Field effect transistors based on semiconductive microbially synthesized chalcogenide nanofibers Ian R. McFarlane a , Julia R. Lazzari-Dean b , Mohamed Y. El-Naggar a,c,⇑ a Department of Physics and Astronomy, University of Southern California, 920 Bloom Walk, Seaver Science Center 215C, Los Angeles, CA 90089-0484, USA b Department of Chemistry, University of Southern California, Los Angeles, CA 90089-0484, USA c Molecular and Computational Biology Section, Department of Biological Sciences, University of Southern California, Los Angeles, CA 90089-0484, USA article info Article history: Received 27 June 2014 Received in revised form 3 October 2014 Accepted 4 November 2014 Available online 11 November 2014 Keywords: Shewanella Arsenic sulfide Nanostructures Field effect transistor Biogenic materials abstract Microbial redox activity offers a potentially transformative approach to the low-temperature synthesis of nanostructured inorganic materials. Diverse strains of the dissimilatory metal-reducing bacteria Shewanella are known to produce photoactive filamentous arsenic sulfide nanomaterials by reducing arsenate and thiosulfate in anaerobic culture conditions. Here we report in situ microscopic observations and measure the thermally activated (79 kJ mol 1 ) precipitation kinetics of high yield (504 mg per liter of culture, 82% of theoretical maximum) extracellular As 2 S 3 nanofibers produced by Shewanella sp. strain ANA-3, and demonstrate their potential in functional devices by constructing field effect transistors (FETs) based on individual nanofibers. The use of strain ANA-3, which possesses both respiratory and detoxification arsenic reductases, resulted in significantly faster nanofiber synthesis than other strains previously tested, mutants of ANA-3 deficient in arsenic reduction, and when compared to abiotic arsenic sulfide precipitation from As(III) and S 2. Detailed characterization by electron microscopy, energy-dis- persive X-ray spectroscopy, electron probe microanalysis and Tauc analysis of UV-vis spectrophotometry showed the biogenic precipitate to consist primarily of amorphous As 2 S 3 nanofibers with an indirect optical band gap of 2.37 eV. X-ray diffraction also revealed the presence of crystalline As 8 S 9-x minerals that, until recently, were thought to form only at higher temperatures and under hydrothermal condi- tions. The nanoscale FETs enabled a detailed characterization of the charge mobility (10 5 cm 2 V 1 s 1 ) and gating behavior of the heterogeneously doped nanofibers. These studies indicate that the biotrans- formation of metalloids and chalcogens by bacteria enables fast, efficient, sustainable synthesis of technologically relevant chalcogenides for potential electronic and optoelectronic applications. Ó 2014 Acta Materialia Inc. Published by Elsevier Ltd. This is an open access article under the CC BY-NC- ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/). 1. Introduction The synthesis of nanomaterials by biological or biomimetic means in physiological conditions offers multiple advantages over traditional physical and chemical strategies that typically require more extreme environments (temperature, pressure and pH). In addition to the promise of cheaper and greener synthesis pro- cesses, the resulting biogenic materials can exhibit unique mor- phologies and physical/chemical properties stemming from the tight control organisms exert over the composition, nucleation, crystallography and desired function of these materials [1]. While significant attention has been dedicated to understanding the syn- thesis and structure–function relations of the most abundant biominerals, especially carbonates and phosphates, the last decade has also witnessed additional interest in exploiting biological, especially microbial, strategies for producing a wider range of syn- thetic materials with technologically relevant mechanical, optical, electronic and magnetic functionalities [2–4]. Towards this goal, recent reports [5–7] demonstrated the synthesis of extracellular chalcogenide nanostructures with unique optoelectronic proper- ties using a bacterial process relying on anaerobic respiration and detoxification activities to alter the oxidation states of the metal, metalloid and chalcogen precursors. Chalcogenide compounds, resulting from the reaction of group VI elements (particularly S, Se and Te) with more electropositive elements (e.g. As, Sb, Si, Ge, Zn, Cd), represent an intriguing target for biogenic synthesis. Chalcogenides have been described as ‘‘cha- meleon’’ compounds because of their remarkable versatility: depending on composition and synthesis techniques they may be crystalline, glassy, metallic, semiconductive or ionic conductors http://dx.doi.org/10.1016/j.actbio.2014.11.005 1742-7061/Ó 2014 Acta Materialia Inc. Published by Elsevier Ltd. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/). ⇑ Corresponding author at: Department of Physics and Astronomy, University of Southern California, 920 Bloom Walk, Seaver Science Center 215C, Los Angeles, CA 90089-0484, USA. Tel.: +1 (213) 740 2394; fax: +1 (213) 740 6653. E-mail address: [email protected](M.Y. El-Naggar). Acta Biomaterialia 13 (2015) 364–373 Contents lists available at ScienceDirect Acta Biomaterialia journal homepage: www.elsevier.com/locate/actabiomat
Field effect transistors based on semiconductive microbially synthesizedchalcogenide nanofibers
http://dx.doi.org/10.1016/j.actbio.2014.11.0051742-7061/� 2014 Acta Materialia Inc. Published by Elsevier Ltd.This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
⇑ Corresponding author at: Department of Physics and Astronomy, University ofSouthern California, 920 Bloom Walk, Seaver Science Center 215C, Los Angeles, CA90089-0484, USA. Tel.: +1 (213) 740 2394; fax: +1 (213) 740 6653.
Ian R. McFarlane a, Julia R. Lazzari-Dean b, Mohamed Y. El-Naggar a,c,⇑a Department of Physics and Astronomy, University of Southern California, 920 Bloom Walk, Seaver Science Center 215C, Los Angeles, CA 90089-0484, USAb Department of Chemistry, University of Southern California, Los Angeles, CA 90089-0484, USAc Molecular and Computational Biology Section, Department of Biological Sciences, University of Southern California, Los Angeles, CA 90089-0484, USA
a r t i c l e i n f o
Article history:Received 27 June 2014Received in revised form 3 October 2014Accepted 4 November 2014Available online 11 November 2014
Microbial redox activity offers a potentially transformative approach to the low-temperature synthesis ofnanostructured inorganic materials. Diverse strains of the dissimilatory metal-reducing bacteriaShewanella are known to produce photoactive filamentous arsenic sulfide nanomaterials by reducingarsenate and thiosulfate in anaerobic culture conditions. Here we report in situ microscopic observationsand measure the thermally activated (79 kJ mol�1) precipitation kinetics of high yield (504 mg per liter ofculture, 82% of theoretical maximum) extracellular As2S3 nanofibers produced by Shewanella sp. strainANA-3, and demonstrate their potential in functional devices by constructing field effect transistors(FETs) based on individual nanofibers. The use of strain ANA-3, which possesses both respiratory anddetoxification arsenic reductases, resulted in significantly faster nanofiber synthesis than other strainspreviously tested, mutants of ANA-3 deficient in arsenic reduction, and when compared to abiotic arsenicsulfide precipitation from As(III) and S2�. Detailed characterization by electron microscopy, energy-dis-persive X-ray spectroscopy, electron probe microanalysis and Tauc analysis of UV-vis spectrophotometryshowed the biogenic precipitate to consist primarily of amorphous As2S3 nanofibers with an indirectoptical band gap of 2.37 eV. X-ray diffraction also revealed the presence of crystalline As8S9-x mineralsthat, until recently, were thought to form only at higher temperatures and under hydrothermal condi-tions. The nanoscale FETs enabled a detailed characterization of the charge mobility (�10�5 cm2 V�1 s�1)and gating behavior of the heterogeneously doped nanofibers. These studies indicate that the biotrans-formation of metalloids and chalcogens by bacteria enables fast, efficient, sustainable synthesis oftechnologically relevant chalcogenides for potential electronic and optoelectronic applications.� 2014 Acta Materialia Inc. Published by Elsevier Ltd. This is an open access article under the CC BY-NC-
The synthesis of nanomaterials by biological or biomimeticmeans in physiological conditions offers multiple advantages overtraditional physical and chemical strategies that typically requiremore extreme environments (temperature, pressure and pH). Inaddition to the promise of cheaper and greener synthesis pro-cesses, the resulting biogenic materials can exhibit unique mor-phologies and physical/chemical properties stemming from thetight control organisms exert over the composition, nucleation,crystallography and desired function of these materials [1]. Whilesignificant attention has been dedicated to understanding the syn-thesis and structure–function relations of the most abundant
biominerals, especially carbonates and phosphates, the last decadehas also witnessed additional interest in exploiting biological,especially microbial, strategies for producing a wider range of syn-thetic materials with technologically relevant mechanical, optical,electronic and magnetic functionalities [2–4]. Towards this goal,recent reports [5–7] demonstrated the synthesis of extracellularchalcogenide nanostructures with unique optoelectronic proper-ties using a bacterial process relying on anaerobic respiration anddetoxification activities to alter the oxidation states of the metal,metalloid and chalcogen precursors.
Chalcogenide compounds, resulting from the reaction of groupVI elements (particularly S, Se and Te) with more electropositiveelements (e.g. As, Sb, Si, Ge, Zn, Cd), represent an intriguing targetfor biogenic synthesis. Chalcogenides have been described as ‘‘cha-meleon’’ compounds because of their remarkable versatility:depending on composition and synthesis techniques they may becrystalline, glassy, metallic, semiconductive or ionic conductors
[8]. This versatility leads to a wide range of tunable functionalitiesin various components including sensors, waveguides, photoniccrystals [9] and photoactive devices [10–12]. Chalcogenide glassesare already commercially important in write-once and rewritableoptical storage disks as well as phase-change memory relying onamorphous–crystalline transformations [8]. Some members of thisfamily (e.g. As2S3) are infrared-transparent (700 nm–11.5 lm) andare therefore candidate materials for applications in infrareddevices [9,13]. From an energy conversion standpoint, chalcoge-nides have been intensively investigated as photovoltaic materialsfor solar cells [12]. In addition, AsxSy and AsxSey glasses have beenreported to exhibit an optomechanical effect [10,11] for directlight-to-mechanical energy conversion when irradiated with polar-ized light matching the band gap energy. This phenomenon hasbeen exploited to generate mechanical strain, resulting in all-opti-cal (electricity-free) actuation of chalcogenide-coated cantilevers[10]. Further interest in unique and tunable size-dependent prop-erties has motivated the development of quasi-one-dimensional(nanotube and nanowire) chalcogenides. The majority of theseefforts, however, focus on physical and chemical synthesis tech-niques requiring templates, precursors, and hydro- or solvothermalmethods, typically under relatively extreme conditions [14].
More recently, Hur et al. reported a microbiological route forsynthesizing chalcogenide nanostructures by exploiting dissimila-tory metal-reducing bacteria, specifically a number of Shewanellaspecies, to produce an extracellular network of filamentous arsenicsulfide nanofibers/nanotubes in anaerobic culture conditions [5,6].This process relies on the remarkable metabolic versatility ofShewanella [15,16] to concomitantly reduce S2O3
2� to S2� andAs(V) to As(III). In this system, the thiosulfate S2O3
2� serves as a ter-minal electron acceptor for respiration in lieu of O2 under anaero-bic conditions, resulting in S2�, while arsenate reduction is thoughtto occur via As reductases tied to either the respiration or detoxi-fication pathways of Shewanella. Rather surprisingly, it wasreported that the resulting amorphous and polycrystalline nano-structures behaved as simultaneously metallic and semiconductivein terms of their electrical and photoconductive properties, respec-tively [5]. An additional report demonstrated the incorporation ofmore elements (Cd, Se) into As–S nanostructures through biogenicdeposition and/or abiotic cation exchange to create ternary (As–S–Se and As–Cd–S) and quaternary (As–Cd–S–Se) composites [7].Using X-ray absorption near-edge structure (XANES) and Fouriertransformed extended X-ray absorption fine structure (EXAFS)analyses, the previous reports also confirmed the elemental oxida-tion states within the biogenic material to be consistent with As2S3
[5,7].In this study, we present in situ microscopic observations to
characterize the microbial synthesis of individual arsenic sulfidenanofibers, measure the synthesis kinetics, and detail the struc-tural, crystallographic, electronic and band gap properties of thenanofibers resulting from the reduction activity of Shewanella.In contrast to previous detailed studies of chalcogenide synthesisby Shewanella sp. strain HN-41 [5,7], we focused our attention onShewanella oneidensis MR-1 as well as Shewanella sp. strain ANA-3and mutants of strain ANA-3 deficient in arsenic reduction. Inaddition to being genetically tractable, which will help shed lighton the genetic and biomolecular mechanisms underlying nano-material synthesis, strain ANA-3 is widely regarded as the proto-typical arsenic reducer, as a result of containing both thedetoxification (encoded by ars genes) and respiratory (encodedby arr genes) As(V) reduction pathways [17–19]. The ars pathwayis well characterized and known to be present in many bacteria[19,20]. Using this pathway, As(V) enters the cytosol by phos-phate transporters and is reduced to As(III) by the arsenate reduc-tase ArsC. As(III) is subsequently extruded from the cells by acytoplasmic membrane efflux pump, ArsB, which may interact
with an ATPase subunit, ArsA, to drive As(III) efflux using ATPhydrolysis. In contrast, the arr pathway has only been recentlydescribed [18,21]. The arrAB operon of strain ANA-3 encodes alarge Mo-containing enzyme, ArrA, and an Fe–S protein, ArrB.Both ArrA and ArrB are required for respiratory reduction ofAs(V) [18,19]. The use of strain ANA-3 resulted in significantlymore rapid precipitation of As–S nanofibers than previouslyreported under similar conditions. Furthermore, we demonstratenovel field-effect transistors (FETs) based on single biogenicnanofibers, and study their charge mobility and switching behav-ior as a function of backgating to identify the doping type andmajority charge carriers in these semiconductors.
2. Materials and methods
2.1. Bacterial growth
A list of the bacterial strains and mutants used in this study isprovided in Table 1. The inocula were grown aerobically in 20 mlof LB medium from a frozen (�80 �C) stock up to an optical densityat 600 nm (OD600) of 1.5 ± 0.15. These aerobic pre-cultures wereinoculated at 0.1% (v/v) into anaerobic serum bottles each contain-ing 80 mL HEPES-buffered (30 mM) medium consisting of: 20 mMsodium DL-lactate, as electron donor; 28 mM ammonium chloride;1.34 mM potassium chloride; 4.35 mM sodium phosphatemonobasic; 20 mM sodium hydroxide; 10 mM thiosulfate, asNa2S2O3�5H2O; and 5 mM arsenate, as Na2HAsO4�7H2O. Vitamins,amino acids and trace mineral stock solutions were used tosupplement the medium as described previously [22]. The mediumwas adjusted to an initial pH of 7.25, and anaerobic conditions werereached by purging with 100% N2 for in excess of 45 min. Theanaerobic serum bottles, sealed with butyl stoppers and aluminumseals, were sterilized by autoclaving at 120 �C for 15 min. Arsenate,thiosulfate and vitamins were added after autoclaving. All cultureswere grown at 30 �C and agitated at a rate of 150 rpm.
2.2. Preparation of nanomaterials for EDS measurements, Taucanalysis and transistor microfabrication
For each preparation, 1 ml of the As–S precipitate was diluted in14 ml of ultrapure water (Millipore Milli-Q Integral 5 purificationsystem) and pelleted by centrifugation at 3000 RCF for 20 min.The samples were allowed to settle for a further 15 min beforeremoving the supernatant. The samples were then washed eighttimes to remove salts, without further centrifugation, allowing15 min between the addition and removal of ultrapure water forthe material to settle at each step. For device fabrication, the result-ing yellow precipitates were drop deposited on oxidized siliconchips for 5–15 min before removing the liquid by capillary actionusing a Kimwipe. The chips were then dried under nitrogen gas.
2.3. In situ microscopy of nanofiber synthesis
Microscopy observation chambers (500 lm height, 3 cm2 sur-face area, 150 ll volume) were constructed by using double-sidedtape (Scotch 3 M Permanent) to separate a coverslip (VWR No. 1)from the glass region of a glass-bottom Petri dish (WillCo Wells,GWSt-5030). A 1 mm diameter filling port was created by placinga 0.5 cm length of plastic microtubing in a drilled hole on the topof the Petri dish lid and fixing it in place with epoxy (Devon 5 Min-ute Gel). All components were sterilized in ethanol and allowed todry before cell injection. An overnight culture of strain ANA-3 wasgrown in LB medium to OD600 2.0 and inoculated into the anaero-bic defined medium described above at 1% (v/v). Subsequentchamber filling and assembly was performed in a nitrogen bag,
Table 1Strains and mutants used in this study and time from inoculation until visible formation of As–S precipitate in medium supplemented with 10 mM thiosulfate and 5 mM arsenate.
Strain/mutant Description Time to visible precipitate (h)
MR-1 Wild-type, Shewanella oneidensis [15] 144 ± 24ANA-3 Wild-type, Shewanella sp. [17] 19 ± 2.5ARRA3 DarrA mutant derived from ANA-3; does not respire As(V) [18] 75 ± 10ARSB1 arsB::Kan(EZ::TN < KAN-2>); arsB mutant derived from ANA-3; deficient in detoxification of As(V) [17] 40 ± 5ARM1 DarrA DarsC mutant derived from ARRA3; does not reduce As(V) under any condition [19] 40 ± 5
and anaerobic conditions were maintained for the duration of theexperiment after transfer to the defined medium. Cells wereinjected slowly into the 150 ll chamber, avoiding bubbles. ThePetri dish lid was sealed to the base with epoxy, and dishes werefilled with additional inoculated medium through the previouslydrilled port to minimize the nitrogen headspace. The filling portwas subsequently sealed with epoxy and the sample was placedon an inverted Nikon Eclipse Ti microscope with a heated stagefor time-lapse transmission or reflection imaging. The microscopewas equipped with a drift correction unit (Nikon Perfect FocusSystem) for maintaining focus at the dish–medium interface duringtime-lapse imaging. Images were captured at 5 min intervalsduring fiber formation and a shutter was used to avoid sampleillumination except while actively capturing images. Fluorescencemicroscopy was also used to assess the localization of the biogenicnanofibers relative to cells as indicated by the NanoOrange proteinstain (Life Technologies, catalog no. N-6666).
2.4. Growth kinetics
To investigate the growth kinetics of the biogenic nanofibers,we systematically investigated the bulk precipitation of As–S as afunction of temperature. For each measurement, Shewanella sp.strain ANA-3 cultures were incubated with arsenate and thiosul-fate, as described above, at 30 �C for 16.5 h in order to seed thereduction and precipitation reactions. The cultures were thenmoved to an incubator already warmed to the temperature ofinterest and the total precipitate was subsequently pelleted,washed and weighed at regular intervals. First, 40 ml of each80 ml experiment was centrifuged at 5750 RCF for 5 min, followedby standing still in the vertical position for an additional 15 min toallow the sample to settle. The supernatant was removed and theprecipitate from the remaining 40 ml volume was then pelletedin the same container. The supernatant was removed again andthe walls of the container were washed with a stream of ultrapurewater to remove any precipitate from the walls. The container wasthen filled to 15 ml with ultrapure water and centrifuged a thirdtime. Each sample was then washed six times to remove salts, asdescribed above for Tauc analysis (Section 2.2) and then driedovernight in air on weigh dishes. Pellets were finally weighed onweigh dishes in triplicate.
2.5. Structural, optoelectronic, and electronic (FET) characterization
The biogenic nanomaterials were studied by scanning electronmicroscopy (SEM) using a JEOL JSM 7001F field-emission micro-scope equipped for energy-dispersive X-ray spectroscopy (EDS)for elemental analysis. Further quantitative analyses (1–2% accu-racy) of stoichiometry were performed by electron probe micro-analysis (EPMA) with a JEOL JXA-8200 interfaced to multiplewavelength dispersive X-ray spectrometers (WDS) and featuringreference standards of known composition. Each EPMA volumesampled is typically a few cubic microns, corresponding to aweight of a few picograms. The nanomaterials were also observedby transmission electron microscopy (TEM) using a JEOL 2100F inboth imaging (bright-field and dark-field) and diffraction modeswith a high tension of 200 kV. The microstructure and crystallinity
of the nanomaterial was also characterized by h–2h X-ray diffrac-tion (XRD) scans on a Rigaku Ultima IV diffractometer.
The optical band gap was estimated from Tauc plots (i.e. (ahm)n
vs. hm where a is the absorption coefficient and hm is the photonenergy) after measuring the UV–vis spectrum using a Thermo Sci-entific NanoDrop 2000c UV–vis spectrophotometer.
For fabrication of nanofiber-based FETs, the nanomaterials weredeposited on oxidized silicon chips (1 lm SiO2 layer on degener-ately doped Si) with photolithographically pre-patterned Au con-tact pads as described previously [23,24]. Nanofibers werelocated by SEM in between the pre-patterned contacts, andsource/drain Pt electrodes were deposited using a focused ionbeam system (JEOL JIB-4500 multibeam SEM/FIB instrument)equipped with a Pt gas injection system (Oxford InstrumentsOmniGIS) to connect the gold contact pads to the nanofibers.The underlying doped Si substrate was used for backgating theresulting single-nanofiber FETs. Drain-to-source current vs. voltage(Ids � Vds) measurements as a function of gating voltage (Vg) wereperformed on a Signatone S1160 probe station instrumented toan Agilent 4156C semiconductor parameter analyzer.
2.6. Detection of arsenic
Aqueous phase As(V) and As(III) concentrations were measuredusing an integrated ion chromatography (IC) and atomic fluores-cence spectrometry (AFS) system. The IC system was a DionexGP40 gradient pump within a Dionex LC20 chromatography enclo-sure connected to a Hamilton PRP-X100 10 lm 4.1 � 250 mm col-umn. The AFS was a PS Analytical PSA 10.055. Solutions used were:mobile phase of 10 mM K2HPO4 and KH2PO4 adjusted to a pH of6.25, reductant base of 100 mM NaOH and 530 mM NaBH4, andacid solution of 1.5 M HCl. All reagents and phases were preparedin ultrapure water. Calibration samples were prepared from stan-dard anaerobic growth media with both arsenite and arsenate in0.63, 0.94, 1.56, 2.19, 3.12 and 5 mM concentrations. Calibrationand data samples were diluted to 10�4 of the original concentra-tion using identical procedures and injected into the IC/AFS sys-tem. Each calibration point was measured twice; each data pointfour times. The calibration curve was fitted quadratically, con-straining zero area to correspond to zero concentration. Analysisinvolved baseline removal, integration of peak areas and subse-quent comparison to the calibration curve. The As(III) signalpeaked from roughly 1.75 to 2.75 min and As(V) from 3.25 to4.5 min. The signal typically returned to baseline between peaks,indicating sufficient fractionation by the column for accurate anal-ysis and quantification.
3. Results
3.1. Synthesis, in situ microscopy, and growth kinetics of As–Snanofibers
The production of As–S was visually detected as a bright yellowprecipitate in all cultures incubated with As(V) and S2O3
2� (Fig. 1).Shewanella sp. strain ANA-3 resulted in rapid precipitation (<1 day)compared to S. oneidensis MR-1 (6 days) (Table 1). Remarkably, all
Fig. 2. False color scanning electron microscopy of Shewanella sp. strain ANA-3 cells(blue) and arsenic sulfide nanofibers (yellow). The nanofibers ranged in diameterfrom 20 to 600 nm and reached 150 lm in length.
three ANA-3 mutants produced As–S precipitate, albeit at latertimes relative to wild-type (1.7–3.1 days, Table 1), despite the dis-ruption of critical components involved in successful As(V) reduc-tion. SEM revealed that the precipitate consisted exclusively ofnanofibers, and nanofiber bundles, ranging in diameter from 20to 600 nm and reaching up to 150 lm in length. While cells werealso routinely observed in the SEM preparations (Fig. 2), the nanof-ibers were located extracellularly.
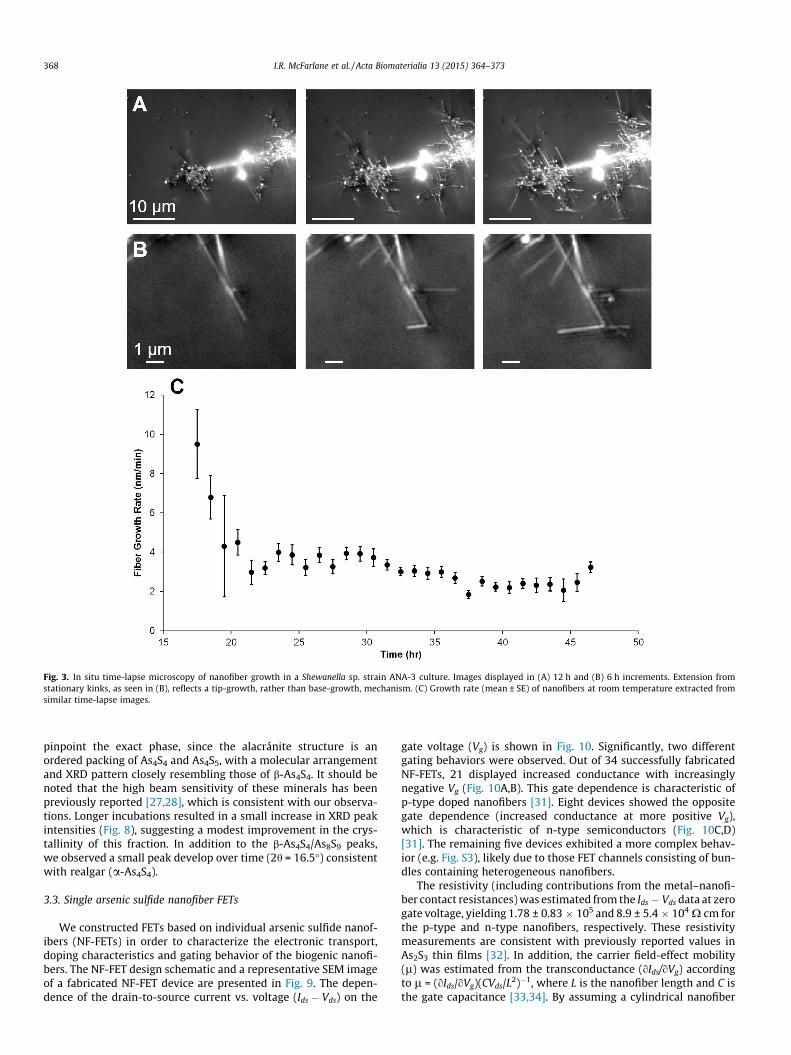
To shed further light on the mechanism of nanofiber formation,we performed in situ microscopy to measure the growth rate andlocalization of nanofibers relative to Shewanella sp. strain ANA-3cells (Fig. 3A–B and Fig. S1). Fiber growth at room temperaturestarted �17 h after the inoculation of the in situ observation cham-ber, with an initial rate of roughly 10 ± 2 nm min�1 (based on sixfibers) (Fig. 3C). The growth rate subsequently declined from thisinitial value, reaching a steady 3.1 ± 0.4 nm min�1 (based on 45fibers) within 4 h. While active bacterial cultures are needed to ini-tiate the reduction (non-inoculated control medium with As(V)and S2O3
2� did not result in precipitation or reduction of As/S, con-sistent with Ref. [5]), we observed that the nucleation of the nanof-ibers did not originate at cell bodies. Furthermore, the subsequentextension of the nanofibers generally occurred in the extracellularspace, and the extension pattern reflected a tip-growth, rather thanbase-growth, mechanism (Fig. 3B).
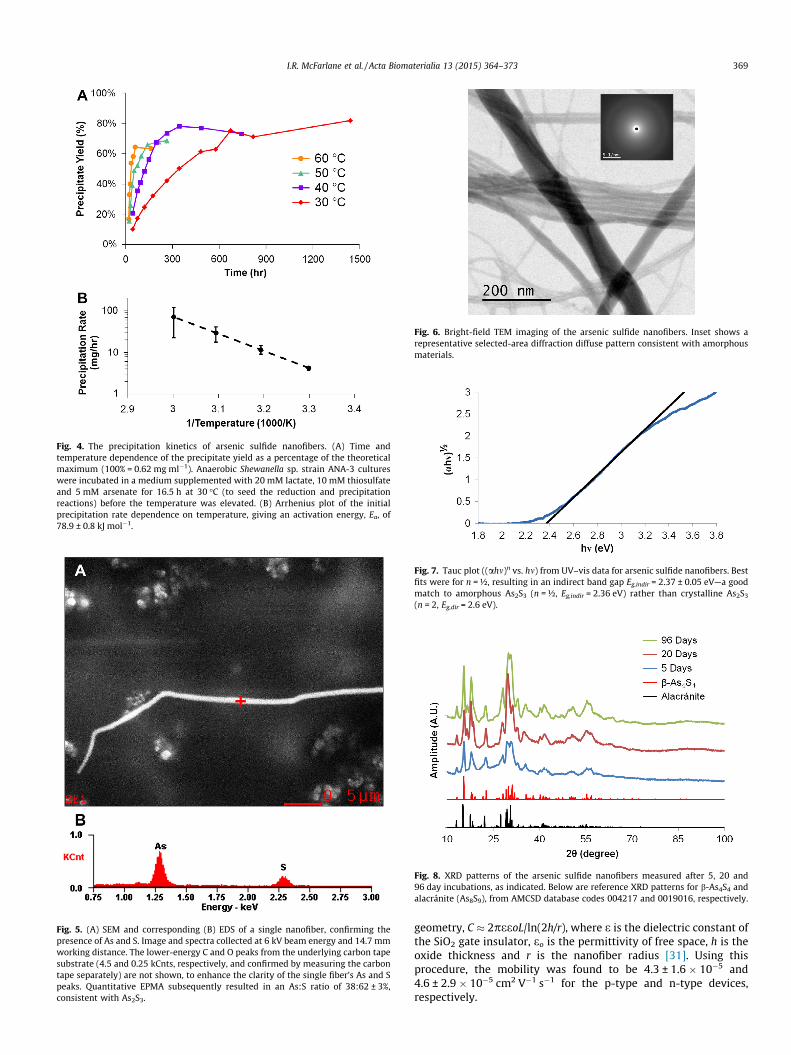
By weighing the total precipitate after 2 months of incubationat 30 �C using Shewanella sp. strain ANA-3, we found that the totalprecipitate yield was 82 ± 8% of the theoretical maximum yield(49.2 mg from each 80 ml batch experiment), calculated based onthe starting As and S concentrations used in our experiments andthe resulting nanofiber stoichiometry (As2S3—see Section 3.2 andDiscussion section below). To investigate the nanofiber growthkinetics, we measured the precipitation rate as a function of tem-perature (Fig. 4). To avoid the adverse effects of higher tempera-tures on the mesophilic Shewanella, and since fiber formationstarted between 16.5 and 21.5 h at 30 �C (Table 1 and Fig. 3), thetemperature was only elevated after 16.5 h of incubation at30 �C. Fig. 4A shows that the nanofiber formation continued atthese higher temperatures, approaching a maximum yield similarto that seen for 30 �C. The initial precipitation rate (extrapolatedfrom the initial slope of Fig. 4A) followed an Arrhenius dependence
on temperature with the rate given by Ae�EaRT , where A is a pre-
exponential factor, Ea is the activation energy, R is the universalgas constant and T is the growth temperature. From the Arrheniusplot displayed in Fig. 4B, the activation energy isEa = 78.9 ± 0.8 kJ mol�1.
3.2. Compositional, structural, and optoelectronic characterization
EDS confirmed the presence of As and S in the biogenic nanofi-bers (Fig. 5). After this initial confirmation of arsenic sulfide forma-tion, the stoichiometry was more quantitatively determined by
Fig. 1. Incubation of Shewanella sp. strain ANA-3 in a medium supplemented with 20 mquantities of arsenic sulfide nanofibers. Images are in increasing magnification taken by
EPMA to give the As and S at.%, using 14 measurements across fourdifferent locations on a dense As–S preparation. The resulting As:Sratio of 38:62 ± 3% was found to be consistent with an As2S3
stoichiometry.Bright-field TEM images (Fig. 6) did not reveal any grain bound-
aries or diffraction-related contrast. In addition, selected-area dif-fraction mode imaging primarily revealed a diffuse ring pattern(inset of Fig. 6) consistent with an amorphous structure [13],except for infrequent diffraction peak patterns found while scan-ning the sample area (see below).
The optical band gap of the As2S3 nanofibers was determined byTauc plots i.e. (ahm)n vs. hm [25,26] (Fig. 7). The best linear fit to theabsorption onset was obtained with n = ½, indicating an indirectband gap semiconductor, and extrapolating to the abscissa gavethe band gap Eg,indir = 2.37 ± 0.05 eV (error is standard error).
A small fraction of the nanofiber morphologies exhibited peri-odic branching with conserved angles (Fig. S2), and occasionalweak diffraction peaks were observed while scanning across vari-ous regions of the precipitate and acquiring TEM diffraction pat-terns from a high-speed CCD camera (Gatan Orius 200D) (datanot shown). The latter approach was used in order to limit theexposure time, since the samples were found to be highly sensitiveto beam exposure, making it difficult to acquire diffraction datawith conventional means. These observations motivated us toexamine the crystallinity of the samples with XRD h–2h scans.The overall XRD pattern presented in Fig. 8 is a close match tothe XRD patterns of b-As4S4 (As8S8) [27,28] and alacránite (As8S9)[29], as well as the previously reported AsxS9-x series spanningthe composition from b-As4S4 to As8S9 [30]. It is difficult to further
M lactate, 10 mM thiosulfate and 5 mM arsenate results in macroscopically visible(A) optical camera and (B, C) scanning electron microscopy.
Fig. 3. In situ time-lapse microscopy of nanofiber growth in a Shewanella sp. strain ANA-3 culture. Images displayed in (A) 12 h and (B) 6 h increments. Extension fromstationary kinks, as seen in (B), reflects a tip-growth, rather than base-growth, mechanism. (C) Growth rate (mean ± SE) of nanofibers at room temperature extracted fromsimilar time-lapse images.
pinpoint the exact phase, since the alacránite structure is anordered packing of As4S4 and As4S5, with a molecular arrangementand XRD pattern closely resembling those of b-As4S4. It should benoted that the high beam sensitivity of these minerals has beenpreviously reported [27,28], which is consistent with our observa-tions. Longer incubations resulted in a small increase in XRD peakintensities (Fig. 8), suggesting a modest improvement in the crys-tallinity of this fraction. In addition to the b-As4S4/As8S9 peaks,we observed a small peak develop over time (2h = 16.5�) consistentwith realgar (a-As4S4).
3.3. Single arsenic sulfide nanofiber FETs
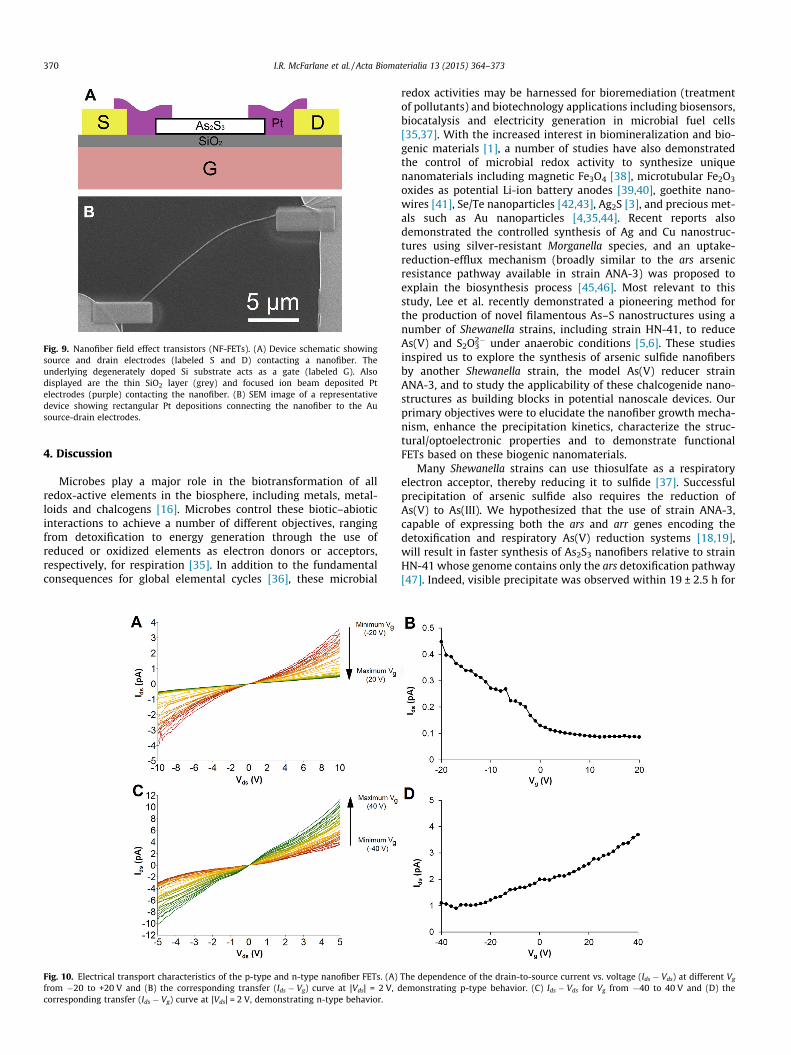
We constructed FETs based on individual arsenic sulfide nanof-ibers (NF-FETs) in order to characterize the electronic transport,doping characteristics and gating behavior of the biogenic nanofi-bers. The NF-FET design schematic and a representative SEM imageof a fabricated NF-FET device are presented in Fig. 9. The depen-dence of the drain-to-source current vs. voltage (Ids � Vds) on the
gate voltage (Vg) is shown in Fig. 10. Significantly, two differentgating behaviors were observed. Out of 34 successfully fabricatedNF-FETs, 21 displayed increased conductance with increasinglynegative Vg (Fig. 10A,B). This gate dependence is characteristic ofp-type doped nanofibers [31]. Eight devices showed the oppositegate dependence (increased conductance at more positive Vg),which is characteristic of n-type semiconductors (Fig. 10C,D)[31]. The remaining five devices exhibited a more complex behav-ior (e.g. Fig. S3), likely due to those FET channels consisting of bun-dles containing heterogeneous nanofibers.
The resistivity (including contributions from the metal–nanofi-ber contact resistances) was estimated from the Ids � Vds data at zerogate voltage, yielding 1.78 ± 0.83 � 105 and 8.9 ± 5.4 � 104 X cm forthe p-type and n-type nanofibers, respectively. These resistivitymeasurements are consistent with previously reported values inAs2S3 thin films [32]. In addition, the carrier field-effect mobility(l) was estimated from the transconductance (oIds/oVg) accordingto l = (oIds/oVg)(CVds/L2)�1, where L is the nanofiber length and C isthe gate capacitance [33,34]. By assuming a cylindrical nanofiber
Fig. 4. The precipitation kinetics of arsenic sulfide nanofibers. (A) Time andtemperature dependence of the precipitate yield as a percentage of the theoreticalmaximum (100% = 0.62 mg ml�1). Anaerobic Shewanella sp. strain ANA-3 cultureswere incubated in a medium supplemented with 20 mM lactate, 10 mM thiosulfateand 5 mM arsenate for 16.5 h at 30 �C (to seed the reduction and precipitationreactions) before the temperature was elevated. (B) Arrhenius plot of the initialprecipitation rate dependence on temperature, giving an activation energy, Ea, of78.9 ± 0.8 kJ mol�1.
Fig. 5. (A) SEM and corresponding (B) EDS of a single nanofiber, confirming thepresence of As and S. Image and spectra collected at 6 kV beam energy and 14.7 mmworking distance. The lower-energy C and O peaks from the underlying carbon tapesubstrate (4.5 and 0.25 kCnts, respectively, and confirmed by measuring the carbontape separately) are not shown, to enhance the clarity of the single fiber’s As and Speaks. Quantitative EPMA subsequently resulted in an As:S ratio of 38:62 ± 3%,consistent with As2S3.
Fig. 6. Bright-field TEM imaging of the arsenic sulfide nanofibers. Inset shows arepresentative selected-area diffraction diffuse pattern consistent with amorphousmaterials.
Fig. 7. Tauc plot ((ahm)n vs. hm) from UV–vis data for arsenic sulfide nanofibers. Bestfits were for n = ½, resulting in an indirect band gap Eg,indir = 2.37 ± 0.05 eV—a goodmatch to amorphous As2S3 (n = ½, Eg,indir = 2.36 eV) rather than crystalline As2S3
(n = 2, Eg,dir = 2.6 eV).
Fig. 8. XRD patterns of the arsenic sulfide nanofibers measured after 5, 20 and96 day incubations, as indicated. Below are reference XRD patterns for b-As4S4 andalacránite (As8S9), from AMCSD database codes 004217 and 0019016, respectively.
geometry, C � 2peeoL/ln(2h/r), where e is the dielectric constant ofthe SiO2 gate insulator, eo is the permittivity of free space, h is theoxide thickness and r is the nanofiber radius [31]. Using thisprocedure, the mobility was found to be 4.3 ± 1.6 � 10�5 and4.6 ± 2.9 � 10�5 cm2 V�1 s�1 for the p-type and n-type devices,respectively.
Fig. 9. Nanofiber field effect transistors (NF-FETs). (A) Device schematic showingsource and drain electrodes (labeled S and D) contacting a nanofiber. Theunderlying degenerately doped Si substrate acts as a gate (labeled G). Alsodisplayed are the thin SiO2 layer (grey) and focused ion beam deposited Ptelectrodes (purple) contacting the nanofiber. (B) SEM image of a representativedevice showing rectangular Pt depositions connecting the nanofiber to the Ausource-drain electrodes.
Microbes play a major role in the biotransformation of allredox-active elements in the biosphere, including metals, metal-loids and chalcogens [16]. Microbes control these biotic–abioticinteractions to achieve a number of different objectives, rangingfrom detoxification to energy generation through the use ofreduced or oxidized elements as electron donors or acceptors,respectively, for respiration [35]. In addition to the fundamentalconsequences for global elemental cycles [36], these microbial
Fig. 10. Electrical transport characteristics of the p-type and n-type nanofiber FETs. (A)from �20 to +20 V and (B) the corresponding transfer (Ids � Vg) curve at |Vds| = 2 V, dcorresponding transfer (Ids � Vg) curve at |Vds| = 2 V, demonstrating n-type behavior.
redox activities may be harnessed for bioremediation (treatmentof pollutants) and biotechnology applications including biosensors,biocatalysis and electricity generation in microbial fuel cells[35,37]. With the increased interest in biomineralization and bio-genic materials [1], a number of studies have also demonstratedthe control of microbial redox activity to synthesize uniquenanomaterials including magnetic Fe3O4 [38], microtubular Fe2O3
oxides as potential Li-ion battery anodes [39,40], goethite nano-wires [41], Se/Te nanoparticles [42,43], Ag2S [3], and precious met-als such as Au nanoparticles [4,35,44]. Recent reports alsodemonstrated the controlled synthesis of Ag and Cu nanostruc-tures using silver-resistant Morganella species, and an uptake-reduction-efflux mechanism (broadly similar to the ars arsenicresistance pathway available in strain ANA-3) was proposed toexplain the biosynthesis process [45,46]. Most relevant to thisstudy, Lee et al. recently demonstrated a pioneering method forthe production of novel filamentous As–S nanostructures using anumber of Shewanella strains, including strain HN-41, to reduceAs(V) and S2O3
2� under anaerobic conditions [5,6]. These studiesinspired us to explore the synthesis of arsenic sulfide nanofibersby another Shewanella strain, the model As(V) reducer strainANA-3, and to study the applicability of these chalcogenide nano-structures as building blocks in potential nanoscale devices. Ourprimary objectives were to elucidate the nanofiber growth mecha-nism, enhance the precipitation kinetics, characterize the struc-tural/optoelectronic properties and to demonstrate functionalFETs based on these biogenic nanomaterials.
Many Shewanella strains can use thiosulfate as a respiratoryelectron acceptor, thereby reducing it to sulfide [37]. Successfulprecipitation of arsenic sulfide also requires the reduction ofAs(V) to As(III). We hypothesized that the use of strain ANA-3,capable of expressing both the ars and arr genes encoding thedetoxification and respiratory As(V) reduction systems [18,19],will result in faster synthesis of As2S3 nanofibers relative to strainHN-41 whose genome contains only the ars detoxification pathway[47]. Indeed, visible precipitate was observed within 19 ± 2.5 h for
The dependence of the drain-to-source current vs. voltage (Ids � Vds) at different Vg
emonstrating p-type behavior. (C) Ids � Vds for Vg from �40 to 40 V and (D) the
ANA-3 cultures incubated with arsenate and thiosulfate; thisresponse is significantly faster than previously observed in otherShewanella strains [6], and our own results with S. oneidensisMR-1 under identical conditions (Table 1). MR-1 does not havean identifiable respiratory As(V) reductase (arrA) and its putativedetoxification As(V) reductase (arsC) has <60% sequence similaritywith arsC from ANA-3 [6].
No precipitation was observed in non-inoculated control mediacontaining As(V) and S2O3
2� or As(V) and S2� [5]. These abiotic con-trols confirm that active microbial cultures are necessary for theprecipitation of As2S3 nanofibers from As(V) and S2O3
2�. With thisin mind, we were quite surprised to observe the delayed formationof nanofibers by ANA-3 mutants disrupted in one or both of thedetoxification and respiratory As(V) reduction pathways (Table 1).In particular, the ability of the ARM1 (missing both the arsC andarrA As(V) reductases) strain to synthesize As2S3 nanofibers,despite being previously shown ([19] and confirmed here) not toreduce As(V), suggests an additional microbially influenced mech-anism of As(V) reduction under our experimental conditions. Toinvestigate this mechanism of delayed nanofiber synthesis bystrains disrupted in As(V) reduction, we performed IC/AFSmeasurements of As ion concentrations in the abiotic controlcontaining As(V) (5 mM) and S2� (10 mM as Na2S), and observeda modest slow reduction of As(V) to As(III) despite the lack ofprecipitate (Fig. S4, 0.5 mM As(III) observed after 6.5 days). Thisfinding is consistent with previous studies of abiotic As(V) reduc-tion by dissolved S2�, demonstrating slow kinetics at neutral pH[48], although we cannot rule out possible reductive contributionsfrom additional chemicals in the growth medium. The abioticreduction of arsenate observed here suggests that microbial strainsunable to reduce As(V) directly, such as ARM1 [19], can influencepartial As(V) reduction indirectly after producing S2� from therespiratory reduction of S2O3
2�. This indirect mechanism has slowerkinetics, however, than direct As(V) reduction by wild-type ANA-3where the presence of As reductases (ArrA and ArsC) was corre-lated with significantly faster precipitation rates (Table 1).
In both the direct and indirect mechanisms, reduced S2� andAs(III) are necessary for the precipitation of the As–S nanofibers.An additional abiotic control containing reduced As and S (As(III)and S2�, as Na2S) did result in the precipitation of nanofibers butat a significantly lower rate and smaller yield compared to incuba-tion of As(V) and S2O3
2� with strain ANA-3, as measured by As2S3
absorption at 375 nm (Fig. S5). In other words, bacterial synthesisof nanofibers was faster and more effective than the abioticprecipitation of already reduced As and S. These results indicatethat, beyond an active role in reduction, Shewanella cells play anadditional role in catalyzing the nucleation of nanofibers fromreduced As(III) and S2�.
We observed a significant reaction yield with strain ANA-3; theAs2S3 precipitate reached >80% of the maximum theoretical yieldof each incubation after 60 days (Fig. 4). This theoretical yield isestimated by considering that the original 5 mM arsenate can yielda maximum 2.5 mM of As2S3 per liter in excess S conditions,and using the As2S3 molecular weight of 246 g mol�1, resulting ina maximum yield of 49.2 mg for each 80 ml incubation(0.62 mg ml�1). In addition to evaluating the overall precipitationyield, we characterized the nanofiber growth rates and precipita-tion rates during active synthesis. Our in situ observations(Fig. 3) revealed a decline in the growth rates of individual nanof-ibers with increased incubation time (from �10 to �3 nm min�1
over �5 h). This observation is consistent with a limitation broughtabout by the diminishing concentration of reactants in solution,namely As(III) and S2�, as they get incorporated into the nanofibersover time. By measuring the temperature dependence of the pre-cipitation in bulk cultures over time (Fig. 4), we were able todescribe the initial precipitation rates with simple thermally
activated Arrhenius kinetics, and estimated an activation energyof 78.9 ± 0.8 kJ mol�1. Remarkably, this activation energy from ourmicrobially driven process is consistent with metal chalcogenidethin film growth (85 and 68 kJ mol�1 for CdS and CuxS, respec-tively) by abiotic chemical bath deposition techniques [49,50].The biological synthesis process, however, avoids the use of toxicsolvents and harsh reaction conditions (e.g. temperature and pH)typical of well-established physicochemical strategies [2].
Taken collectively, the EPMA stoichiometry, TEM imaging,diffraction patterns and the indirect (n = ½) 2.37 ± 0.05 eV bandgap, in excellent agreement with amorphous As2S3 (n = ½, Eg,indir =2.36 eV [51]) but not the direct band gap of crystalline As2S3 (n = 2,Eg,dir = 2.6 eV [52]), point to the nanofibers as being primarilyamorphous As2S3 (Figs. 5–7). However, we also observed occa-sional diffraction patterns containing peaks while scanningsamples in the TEM, suggesting a crystalline fraction of theprecipitate, which was subsequently confirmed by XRD (Fig. 8).This XRD pattern matched the expected patterns of b-As4S4 andAs8S9 (alacránite), with both microdomains possibly contained inthe previously characterized As8S9-x series [29]. While the presenceof these crystalline stoichiometries is clear, they appear to repre-sent only a small fraction of the precipitate, given the overall38:62 ± 3% As:S stoichiometry measured by EPMA. Until recently,it was thought that b-As4S4 and As8S9 are only produced abioticallyat higher temperatures and hydrothermal conditions. Two recentstudies, however, demonstrated the formation of these mineralsthrough the action of newly isolated As(V)-reducing bacteria[53,54]. The present study adds a new example of the formationof these minerals by bacteria, specifically Shewanella.
The construction of NF-FETs allowed us to both characterize theelectronic properties and test the applicability of microbially syn-thesized nanofibers in a device context. The Ids � Vds behavior,especially in the low-bias regime, was roughly linear, suggestingnear-ohmic contacts with the electrodes [31]. 62% of the devicestested exhibited the p-type behavior of decreased conductance asVg becomes more positive. This is explained by the depletion ofholes as the p-type semiconductor bands (bent up at the nanofi-ber–metal interface) are lowered at Vg > 0, thereby suppressingthe conductivity [31,34]. The observation of p-type behavior inthe majority of devices is again consistent with the behavior ofamorphous chalcogenide glasses, including As2S3, that typicallyexhibit hole conduction; this is in contrast to crystalline As2S3
where electrons are more mobile [55,56]. 24% of the FETs demon-strated the reverse characteristics of increased conductance atVg > 0; this behavior is explained by the accumulation of electronsin n-type nanofibers as the bands (bent down at the nanofiber-metal interface) are lowered [31,34]. The heterogeneity of thenanofibers, in terms of dopant type, may reflect the complexityof the bacterial growth medium containing multiple elements inaddition to As and S (see Materials and methods section). Whileany residual salt and nutrient content appears to be below thedetection limit of EDS (�0.1 wt.% [57]) as a result of the extensivewashing procedure (Section 2.2), it is possible that trace elementsmay impact the electronic properties. Specifically, the residual ele-ments may be responsible for the observed n-type doping in a frac-tion of the tested As2S3 nanofibers, which are otherwise known toexhibit p-type conduction intrinsically [55]. These observationsalso raise the interesting prospect of ‘biological doping’ that mayinfluence, or tune, the properties of the biogenic nanomaterialsdepending on the constituents of the growth medium.
In addition to the p-type and n-type behavior of individualnanofibers, a number of devices exhibited more complex gatedependencies, such as the mixed behavior seen in Fig. S3A,B. Thisunusual gate dependence was explained by a close inspection ofsuch devices at the electrode–fiber interface (Fig. S3C), revealingthat the semiconducting channel consisted of a bundle of
heterogeneous fibers. While the underlying reason for the bundlingof smaller fibers is not yet clear, this phenomenon partiallyexplains the significant diversity of diameters (20–600 nm)observed in our study, since smaller-diameter nanofibers maycoalesce into larger-diameter structures.
It should be noted that no effort was made to improve theoverall device performance by annealing, which may furtherdecrease both the nanofiber and nanofiber–electrode contact resis-tances, since our primary goal was to assess the as-synthesizednanomaterials. From the transconductance of the NF-FETs, weextracted field-effect hole and field-effect electron mobilities of4.3 ± 1.6 � 10�5 and 4.6 ± 2.9 � 10�5 cm2 V�1 s�1 for the p-typeand n-type nanofibers, respectively. The p-type mobility is inexcellent agreement with the previously observed hole mobility(10�4–10�5 cm2 V�1 s�1) in amorphous As2S3 glasses [55,56]. Thebiogenic NF-FET mobility demonstrated here is significantly lowerthan many well-studied FETs based on crystalline or organic semi-conductors [34,58]. While this results in small (pA) on currents andon/off current ratios, it should be noted that the p-type devicesshowed attractive threshold voltages (Vg � 0 V). Future applica-tions may also rely on the 2.37 eV optical band gap of the biogenicAs2S3, which compares well to the 2.42 eV band gap of CdS, a tech-nologically important chalcogenide widely used in commercialphotoresistors, and actively pursued for applications in photonics,photovoltaics and transistors [59]. For this reason, we are currentlyinvestigating the use of biogenic As2S3 as the active component inphotoelectrochemical cells. Our results motivate further investiga-tions geared towards improving the electronic and optoelectroniccharacteristics of cheaply and greenly produced biogenicnanomaterials.
5. Conclusions
Biological, including microbial, systems present unique oppor-tunities for the economic and eco-friendly fabrication of nanoma-terials with technologically relevant mechanical, optical,electronic and magnetic functionalities. In this study, we harnessedthe As and S reductase activity of Shewanella sp. strain ANA-3 todrive the high yield precipitation (504 mg per liter of culture) ofextracellular As2S3 nanofibers. The synthesis rates by strainANA-3, which possesses both the detoxification and respiratoryAs(V) reduction pathways, were more rapid than previous observa-tions using other Shewanella strains. Direct in situ observations ofnanofiber growth, and bulk measurements of the precipitationrates, demonstrated thermally activated kinetics with an activationenergy of 79 kJ mol�1. The compositional, structural and optoelec-tronic properties of the nanofibers were characterized by SEM, EDS,EPMA, TEM, XRD and Tauc analysis of UV–vis spectrophotometry,revealing a primarily amorphous As2S3 structure. In addition, wedemonstrated and characterized novel FETs based on individualbiogenic nanofibers, and detailed the carrier type, mobility andswitching behavior of these devices. Given the remarkable techno-logical versatility of chalcogenides, the rapid and high-yield biofab-rication process described here may help realize the potential ofthese nanomaterials in (opto)electronics, sensors, waveguides,photovoltaics and storage devices.
Acknowledgments
The authors acknowledge Andrea Cheung for input into the bac-terial cultivation, Frank Devlin for training and access to XRD, andDr. Tom Yuzvinsky for microfabrication of the pre-patterned Sichips. EPMA measurements were performed at the Caltech GPSdivision analytical facility with the help of Dr. Chi Ma. Wild-typeANA-3 and mutants were kindly provided by Profs. DianneNewman (Caltech) and Chad Saltikov (UCSC). SEM, TEM and FIB
deposition were performed at the USC Center for ElectronMicroscopy and Microanalysis, where John Curulli and Dr. MattMecklenburg provided valuable assistance. We are also gratefulto Guang-Sin Lu and Prof. Jan Amend for help and access to theIC/AFS system. Michael Qian and Brian Zukotynski provided assis-tance in overnight UV–vis measurements. M.E.-N. acknowledgessupport from NASA through the NASA Astrobiology Institute undercooperative agreement NNA13AA92A.
Appendix A. Figures with essential colour discrimination
Certain figures in this article, particularly Figs. 4, 5, 7, 9 and 10are difficult to interpret in black and white. The full colour imagescan be found in the on-line version, at http://dx.doi.org/10.1016/j.actbio.2014.11.005.
Appendix B. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.actbio.2014.11.005.
References
[1] Estroff LA. Introduction: biomineralization. Chem Rev 2008;108:4329–31.[2] Bansal V, Bharde A, Ramanathan R, Bhargava SK. Inorganic materials using
‘unusual’ microorganisms. Adv Colloid Interfac 2012;179:150–68.[3] Suresh AK, Doktycz MJ, Wang W, Moon JW, Gu BH, Meyer HM, et al.
Monodispersed biocompatible silver sulfide nanoparticles: facile extracellularbiosynthesis using the gamma-proteobacterium, Shewanella oneidensis. ActaBiomater 2011;7:4253–8.
[4] Suresh AK, Pelletier DA, Wang W, Broich ML, Moon JW, Gu BH, et al.Biofabrication of discrete spherical gold nanoparticles using the metal-reducing bacterium Shewanella oneidensis. Acta Biomater 2011;7:2148–52.
[5] Lee JH, Kim MG, Yoo BY, Myung NV, Maeng JS, Lee T, et al. Biogenic formationof photoactive arsenic-sulfide nanotubes by Shewanella sp strain HN-41. ProcNatL Acad Sci USA 2007;104:20410–5.
[6] Jiang S, Lee JH, Kim MG, Myung NV, Fredrickson JK, Sadowsky MJ, et al.Biogenic formation of As-S nanotubes by diverse Shewanella strains. ApplEnviron Microbiol 2009;75:6896–9.
[7] Jiang S, Liu F, Kim MG, Lim JH, Lee KJ, Choa YH, et al. Synthesis of chalcogenideternary and quaternary nanotubes through directed compositional alterationsof bacterial As-S nanotubes. J Mater Chem 2011;21:10277–9.
[8] Greer AL, Mathur N. Materials science—changing face of the chameleon. Nature2005;437:1246–7.
[9] Sundaram SK, Johnson BR, Schweiger MJ, Martinez JE, Riley BJ, Saraf LV, et al.Chalcogenide glasses and structures for quantum sensing. Quantum SensNanophoton Devices 2004;5359:234–45.
[10] Krecmer P, Moulin AM, Stephenson RJ, Rayment T, Welland ME, Elliott SR.Reversible nanocontraction and dilatation in a solid induced by polarized light.Science 1997;277:1799–802.
[11] Stuchlik M, Elliott SR. All-optical actuation of amorphous chalcogenide-coatedcantilevers. J Non-Cryst Solids 2007;353:250–62.
[14] Rao CNR, Govindaraj A. Synthesis of inorganic nanotubes. Adv Mater2009;21:4208–33.
[15] Myers CR, Nealson KH. Bacterial manganese reduction and growth withmanganese oxide as the sole electron-acceptor. Science 1988;240:1319–21.
[16] Nealson KH, Belz A, McKee B. Breathing metals as a way of life: geobiology inaction. Antonie Van Leeuwenhoek Int J Gen Mol Microbiol 2002;81:215–22.
[17] Saltikov CW, Cifuentes A, Venkateswaran K, Newman DK. The arsdetoxification system is advantageous but not required for As(V) respirationby the genetically tractable Shewanella species strain ANA-3. Appl EnvironMicrobiol 2003;69:2800–9.
[18] Saltikov CW, Newman DK. Genetic identification of a respiratory arsenatereductase. Proc Natl Acad Sci USA 2003;100:10983–8.
[19] Saltikov CW, Wildman RA, Newman DK. Expression dynamics of arsenicrespiration and detoxification in Shewanella sp strain ANA-3. J Bacteriol2005;187:7390–6.
[20] Rosen BP. Biochemistry of arsenic detoxification. Febs Lett 2002;529:86–92.[21] Krafft T, Macy JM. Purification and characterization of the respiratory arsenate
reductase of Chrysiogenes arsenatis. Eur J Biochem 1998;255:647–53.[22] Bretschger O, Obraztsova A, Sturm CA, Chang IS, Gorby YA, Reed SB, et al.
Current production and metal oxide reduction by Shewanella oneidensis MR-1wild type and mutants. Appl Environ Microbiol 2007;73:7003–12.
[23] El-Naggar MY, Wanger G, Leung KM, Yuzvinsky TD, Southam G, Yang J, et al.Electrical transport along bacterial nanowires from Shewanella oneidensisMR-1. Proc Natl Acad Sci 2010;107:18127–31.
[24] Leung KM, Wanger G, El-Naggar MY, Gorby Y, Southam G, Lau WM, et al.Shewanella oneidensis MR-1 bacterial nanowires exhibit p-type, tunableelectronic behavior. Nano Lett 2013;13:2407–11.
[25] Tauc J, Menth A, Wood DL. Optical and magnetic investigations of localizedstates in semiconducting glasses. Phys Rev Lett 1970;25:749–52.
[26] Tauc J, Menth A. States in the gap. J Non-Cryst Solids 1972;8–10:569–85.[27] Clark AH. Alpha-arsenic sulfide, from Mina-Alacran, Pampa-Larga, Chile. Am
Mineral 1970;55:1338.[28] Burns PC, Percival JB. Alacranite, As4S4: a new occurrence, new formula, and
determination of the crystal structure. Can Mineral 2001;39:809–18.[29] Bonazzi P, Bindi L, Popova V, Pratesi G, Menchetti S. Alacranite, AS(8)S(9):
structural study of the holotype and re-assignment of the original chemicalformula. Am Mineral 2003;88:1796–800.
[30] Bonazzi P, Bindi L, Olmi F, Menchetti S. How many alacranites do exist? Astructural study of non-stoichiometric As8S9-x crystals. Eur J Mineral2003;15:283–8.
[31] Cui Y, Duan XF, Hu JT, Lieber CM. Doping and electrical transport in siliconnanowires. J Phys Chem B 2000;104:5213–6.
[32] Mane RS, Todkar VV, Lokhande CD. Low temperature synthesis ofnanocrystalline As2S3 thin films using novel chemical bath deposition route.Appl Surf Sci 2004;227:48–55.
[33] Sze SM. Physics of semiconductor devices. 2nd ed. New York: Wiley; 1981.[34] Lu W, Xie P, Lieber CM. Nanowire transistor performance limits and
applications. IEEE T Electron Dev 2008;55:2859–76.[35] Gadd GM. Metals, minerals and microbes: geomicrobiology and
bioremediation. Microbiol-Sgm 2010;156:609–43.[36] Kowalchuk GA, Jones SE, Blackall LL. Microbes orchestrate life on Earth. Isme J
et al. Towards environmental systems biology of Shewanella. Nat RevMicrobiol 2008;6:592–603.
[38] Lovley DR, Stolz JF, Nord GL, Phillips EJP. Anaerobic production of magnetite bya dissimilatory iron-reducing microorganism. Nature 1987;330:252–4.
[39] Hashimoto H, Fujii T, Kohara S, Asaoka H, Kusano Y, Ikeda Y, et al. Amorphousstructure of iron oxide of bacterial origin. Mater Chem Phys 2012;137:571–5.
[40] Hashimoto H, Kobayashi G, Sakuma R, Fujii T, Hayashi N, Suzuki T, et al.Bacterial nanometric amorphous Fe-based oxide: a potential lithium-ionbattery anode material. ACS Appl Mater Interfac 2014;6:5374–8.
[41] Jiang S, Kim MG, Kim IY, Hwang SJ, Hur HG. Biological synthesis of free-standing uniformed goethite nanowires by Shewanella sp HN-41. J MaterChem A 2013;1:1646–50.
[42] Oremland RS, Herbel MJ, Blum JS, Langley S, Beveridge TJ, Ajayan PM, et al.Structural and spectral features of selenium nanospheres produced by se-respiring bacteria. Appl Environ Microbiol 2004;70:52–60.
[43] Baesman SA, Bullen TD, Dewald J, Zhang DH, Curran S, Islam FS, et al.Formation of tellurium nanocrystals during anaerobic growth of bacteria thatuse te oxyanions as respiratory electron acceptors. Appl Environ Microbiol2007;73:2135–43.
[44] Reith F, Etschmann B, Grosse C, Moors H, Benotmane MA, Monsieurs P, et al.Mechanisms of gold biomineralization in the bacterium Cupriavidusmetallidurans. Proc Natl Acad Sci USA 2009;106:17757–62.
[46] Ramanathan R, Field MR, O’Mullane AP, Smooker PM, Bhargava SK, Bansal V.Aqueous phase synthesis of copper nanoparticles: a link between heavy metalresistance and nanoparticle synthesis ability in bacterial systems. Nanoscale2013;5:2300–6.
[47] Kim DH, Jiang S, Lee JH, Cho YJ, Chun J, Choi SH, et al. Draft genome sequence ofShewanella sp strain HN-41, which produces arsenic-sulfide nanotubes. JBacteriol 2011;193:5039–40.
[48] Rochette EA, Bostick BC, Li GC, Fendorf S. Kinetics of arsenate reduction bydissolved sulfide. Environ Sci Technol 2000;34:4714–20.
[49] Dona JM, Herrero J. Chemical bath deposition of CdS thin films: an approach tothe chemical mechanism through Study of the film microstructure. JElectrochem Soc 1997;144:4081–91.
[50] Vas-Umnuay P, Chang CH. Growth kinetics of copper sulfide thin films bychemical bath deposition. Ecs J Solid State Sc 2013;2:P120–9.
[51] Mott NF, Davis EA. Electronic processes in non-crystalline materials. 2nded. Oxford: Clarendon Press; 2012.
[52] Zallen R, Blossey D. The optical properties, electronic structure, andphotoconductivity of arsenic chalcogenide layer crystals. In: Lee P, editor.Optical and Electrical Properties. Springer: Netherlands; 1976. p. 231–72.
[53] Ledbetter RN, Connon SA, Neal AL, Dohnalkova A, Magnuson TS. Biogenicmineral production by a novel arsenic-metabolizing thermophilic bacteriumfrom the Alvord Basin, oregon. Appl Environ Microbiol 2007;73:5928–36.
[54] Mumford AC, Yee N, Young LY. Precipitation of alacranite (As8S9) by anovel As(V)-respiring anaerobe strain MPA-C3. Environ Microbiol 2013;15:2748–60.
[55] Tanaka K. Wrong bond in glasses: a comparative study on oxides andchalcogenides. J Optoelectron Adv Mater 2002;4:505–12.
[56] Tanaka K, Shimakawa K. Amorphous Chalcogenide Semiconductors andRelated Materials. New York: Springer Science+Business Media; 2011.
[57] Electronic Device Failure Analysis Society Desk Reference Committee.Microelectronics Failure Analysis: Desk Reference. 5th ed. Materials Park,OH: ASM International; 2004.
[58] Grozema FC, Siebbeles LDA. Mechanism of charge transport in self-organizingorganic materials. Int Rev Phys Chem 2008;27:87–138.
[59] Kar S, Chaudhuri S. Cadmium sulfide one-dimensional nanostructures:synthesis, characterization and application. Synth React Inorg M 2006;36:289–312.











![[Chapter III] Basic Knowledge of Discrete Semiconductor ......transistors (IGBTs) Power transistors (2SAxx,2SBxx,2SCxx,2SDxx, TTAxx,TTBxx,TTCxx,TTDxx) Types of Transistors Transistors](https://static.documents.pub/doc/80x56/5e766014341a1a707d5f4c34/chapter-iii-basic-knowledge-of-discrete-semiconductor-transistors-igbts.jpg)












