in the superfusion medium at the concentra-tions indicated.
Statistical Analyses. Raw data were tested for normalitywith either the Kolmogorov-Smirnov or the Wilk-Shapirotest. All samples were obtained from populations whosedistributions were normal. The paired t-test was used foranalysis of significant overall effects (Bolt, Beranek, andNewman Research Systems RS/1 Biocomputing Software).All data are presented as the mean ± SEM.
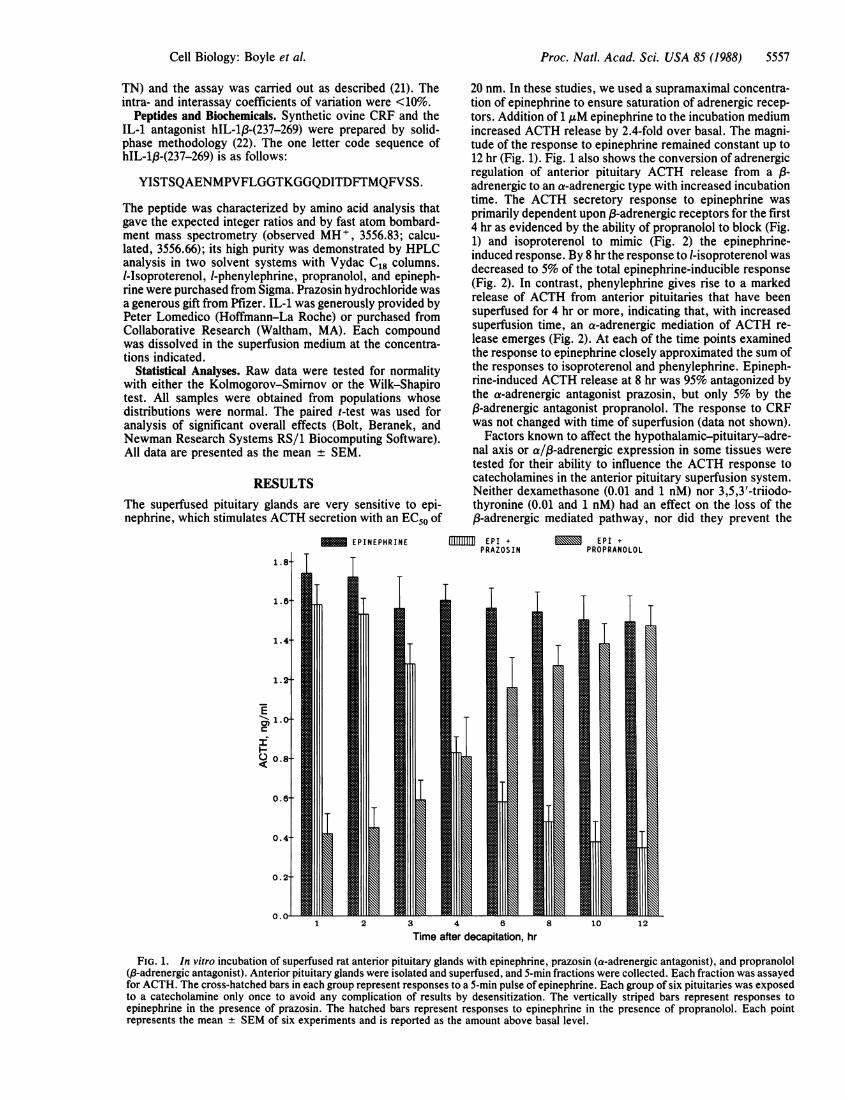
RESULTSThe superfused pituitary glands are very sensitive to epi-nephrine, which stimulates ACTH secretion with an EC50 of
20 nm. In these studies, we used a supramaximal concentra-tion of epinephrine to ensure saturation of adrenergic recep-tors. Addition of 1 AM epinephrine to the incubation mediumincreased ACTH release by 2.4-fold over basal. The magni-tude of the response to epinephrine remained constant up to12 hr (Fig. 1). Fig. 1 also shows the conversion of adrenergicregulation of anterior pituitary ACTH release from a /8-adrenergic to an a-adrenergic type with increased incubationtime. The ACTH secretory response to epinephrine wasprimarily dependent upon P3-adrenergic receptors for the first4 hr as evidenced by the ability of propranolol to block (Fig.1) and isoproterenol to mimic (Fig. 2) the epinephrine-induced response. By 8 hr the response to l-isoproterenol wasdecreased to 5% of the total epinephrine-inducible response(Fig. 2). In contrast, phenylephrine gives rise to a markedrelease of ACTH from anterior pituitaries that have beensuperfused for 4 hr or more, indicating that, with increasedsuperfusion time, an a-adrenergic mediation of ACTH re-lease emerges (Fig. 2). At each of the time points examinedthe response to epinephrine closely approximated the sum ofthe responses to isoproterenol and phenylephrine. Epineph-rine-induced ACTH release at 8 hr was 95% antagonized bythe a-adrenergic antagonist prazosin, but only 5% by theP-adrenergic antagonist propranolol. The response to CRFwas not changed with time of superfusion (data not shown).
Factors known to affect the hypothalamic-pituitary-adre-nal axis or a/p3-adrenergic expression in some tissues weretested for their ability to influence the ACTH response tocatecholamines in the anterior pituitary superfusion system.Neither dexamethasone (0.01 and 1 nM) nor 3,5,3'-triiodo-thyronine (0.01 and 1 nM) had an effect on the loss of the03-adrenergic mediated pathway, nor did they prevent the
- EPINEPHRINE
TEPI +
PRAZOSIN
I T
1 2 3 4 6
Time after decapitation, hr
EPI +PROPRANOLOL
IT
8 10 12
FIG. 1. In vitro incubation of superfused rat anterior pituitary glands with epinephrine, prazosin (a-adrenergic antagonist), and propranolol(,3-adrenergic antagonist). Anterior pituitary glands were isolated and superfused, and 5-min fractions were collected. Each fraction was assayedfor ACTH. The cross-hatched bars in each group represent responses to a 5-min pulse of epinephrine. Each group of six pituitaries was exposedto a catecholamine only once to avoid any complication of results by desensitization. The vertically striped bars represent responses toepinephrine in the presence of prazosin. The hatched bars represent responses to epinephrine in the presence of propranolol. Each pointrepresents the mean SEM of six experiments and is reported as the amount above basal level.
1. 4
1.6
1.4+
1 .2
C
I-I-
1.0o
0.8+
0. 6B
0.4+
_;
Cell Biology: Boyle et al.
0.2-
o. ol I WIN WIN Imill
Proc. Natl. Acad. Sci. USA 85 (1988)
|_ EPINEPHRINE
1.8{ T
~ ISOPROTERENOL PHENYLEPHRINE
TT
1.6t I
1.2+
1.*0
0.8+
0. 6t
0.4+
0.2+
O.0-'-10
iilllll l ml tt AmEI 1 -i 91 11 m _sm- _ m _Emm _amm _mm _i HEmmIllSlHffl Hm _immi Illmm _sm_ _ _ _ worm _sm. mam91 tmiEI! m Ami i m Grimmi 53 i ffi A.
1 2 3 4 6 8Time after decapitation, hr
appearance of the a-adrenergic mediated pathway (data notshown). In the presence of 100 pM rhIL-1, the ACTHresponse to the B-adrenergic agonist l-isoproterenol wasmaintained for at least 8 hr without any change in the onsetor magnitude ofthe response to phenylephrine (Figs. 3 and 4).In the presence of rhIL-1 the response to epinephrineincreased significantly but not to the same degree as theresponse to isoproterenol increased. The effect of rhIL-1 on
ISOPROTERENOL IL-1 + ISOPROTERENOL
FIG. 2. In vitro incubation of superfused ratanterior pituitary glands with l-isoproterenol andphenylephrine. Conditions were as reported in Fig.1. The cross-hatched bars represent responses to a5-min pulse ofepinephrine. The solid bars representresponses to a 5-min pulse of l-isoproterenol. Thecross-striped bars represent responses to a 5-minpulse of phenylephrine. Each point represents themean ± SEM of six experiments and is reported asthe amount above basal level.
l-isoproterenol stimulated release of ACTH was blocked bythe IL-1 antagonist hIL-1p-(237-269) (23) (Fig. 3). Pretreat-ment with rhIL-1 had no effect on secretion of ACTH thatoccurred in response to CRF.
DISCUSSIONIn rats, catecholamines appear to contribute to stress-induced ACTH secretion in at least two ways. Adrenergic
IL-1 ANTAGONIST+ IL-1 + ISOPROTERENOL
I0:
1 2 4 6
Time after decapitation, hr
FIG. 3. In vitro incubation of superfused ratanterior pituitary glands with l-isoproterenol in thepresence and absence of rhIL-1. The solid barsrepresent responses to a 5-min pulse of 1-
isoproterenol alone. The horizontally striped barsrepresent responses to l-isoproterenol when rhIL-1
has been present since the start of superfusion. Thehatched bars represent the response to l-isoprote-renol when rhIL-1 and the IL-1 antagonist hIL-1l3-(237-269) have been present since the start ofsuperfusion. Each group of six pituitaries, eachpoint, was exposed to a catecholamine only once toavoid any complication of results by desensitiza-tion. Each point represents the mean SEM of six
8 experiments and is reported as the amount abovebasal level.
I-0
I
5558 Cell Biology: Boyle et al.
1. 4t
12
Proc. Natl. Acad. Sci. USA 85 (1988) 5559
I-0
ISOVLL
Phe EpiFIG. 4. Effect of rhIL-1 on catecholamine-induced ACTH release
from superfused rat anterior pituitary glands at 8 hr. Conditions are thesame as those reported in Fig. 3. Open bars represent samples incubatedwithout IL-1. Hatched bars represent samples incubated with IL-1.Each point represents the mean ± SEM of six experiments and isreported as the amount above basal level. Asterisks indicate a signifi-cant difference (P < 0.05) between groups untreated and treated withhIL-1. Iso, I-isoproterenol; Phe, phenylephrine; Epi, epinephrine.
afferents originating in the midbrain terminate in the CRF-containing region of the paraventricular nucleus (24-28).Electrical stimulation of these afferents causes a release ofCRF into portal blood, which is blocked by chemical orelectrical lesions of the paraventricular nucleus (12, 13, 28).Such lesions prevent the release of CRF into hypothalamic-portal blood (13). Moreover, intracerebroventricular injec-tions of low doses of norepinephrine into rats produce adramatic increase in CRF release-an effect that is blockedby a1-adrenergic antagonists (13, 15). Apart from centraleffects in the hypothalamus, catecholamines also appear tostimulate ACTH release directly from the anterior pituitary inrats because rats with pituitary-stalk transection respond toperipherally injected fB-adrenergic agonists with an increasein ACTH secretion (9, 10). A number of studies from severallaboratories have demonstrated that the in vivo pituitaryresponse to catecholamines is mediated by /-adrenergicreceptors (7-10), although some uncertainty remains (29).More direct evidence supporting effects of catecholamines
on anterior pituitary release ofACTH has come from studieswith primary cultures of rat anterior pituitary cells. In thesestudies, norepinephrine and epinephrine stimulate ACTHrelease as much as 3-fold (2). However, in contrast to thefindings in vivo, the ACTH secretory response of culturedcells has been observed to be mediated by a1-adrenergicreceptors (16, 17). It has been reported that a- and /-adrenergic-mediated ACTH responses coexist in undi-spersed rat anterior pituitary glands in short-term incuba-tions.t In the present study, we have shown that the rat
anterior hemipituitaries respond in a biphasic manner tocatecholamines in a superfusion system. From 0 to 4 hr afterremoval of the anterior pituitary glands from the rat, thecatecholamine response is mediated predominantly by a3-adrenergic pathway; after 4 hr, an a-adrenergic pathwaydominates.The importance of other hormonal factors for the activity of
one or the other of the catecholamine pathways has beendemonstrated many times in vivo. Results from several studieshave suggested that both glucocorticoid and thyroid hormonescan influence the nature of adrenergic responses by alteringeither the receptor expression or the effectiveness of theguanine nucleotide-binding regulatory proteins (30, 31). Hy-drocortisone increases /3-adrenergic receptor synthesis in lungcells (32). In the liver, in vivo, 3,5,3'-triiodothyronine levelsalter expression of a- and /3-adrenergic pathways (33-35). Asin the pituitary, the conversion in liver cells is triggered by thechange from in vivo to in vitro conditions, develops rapidly (in<4 hr) in a serum-free medium, and cannot be reversed by invitro exposure to hormones (36). In hypothyroid rats, a1-adrenergic receptor-mediated secretion of P3-endorphin andother corticotrophic products is suppressed (37). However, inour studies, neither dexamethasone nor 3,5,3'-triiodothyro-nine had a stimulus-selective effect on the loss of the 13-adrenergic-mediated pathway in our superfusion system, nordid they prevent the appearance of the a-adrenergic-mediatedpathway.Evidence strongly suggests that just as the adrenocortical
axis is capable of regulating immune function, the immunesystem reciprocally exerts neuroendocrine control over theadrenocortical axis (38). The monokine IL-1 has been foundto stimulate the release of cortisol (38). It is believed that thisactivity of IL-1 is mediated through the higher levels of thehypothalamic-pituitary-adrenal axis since IL-1 has no directeffect on the adrenal gland (20, 38), and injection of IL-1 hasbeen demonstrated to elevate plasma ACTH levels in vivo(18, 20, 39, 40). Woloski et al. (19) have reported an acuteACTH response to IL-1 by cultured AtT-20 pituitary tumorcells, and Bernton et al. (41) have reported a similar responsein primary cultures of rat anterior pituitary cells. However,several other groups, including our own, have found no effectof IL-1 on basal or CRF-mediated secretion of ACTH bycultured anterior pituitary cells (18, 40). Furthermore, we (18)and Berkenbosch et al. (40) have shown that the IL-i-inducedrelease ofACTH was decreased by anti-CRF serum, showingthat the response to IL-1 was mediated by or dependent uponthe presence of CRF. We have also demonstrated thatinjection of IL-1 into rats increased pituitary-portal bloodlevels of CRF (18), and Berkenbosch et al. (40) observed anIL-i-mediated depletion of CRF in the hypothalamus ofcolchicine-pretreated rats. To further elucidate the action ofIL-1 on the hypothalamic-pituitary-adrenal axis, we exam-ined the effect of IL-1 on catecholamine-induced ACTHrelease from superfused anterior pituitary glands. We reporthere that in the presence of IL-1, the ACTH response of ratanterior pituitaries to P-adrenergic agents is maintained invitro, whereas the response to CRF and the emergence of ana-adrenergic-mediated response, with increased superfusiontime, are unaltered. In contrast to the findings of Woloski etal. (19) and Bernton et al. (41) but in agreement with Ueharaet al. (39) and Berkenbosch et al. (40), we do not find a directstimulation of ACTH release by IL-1.Our findings suggest a possible resolution to the conflicting
data that demonstrate 83-adrenergic mediation of ACTHsecretion in vivo and a-adrenergic mediation in vitro: Thelack of IL-1, or an IL-1 by-product, under culture conditions,facilitates the loss of the 83-adrenergic-mediated response.This function of IL-1 may represent an interaction betweenthe immune system and the endocrine system. The facilita-tion of the catecholamine-induced ACTH response by the
tChen, F. M., Proceedings of the 68th Annual Meeting of theEndocrine Society, Anaheim, CA, June 25-27, 1986, p. 196, Abstr.663.
Cell Biology: Boyle et al.
Proc. Natl. Acad. Sci. USA 85 (1988)
anterior pituitary that we report here would appear to besynergistic with the IL-1-induced CRF release we (18) andothers (39, 40) have reported.
The assistance of Drs. Myriam Baes and Eric Widmaier isappreciated. We also thank Robert Galyean, Ron Kaiser, and JohnDykert for technical assistance, and Susan McCall for assistancewith the manuscript. This work was supported by Grants DK26741and AA07454 from the National Institutes of Health and the MandelFamily Foundation. Research was conducted in part by the ClaytonFoundation for Research, California Division. W.V. is a ClaytonFoundation Investigator.
1. Axelrod, J. & Reisine, T. D. (1984) Science 224, 452-459.2. Vale, W., Vaughan, J., Smith, M., Yamamoto, G., Rivier, J. &
Rivier, C. (1983) Endocrinology 113, 1121-1131.3. Plotsky, P., Bruhn, T. & Vale, W. (1985) Endocrinology 116,
633-639.4. Johnston, C. A., Gibbs, D. M. & Negro-Vila, A. (1983) Endo-
crinology 113, 819-821.5. Gibbs, D. M. (1985) Brain Res. 335, 360-364.6. Labrie, F., Giguere, V., Proulx, L. & Fefevre, G. (1984) J.
Steroid. Biochem. 20, 153-160.7. Berkenbosch, F., Vermes, I., Binnekade, R. & Tilders,
F. J. H. (1981) Life Sci. 29, 2249-2256.8. Tilders, F. J. H., Berkenbosch, F. & Smelik, P. G. (1982)
Endocrinology 110, 114-120.9. Mezey, E., Reisine, T., Palkovits, T., Brownstein, M. J. &
Axelrod, J. (1983) Proc. Natl. Acad. Sci. USA 80, 6728-6731.10. Mezey, E., Reisine, T., Brownstein, M. J., Palkovits, M. &
Axelrod, J. (1984) Science 226, 1085-1087.11. Weiner, R. I. & Ganong, W. F. (1978) Physiol. Rev. 58, 905-
976.12. Guillaume, V., Conte-Devolx, B., Szafarczyk, A., Malaval, F.,
Pares-Herbute, N., Grino, M., Alonso, G., Assenmacher, I. &Oliver, C. (1987) Neuroendocrinology 46, 143-146.
13. Plotsky, P. M. (1987) Endocrinology 121, 924-930.14. Proulx-Ferland, L., Breault, M. & Cote, J. (1982) Prog.
Neuropsychopharmacol. Biol. Psychiatry 6, 433-438.15. Plotsky, P. M., Otto, S. & Sutton, S. (1987) Life Sci. 41, 1311-
1317.16. Vale, W. & Rivier, C. (1977) Fed. Proc. Fed. Am. Soc. Exp.
Biol. 36, 2094-2099.17. Giguere, V., Cote, J. & Labrie, F. (1981) Endocrinology 109,
757-762.18. Sapolsky, R., Rivier, C., Yamamoto, G., Plotsky, P. & Vale,
W. (1987) Science 238, 522-524.
19. Woloski, B. M. R. N. J., Smith, E. M., Meyer, W. J., III,Fuller, G. M. & Blalock, J. E. (1985) Science 230, 1035-1037.
20. Besedovsky, H., Del Rey, A., Sorkin, E. & Dinarello, C. A.(1986) Science 233, 652-654.
21. Rivier, C., Brownstein, M., Spiess, J., Rivier, J. & Vale, W.(1982) Endocrinology 110, 272-278.
22. Rivier, J., Spiess, J. & Vale, W. (1983) Proc. Natl. Acad. Sci.USA 80, 4851-4855.
23. Palaszynski, E. W. (1987) Biochem. Biophys. Res. Commun.147, 204-211.
24. Swanson, L. W., Sawchenko, P. E., Rivier, J. & Vale, W. W.(1983) Neuroendocrinology 36, 165-186.
25. Mezey, E., Kiss, J. Z., Skirboll, L. R., Goldstein, M. &Axelrod, J. (1984) Nature (London) 310, 140-141.
26. Bruhn, T. O., Plotsky, P. M. & Vale, W. W. (1984) Endocri-nology 114, 57-62.
27. Antoni, F. A., Palkovits, M., Makara, G. B., Linton, E. A.,Lowry, P. J. & Kiss, J. Z. (1983) Neuroendocrinology 36, 415-423.
28. Szafarczyk, A., Malaval, F., Laurent, A., Gibaud, R. &Assenmacher, I. (1987) Endocrinology 121, 883-892.
29. Makara, G. B., Kvetnansky, R., Jezova, D., Jindra, A., Ka-kucska, I. & Oprsalova, Z. (1986) Endocrinology 119, 1757-1762.
30. Itoh, H., Okajima, F. & Ui, M. (1984) J. Biol. Chem. 259,15464-15473.
31. Kunos, G. & Ishac, E. J. N. (1987) Biochem. Pharmacol. 36,1185-1191.
32. Fraser, C. M. & Venter, J. C. (1980) Biochem. Biophys. Res.Commun. 94, 390-397.
33. Malbon, C. C., Li, S. & Fain, J. N. (1978) J. Biol. Chem. 253,8820-8825.
34. Preiksaitis, H. G. & Kunos, G. (1979) Life Sci. 24, 35-42.35. Preiksaitis, H. G., Kan, W. H. & Kunos, G. (1982) J. Biol.
Chem. 257, 4321-4327.36. Kunos, G., Hirata, F., Ishac, E. J. N. & Tchakarov, L. (1984)
Proc. Natl. Acad. Sci. USA 81, 6178-6182.37. Ishac, E. J. N., Eskay, R., Hirata, F., Axelrod, J. & Kunos, G.
(1987) Endocrinology 120, 1073-1078.38. Beise, W. R. (1985) Prog. Leukocyte Biol. 2, 3-12.39. Uehara, A., Gottschall, P. E., Dahl, R. R. & Arimura, A.
(1987) Endocrinology 121, 1580-1582.40. Berkenbosch, F., van Oers, J., Del Rey, A., Tilders, A. &
Besedovsky, H. (1987) Science 238, 524-526.41. Bernton, E. W., Beach, J. E., Holaday, J. W., Smallridge,