Revista Brasil. Bot., V.29, n.1, p.163-174, jan.-mar. 2006 Fenologia da floração e biologia floral de bromeliáceas ornitófilas de uma área da Mata Atlântica do Sudeste brasileiro CAIO GRACO MACHADO 1,3 e JOÃO SEMIR 2 (recebido: 7 de outubro de 2004; aceito: 2 de fevereiro de 2006) ABSTRACT – (Flowering phenology and floral biology of some ornitophilous Bromeliaceae of an Atlantic forest area in southeastern Brazil). This study dealt with phenology and flower visitors of some Bromeliaceae species in an Atlantic Forest area at Parque Estadual Intervales, southeastern Brazil. The taxa studied belonged to the genera Tillandsia L. (3 spp.), Vriesea Lindl. (5 spp.), Aechmea Ruiz & Pav. (3 spp.), Billbergia Thunb. (1 sp.) and Nidularium Lem. (2 spp.). Sample transects were established in areas with different succession stages, where the bromeliads were localized and marked. Flowering phenology surveys were made monthly. The bromeliad community showed a sequential flowering all along the year and most species flowered during the rainy season. The floral visitors were recorded by naturalistic observations. Eight hummingbird species visited the bromeliads flowers. Among them, Phaethornis eurynome Lesson and Thalurania glaucopis Gmelin were the most frequent visitors. Bromeliads were divided in two groups by similarity analysis of floral visitors (hummingbirds): a group represented by species visited by Trochilinae hummingbirds and another visited by Phaethornis eurynome (subfamily Phaethornithinae). The competition among bromeliad species for pollinators is reduced due to their spatial distribution and the flowering periods. Key words - Atlantic Forest, Bromeliaceae, floral biology, hummingbirds, phenology RESUMO – (Fenologia de floração e biologia floral de bromeliáceas ornitófilas de uma área da Mata Atlântica do Sudeste brasileiro). Foi investigada a fenologia e os visitantes florais de espécies da família Bromeliaceae em uma área de Mata Atlântica, no Parque Estadual Intervales, no Sudeste brasileiro. Os táxons estudados pertencem aos gêneros Tillandsia L. (3 spp.), Vriesea Lindl. (5 spp.), Aechmea Ruiz & Pav. (3 spp.), Billbergia Thunb. (1 sp.) e Nidularium Lem. (2 spp.). Foram estabelecidas transecções amostrais em áreas com diferentes estádios sucessionais, onde foi registrada a localização dos indivíduos nos estratos da vegetação e realizados censos mensais da fenologia de floração. As bromeliáceas apresentaram padrão de floração seqüencial ao longo do ano, com maior número de espécies floridas na estação chuvosa. Os visitantes florais foram registrados por observações naturalísticas. Oito espécies de beija-flores visitaram as flores das bromélias. Destas, Phaethornis eurynome Lesson e Thalurania glaucopis Gmelim foram os visitantes mais freqüentes. A análise da similaridade dos beija-flores visitantes florais indicou a existência de dois conjuntos de espécies de bromeliáceas: o primeiro polinizado principalmente por espécies da sub-família Trochilinae e outro por Phaethornis eurynome (sub-família Phaethornithinae). A distribuição espacial diversa e, sobretudo os diferentes picos de floração foram os principais fatores que minimizaram a competição por polinizadores entre as espécies de Bromeliaceae estudadas. Palavras-chave - beija-flores, biologia floral, Bromeliaceae, fenologia, Mata Atlântica 1. Universidade Estadual de Feira de Santana, Departamento de Ciências Biológicas, Km 03 - BR 116, 44031-460 Feira de Santana, BA, Brasil. 2. Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil. 3. Autor para correspondência: [email protected]Introdução Bromeliaceae está entre as poucas famílias em que a polinização por vertebrados predomina sobre a polinização por insetos (Sazima et al. 1989). Sick (1984) sugere que a diversificação das famílias Bromeliaceae e Trochilidae tenha se processado paralelamente, pois esta última contém os agentes polinizadores mais importantes para as bromélias. Essa idéia tem sido corroborada por diversos estudos que apontam os beija- flores como vetores de pólen de cerca de 85% das bromeliáceas nas diferentes comunidades investigadas, sendo as demais espécies visitadas por morcegos e, em menor quantidade, abelhas (Snow & Snow 1980, 1986, Snow & Teixeira 1982, Gardner 1986, Fischer & Araujo 1995, Sazima et al. 1995, 1996, Canela & Sazima 2003, Araujo et al. 2004). A fenologia e polinização de Bromeliaceae têm sido relativamente pouco investigadas. O padrão fenológico de uma comunidade vegetal é relevante no estudo da interação planta-animal, pois propicia importante ferramenta para o entendimento da reprodução das plantas e da organização espaço-temporal dos recursos disponíveis no ambiente aos animais associados
Transcript
Revista Brasil. Bot., V.29, n.1, p.163-174, jan.-mar. 2006
Fenologia da floração e biologia floral de bromeliáceas ornitófilas de umaárea da Mata Atlântica do Sudeste brasileiro
CAIO GRACO MACHADO1,3 e JOÃO SEMIR2
(recebido: 7 de outubro de 2004; aceito: 2 de fevereiro de 2006)
ABSTRACT – (Flowering phenology and floral biology of some ornitophilous Bromeliaceae of an Atlantic forest area insoutheastern Brazil). This study dealt with phenology and flower visitors of some Bromeliaceae species in an Atlantic Forestarea at Parque Estadual Intervales, southeastern Brazil. The taxa studied belonged to the genera Tillandsia L. (3 spp.), VrieseaLindl. (5 spp.), Aechmea Ruiz & Pav. (3 spp.), Billbergia Thunb. (1 sp.) and Nidularium Lem. (2 spp.). Sample transects wereestablished in areas with different succession stages, where the bromeliads were localized and marked. Flowering phenologysurveys were made monthly. The bromeliad community showed a sequential flowering all along the year and most speciesflowered during the rainy season. The floral visitors were recorded by naturalistic observations. Eight hummingbird speciesvisited the bromeliads flowers. Among them, Phaethornis eurynome Lesson and Thalurania glaucopis Gmelin were the mostfrequent visitors. Bromeliads were divided in two groups by similarity analysis of floral visitors (hummingbirds): a grouprepresented by species visited by Trochilinae hummingbirds and another visited by Phaethornis eurynome (subfamilyPhaethornithinae). The competition among bromeliad species for pollinators is reduced due to their spatial distribution and theflowering periods.
Key words - Atlantic Forest, Bromeliaceae, floral biology, hummingbirds, phenology
RESUMO – (Fenologia de floração e biologia floral de bromeliáceas ornitófilas de uma área da Mata Atlântica do Sudestebrasileiro). Foi investigada a fenologia e os visitantes florais de espécies da família Bromeliaceae em uma área de MataAtlântica, no Parque Estadual Intervales, no Sudeste brasileiro. Os táxons estudados pertencem aos gêneros Tillandsia L. (3spp.), Vriesea Lindl. (5 spp.), Aechmea Ruiz & Pav. (3 spp.), Billbergia Thunb. (1 sp.) e Nidularium Lem. (2 spp.). Foramestabelecidas transecções amostrais em áreas com diferentes estádios sucessionais, onde foi registrada a localização dosindivíduos nos estratos da vegetação e realizados censos mensais da fenologia de floração. As bromeliáceas apresentarampadrão de floração seqüencial ao longo do ano, com maior número de espécies floridas na estação chuvosa. Os visitantesflorais foram registrados por observações naturalísticas. Oito espécies de beija-flores visitaram as flores das bromélias. Destas,Phaethornis eurynome Lesson e Thalurania glaucopis Gmelim foram os visitantes mais freqüentes. A análise da similaridadedos beija-flores visitantes florais indicou a existência de dois conjuntos de espécies de bromeliáceas: o primeiro polinizadoprincipalmente por espécies da sub-família Trochilinae e outro por Phaethornis eurynome (sub-família Phaethornithinae). Adistribuição espacial diversa e, sobretudo os diferentes picos de floração foram os principais fatores que minimizaram acompetição por polinizadores entre as espécies de Bromeliaceae estudadas.
Palavras-chave - beija-flores, biologia floral, Bromeliaceae, fenologia, Mata Atlântica
1. Universidade Estadual de Feira de Santana, Departamento deCiências Biológicas, Km 03 - BR 116, 44031-460 Feira deSantana, BA, Brasil.
2. Universidade Estadual de Campinas, Departamento deBotânica, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil.
Bromeliaceae está entre as poucas famílias em quea polinização por vertebrados predomina sobre apolinização por insetos (Sazima et al. 1989). Sick (1984)sugere que a diversificação das famílias Bromeliaceaee Trochilidae tenha se processado paralelamente, poisesta última contém os agentes polinizadores mais
importantes para as bromélias. Essa idéia tem sidocorroborada por diversos estudos que apontam os beija-flores como vetores de pólen de cerca de 85% dasbromeliáceas nas diferentes comunidades investigadas,sendo as demais espécies visitadas por morcegos e, emmenor quantidade, abelhas (Snow & Snow 1980, 1986,Snow & Teixeira 1982, Gardner 1986, Fischer & Araujo1995, Sazima et al. 1995, 1996, Canela & Sazima 2003,Araujo et al. 2004).
A fenologia e polinização de Bromeliaceae têm sidorelativamente pouco investigadas. O padrão fenológicode uma comunidade vegetal é relevante no estudo dainteração planta-animal, pois propicia importanteferramenta para o entendimento da reprodução dasplantas e da organização espaço-temporal dos recursosdisponíveis no ambiente aos animais associados
C.G. Machado & J. Semir: Fenologia e biologia floral de bromeliáceas ornitófilas164
(Morellato & Leitão Filho 1992, Talora & Morellato2000). A floração seqüencial das bromeliáceas em umaregião pode ser de extrema importância para amanutenção dos agentes polinizadores na área,contribuindo para a eficiência no sistema de polinizaçãode espécies ornitófilas da comunidade (Waser & Real1979, Feinsinger 1983, Araujo et al. 1994, Fischer &Araujo 1995).
Este estudo objetivou investigar o padrão fenológicode floração e atributos florais de bromélias ornitófilasem uma área da Mata Atlântica de região serrana e autilização diferencial destes recursos pelas espécies debeija-flores.
Material e métodos
Este estudo foi realizado no Parque Estadual Intervales(PEI), em Ribeirão Grande, SP, Brasil. O PEI situa-se a 24º12’- 24º25’ S e 48º03’ - 48º30’ W, com área de 49.000 ha. Osregistros foram feitos na região da sede administrativa doPEI, a 950 m de altitude, de setembro de 1994 a março de1996, com 19 visitas mensais de oito a 20 dias cada. O relevona área é do tipo “montanha com vales profundos” (Ponçano1981). A vegetação é típica da zona ecotonal dos domíniosTropical Atlântico e dos Planaltos de Araucária. Devido aorelevo e aos estádios sucessionais diversos, existemdiferenças entre as florestas que ocorrem nos váriosambientes da região. Para maior detalhamento sobre avegetação local veja Nascimento (1994). O clima éclassificado como Cfb (Köepen 1948). A precipitação anualé entorno de 1.800 mm, com pequenas diferenças sazonais,ocorrendo uma estação chuvosa, de outubro a março, eoutra subseca, de abril a setembro. A temperatura médiaanual é de 22 ºC, com mínima registrada de 0 ºC e máxima de32 ºC (Machado 1999).
Foram estabelecidos sete transectos (com extensões de1.500 m, 600 m, 600 m, 2.400 m, 600 m, 1.500 m e 800 m,totalizando 8.000 m), onde foram amostradas áreas deambientes distintos, determinados pelo relevo (encostas evales). A área de amostragem (48.000 m2) foi calculada a partirdo comprimento dos transectos multiplicado por suaslarguras, visto que a amostragem era feita até três metros decada lado do eixo dos transectos.
Das espécies de Bromeliaceae ocorrentes na área, foramestudadas as que floresceram no período de estudo econsideradas ornitófilas.
No campo, as bromeliáceas foram identificadas apósformação da inflorescência. Muitas espécies apresentamindivíduos que ocorrem agregados, formando densastouceiras. Nesse caso, como procedeu Fischer (1994), foiconsiderado que cada inflorescência correspondia a umindivíduo.
A determinação da fenofase de floração foi feita atravésde registros mensais, quando foram anotados o número de
indivíduos floridos de cada espécie, o estrato da vegetaçãoem que estavam estabelecidos (solo, sub-bosque e dossel) eo respectivo hábito (terrestre, epifítico ou facultativo).
As espécies de bromélias foram classificadas comoheliófilas quando ocorriam preferencialmente em locais comalto grau de luminosidade, com incidência direta de luz solar,ou esciófilas, quando se desenvolviam em ambientessombreados e de luz difusa (Fischer & Araujo 1995).
A classificação dos padrões fenológicos seguiuNewstrom et al. (1994) e a categorização quanto à síndromede polinização foi feita através da análise dos atributos florais(Faegri & Pijl 1980). Neste estudo, considerou-se como picode floração o período no qual mais que 50% dos indivíduosamostrados de cada espécie estavam floridos.
Durante o período de floração foram registrados (em nomínimo três indivíduos diferentes) o número de flores abertaspor dia, o período de disponibilidade da flor (o período emque a flor permanece aberta, disponível aos visitantes, desdesua antese até a senescência - em algumas espécies as florespodem durar mais que um dia, apresentando fechamento eabertura das flores, uma ou mais vezes, entre o período deantese e senescência), a duração (em dias) das flores, ocomprimento do tubo da corola, o período de deiscência dasanteras com o pólen disponível (Dafni 1992) e o período dereceptividade do estigma (Kaerns & Inouye 1993), além dacoloração do cálice, corola e brácteas. A coleta de néctar edeterminação de seu volume foi feita com uso demicrosseringa ou microcapilares graduados, em florespreviamente isoladas com sacos de papel, em intervalos deduas horas durante todo o período de abertura da flor. Aconcentração de solutos no néctar (açúcares hidrossolúveis)foi determinada com o uso de refratômetro de bolso.
As observações dos beija-flores visitantes foram feitascom auxílio de binóculos 8 X 35 ou à vista desarmada. Assessões foram feitas da aurora ao crepúsculo ou até ofechamento ou a senescência das flores. Para o registro dosbeija-flores visitantes foi adotado o método de observaçãoindivíduo-focal (Altmann 1974), com no mínimo três repetiçõespara cada espécie. A variação do número de horas deobservação em cada espécie decorreu do período dedisponibilidade da flor, número de dias de duração da flor ediferenças nos horários da aurora e crepúsculo nas diferentesestações do ano. Em cada sessão de observação-focal, foramregistradas quais espécies de beija-flor visitavam as floresdas bromeliáceas, o horário e o número de visitas(considerou-se “visita” toda vez que um beija-flor sealimentava do néctar de uma flor) e o local de impregnação depólen nas aves. Todas as ocorrências de agonismo intra ouinterespecífica entre os beija-flores foram registradas. Aidentificação das aves foi feita em campo e a classificaçãoseguiu Sick (1984).
Para determinar a similaridade entre a guilda de beija-flores polinizadores associada às espécies de bromeliáceasestudadas, foi utilizado o Índice de Jaccard (Magurran 1988).Todos os cálculos estatísticos seguiram as determinações
Revista Brasil. Bot., V.29, n.1, p.163-174, jan.-mar. 2006 165
de Sokal & Rohlf (1981), sendo testada a distribuição normaldos dados. No caso do volume e concentração de néctarproduzido foram excluídos os “out liers”, uma vez que, emalgumas flores, a presença de muco ou água de chuvaacumulada, causaram aumento do volume e diluição do néctar.Desta forma, todas as análises foram paramétricas. Utilizou-se a correlação de Spearman para correlacionar comprimentode bico de beija-flor e de tubo da corola, assim comofreqüência de visita de beija-flor e produção de néctar pelaflor. A análise de agrupamento foi feita pelo método dasdistâncias médias entre os grupos (UPGMA) sobre ocoeficiente de distância métrica Euclidiana. Utilizou-se nestasanálises o pacote estatístico SPSS – Statistical Package forSocial Sciences (SPSS 1983).
Resultados
Quatorze espécies de bromeliáceas ornitófilasfloresceram durante este estudo (tabela 1), sendo cincopertencentes à sub-família Bromelioideae (gênerosNidularium, Billbergia e Aechmea) e as demais à sub-família Tillandsioideae (Vriesea e Tillandsia). Outrasespécies de bromélias ornitófilas podem ocorrer na áreade estudo, como Vriesea vagans L.B. Smith, Vriesea
friburguensis Mez e Canistrum lindenii (Regel) Mez,porém não floresceram durante o período de coletas dedados. Apesar de Vriesea philippocoburgii não terocorrido dentro da área amostral, foram registradosdados sobre sua fenologia, biologia floral e sobre os seusvisitantes florais, pois é uma espécie comum na região.
Das espécies de bromélias estudadas, nenhumaapresentou hábito exclusivamente terrestre (tabela 1) emesmo as de hábito facultativo tenderam ao epifitismo,com raros indivíduos crescendo junto ao solo, muitasvezes sobreviventes de quedas de galhos de estratosmais altos da vegetação, nos quais estavamestabelecidas. Nidularium rubens foi a espécie maisfreqüentemente observada estabelecendo-sediretamente sobre o solo.
Nove espécies foram classificadas como heliófilas(tabela 1) e ocorreram preferencialmente nos estratosmais altos da vegetação ou, quando em estratos maisbaixos, em locais mais ensolarados, como clareiras ebordas de mata. As espécies esciófilas ocorreram nosestratos mais baixos e dentro do sub-bosque; quandopresentes no dossel, ocorriam dentro das copas densas,que proporcionavam ambiente sombreado.
Tabela 1. Bromeliáceas ornitófilas estudadas na região da sede do Parque Estadual Intervales, Ribeirão Grande, SP, seushábitos, tolerância à luminosidade e as espécies de beija-flores que visitaram suas flores (TG = Thalurania glaucopis Gmelin;CR = Clytolaema rubricauda Boddaert; PE = Phaethornis eurynome Lesson; AV = Amazilia versicolor Vieillot;CA = Chlorostilbon aureoventris Orbigny & Lafresnaye; LA = Leucochloris albicollis Vieillot; AN = Anthracothorax nigricollisVieillot; MF = Melanotrochilus fuscus Vieillot).
Table 1. Ornitophilous species of Bromeliceae at the Parque Estadual Intervales (Ribeirão Grande, SP, Brasil), their habits,tolerance to light and visitor humminbirds (TG = Thalurania glaucopis Gmelin; CR = Clytolaema rubricauda Boddaert;PE = Phaethornis eurynome Lesson; AV = Amazilia versicolor Vieillot; CA = Chlorostilbon aureoventris Orbigny & Lafresnaye;LA = Leucochloris albicollis Vieillot; AN = Anthracothorax nigricollis Vieillot; MF = Melanotrochilus fuscus Vieillot).
Espécies Hábito Tolerância à luminosidade Espécies de beija-floresvisitantes florais
C.G. Machado & J. Semir: Fenologia e biologia floral de bromeliáceas ornitófilas166
A comunidade de bromélias apresentou floraçãoseqüencial e contínua (figura 1). Todas as espéciesapresentaram padrão anual de floração e, quanto àduração da fenofase de floração, a maioria foi do tipocurta (figura 1). Foi verificado em Aechmeaorganensis, Vriesea incurvata e V. philippocoburgiique o período de floração foi mais extenso; nas duasprimeiras espécies, a fenofase de floração individual écurta, porém os diferentes indivíduos vão se sucedendoem floração, enquanto em V. philippocoburgii afloração individual é extensa, com grande quantidadede flores por indivíduo (0 = 146,78 ± 13,46; n = 10), mascom um número baixo delas abertas por dia(0 = 4,40 ± 2,03; n = 10).
O maior número de espécies (n = 10) floresceu naestação chuvosa, entre outubro e março (figura 1). Onúmero total de indivíduos floridos nos transectosamostrais variou durante o ano (figura 2), sendosignificativamente menor durante a estação subseca(P ≤ 0,05). Os picos com maior número de indivíduosfloridos nos meses de junho, agosto e setembro foramconseqüência do período de pico de floração deV. carinata (junho) e T. stricta (agosto e setembro),espécies abundantes na área de estudo.
Tillandsia stricta e T. geminiflora apresentaramflores que perduraram por três dias enquantoT. tenuifolia, Nidularium rubens e N. innocentii, floresque duraram dois dias. Nas Tillandsia ocorreu ofechamento da flor no final do dia e abertura na manhãseguinte. Em Nidularium as flores permaneceram como ápice das pétalas fechado, não expondo anteras eestigma. As demais espécies estudadas apresentaramflores com duração de apenas um dia (tabela 2). Todasas bromélias estudadas apresentaram antese diurna, porvolta 4 e 6 h da manhã e senescência entre 17 e 20 h(tabela 2). No caso de Nidularium, considerou-se comoperíodo de disponibilidade o período de produção denéctar do início da manhã, por volta das 05 h e 30 min.,quando as anteras já estavam abertas e os grãos depólen expostos e facilmente removíveis, até por voltadas 12 h (tabela 2).
Nas espécies estudadas, o volume médio de néctarfoi 132,0 ± 138,7 mL por flor. O alto desvio padrãoocorreu em algumas espécies que secretaram néctarem grande quantidade, como Vriesea altodaserrae eV. incurvata (tabela 2). A concentração do néctarvariou de 20% a 30% (tabela 2), com média de25,7 ± 3,8% (n = 14). Foi significativa a correlação entre
Figura 1. Fenologia de floração de 14 espécies de bromeliáceas que ocorrem na região da sede do Parque Estadual Intervales,Ribeirão Grande, SP, entre abril de 1995 e março de 1996. A linha simples representa o período total de cada florada e a maisgrossa, o pico de floração.
Figure 1. Floral phenology of 14 Bromeliaceae species of the Parque Estadual Intervales (Ribeirão Grande, SP, Brasil), April/1995-March/1996. (Fine line = flowering period; thick line = flowering peak).
Estação subseca Estação chuvosa
Meses A M J J A S O N D J F M
Billbergia amoena
Vriesea carinata
Vriesea philippocoburgiiTillandsia stricta
Tillandsia geminiflora
Aechmea organensis
Vriesea altodaserrae
Aechmea nudicaulis
Vriesea incurvata
Nidularium innocentii
Nidularium rubens
Vriesea flammea
Tillandsia tenuifoliaAechmea ornata
Revista Brasil. Bot., V.29, n.1, p.163-174, jan.-mar. 2006 167
0
50
100
150
J F M A M J J A S O N D
comprimento do tubo e volume de néctar produzido nasespécies estudadas (r = 0,51; P ≤ 0,05; n = 14). Acorrelação entre tamanho do tubo da corola econcentração de néctar das bromélias foi positiva, masnão significativa (r = 0,21; P ≥ 0,05; n = 14).
Os tubos das corolas de Nidularium rubens eN. innocentii são brancos e longos. As flores sedesenvolvem dentro da roseta, em posição terminal ouaxial, imersas na água acumulada. O contraste com overmelho das folhas mais internas as tornam conspícuas.O estigma se tornou receptivo duas horas após o iníciode produção de néctar da flor e permaneceu receptivoaté a sua senescência, no segundo dia de duração daflor.
Aechmea ornata e A. organensis têm floresvioleta a lilás, enquanto em A. nudicaulis estas sãoamarelas. Em A. ornata, as flores contrastam com asbrácteas do escapo, que são grandes e vermelhas. EmA. organensis há contraste com escapo e brácteas dospedúnculos, de coloração vinácea. Por sua vez, ocontraste das flores amarelas de Aechmea nudicaulisse dá com as grandes brácteas vermelhas do escapo.Em todas as espécies, a liberação do pólen das anterasocorreu logo após a antese, enquanto o estigma setornou receptivo em torno de três horas depois.
Billbergia amoena tem corola amarela,ligeiramente translúcida, com o ápice esverdeado,contrastando com grandes brácteas vermelhas. Cadainflorescência produziu cinco ou seis flores e metadedelas abriu no mesmo dia e as restantes, dois dias depois.Verificou-se que em algumas flores (n = 15) de diversasinflorescências (n = 10), não houve a produção de
néctar, apesar dos nectários desenvolvidos próximo aoovário. Os grãos de pólen amadureceram após a anteseda flor, se desprendendo facilmente das anteras,enquanto o estigma, assim como nas espéciescongêneres, se tornou receptivo cerca de três horas apósa antese.
Tillandsia stricta, T. geminiflora e T. tenuifoliapossuem flores relativamente pequenas, azul-arroxeadasnas duas primeiras e brancas na última, todas combrácteas rosadas. Nessas espécies as anteras abriram-se logo após a antese, liberando grãos de pólen, mas areceptividade do estigma foi detectada no segundo dia,após sua abertura e no caso de Tillandsia stricta eT. geminiflora, também no terceiro dia de abertura daflor.
As corolas das flores das espécies de Vriesea sãode coloração amarelo intenso, contrastando com overmelho das sépalas, brácteas e escapo, com exceçãode V. flammea, cuja corola branca é conspícua pelocontraste do cálice, brácteas e escapo de coloraçãovermelho-vinácea. As flores de Vriesea secretaramnéctar com concentração média de 24%. Em todas elas,as anteras se abriram cerca de uma hora após a antese,liberando os grãos de pólen. O estigma se tornoureceptivo, em todas as espécies, logo após a antese daflor.
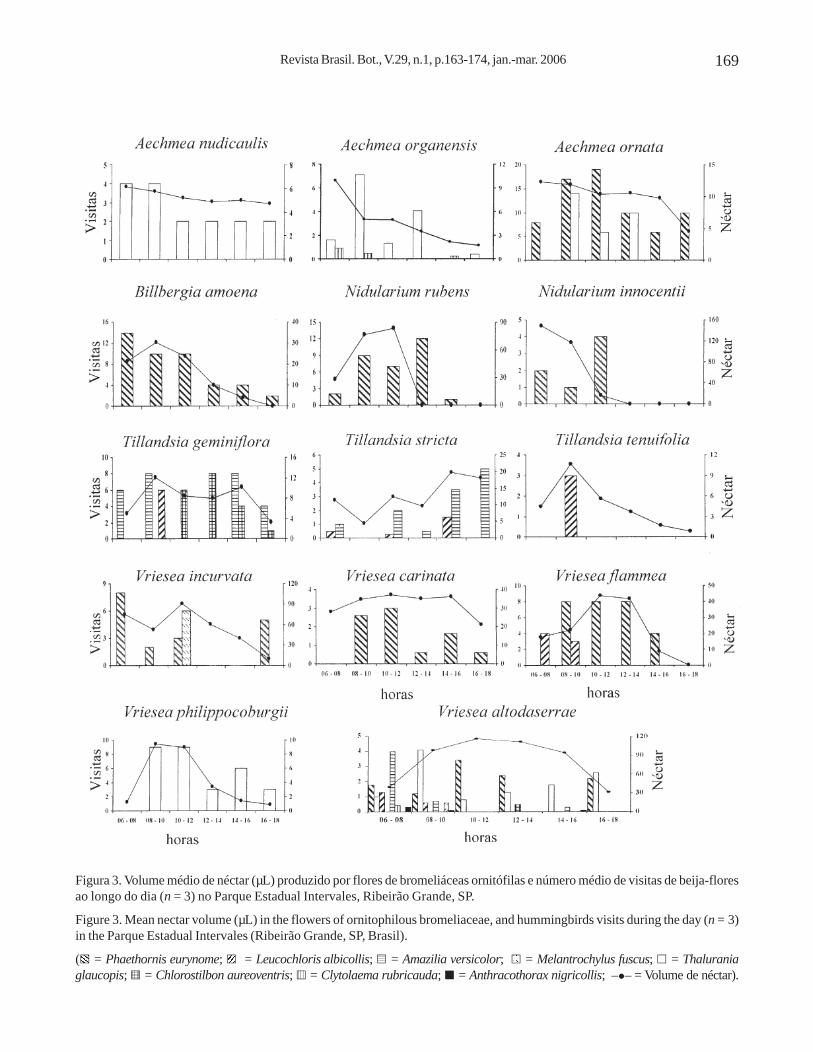
Oito espécies de bromeliáceas apresentaram umaprodução maior de néctar no período da manhã(figura 3). Aechmea nudicaulis, Vriesea carinata eTillandsia geminiflora produziram néctar maisuniformemente ao longo do dia, sendo que a últimaapresentou dois picos de maior produção, um pela manhãe outro pela tarde. Tillandsia stricta teve uma maiorprodução no período da tarde, enquanto Vrieseaflammea e V. altodaserrae produziram mais néctar das10 às 14 h (figura 3).
Quanto à correlação entre produção de néctar e àfreqüência de visitas de beija-flores ao longo do dia,Aechmea nudicaulis, Billbergia amoena, Tillandsiageminiflora, T. stricta, T. tenuifolia, Vriesea carinata,V. incurvata, V. flammea e V. philippocoburgiiapresentaram correlação positiva e estatisticamentesignificativa (P ≤ 0,05) (respectivamente: r = 0,92;r = 0,85; r = 0,88; r = 0,96; r = 0,86; r = 0,64; r = 0,46;r = 0,73 e r = 0,83). Aechmea organensis, A. ornata,Nidularium innocentii e N. rubens apresentaramcorrelação positiva, porém não significativa (P ≥ 0,05)(respectivamente: r = 0,39; r = 0,35; r = 0,37 e r = 0,27);por sua vez, V. altodaserrae apresentou correlaçãonegativa não significativa (r = -0,40; P ≥ 0,05).
Oito espécies de beija-flores foram registradas
Meses
Indi
vídu
os e
m fl
or (N
)
Figura 2. Número total de indivíduos de bromélias floridos,registrados mensalmente nos transectos amostraisestabelecidos na região da sede administrativa do ParqueEstadual Intervales, Ribeirão Grande, SP, durante 1995.
Figure 2. Number of flowering individuals of Bromeliaceaerecorded monthly in the sampled transect of the ParqueEstadual Intervales (Ribeirão Grande, SP, Brasil), in 1995.
C.G. Machado & J. Semir: Fenologia e biologia floral de bromeliáceas ornitófilas168
visitando as flores das bromeliáceas (tabela 1, figura 3).Desses, apenas cinco delas ocorreram na região aolongo do ano todo (Phaethornis eurynome, Thaluraniaglaucopis, Amazilia versicolor, Leucochlorisalbicollis e Chlorostilbon aureoventris). As outrastrês espécies estiveram ausentes em toda a estação sub-seca ou em grande parte dela, como Anthracothoraxnigricollis, presente de outubro a março,Melanotrochilus fuscus, presente de outubro até abrile Clytolaema rubricauda, de outubro a maio.
Todas as espécies efetuaram visitas legítimas pelafrente e tocando as partes reprodutivas das flores, nãotendo sido observadas tentativas de perfuração da baseda flor para pilhar néctar. A correlação entrecomprimento dos bicos de beija-flores e comprimentodos tubos das corolas das bromeliáceas que visitam nãofoi significativa (r = 0,41; P ≥ 0,05; n = 29).
As únicas espécies de beija-flores que visitarammais de cinco espécies de bromélias foram Phaethorniseurynome, Thalurania glaucopis e Leucochlorisalbicollis (tabela 1). Phaethornis eurynome visitouoito espécies, sendo que em quatro delas foi único vetorde pólen (Billbergia amoena, Nidularium rubens,N. innocentii e Vriesea carinata); as demais espéciesforam visitadas também por outros troquilídeos, porémnestas, P. eurynome foi o visitante mais freqüente(V. flammea, V. incurvata e A. ornata) ou o segundomais freqüente (V. altodaserrae). Esse beija-flor tendeua forragear nas espécies do sub-bosque e na porçãoinferior das copas (Billbergia amoena). Na maioria dasespécies de bromélias que explorou as visitas às floresforam em intervalos de 40 minutos a uma hora. Exceçãoocorreu durante a floração de Vriesea altodaserrae,quando permaneceu próximo à área de observação,entre as 10 e 14 h, período em que se verificou nesta aprodução de maior volume de néctar (figura 3). O localde deposição de pólen das bromélias visitadas emPhaethornis eurynome foi o bico, sendo que apenas opólen de Billbergia amoena foi depositado na fronte.
Thalurania glaucopis exibiu comportamentoterritorialista, permanecendo próxima a área onde haviaplantas floridas. Visitou cinco espécies de bromélias(tabela 1), sendo o único visitante de Aechmeanudicaulis e Vriesea philippocoburgii. Foi também aespécie com maior número de visitas em Aechmeaorganensis e Vriesea altodaserrae. Nessa última, opólen foi depositado na fronte da ave, porém o localcom maior freqüência de impregnação foi o bico, emboraem Vriesea philippocoburgii o pólen também foidepositado no vértice da ave. Esse troquilídeo ocorreutanto em áreas mais abertas como no interior da mata,Ta
bela
2. D
ados
sob
re a
bio
logi
a fl
oral
de
brom
eliá
ceas
que
oco
rrem
na
Mat
a A
tlân
tica
da
regi
ão d
a se
de d
o P
arqu
e E
stad
ual I
nter
vale
s, R
ibei
rão
Gra
nde,
SP,
Bra
sil.
Os
dado
s fo
ram
col
etad
os e
ntre
set
embr
o de
199
4 e
feve
reir
o de
199
6. (s
= d
esvi
o pa
drão
).
Tabl
e 2.
Flo
ral b
iolo
gy d
ata
for t
he B
rom
elia
ceae
spec
ies o
f the
Atla
ntic
fore
st o
f the
Par
que
Est
adua
l Int
erva
les (
Rib
eirã
o G
rand
e, S
P, B
rasi
l). S
epte
mbe
r/19
94-F
ebru
ary/
1996
.(s
= s
tand
ard
devi
atio
n).
Esp
écie
sC
once
ntra
ção
doV
olum
e di
ário
de
Per
íodo
de
disp
onib
ilid
ade
Dia
s de
Núm
ero
de fl
ores
Com
prim
ento
do
Hor
as d
ené
ctar
± s
%né
ctar
± s
µL
diár
io d
a fl
ordu
raçã
o da
flo
r a
bert
as p
or d
iatu
bo fl
oral
(mm
)ob
serv
ação
foc
al
Aec
hmea
nud
icau
lis
29,2
±0,
8 (6
)25
,1±
3,8
(6)
06:0
0 - 1
8:15
11
- 312
,1 +
0,1
48A
echm
ea o
rgan
ensi
s28
,0±
0,6
(6)
31,5
±3,
5 (6
)04
:00
- 17:
301
2 - 4
14,5
+ 0
,260
Aec
hmea
orn
ata
28,7
±1,
0 (5
)64
,2±
9,5
(5)
06:3
0 - 1
8:00
12
- 520
,3 +
0,1
60B
illb
ergi
a am
oena
27,3
±3,
5 (5
)81
,0±
5,0
(5)
04:3
0 - 1
7:30
12
- 340
,7 +
0,3
44N
idul
ariu
m i
nnoc
enti
i 2
9,7
±1,
1 (1
2) 1
49,4
±10
,6 (1
2)05
:30
- 12:
002
2 - 5
43,5
+ 0
,540
Nid
ular
ium
rub
ens
30,7
±1,
4 (9
)18
8,8
±14
,7 (9
)05
:30
- 13:
002
2 - 3
36,4
+ 1
,648
Till
ands
ia g
emin
iflo
ra21
,1±
1,5
(6)
48,8
±3,
9 (6
)06
:00
- 17:
303
2 - 6
17,9
+ 0
,260
Till
ands
ia s
tric
ta 2
0,9
±1,
5 (8
) 7
2,2
±10
,3 (8
)15
:00
- 18:
003
2 - 4
18,3
+ 0
,960
Till
ands
ia t
enui
foli
a20
,5±
0,6
(5)
30,7
±4,
8 (5
)05
:00
- 19:
002
1 - 3
17,0
+ 0
,960
Vrie
sea
alto
dase
rrae
30,0
±0,
6 (9
)49
0,5
±28
,7 (9
)04
:30
- 21:
001
1 - 3
31,3
+ 0
,548
Vrie
sea
cari
nata
26,2
±1,
5 (7
)21
4,9
±14
,5 (7
)07
:30
- 17:
001
1 -2
39,0
+ 0
,344
Vrie
sea
flam
mea
21,
5±
1,9
(13)
134
,5±
17,9
(13)
06:1
5 - 2
0:00
11
- 237
,0 +
0,1
45Vr
iese
a in
curv
ata
24,
5±
2,3
(17)
340
,1±
16,5
(17)
06:3
0 - 1
9:00
11
- 243
,2 +
0,9
44Vr
iese
a ph
ilip
poco
burg
ii 2
2,2
±1,
1 (1
0)26
,8±
2,5
(10)
04:0
0 - 1
9:00
1 3
- 8
28,7
+ 1
,148
Revista Brasil. Bot., V.29, n.1, p.163-174, jan.-mar. 2006 169
Figura 3. Volume médio de néctar (µL) produzido por flores de bromeliáceas ornitófilas e número médio de visitas de beija-floresao longo do dia (n = 3) no Parque Estadual Intervales, Ribeirão Grande, SP.
Figure 3. Mean nectar volume (µL) in the flowers of ornitophilous bromeliaceae, and hummingbirds visits during the day (n = 3)in the Parque Estadual Intervales (Ribeirão Grande, SP, Brasil).
C.G. Machado & J. Semir: Fenologia e biologia floral de bromeliáceas ornitófilas170
onde foi mais freqüente.Leucochloris albicollis foi observada mais
freqüentemente em áreas abertas, capoeiras e matasde vegetação secundária recente e antiga, porémtambém pôde ser avistado em área de mata primária,onde forrageou em estratos mais altos da vegetação.Visitou Vriesea altodaserrae, V. flammea e todas astrês espécies de Tillandsia estudadas, sendo o visitanteexclusivo de T. tenuifolia (tabela 1). Todas essasespécies tiveram seu pólen depositado no bico deLeucochloris albicollis.
Amazilia versicolor, espécie típica de áreasabertas, foi observada em capoeiras e áreas de matasecundária, sendo a principal visitante de Tillandsiastricta e T. geminiflora. Exibiu comportamentoagonístico, expulsando outros visitantes, principalmentedurante os períodos de maior produção de néctar emTillandsia.
A partir do agrupamento (“cluster”), elaborado comos valores dos índices de similaridade (Jaccard) de usodas espécies de bromeliáceas pelas diferentes espéciesde beija-flores, observou-se dois grandes conjuntos deespécies de bromélias (figura 4), sendo o primeirocomposto por dois sub-conjuntos: o primeiro (Vrieseaphilippocoburgii, Aechmea nudicaulis eA. organensis), cujo único ou principal visitante floralfoi Thalurania glaucopis e o segundo (Tillandsiageminiflora, T. stricta, T. tenuifolia eV. altodaserrae), cujas espécies foram visitadasprincipalmente por Amazilia versicolor, Leucochlorisalbicollis, Chlorostilbon aureoventris (tabela 1). Osegundo conjunto de bromélias foi formado pelasespécies que utilizam Phaethornis eurynome como seuúnico vetor de pólen (Nidularium rubens, N. innocentii,Vriesea carinata e Billbergia amoena) ou como o maisfreqüente agente polinizador (Vriesea incurvata,V. flammea e Aechmea ornata) (tabela 1, figura 4).
Discussão
Em estudos realizados com bromeliáceas ornitófilasem diferentes áreas de Mata Atlântica da planícielitorânea no Estado de São Paulo foram registradas 22espécies em Caraguatatuba (Buzato et al. 2000), 15espécies em Picinguaba (Araujo 1996) e 24 espécies naJuréia (Araujo et al. 2004); na Serra do Mar, ainda noEstado de São Paulo, Buzato et al. (2000) registraram12 espécies no município de Cunha, com altitude decerca de 1.000 m, e sete espécies em Campos do Jordão(ca. de 1.500 m de altitude), de bromélias ornitófilas.Estes dados corroboram Snow & Snow (1986), que
sugerem que a maior diversidade e riqueza de espéciesde bromeliáceas ocorrem em áreas de pouca altitude.Benzing (1980) e Reitz (1983) consideram que as baixastemperaturas de inverno, que ocorrem em regiões dealtitude maior, podem ser um fator limitante para oestabelecimento de várias espécies de bromélias.
Na comunidade estudada, o padrão de floração dacomunidade de bromélias ornitófilas foi sequencial econtínuo. Estudos realizados na Mata Atlântica doSudeste brasileiro têm constatado que as bromeliáceasformam o maior grupo de plantas ornitófilas (em riquezade espécies) na comunidade e que também apresentamo mesmo padrão de floração (Araujo et al. 1994, Buzatoet al. 2000). A disponibilidade de flores ornitófilas aolongo do ano possibilita a permanência beija-florespolinizadores na área. Isso propicia uma maximizaçãodo sucesso reprodutivo das espécies de plantasenvolvidas, pois caso haja deslocamentos populacionaisdestas aves, devido a uma pausa na oferta de recursos,as primeiras espécies de plantas ornitófilas que floriremnão encontrarão disponíveis seus agentes polinizadores(Poulin et al. 1992).
Distância Euclidiana
Figura 4. Agrupamento das espécies de bromélias ornitófilasdo Parque Estadual Intervales (Ribeirão Grande, SP, Brasil),pela similaridade das espécies de beija-flores que as visitam(I – principalmente visitadas por Trochilinae; II –predominantemente visitadas por Phaethornitinae.
Figure 4. Ornitophilous bromeliad groups in the ParqueEstadual Intervales (Ribeirão Grande, SP, Brasil), accordingto similarity of hummingbird visitors (I – mainly visited byTrochilinae visitors; II – mainly visited by Phaethornitinae).
25 20 15 10 5 0
V. philippocoburgii
A. nidicaulis
A. organensis
T. geminiflora
T. stricta
T. tenuifolia
V. altodaserrae
N. rubens
V. carinata
B. amoena
N. innocentii
V. incurvata
V. flammea
A. ornata
I
II
Revista Brasil. Bot., V.29, n.1, p.163-174, jan.-mar. 2006 171
Dessa forma, as bromeliáceas devem serconsideradas fundamentais para a manutenção local dafauna de beija-flores residentes na Mata Atlântica,beneficiando não somente estas aves como também aspróprias bromeliáceas ornitófilas e outras espécies dacomunidade que utilizam estas aves como seus vetoresde pólen.
Neste estudo, cerca de 70% das espécies debromélias floresceram durante a estação chuvosa,período no qual também foi verificado maior número deindivíduos floridos. Fischer & Araujo (1995) observaramproporção semelhante neste período em região estuarinado Estado de São Paulo, assim como Sazima et al.(1995) observaram em todas as comunidades devegetais ornitófilos com que trabalharam. Essa tendênciaassemelha-se ao encontrado por Wolf et al. (1976) comflores polinizadas por beija-flores em floresta estacionalda América Central e corrobora o padrão esperado emambientes tropicais, onde os fatores climáticos maisfavoráveis, como temperatura e pluviosidade maiores,resultam em uma alta produtividade primária (Pandey& Singh 1992). A maior disponibilidade de floresornitófilas como recurso na estação chuvosa também écoincidente com o período de reprodução da maioria deespécies de beija-flores (Sick 1984), o que demonstra aestreita inter-relação entre estes organismos.
Vriesea carinata apresentou pico de floraçãodurante o mês de junho. Em outra área da MataAtlântica, Araujo et al. (1994) observaram um períodode floração de V. carinata mais longo, de abril a outubro,com o pico em julho e agosto. Comparado aos dadosregistrados por Fischer (1994), Araujo (1996) e Buzatoet al. (2000), com Bromeliaceae da Mata Atlântica emSão Paulo, verificou-se que Nidularium innocentii,Aechmea nudicaulis, A. organensis, Vrieseaaltodaserrae, V. incurvata, Tillandsia stricta,T. tenuifolia e T. geminiflora apresentaram, no PEI,período e pico de floração diferentes das áreasestudadas por aqueles autores. Essas diferenças quantoao início e extensão do período de floração das mesmasespécies em áreas diversas indicam que fatoresambientais são mais importantes para a fenologia destasespécies que fatores intrínsecos, como relaçõesfilogenéticas e história evolutiva do grupo (Kochmer &Handel 1986).
Todas essas espécies apresentaram atributos dasíndrome da ornitofilia (Faegri & Pijl 1980), como:inflorescências expostas, flores tubulosas, antese diurna,cores conspícuas ou contrastantes, néctar comconcentração entre 20% e 32%, inodoras, estigma eanteras afastadas dos nectários e posicionados para o
contato com as aves e ausência de plataforma de pouso.Feinsinger (1983) e Fischer (1994) encontraram
alta correlação positiva entre tamanho de corola econcentração de solutos (açúcares) no néctar. Isso podeestimular, em flores de corola longa, a maior freqüênciade beija-flores de bico longo, uma vez que a recompensaé melhor. No entanto, essa correlação entre tamanhodo tubo da corola e concentração de néctar, ainda quepositiva, não foi significativa nas bromélias ornitófilasdeste estudo. A correlação positiva entre o tamanho dotubo da corola com o volume de néctar produzido nasespécies aqui estudadas corrobora a idéia de que florescom corolas de tubo longo selecionam visitantes de bicomais longo conferindo-lhes melhor recompensa(Arizmendi & Ornelas 1990).
Espera-se que haja alta correlação entre ocomprimento dos bicos dos beija-flores e o comprimentoda corola das flores que visitam, pois corolas longastendem a impedir que visitantes de bico curto alcancemo néctar contido na câmara nectarífera no fundo do tubo,selecionando, desta forma, aves com bico mais longo(Fenster 1991). Neste estudo, esta correlação não foisignificativa. Este resultado provavelmente foiinfluenciado pelo fato de que espécies de bico longotambém visitaram espécies de bromélias de corola curta.
Billbergia amoena pode apresentar mimetismointrapopulacional, pois apenas algumas flores em cadaindivíduo produzem néctar. Nesse caso, flores semnéctar seriam visitadas por beija-flores que poderiamconfundi-las com aquelas que efetivamente produzemnéctar (modelos). Espécies com flores em padrões auto-miméticos também foram descritas em regiõestemperadas (Brown & Kodric-Brown 1979).
Entre os beija-flores, apenas um representante dasub-família Phaethornithinae foi registrado (Phaethorniseurynome), enquanto as demais espécies pertencem àsub-família Trochilinae. Esses dados, assim como os deBuzato et al. (2000), corroboram Snow & Snow (1986),que sugeriram que os trochilíneos são mais abundantesem altitudes mais elevadas.
O longo período entre as visitas de Phaethorniseurynome às flores, somada ao fato desta ave não terpermanecido próxima às flores que visitava, sugere queeste apresente técnica de forrageamento do tipo “linhade captura” ou seja, forrageia efetuando um longo roteirode visitas, não demarcando território definido. Estaestratégia de forrageamento é típica das espécies dasub-família Phaethornithinae (Gottsberger 1986, TieboutIII 1993). No entanto, durante a floração de Vrieseaaltodaserrae, Phaethornis eurynome apresentouestratégia de forrageamento do tipo territorialista durante
C.G. Machado & J. Semir: Fenologia e biologia floral de bromeliáceas ornitófilas172
o período de maior produção de nectar (figura 3),embora não tenha apresentado nenhuma interaçãoagonística interespecífica. Essa alteração temporária decomportamento tem sido reportada em outros estudos(Stiles 1978, Feisinger & Colwell 1978, Canela & Sazima2003). Provavelmente, a alta densidade de flores queproduziam grande volume de néctar e a ausência deoutras espécies competidoras influenciou essecomportamento em Phaethornis eurynome duranteaquele período.
Em relação ao agrupamento das espécies debromélias quanto à similaridade de seus visitantes florais,no primeiro conjunto de bromélias foram mais comunsos beija-flores da sub-família Trochilinae como agentespolinizadores (figura 4, conjunto I). Entre as espéciespolinizadas exclusivamente por Thalurania glaucopis,ou que tenham tido este troquilídeo como o visitantemais freqüente, foi observado que seus picos de floraçãonão foram coincidentes (figura 1), o que pode evitarcompetição pelos beija-flores, diminuindo a probabilidadede mistura de pólen de diferentes espécies no corpodas aves, o que reduziria a transferência correta depólen.
Todas as espécies de bromeliáceas do conjunto Isão heliófilas e a predominância de visitas de beija-florestroquilíneos foi esperada, pois as espécies dessa sub-família, ao contrário de Phaethornis eurynome,forrageiam em locais mais abertos (Piratelli 1997).
Tillandsia geminiflora e T. stricta apresentaramalto grau de similaridade, ou seja, partilharam quase osmesmos agentes polinizadores, sendo Amaziliaversicolor o mais freqüente deles. São plantas quepodem ocorrer em ambientes semelhantes, porém seusperíodos de pico de floração também não foramcoincidentes. A relativa semelhança entre essas duasplantas (flores azuis arroxeadas e brácteas rosadas)pode ainda reforçar nos beija-flores uma imagem deprocura das flores, aumentando as chances de seremvisitadas, como sugeriram Araujo et al. (1994).
O segundo conjunto de bromélias (figura 4,conjunto II) foi formado pelas espéciespreferencialmente esciófilas, sendo Phaethorniseurynome seu único ou o mais freqüente agentepolinizador (figura 3). Nesse conjunto de espécies foiobservado um padrão de floração seqüencial econtínuo, de novembro a maio. Esse padrão fenológicoocorreu também em florestas da América Central nasplantas polinizadas por Phaethornithinae (Stiles 1978).As bromeliáceas que foram exclusivamente visitadaspor Phaethornis eurynome e são todas esciófilas,podendo ocorrer no mesmo ambiente. Essas espécies,
porém, apresentam picos de floração em períodosdistintos (figura 1). Assim, a competição porpolinizadores entre as bromeliáceas que ocorrem naárea de estudo é minimizada, como também apossibilidade de hibridação interespecífica.
Ainda dentro desse segundo conjunto de plantas,observaram-se espécies (como Vriesea incurvata,V. flammea e Aechmea ornata) que foram visitadaspor outras espécies de beija-flores, tendo Phaethorniseurynome como seu principal visitante (figura 3). Aprimeira delas, Vriesea incurvata, é uma espécieesciófila, que apresentou floração durante quase toda aestação chuvosa, com pico em janeiro e fevereiro. Noentanto, esteve restrita a ambientes bastante úmidos,próximos a cursos d’água, possibilitando as visitas dePhaethornis eurynome, que forrageia preferencialmenteem ambientes sombreados (Piratelli 1997). Já Vrieseaflammea e Aechmea ornata diferiram em seus períodosde pico de floração (figura 1) e também ocorreram emestratos vegetacionais distintos (a primeira no estratomédio da vegetação e em borda de mata e a última, nodossel). Apesar de ambas serem heliófilas podiamocorrer sombreadas por copas das árvores,possibilitando as visitas constantes de Phaethorniseurynome.
A deposição de pólen em diversos locais do corpodos beija-flores poderia ser um mecanismo para evitara mistura de pólen das diferentes espécies. No entanto,dada a estrutura mais comum das flores destasbromélias, na qual as anteras ocorrem no interior dacorola tubulosa, o bico dos beija-flores tendeu a ser olocal mais freqüente de impregnação de pólen, o quetambém foi observado por Fischer (1994), Sazima et al.(1995, 1996), Araujo (1996) e Buzato et al. (2000).Assim, o período de floração e tipos distintos deambientes onde ocorreram as bromeliáceas pareceramser os principais meios de se evitar a competição porvetores de pólen.
Agradecimentos – À Capes, pelo apoio financeiro; à Fundaçãopara a Conservação e Produção Florestal do Estado de SãoPaulo e ao Parque Estadual Intervales, pelo apoio logístico;aos Departamentos de Botânica (Unicamp) e de CiênciasBiológicas (UEFS); ao Dr. Wesley R. Silva, à Dra. SilvanaBuzato, à Dra. Julie H.A. Dutil e ao Dr. Marco Aurélio Pizo,pelas valiosas críticas; à Dra. Maria da Graça L. Wanderley,pela identificação de algumas bromélias deste estudo; aoscompanheiros do Depto. de Botânica/Unicamp e DCBio/UEFSpela ajuda providencial nas diversas fases deste estudo; e aNívea Mara Ribeiro Rodrigues, pelo precioso auxílio nostrabalhos de campo e acompanhamento deste projeto; aosrevisores anônimos, pelas críticas e sugestões.
Revista Brasil. Bot., V.29, n.1, p.163-174, jan.-mar. 2006 173
Referências bibliográficas
ALTMANN, S.A. 1974. Observacional study of behavior:sampling methods. Behavior 49:227-265.
ARAUJO, A.C. 1996. Beija-flores e seus recursos florais numaárea de planície costeira do litoral norte de São Paulo.SP. Dissertação de mestrado, Universidade Estadual deCampinas, Campinas.
ARAUJO, A.C., FISCHER, E.A. & SAZIMA, M. 1994.Floração seqüencial e polinização de três espécies deVriesea (Bromeliaceae) na região da Juréia, sudeste doBrasil. Revista Brasileira de Botânica 17:113-118.
ARAUJO, A.C., FISCHER, E. & SAZIMA, M. 2004. Asbromélias na região do Rio Verde. In Estação EcológicaJuréia-Itatins: Ambiente físico, flora e fauna (O.A.V.Marques & W. Duleba, eds.). Editora Holos, RibeirãoPreto.
ARIZMENDI, M.C. & ORNELAS, J.F. 1990. Hummingbirdsand their floral resources in tropical dry forest in Mexico.Biotropica 22:172-180.
BENZING, D.H. 1980. The biology of the bromeliads. MadRiver Press, Eureka.
BROWN, J.H. & KODRIC-BROWN, A. 1979. Convergence,competition, and mimicry in a temperate community ofhummingbird-pollinated flowers. Ecology 60:1022-1035.
BUZATO, S., SAZIMA, M. & SAZIMA, I. 2000.Hummingbird-pollinated floras at three Atlantic forestsites. Biotropica 32:824-841.
CANELA, M.B.F. & SAZIMA, M. 2003. Aechmea pectinata:a hummingbird-dependent Bromeliad withinconspicuous flowers from the rainforest in South-eastern Brazil. Annals of Botany 92:731-737.
DAFNI, A. 1992. Pollination Ecology. Oxford UniversityPress, Oxford.
FAEGRI, K. & PIJL, L. 1980. The principles of pollinationecology. Pergamon Press, New York.
FEINSINGER, P. 1983. Variable nectar secretion in a Heliconiaspecies pollinated by hermit hummingbirds. Biotropica15:48-52.
FEINSINGER, P. & COWELL. R.K. 1978. Communityorganization among Neotropical nectar feeding birds.American Zoologist 18:779-795.
FENSTER, C.B. 1991. Selection on floral morphology byhummingbirds. Biotropica 23:98-101.
FISCHER, E.A. 1994. Polinização, fenologia e distribuiçãoespacial de Bromeliaceae numa comunidade de MataAtlântica, litoral sul de São Paulo. Dissertação demestrado, Universidade Estadual de Campinas,Campinas.
FISCHER, E.A. & ARAUJO, A.C. 1995. Spatial organizationof a bromeliad community in the Atlantic Rainforest,South-Eastern Brazil. Journal of Tropical Ecology11:559–567.
GARDNER, C.S. 1986. Inferences about pollination inTillandsia (Bromeliaceae). Selbyana 9:76-87.
GOTTSBERGER, G. 1986. Some pollination strategies inNeotropical savannas and forests. Plant SystematicEvolution 152:26-45.
KAERNS, C.A. & INOUYE, D.W. 1993. Techniques forpollination biologists. University Press of Colorado,Niwot.
KOCHMER, J.P. & HANDEL, S.N. 1986. Constraints andcompetition in the evolution of flowering phenology.Ecological Monographs 56:303-325.
KÖEPEN, W. 1948. Climatología. Fondo de Cultura Econômica,Mexico.
MACHADO, C.G. 1999. Composição e estrutura de bandosmistos de aves na Mata Atlântica do alto da Serra doParanapiacaba, SP. Revista Brasileira de Biologia 1:62-65.
MAGURRAN, A.E. 1988. Ecological diversity and itsmeasurement. Croom Helm, Sydney.
MORELLATO, L.P.C. & LEITÃO FILHO, H.F. 1992. Padrõesde frutificação e dispersão na Serra do Japi. In HistóriaNatural da Serra do Japi: ecologia e preservação de umaárea florestal no Sudeste do Brasil (L.P.C. Morellato,org.). Editora da Unicamp, Campinas, p.112-140.
NASCIMENTO, F.H. 1994. A sucessão secundária inicial naMata Atlântica sobre a Serra de Paranapiacaba, RibeirãoGrande, SP. Dissertação de mestrado, Universidade deSão Paulo, São Paulo.
NEWSTROM, L.E., FRANKIE, G.W. & BAKER, H.G. 1994. Anew classification for plant phenology based onflowering patterns in Lowlands tropical rain forest treesat La Selva, Costa Rica. Biotropica 26:141-159.
PANDEY, C.B. & SINGH, S.J. 1992. Rainfall and grazing effectson net primary productivity in tropical savanna India.Ecology 73:2007-2021.
PIRATELLI, A.J. 1997. Comportamento alimentar de beija-flores em duas espécies de Hippeastrum Herb.(Amaryllidaceae). Revista Brasileira de Biologia57:261-273.
PONÇANO, W.L. 1981. Mapa geomorfológico do Estado deSão Paulo. v.1. Secretaria da Indústria, Comércio eTecnologia do Estado de São Paulo, São Paulo.
POULIN, B., LEFEBVRE, G. & MCNEIL, R. 1992. Tropical avianphenology in relation to abundance and exploitation offood resources. Ecology 73:2295-2309.
REITZ, R. 1983. Bromeliáceas e a malária-bromélia endêmica.In Flora ilustrada catarinense (R. Reitz, ed.). HerbárioBarbosa Rodrigues, Itajaí.
SAZIMA I., BUZATO, S. & SAZIMA, M. 1995. The saw-billed hermit Ramphodon naevius and its flowers insoutheastern Brazil. Journal of Ornithology 136:195-206.
SAZIMA I., BUZATO, S. & SAZIMA, M. 1996. Anassemblage of hummingbird-pollinated flowers inMontane Forest in Southeastern Brazil. Botanica Acta109:149-160.
SAZIMA, I., VOGEL, S. & SAZIMA, M. 1989. Bat pollinationof Encholirium glaziovii, a terrestrial bromeliad. PlantSystematic Evolution 168:167-179
C.G. Machado & J. Semir: Fenologia e biologia floral de bromeliáceas ornitófilas174
SICK, H. 1984. Ornitologia brasileira: uma introdução.Universidade de Brasília, Brasília, DF.
SNOW, D.W. & SNOW, B.K. 1980. Relationships betweenhummingbirds and flowers in the Andes of Colombia.Bulletin of the British Museum of Natural History(Zoology) 38:105-139.
SNOW, D.W. & SNOW, B.K. 1986. Feeding ecology ofhummingbirds in the Serra do Mar, southeastern Brazil.El Hornero 12:286-296.
SNOW, D.W. & TEIXEIRA, D.L. 1982. Hummingbirds andtheir flowers in the coastal montains of southeasternBrazil. Journal of Ornithology 123:446-450.
SOKAL, R.R. & ROHLF, F.J. 1981. Biometry. FreemanCompany, San Francisco.
SSPS. 1983.Statistical package for social sciences - user’sguide. McGraw-Hill, Chicago.
STILES, F.G. 1978. Temporal organization of flowering amongthe hummingbird foodplants of a tropical wet forest.Biotropica 10:194-210.
TALORA, D.C. & MORELLATO, L.P.C. 2000. Fenologia deespécies arbóreas em floresta de planície litorânea dosudeste do Brasil. Revista Brasileira de Botânica 23:13-26.
TIEBOUT III, H.M. 1993. Mechanisms of competition intropical hummingbirds: metabolic costs for loosers andwinners. Ecology 74:405-418.