Formation of Bacterial FlagellaI. Demonstration of a Functional Flagellin Pool in Spirillum serpens
and Bacillus subtilisR. J. MARTINEZ AND E. Z. GORDEE
Department ofBacteriology, University of California, Los Angeles, California
Received for publication 24 September 1965
ABSTRACT
MARTINEZ, R. J. (University of California, Los Angeles), AND E. Z. GORDEE.Formation of bacterial flagella. I. Demonstration of a functional flagellin pool inSpirillum serpens and Bacillus subtilis. J. Bacteriol. 91:870-875. 1966-Exponentiallygrowing cultures of Spirillum serpens and Bacillus subtilis regained motility andflagella within one generation after mechanical deflagellation. Regeneration offlagella occurred in both cultures in the presence of chloramphenicol at concentra-tions shown to inhibit flagellin synthesis. Cells labeled with C'4-amino acids regen-erated radioactive flagella in the presence of chloramphenicol. A conditional mutantof S. serpens (T-45) was isolated. This strain did not produce flagella when grown at45 C, but formed the organelles upon temperature shift to 30 C, even in the presenceof chloramphenicol. A reduction of intracellular antibody-precipitable flagellincounts in labeled S. serpens T-45 occurred concomitant with the generation offlagella at 30 C. The data suggest that the flagella of S. serpens and B. subtilis areformed from a pool of intracellular flagellin proteins.
The bacterial flagellum, because of its relativechemical (16) and structural simplicity (9, 10),ease of isolation and purification (12), and itsrapid rate of synthesis (15), offers a convenienttest system to investigate the biosynthesis of sub-cellular structures. Pertinent to the mechanism ofbiosynthesis of the bacterial flagellum is the pos-sible existence of a functional pool of flagellarprecursor proteins, the flagellins (3). Kerridge (7)reported that Salmonella typhimurium regeneratedflagella under conditions where protein or ribo-nucleic acid (RNA) synthesis, or both, were in-hibited. His results suggest either (i) that flagellaare assembled from a pool of flagellin or (ii) thatthe biosynthesis of the protein subunits is resistantto the conventional inhibitors of protein synthe-sis. The experiments described in this communica-tion were designed to distinguish between thesetwo possibilities.
MATERIALS AND METHODS
Organisms and growth media. The organisms usedin this study were Spirillum serpens, a bipolarlyflagellated bacterium; a mutant of S. serpens, S.serpens T-45, which is nonflagellated at 45 C but whichforms flagella upon temperature shift to 30 C; andBacillus subtilis 168-15, a peritrichously flagellatedorganism requiring tryptophan and uracil for growth
(provided by W. R. Romig). The spirilla were grownin nutrient broth-0.3% yeast extract (N-Y) in ashaker water bath maintained at 30 C for the parentculture and at 45 C for the mutant. Rapid temperatureshifts were made by diluting exponentially growingcultures of S. serpens T-45 with two volumes ofmedium at 25 C. B. subtilis was grown in a shakerwater bath at 37 C in a medium containing (per liter):glucose, 5.0 g; (NH4)2SO4, 2.0 g; K2HPO4, 0.14 g;KH2PO4, 0.06 g; MgSO4c7H20, 0.2 g; KCl, 7.5 g;sodium citrate, 1.0 g; and 0.05 M tris(hydroxymethyl)-aminomethane (Tris)-HCl buffer (pH 7.2); supple-mented with 15 jg/ml of uracil, 20 ,ug/ml of L-tryptophan, and 0.1% Vitamin Free Casamino Acids.The organisms were used while in the exponentialphase of growth.
Incorporation of labeled amino acids into flagellinand flagella. The incorporation of C14- or H3-labeledamino acids was carried out in N-Y medium diluted1: 5 for the spirilla and in the growth medium for B.subtilis. The labeled amino acids were used at 0.25 to0.5 ,c/ml at different specific activities; incubationswere in a shaker water bath at temperatures specifiedfor the individual experiments. At timed intervals,samples of the cultures were rapidly chilled to 0 C anddeflagellated by mechanical shearing in a ServallOmni-mixer at 24,000 rev/min for 1 min; the cellswere separated from the flagella by centrifugation(6,000 X g for 10 min), and the flagella-containingsupematant fluid was treated with an excess of
specific antiflagellar antibody. The antigen-antibodycomplex was harvested by centrifugation, washed, andresuspended in 5% trichloroacetic acid containing 1%Casamino Acids (8). The trichloroacetic acid pre-cipitate was collected on a Gelman type GM-6 mem-brane filter (2.5-cm diameter) and washed, and itsradioactivity was determined with a liquid scintillationspectrometer by use of appropriate channels. In-corporation of labeled amino acids into cellular pro-teins was determined by a procedure similar to thatdescribed by Hartwell and Magasanik (6).
Determination of flagella regeneration. Fourmethods were used for measuring the formation offlagella: (i) resumption of motility by direct phase-contrast microscopic observation, (ii) detection offlagella by electron microscopy in either phospho-tungstic acid- or uranyl acetate-stained preparations,(iii) appearance of specific flagellar antigen with theagglutination-inhibition technique (12), and (iv)appearance of flagellar antibody-precipitable countsfrom radioactive cells. The latter assay was used byKerridge and was found to give only a minimalnumber of flagellar-associated counts (8).
Materials. Uniformly C'4-labeled L-leucine, lysine,and valine, and H3-labeled lysine were obtained fromNew England Nuclear Corp., Boston, Mass., or VolkRadiochemical Co., Burbank, Calif. Chloramphenicolwas obtained from Worthington Biochemical Corp.,Freehold, N.J. Antisera were prepared as previouslydescribed (12).
RESULTS
Exponentially growing cultures of S. serpensand B. subtilis, when deflagellated and subculturedinto their respective media, regenerated flagellawithin one doubling time. Similar results wereobtained when S. serpens T-45 was incubated at30 C; i.e., within one doubling time (70 min at30 C) approximately 40 to 50% of the cellsshowed motility. To determine whether nondivid-ing cells of S. serpens regained motility after de-flagellation, chloramphenicol was used at concen-trations which inhibit cell division (20 to 40,ug/ml). Under these conditions, a portion of thepopulation regained motility, flagellar antigenwas detected, and flagellated cells were observedin electron micrographs (Fig. 1). S. serpens T-45,grown at 45 C, was found to behave in a similarfashion when incubated at 30 C (Table 1); i.e.,in the presence of 40 Ag/ml of chloramphenicol,approximately 50% of the viable cells becamemotile after 2 hr of incubation.To ascertain whether chloramphenicol inhibits
the biosynthesis of flagellin, the following experi-ments were carried out. Exponentially growingcultures of S. serpens were deflagellated, the cellswere separated from the flagella by centrifugation
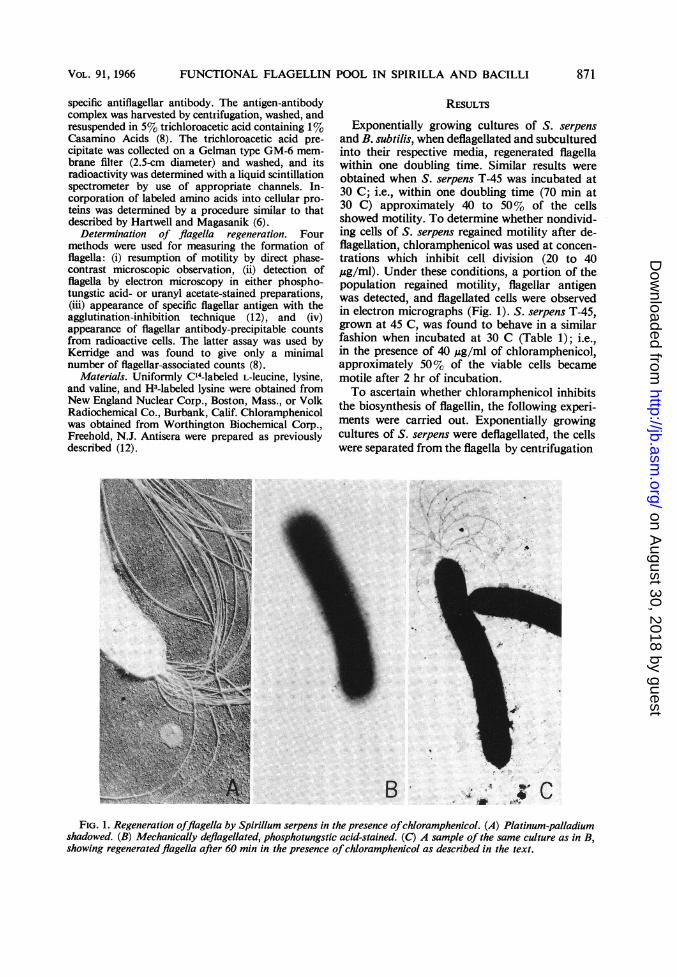
BFIG. 1. Regeneration offlagella by Spirillum serpens in the presence ofchloramphenicol. (A) Platinum-palladium
shadowed. (B) Mechanically deflagellated, phosphotungstic acid-stained. (C) A sample of the same culture as in B,showing regeneratedflagella after 60 min in the presence ofchloramphenicol as described in the text.
TABLE 1. Effect of chloramphenicolon the growth of Spirillum
serpens T-45*
Viable-cell count/ml Per centmotile cells
Condition after 2 hrCondition ~~~~~~~at30 C
Initial Final (based ontotal count)
Control......... 1.23 X 108 3.2 X 108 30-40Chlorampheni-
col treated.... 1.33 X 108 8.0 X 107 25-35
* Exponential-phase cultures of S. serpens T-45growing at 45 C were stepped down to 30 C by di-lution with two parts of cold medium; viablecounts were made immediately and after incuba-tion at 30 C for 2 hr, and the per cent motile cellswas determined at the end of the experiment bydirect phase-microscopic observation.
and the cell paste was resuspended in diluted N-Ybroth. To this culture, chloramphenicol wasadded to a final concentration of 40 ,ug/ml, andthe culture was incubated at 30 C with shaking for20 min. Valine-, lysine-, or leucine-C'4 was thenadded, and the incubation was continued foranother 2 hr. During this latter period, flagellawere regenerated, as evidenced by the resumptionof motility in 25 to 40% of the population. Thenewly formed flagella were removed from thecells, and radioactivity of the cell paste and theflagella was determined (Table 2). It can be seenin Table 2 that chloramphenicol inhibited aminoacid incorporation into flagellar proteins as wellas into other cellular proteins; the extent of in-hibition into cellular proteins ranged from 65 to98% and into flagellin proteins from 76 to 97%in the three experiments shown.
In an attempt to test directly for a flagellin pool,exponentially growing cultures of S. serpens wereincubated with Cl4-valine for 2 hr. At this timethe labeled amino acid was diluted with an excessof nonradioactive compound (1% CasaminoAcids); the culture was deflagellated, and the cellswere separated from the flagella. The deflagellatedculture was permitted to regenerate flagella byincubation in diluted N-Y broth containing 40,ug/ml of chloramphenicol. At intervals, 5-mlsamples (-5 X 108 cells per milliliter) were re-moved and deflagellated, and the radioactivity inthe flagella was determined. Assuming that chlor-amphenicol inhibits the synthesis of flagellin, as issuggested from Table 2, then it is reasonable toassume that antibody-precipitable counts, whichappear during the incubation with chloramphen-icol, arise from flagellin synthesized in the initialincubation with the labeled amino acids. After 2hr of incubation with chloramphenicol, the anti-
TABLE 2. Inhibition of amino acid incorpo-rationt inito cellular and flagellar
protein ofSpirillum serpensby chloramphenicol
Labeled precursor
L-Leucine
L-Lysine
L-Valine
Condition
ControlChloram-phenicol
ControlChloram-phenicol
ControlChloram-phenicol
aa
0.-
2,040
639
993
20
1,330
461
* Determined in 5 ml of culture.
00.
v ,=
6D-
69
98
65
co
D._
700
138
33
8
56
2
0
*:5W._8.0
PS
80
76
97
body-precipitable counts in flagella were essen-tially equal to those obtained in control cultureswithout the drug (Table 3). Similar experimentswith B. subtilis 168-15 have shown essentially thesame results.The data suggest that cells of S. serpens and B.
subtilis possess a pool of preformed flagellin pro-teins which is functional and serves as a precursorof the ordered flagellar structure.To further test this hypothesis, a double-label
experiment was carried out as follows. Log-phasecultures of B. subtilis were incubated with 0.25,uc/ml of C'4-valine and 12.5 ,uc/ml of H3-lysinefor 2 hr. Incorporation of the labeled amino acidsinto proteins was stopped by adding an excess ofCasamino Acids and centrifuging the cells. Thecells were resuspended in phosphate-bufferedsaline and deflagellated, and the isotopes werefurther diluted with Casamino Acids. After sep-arating the cells from the flagella, the cells weresuspended in growth medium supplemented with1% Casamino Acids containing 12.5 ,uc/ml ofH3-lysine. Chloramphenicol was added to one halfof the culture (40 ,g/ml, final concentration).Both cultures were incubated with shaking at37 C; samples were removed at intervals and de-flagellated, the cells were separated from theflagella, and the C14 and H3 in the antigen-anti-body pellet were determined (Table 4). In thecontrol culture incubated without chlorampheni-col, there was a decrease in the C14-H3 ratio duringthe 1.5-hr incubation with H3-lysine, implyingthat flagellin was constantly being synthesized,and that the newly synthesized protein mixedfreely with the pre-existing pool of flagellin. In thechloramphenicol-treated culture, the C14-H3 ratio
remained relatively constant after the first 30 min.The incorporation of H3-lysine during the first30 min in the chloramphenicol-treated culturemay be ascribed to the lack of preincubation withthe drug prior to the addition of the H3 aminoacid. These experimental findings support thehypothesis that a flagellin pool exists in B. subtilis168-15 and S. serpens cells.
S. serpens T-45, which is nonflagellated whengrown at 45 C, was also found to possess a poolof flagellin proteins at the elevated temperature;this pool, upon temperature shift to 30 C, servedas a precursor to the ordered flagellar structure(Table 5). The number of antibody-precipitablecounts obtained after 3 hr of incubation withchloramphenicol at 30 C was similar to that ob-tained in the control culture incubated withoutchloramphenicol.
Since we are proposing that flagella are formedfrom a pool of flagellin in the cells, it should bepossible to demonstrate a reduction in the internalflagellin pool of S. serpens T-45 as a result of
TABLE 5. Formationi of radioactive flagellaat 30 C from prelabeled flagellinby Spitillum serpenis T-45 in thepresenice of chloramphenicol*
Time of incubation at 30 C Count/mi in flagellatwith chloramphenicol
Ihr
1 2,2212 1,2792.5 2,5623 3,5863 t 3,039
* Log-phase cultures of S. serpenis T-45 wereincubated in 1:5 N-Y broth containing 0.5 uc/mlof C'4-valine for 2 hr at 45 C. After isotope dilu-tion, the cultures were incubated at 30 C in N-Ybroth containing 40 ,ug/ml of chloramphenicol.Samples were removed at intervals and deflagel-lated, and the radioactivity in the flagella was de-termined.
t Determined in 5 ml of culture. The zero-timecounts (182) have been subtracted.
t Control (no chloramphenicol).
temperature shift and flagella production in thepresence of chloramphenicol. The following ex-periment was designed to test this possibility. S.serpens was incubated in diluted N-Y broth con-taining 0.5 ,uc/ml of C'4-lysine for 3 hr at 45 C.After diluting the isotope, a sample of the culturewas lysed with lysozyme-ethylenediaminetetra-acetate (12), and the lysate was centrifuged at100,000 X g for 1.5 hr. The supernatant fluid wastreated with antiflagellin antiserum, and the anti-body precipitable counts were determined. Anequal portion of the culture was permitted to formflagella at 30 C in the presence of chlorampheni-col; the flagella were then removed from the cells,precipitated with antibody, and counted for radio-activity. The deflagellated cells were washed andlysed as above, and the 100,000 X g supernatantfluid was similarly treated with antiflagellin anti-serum and was counted. It is apparent (Table 6)that there was a drastic reduction (88%;7) in theintracellular antibody-precipitable counts con-comitant with the generation of flagella at 30 C.We interpret this to mean that S. serpens T-45 iscapable of producing a pool of flagellin at 45 C,but is incapable of organizing the protein sub-units to the ordered structure at the elevated tem-perature.
DISCUSSION
The data presented support the view that theflagella of S. serpens and B. subtilis 168-15 areaggregated from a pool of intracellular flagellinproteins. This conclusion is based on the following
TABLE 6. Reduction offlagellin antibody-precipitable counts from prelabeled
Spirillum serpens T-45 aftertemperature shift and flagella
formation
Sample Count/min inantigen-antibody pellet*
45 C cell lysate ............. 52245 C - 30 C cell lysate ..... 6345 C 30 C flagella ....... 839
* Determined in 5 ml of culture.
observations. (i) In the presence of chlorampheni-col at concentrations which inhibit growth, thereis a resumption of motility, appearance of specificflagellar antigen, and regeneration of flagella asseen in electron micrographs. (ii) Cells prelabeledwith C14-amino acids produce radioactive flagellain the presence of chloramphenicol. (iii) A reduc-tion of intracellular antibody-precipitable flagellincounts in labeled S. serpens T-45 occurs concom-itant with the generation of flagella at 30 C in thepresence of chloramphenicol.To explain the formation of flagella under con-
ditions which inhibit flagellin synthesis, two proc-esses must be involved which can be schematicallyrepresented as follows:
Amino acids - (-)- 4 flagellin flagella
Chloramphenicol presumably inhibits only thefirst process; thus, flagella can be synthesized inthe presence of the drug provided that a pool offlagellin is present in the cell.
Weinstein, Koffler, and Maskowitz (Bacteriol.Proc., p. 63, 1960) and Kerridge (8) examined byserological means Proteus vulgaris and Salmonellatyphimurium, respectively, for the presence offlagellin. Both of these groups were successful indemonstrating the existence of an antigen capableof reacting with antisera (prepared against flagellaor flagellin) in the soluble fraction of cells. Ker-ridge (8), however, has cast some doubt on thevalidity of interpreting these data as evidence forthe existence of a flagellin pool in Salmonella.Further, from measurement of the kinetics ofisotope incorporation into the flagella of Sal-monella, Kerridge (8) has concluded that a func-tional pool of flagellin proteins is probably notpresent in these organisms. The divergence be-tween our results and those reported by Kerridgemay be due to significant differences in experi-mental approach and to the different organismsemployed. By using procedures similar to ours,however, Brinton (personal communication) wasable to demonstrate that the pih of Escherichiacoli may be removed by mechanical means and
regenerate in the presence of chloramphenicol.Further, it appears that the pool of pili precursorproteins is considerable, since depilation and sub-sequent regeneration take place several times inthe presence of chloramphenicol. It is possiblethat other bacterial organelles, besides pili andflagella, may be assembled from pools of pre-cursor proteins.
Recently, Hartwell and Magasanik (6) demon-strated that B. subtilis produces the inducible en-zyme histidase as an inactive protein which canbe converted to the active enzyme by reactionsinsensitive to chloramphenicol inhibition. Theconversion of the histidase precursor to the activeenzyme parallels the conversion of pill and flagellaprecursor proteins to their respective structuresin that the processes apparently do not involvepeptide bond formation.Assuming that the conversion of amino acids
to flagellin occurs via a set of reactions normallyassociated with protein synthesis, we must stillaccount for the second reaction which is necessaryfor the synthesis of the ordered structure, i.e., theaggregation of flagellin molecules to form theflagellum. The experiments of Abram and Koffler(1), Asakura, Eguchi, and Iino (2), Lowy andMcDonough (11), and Nossal, Ada, and Austin(14) show that flagellin molecules may undergoself-assembly to form a structure which is almostindistinguishable from the native flagellum (11).Both Abram and Koffler (1) and Asakura et al.(2) have suggested that the in vivo aggregation ofthe subunits to form the flagellum is a spontane-ous phenomenon. In the experiments describedby Abram and Koffler (1), aggregation of thesubunits appears to be controlled by the pH ofthe solution, as is the final product of the reaction,i.e., straight structures or flagellalike filaments.Asakura et al. (2), on the other hand, reportedthat "seeds" (flagella fragments) are required assites of aggregation and growth of the flagellum,and they compared the process of flagellar growthwith that of crystallization. The fact that sub-units of supermolecular structures can spontane-ously aggregate in vitro to form native structuresdoes not apply only to the flagellins; it has longbeen known that tobacco mosaic virus proteinand RNA aggregate abiologically to form in-fectious particles (5). This ability to aggregatemust be inherent in the structure of the subunits(4).
If, indeed, the in vivo biosynthesis of structuresoccurs by self-assembly independent of biologicalregulation, then several observations must beexplained. Cellular division of a bipolarly flagel-lated cell such as S. serpens results in two daughtercells, each of which has inherited one maternaltuft of flagella. Shortly thereafter, a new tuft of
flagella appears at the nonflagellated pole. Theinherited maternal tuft, however, increases neitherin length nor in numbers; nor is there any disag-gregation at the ends of these flagella permittinguniform growth at both poles. This is verified bythe fact that growth media do not contain flagellaror flagellin antigen. Further, if cultures of spirillaare deflagellated and then allowed to regenerateflagella, both poles of the cells appear to beequally competent in regenerating the organelles.These, and other observations, suggest that abiologically controlled mechanism is involved inaggregating the flagellins to an ordered structure.Such a control mechanism is difficult to envisionif the final step in the production of the flagellumis a spontaneous phenomenon, especially since apool of flagellin has been shown to be presentintracellularly in spirilla and B. subtilis.
Finally, the nonflagellated S. serpens T-45,which we have shown to possess a pool of flagel-lin proteins, but which is incapable of organizingthese proteins to the organelle at 45 C, must beexplained. We have made several observationswhich bear on this problem. It is possible that at45 C flagellin is aggregated to a structure whichis unstable at the elevated temperature and disag-gregates upon extrusion. If this were the case, wewould anticipate finding flagellin antigen ingrowth media. This has not been found, nor arenascent flagella formed upon temperature shift to30 C dissociated upon elevating the temperatureof the culture to 45 C. Rather, we found a gradualloss of motility of the cells concomitant with anincrease in cell numbers, i.e., a dilution of thepre-existing organelles.
In conclusion, we suggest that the in vivo ag-gregation of flagellins (and possibly other subunitproteins) to the ordered structure is not a spon-taneous occurrence, but rather that it is mediatedby a biological organizing principle.
ACKNOWLEDGMENTSThis investigation was supported by the National
Science Foundation grant G-21378, and by theUniversity of California Cancer Research Committee.The expert technical assistance of Marie Hagstedt
and Barbara Hayes in the performance of some ofthese experiments is greatly appreciated.
LITERATURE CITED1. ABRAM, D., AND H. KOFFLER. 1964. Int vitro
formation of flagella-like filaments and otherstructures from flagellin. J. Mol. Biol. 9:168-185.
2. ASAKURA, S., G. EGUCHI, AND T. IINO. 1964.Reconstitution of bacterial flagella in vitro. J.Mol. Biol. 10:42-56.
3. ASTBURY, W. T., E. BEIGHTON, AND C. WEIBULL.1955. The structure of bacterial flagella. Symp.Soc. Exptl. Biol. 9:282-305.
4. CASPER, D. L. D., AND A. KLUG. 1962. Physicalprinciples in the construction of regular viruses.Cold SpIing Harbor Symp. Quant. Biol. 27:1-24.
5. FRAENKEL-CONRAT, H., AND R. C. WILLIAMS.1955. Reconstitution of active tobacco mosaicvirus from its inactive protein and nucleic acidcomponents. Proc. Natl. Acad. Sci. U.S. 41:690-698.
6. HARTWELL, L. H., AND B. MAGASANIK. 1963. Themolecular basis of histidase induction inBacillus subtilis. J. Mol. Biol. 7:401-420.
7. KERRIDGE, D. 1960. The effect of inhibitors on theformation of flagella by Salmonella typhi-murium. J. Gen. Microbiol. 23:519-538.
8. KERRIDGE, D. 1963. Flagellar synthesis in Sal-montella typhimurium: the incorporation ofisotopically-labeled amino acids into flagellin.J. Gen. Microbiol. 33:63-76.
9. KERRIDGE, D., R. W. HORNE, AND A. M.GLAUERT. 1962. Structural components offlagella from Salmoltella typhimurium. J. Mol.Biol. 4:227-238.
10. Lowy, J., AND J. HANSON. 1964. Structure ofbacterial flagella. Nature 202:538-540.
11. Lowy, J., AND M. W. MCDONOUGH. 1964.Structure of filaments produced by re-aggrega-tion of Salmoniella flagellin. Nature 204:125-127.
12. MARTINEZ, R. J. 1963. A method for the purifica-tion of bacterial flagella by ion exchange chro-matography. J. Gen. Microbiol. 33:115-120.
13. MARTINEZ, R. J. 1963. On the nature of thegranules of the genus Spirillum. Arch. Mikro-biol. 44:334-343.
14. NOSSAL, G. J. V., G. L. ADA, AND C. M. AUSTIN.1963. Behaviour of active bacterial antigensduring the induction of the immune response.Nature 199:1257-1262.
15. STOCKER, B. A. D., AND J. C. CAMPBELL. 1959.The effect of nonlethal deflagellation on bac-terial motility and observations on flagellarregeneration. J. Gen. Microbiol. 20:670-685.
16. WEIBULL, C. 1960. Movement, p. 153-205. InI. C. Gunsalus and R. Y. Stanier [ed.], Thebacteria, vol. 1. Academic Press, Inc., NewYork.