Refer to: Locksley R: Fuel utilization in marathons: Implications for performance-Medical Staff Conference, University of California, San Francisco. West J Med 133:493-502, Dec 1980 Medical Staf Conference Fuel Utilization in Marathons: Implications for Performance These discusslons are selected from the weekly staff conferences In the Department of Medllne, University of Callfornla, San Francisco. Taken from transcriptions, they are prepared by Drs. David W. Martin, Jr, Pro- fessor of Medicine, and James L. Naughton, Assistant Professor of Medl- cine, under the directlon of Dr. Lloyd H. Smith, Jr, Professor of Medllne and Chalrman of the Department of Mediclne. Requests for reprlnts should be sent to the Department of Medlilne, University of Callfornla, San Fran- cisco, School of Medicine, San Francisco, CA 94143. DR. SMITH: * In keeping with our yearly tradition, this Medical Grand Rounds will be presented by our chief resident. Dr. Richard Locksley will dis- cuss fuel utilization in marathons, and its impli- cations for performance. This is not just of aca- demic interest to Dr. Locksley because he has run the marathon in less than 2 hours and 30 minutes himself. DR. LOCKSLEY:t Marathons have become increas- ingly popular races in the United States, with more than 50,000 men and women runners completing such races in 1979. The high intensity and pro- longed duration of these grueling 42 km (26.2 mile) road races clearly illustrate the role of body fuels in sustained exercise. Furthermore, a study of high-intensity prolonged aerobic exercise can shed light on many facets of intermediary metabo- lism that are broadly applicable to other hyper- catabolic states, such as occur in trauma, starva- tion or during surgical procedures. Recent data increasingly suggest several metabolic benefits of long-distance running." 2 The following review *Lloyd H. Smith, Jr, MD, Professor and Chairman, Department of Medicine. tRichard Locksley, MD, Chief Medical Resident. was undertaken to provide background for these emerging concepts, as well as to assist runners preparing for such an event. The energy expenditure of a marathon is roughly 60 kcal per km (96 kcal per mile). Pace has little effect on the caloric cost per mile."," The intensity of a competitive marathon is between 70 percent and 80 percent of the maximal oxygen uptake (Vo2 max). Vo2 max defines the point at which oxygen consumption and combustion can no longer keep up with the breakdown of adeno- sine triphosphate (ATP) by contracting muscles; that is, when oxidative (or aerobic) exercise can no longer continue. Thus, a marathon is an aero- bic event. This contrasts to the sprint events, in which ATP is supplied principally by anaerobic glycolysis and consumption of the stored phos- phagens ATP and creatine phosphate. The fuel stores in an average man in the rest- ing postabsorptive state are given in Table 1. At the pace of a competitive marathon, if only car- bohydrate stores (glucose and glycogen) were used, the blood glucose would be consumed in less than half a mile, the hepatic glycogen by two miles, and the entire muscle glycogen supply in an THE WESTERN JOURNAL OF MEDICINE 493
Transcript
Refer to: Locksley R: Fuel utilization in marathons: Implicationsfor performance-Medical Staff Conference, Universityof California, San Francisco. West J Med 133:493-502,Dec 1980
Medical Staf Conference
Fuel Utilization in Marathons:Implications for Performance
These discusslons are selected from the weekly staff conferences In the
Department of Medllne, University of Callfornla, San Francisco. Taken
from transcriptions, they are prepared by Drs. David W. Martin, Jr, Pro-
fessor of Medicine, and James L. Naughton, Assistant Professor of Medl-cine, under the directlon of Dr. Lloyd H. Smith, Jr, Professor of Medllneand Chalrman of the Department of Mediclne. Requests for reprlnts should
be sent to the Department of Medlilne, University of Callfornla, San Fran-cisco, School of Medicine, San Francisco, CA 94143.
DR. SMITH: * In keeping with our yearly tradition,this Medical Grand Rounds will be presented byour chief resident. Dr. Richard Locksley will dis-cuss fuel utilization in marathons, and its impli-cations for performance. This is not just of aca-demic interest to Dr. Locksley because he hasrun the marathon in less than 2 hours and 30minutes himself.
DR. LOCKSLEY:t Marathons have become increas-ingly popular races in the United States, with morethan 50,000 men and women runners completingsuch races in 1979. The high intensity and pro-longed duration of these grueling 42 km (26.2mile) road races clearly illustrate the role of bodyfuels in sustained exercise. Furthermore, a studyof high-intensity prolonged aerobic exercise canshed light on many facets of intermediary metabo-lism that are broadly applicable to other hyper-catabolic states, such as occur in trauma, starva-tion or during surgical procedures. Recent dataincreasingly suggest several metabolic benefits oflong-distance running." 2 The following review
*Lloyd H. Smith, Jr, MD, Professor and Chairman, Departmentof Medicine.
tRichard Locksley, MD, Chief Medical Resident.
was undertaken to provide background for theseemerging concepts, as well as to assist runnerspreparing for such an event.The energy expenditure of a marathon is
roughly 60 kcal per km (96 kcal per mile). Pacehas little effect on the caloric cost per mile."," Theintensity of a competitive marathon is between70 percent and 80 percent of the maximal oxygenuptake (Vo2 max). Vo2 max defines the point atwhich oxygen consumption and combustion canno longer keep up with the breakdown of adeno-sine triphosphate (ATP) by contracting muscles;that is, when oxidative (or aerobic) exercise canno longer continue. Thus, a marathon is an aero-bic event. This contrasts to the sprint events, inwhich ATP is supplied principally by anaerobicglycolysis and consumption of the stored phos-phagens ATP and creatine phosphate.
The fuel stores in an average man in the rest-ing postabsorptive state are given in Table 1. Atthe pace of a competitive marathon, if only car-bohydrate stores (glucose and glycogen) were
used, the blood glucose would be consumed inless than half a mile, the hepatic glycogen by two
miles, and the entire muscle glycogen supply in an
THE WESTERN JOURNAL OF MEDICINE 493
FUEL UTILIZATION IN MARATHONS
TABLE 1.-Fuel Reserves in an Average Man*
Fuel Reserves in anA erage Man
Fuel Tissue Kcal Grams
Triglyceride . . Adipose tissue 100,000
Glycogen .... Liver 200
Muscle 400Glucose ... Body fluids 40
Protein . Muscle 25,000
*Reprinted by permission from Newsholme,34 p 88.
15,0007012020
6,000
additional four miles. Obviously the large adiposereserve must be mobilized for fuel. Protein islargely, although not entirely,5 inaccessible as fuel.
Muscle fuels are either endogenous (glycogenand triglyceride) or exogenous (free fatty acids[FFA] and glucose)., Endogenous fuels are han-dled more efficiently because they do not re-quire mobilization or uptake, and are instantlyavailable for energy when the contractile appara-tus activates.
The reservoir for blood glucose is hepatic gly-cogen, which is converted through a series ofenzymatic steps to glucose-6-phosphate. This inturn is converted to glucose by the enzyme glu-cose-6-phosphatase, and delivered into the bloodfor use by body tissues. Muscle lacks glucose-6-phosphatase. Hence, muscle glycogen can onlyby utilized by the individual muscle fiber in which
TABLE 2.-Characteristics of Type I and Type 11 Fibers in Human Skeletal MuscleStudied in Needle-Biopsy Specimens of Quadriceps*
Characteristic
Reaction to myosin ATPase stain .....................................
Morphology and histochemistryFiber size, j (untrained men) .....................................Relative fiber frequency, % (untrained men) ........................Capillary, no. of capillary contacts per unit cross-section (,u210-).a-Glycerophosphate dehydrogenase, microphotometry .................NADH tetrazolium reductase, microphotoinetry ......................
ContractilityRelaxation rate (% force loss per 10 milliseconds) ....................Contraction time (milliseconds to peak twitch)t ......................
Energy stores (pooled or individual fibers, freeze dried)Glycogen, mmol * kg-1... .......................................... 355Triglyceride, mmol * kg-1.. ........................................ 207
NADH = nicotinamide adenine dinucleotide
ntnt
Dark staining,high activity
69.2420.85
Higher activityLower activity
15.744,62(2 motor units)
FatigableFatigable
2.15
0.85
16.6
12.1
3.66
49.4
19.3
35974
*Source: Edwards R, Young A, Wiles M: Needle biopsy of skeletal muscle in the diagnosis of myopathy and the clinical study ofmuscle function and repair. N Engl J Med 302:261-271, Jan 31, 1980fDetermined in medial gastrocnemius, peroneus longus.
A ypr, Al a sucta I ype I r luersTvve 11Fibers Tvne I Fihpr.vr
FUEL UTILIZATION IN MARATHONS
the glycogen is stored. The glucose-6-phosphateunits enter the Embden-Meyerhof pathway forcellular combustion. Similarly, the reservoir forthe plasma free fatty acids is the body adiposetissue. Hormonal stimulation of adipose lipasecleaves stored triglyceride into glycerol and threeFFA, which are released into the blood. As is thecase for muscle glycogen, muscle triglyceride isnot exportable, as shown by the inability to showglycerol efflux from working muscle.7
Fiber TypesHuman skeletal muscle is a mosaic of two fiber
types, designated type I and type II on the basisof myofibrillar ATPase activity measured at alka-line pH.8 Type II fibers (high ATPase activity)are further subdivided into types Ila and Ilb(Table 2). The proportion of fiber types is ge-netically inherited and independent of sex, al-though variability is greater in males.9 Theaverage composition of the human quadricepsmuscle is 52 percent type I, 33 percent type Ilaand 14 percent type lIb.As shown in Table 2, type I, or slow twitch
fibers, are characterized by higher oxidative en-zyme activities, greater vascularization, highermyoglobin content and higher lipid stores.10 Thesefibers are ideally suited for activity of myofibrillarATPase and glycolytic enzymes. Type II fibersdevelop peak tension more rapidly than type 1fibers, but fatigue more rapidly, because of theirlesser oxidative capabilities. Type Ila fibers havegreater oxidative capacity than type Ilb fibers.10
Glycogen content is the same in both types ofmuscle fiber. By doing biopsies on muscle afterdifferent types of exercise, the degree of glycogendepletion in each of the fiber types gives an indi-cation of which type is used preferentially."1"12 Asexpected, type I fibers are utilized for prolonged,aerobic work. Type II fibers are used for highintensity work of short duration. In exercise atmarathon intensity (70 percent to 80 percentVo, max) type I fibers are used primarily. TypeII fibers are utilized at the initiation of exercise,before vasodilatation and substrate delivery totype I fibers are maximal, and are recruited asexercise is prolonged and type I fibers fatigue.Thus, recruitment of type II fibers is dependent onthe intensity and duration of exercise.As might be expected, an increased proportion
of type I fibers would confer a metabolic advan-tage in an event like the marathon, because of theincreased aerobic capacity of the muscle. Several
studies have documented positive correlations be-tween Vo, max and percent of type I fibers, andbetween time to fatigue and percent of type IIfibers.13'4 That elite athletes gravitate to eventsat which they have genetic favorability is sub-stantiated by biopsy data showing a high propor-tion of type I fibers (up to and over 90 percent)in successful long-distance runners versus the pre-dominance of type II fibers in sprinters.'5
Training does not change the inherited propor-tion of fiber types,'6 although there are data thatshow conversion of type Ilb fibers to their moreoxidative type Ila profile with endurance train-ing.'7 Training, however, is much more importantthan genetics in predicting success. Endurancetraining further increases oxidative capacity byinducing mitochondrial proliferation in the type Ifibers,'8 changing type II fibers to a more oxida-tive profile,'7 enhancing the activity of enzymeswhich mobilize and transport fatty acids to themuscle,19-24 and increasing sensitivity to thevarious hormones which orchestrate the metabolicresponse.2 This increased oxidative capacity isreflected by the increased Vo., max in elite dis-tance runners. However, fuel utilization charac-teristics are applicable to all runners of all abili-ties, and the following comments apply to allrunners.
Endogenous FuelsTriglyceride
Although muscle triglyceride can contribute upto between 50 percent and 70 percent of the totalfatty substrate with exercise,6 the endogenous tri-glyceride levels in muscle seem not to affect per-formance. Depletion rates are related to theamount stored, and not to intensity or durationof exercise.26 In fact, elite runners have lowerendogenous muscle lipid levels. The lesser im-portance of endogenous lipid supply to total fatmetabolism may be related to the fact that FFAuptake is not rate-limiting in muscle.27
ing the course of running at a steady intensity toexhaustion show a curvilinear decline in muscleglycogen as exercise is continued. The amount ofmuscle glycogen decreases from a normal level of9 to 20 grams per kg of muscle to 0.6 to 1.0gram per kg (wet weight).28 With muscle gly-cogen depletion, exhaustion occurs and the run-ning pace can no longer be sustained. Regardless
THE WESTERN JOURNAL OF MEDICINE 495
FUEL UTILIZATION IN MARATHONS
gIioogmuscle2.or
Glycogen utilized
1 .,
1. 01
0.51
Oxygen uptake C/. of maximal)50 75 100
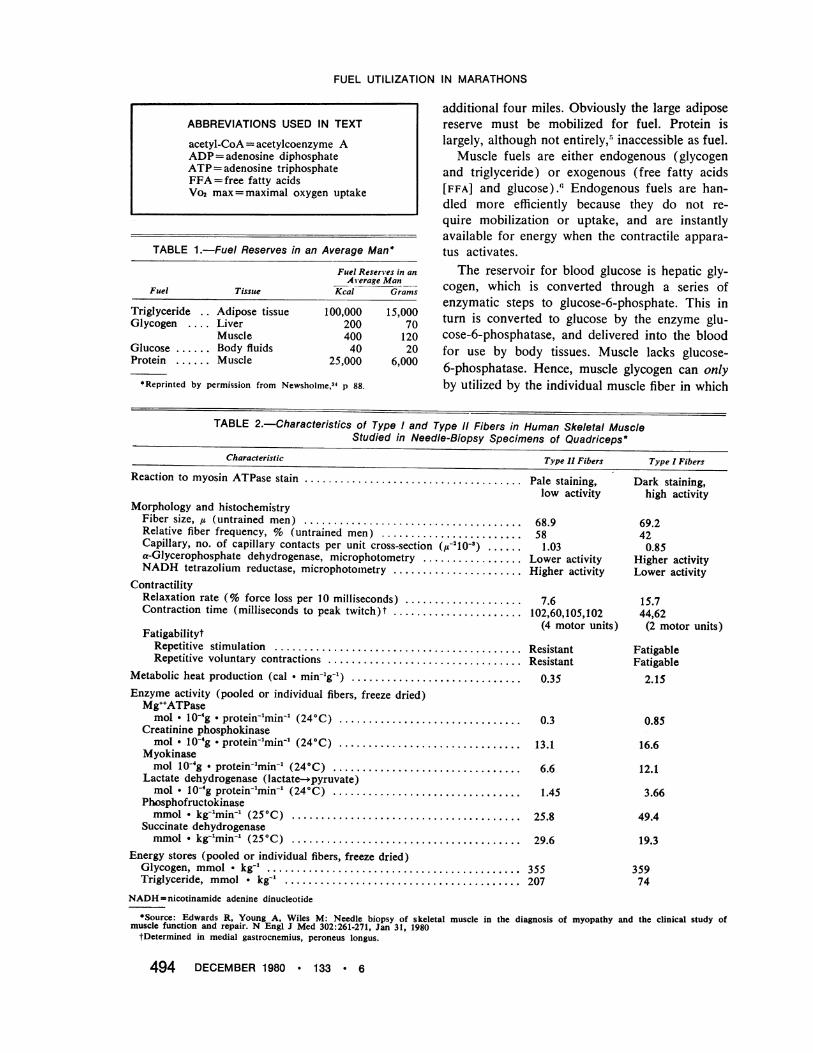
Figure 1.-Decrease of muscle glycogen after onehour of exercise at three different work levels (mean±SD, n=8). (Reprinted by permission from Hermansenet al,28 p 129.)
of the pre-exercise glycogen level, fatigue occurswith glycogen depletion.29 At intensities of 70percent Vo2 max, pace cannot be sustained whenmuscle glycogen content falls below 3 to 5 gramsper kg of muscle.30 No other variable, includingblood glucose, lactate, pyruvate, change in heartrate or oxygen consumption, or fall in body weight(with fluid depletion), correlates so consistentlywith fatigue during sustained aerobic exercise.Glycogen depletion occurs in a characteristic pat-tern during aerobic exercise of this intensity, firstfrom the aerobic type I fibers and only later fromtype II fibers as they become recruited."Two important points about the glycogen de-
pletion pattern deserve emphasis. First, at a givenexercise intensity (70 percent to 80 percent Vo2max for the marathon), the pre-exercise glycogencontent of the muscle will determine the durationof sustained optimal pace. Second, the shape ofthe curve shows a disproportionate depletion oc-curring at the initiation of exercise. Much of this,up to 20 percent of the total glycogen utilized, isconsumed in the first five minutes of exercise.6This "glycogen burst," perceived by the runneras discomfort before the "second wind" begins, isdue to the recruitment of type II glycolytic fibersfor anaerobic glycolysis at the beginning of run-ning, before blood flow is redistributed to theworking muscles bringing oxygen and substratesfor oxidative metabolism. Whether true local hy-poxemia or high catecholamine levels (principallyepinephrine) at the beginning of the race mediatethe glycogen burst is unclear. The glycogen burstis reflected by the rising levels of lactate in bloodand muscle at the onset of exercise, which gradu-ally clear as exercise continues.6'3'
The other critical point about glycogen deple-tion is that the rate of utilization is directly pro-portional to exercise intensity (percent Vo2 max),or pace (Figure 1) .28 Whereas fats alone cansupply most of the needs for resting muscle, exer-cising muscle has an obligate requirement for car-bohydrate substrate, which is met primarily byglycogen, and this obligate requirement increaseswith intensity of effort.The reason for this obligate use of glycogen is
not entirely clear, but may be related to the fol-lowing: increasing recruitment of glycolytic typeII fibers as pace quickens,1"2; the greater high-energy phosphate yield per mole of oxygen con-sumed for carbohydrates (P:O= 3.0) in compari-son to fats (P:O=2.8 for palmitic acid, the mostabundant saturated fatty acid) 32; the increasinghormonal stimulus to glycogenolysis as intensityincreases, principally mediated by increasing cate-cholamine levels,33 and the increasing cellularadenosine diphosphate (ADP) to adenosine tri-phosphate ratio, which favors glycolysis over fattyacid oxidation.34
The performance corollary is that increasingintensity (pace) will shift the glycogen depletioncurve to the lfft, and for a given exercise intensity,shorten the duration of sustained effort. The rateof muscle glycogen depletion in the marathoneffort (70 percent to 80 percent Vo2 max) is ap-proximately 0.5 gram per kg of muscle per km.30
Realizing that glycogen depletion determinesduration of exercise, a simple calculation under-scores the marathon runner's dilemma. Assuminga working muscle mass of 20 to 25 kg, and a nor-mal glycogen content of about 18 grams per kg ofmuscle, the available muscle glycogen will be ap-proximately 400 grams. At the known glycogendepletion rate for marathon pace, 10 to 12.5grams of glycogen will be used per km as obligatesubstrate. Therefore, total glycogen depletion willoccur between 32 and 40 km (19.8 and 24.8miles) of a 42 km (26.2 mile) race.What happens then is known in runner's par-
lance as "hitting the wall." Although other factorscontribute to the "wall" in the marathon, includ-ing dehydration and mental fatigue, glycogenstores are the final determinant of exercise dura-tion at this intensity.
It follows from the shape of the glycogen de-pletion curve that attempts to either shift thecurve upwards and to the right, or change therate of depletion, would enhance endurance.The influence of diet on exercise performance
496 DECEMBER 1980 * 133 * 6
FUEL UTILIZATION IN MARATHONS
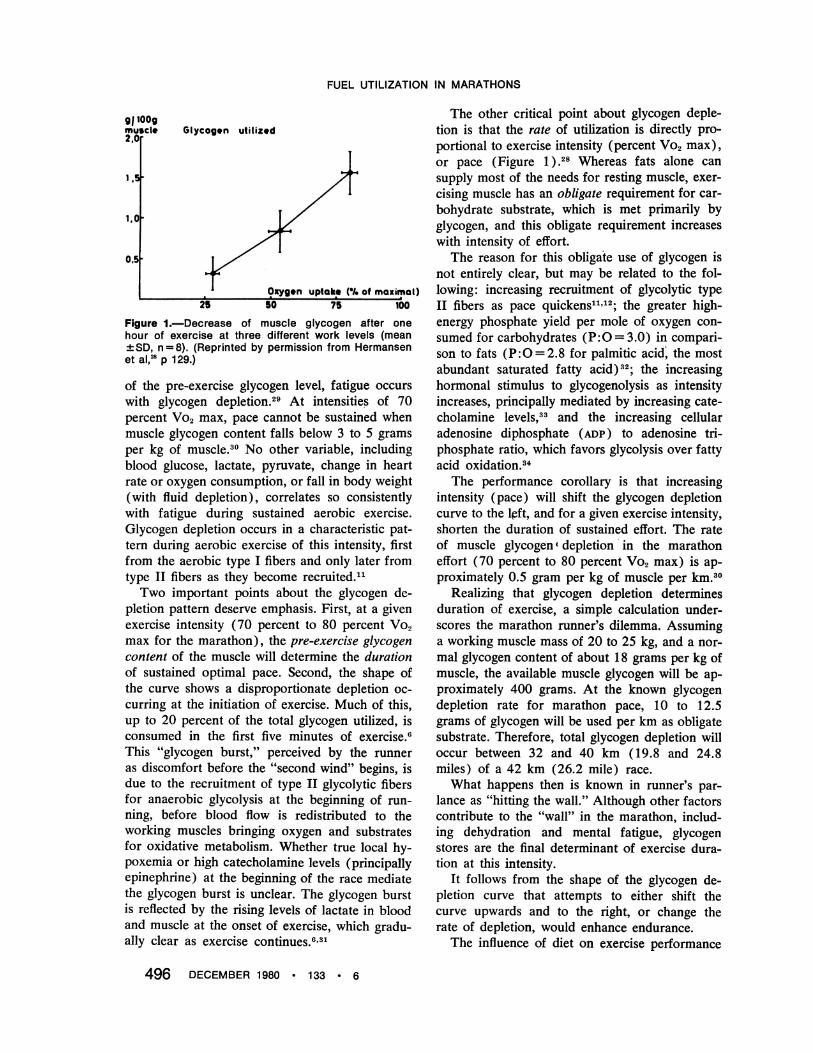
was described by Christensen and Hansen in1939.35 They showed that subjects ingesting acarbohydrate-rich diet could work substantiallylonger than those ingesting a protein-fat diet (210minutes to exhaustion versus 80). More recently,Bergstrom and Hultman explored this relationshipusing muscle biopsy specimens.28'36 These inves-tigators exercised subjects to glycogen depletionat 75 percent Vo2 max, and then fed them eithera carbohydrate-poor or a carbohydrate-rich dietfor three days. They then re-exercised their sub-jects.As seen in Figure 2, duration of work corre-
lated well with the initial glycogen level, as pre-dicted. Surprisingly, a pronounced effect of dieton glycogen storage was documented, with indi-vidual subjects storing up to 45 grams per kg ofmuscle, more than double the normal upper limit.This allowed a doubling of work duration at anintensity of 75 percent Vo2 max. Note that thesedata predict that a marathon run in less than 2hours 30 minutes would be nearly impossiblewithout elevated glycogen stores.
Four important observations have been maderegarding this supercompensation in glycogenstorage.
* Supercompensation occurs only followingexercise-induced glycogen depletion. Carbohy-drate-rich diets following a fast or low-carbohy-drate diet only replenish muscle stores to normallevels, and over several days.36
* If depletion is followed by two to three daysof low-carbohydrate diet, and then by a high-carbohydrate diet, glycogen storage is significantlyhigher.29
* Glycogen storage occurs only in those mus-cles which have been depleted by previous exer-cise, and only in those fibers which have beendepleted. Thus, in experiments studying one-legged exercise to exhaustion, only the depleted(exercised) leg supercompensated after high-carbohydrate feeding.37 For a depleting run to beeffective, it must tax the same muscles and fibertypes to be used in the race.
* Finally, fully 60 percent of the total storageoccurs in the first 10 hours following high-carbo-hydrate intake.38 Normal levels can be reachedby 24 hours. Nearly all of the ingested carbohy-drate over the first 24 hours is stored in muscle.Hepatic glycogen is also rapidly restored. Muscleloading will continue for three to four days, butincreasingly, excess caloric intake will be storedas adipose rather than in carbohydrate deposits.
WORK TIME
300 /OI ETS:Is liic.d0 Pmofesin * fofis1
: ;XCaarbhydrt ar 2,800'I"&^fo.ej *F.-
ieol-
720_
601-
I I I II1 2 3 4 5
GLYCOGE0N %1X g MX scZe
Figure 2.-Relationship between diet, initial glycogencontent in quadriceps femoris and work time in ninesubjects exercised at the same intensity. (Reprintedpermission from Bergstrom & Hultman: JAMA 221:999,197236; copyright 1978, American Medical Association.)
Frequency of meals may also enhance carbohy-drate loading, with improved storage observedwith two meals per day versus multiple smallerfeedings with the same total caloric value.39
The performance aspects of glycogen loadinghave been documented. Before competing in a 30km (18.6 mile) race, runners ate either a mixeddiet or followed the three-phase glycogen loadingprotocol, that is, a depleting run, followed by a
low-carbohydrate intake for two days and a car-
bohydrate-rich intake for three days. Three weekslater, the same race was repeated under the same
conditions, but all diets were switched. In everycase, the glycogen-loaded runner had a fastertime. Fatigue and inability to maintain the initialpace occurred near the time that muscle glycogencontent fell below 3 to 5 grams per kg of muscle.30Of note, glycogen loading did not increase theinitial pace for any of the runners, but only theduration for which optimal pace could be sus-
tained.The reasons for supercompensated storage after
exercise are unknown, but may be related to ob-served affinity changes in the insulin receptor withexercise,40 starvation4' and high-carbohydratefeeding.4' An additional benefit of glycogen load-ing is the 2.7 grams of water stored intracellularlywith each gram of glycogen, and the 0.6 gram of
THE WESTERN JOURNAL OF MEDICINE 497
in.
2c0 _
FUEL UTILIZATION IN MARATHONS
water formed by the metabolisni of 1 gram ofglycogen. Thus, each gram of glycogen stored inmuscle yields just over 3 grams of water for re-pleting evaporative losses incurred with exercise.';
Exogenous FuelsGlucose
The blood glucose represents a relatively smallcaloric source (20 grams) which must be pre-served for utilization by the brain, blood cells andanaerobic tissues of the body. With exercise, pe-ripheral uptake of glucose is greatly enhanced,despite the falling insulin levels, a feature ap-preciated by diabetes specialists years ago. Theexercise-induced glucose uptake does requiresmall amounts of insulin, probably less than 10 to12 mU per mi.4' As exercise is continued, glucoseturnover can increase 10-fold to 15-fold, and bythree hours at moderate intensity can account forabout a third of the total oxidative metabolism ofthe let in human subjects."' In trained runners,who display improved FFA mobilization and utili-zation, less glucose is used, but it may still accountfor 10 percent to 20 percent of total metabolicnceds.
Hepatic glucose output increases progressivelyup to fivefold as exercise continues, first throughglycogenolysis, and later through enhanced glu-concogenesis, as three-carbon substrates (prin-cipally alanine, lactate, glycerol and pyruvate)become available.4";
All the major hormonal changes of exerciseserve to preserve the blood glucose, and to mobi-lize stored complex fuels to simpler forms thatcan be utilized by working muscle. Of major im-portance are the rise in catecholamines and fallin insulin levels. Thus, epinephrine stimulatesglycogenolysis, inhibits insulin release and inter-feres with insulin action peripherally740 nor-epinephrine mobilizes FFA from adipose tissue;falling insulin levels minimize peripheral glucoseuptake while allowing lipolysis and glycogenolysis,and glucagon levels rise to stimulate splanchnicglucose output in concert with increased epineph-rine and falling insulin levels.' 52
If liver glycogen is depleted, either throughstarvation or prolonged exercise, and the intensityof effort is such that the obligate carbohydraterequirement for the working muscle exceeds theglycogen content, blood glucose will be utilized.Gluconeogenesis alone cannot maintain the bloodglucose at high work intensities, and hypoglycemiaoccurs, stopping exercise.5 3
At moderate work levels ingested glucose canbe utilized by working muscle, thus sparingmuscle glycogen. But at marathon race pace,ingested glucose has little sparing effect on gly-cogen.55 This may be due to the increased epi-nephrine levels at high work loads. In fact, largedoses of glucose preceding exercise will raiseportal vein insulin levels and substantially lowerblood sugar by impairing hepatic glucose output.56
Fr-ee Faux Acids
Free fatty acids are the major exogenous sub-strate for working muscle. They are taken up andoxidized almost to completion in direct relation-ship to arterial concentration.7'57 The rate-limitingstep in muscle FFA use is in mobilization fromadipose stores. The latter occurs principallythrough norepinephrine stimulation of adiposelipase,5 which cleaves triglyceride, yielding glyc-erol and three FFA molecules, which are trans-ported in blood bound to albumin.
At initiation of running, FFA levels declineowing to uptake by working muscle.5' SubsequentFFA mobilization is rapid, mediated by sympa-thetic outflow and falling insulin levels. After theinitial lag, mobilization outstrips uptake as docu-mented by rising levels of plasma FFA and glyc-erol. After 40 to 60 minutes of running, FFA levelshave risen to six times basal levels. Turnover ofFFA rises as well, reflecting increased muscle uti-lization of fatty substrate as exercise continues,shown by the falling respiratory quotient in exer-cising subjects. Except for the initial lag period,mobilization continues to supply FFA faster thandemand, and fractional extraction of FFA actuallydecreases as exercise is continued.60 When exer-cise stops, mobilizing forces are unmasked as anovershoot in the FFA level, which is responsiblefor the ketosis after exercise.
The relationship of FFA utilization to exerciseintensity (percent Vo., max) is the reverse ofthat for glycogen.'' Therefore, at rest and withprolonged light exercise, fats can supply over 90percent of the oxidative needs of the muscle. Atmoderate work intensities, free fatty acids becomethe major blood-borne fuel, supplying up to 50percent of the needs of the muscle.60 However, asintensity increases above 55 percent to 60 percentVo2 max, the amount of fat put to use declines indirect proportion to the amount of obligate carbo-hydrate substrate required for reasons previouslydiscussed. This reciprocal relationship creates acontinuum of fuel utilization from fats (principally
498 DECEMBER 1980 * 133 * 6
FUEL UTILIZATION IN MARATHONS
as exogenous FFA) for low-intensity prolongedenergy use, to carbohydrate (principally as endog-enous glvcogen) for high-intensity short-durationuse.
At marathon pace (70 percent to 80 percentVo., max), the utilization of carbohydrate relativeto fat is approximately 70:30, although it may becloser to 55:45 in trained long-distance runners.Inmproved fat metabolism in trained runners isdue to enhancement of enzymatic and hormonalactivities controlline mobilization and metabolismof FFA.-"'
The relative importance of endogenous glyco-geni and exogenous FFA in the fuel spectrum hasbccn investi>ated.';; At the intensity of the mara-thoul, blocking-FFA mobilization with nicotinicacid does not impair the ability to maintain pace.However, muscle glycogen utilization increasessionificantly to cover that portion of oxidativemetabolisnm previously supported by beta-oxida-tion of fatty acids.' This allows two conclusions:first, if free fatty acids are unavailable as sub-strate, glycogen metabolism will accelerate tocover energy needs at no cost to pace; and second,because glycogen is metabolized more quickly, thetotal dluraltioni of sustained effort will be less.
However, we have already seen that at thispace glycogen is an obligate substrate, and FFAalone cannot be used to sustain pace because ofits less favorable high-energy phosphate to oxygenratio. If the legs are first glycogen depleted by a90 percent Vo2 max effort, subsequent FFA-SUp-ported effort can only achieve 60 percent Vo2 max,and then with a 45 percent reduction in time toexhaustion. If nicotinic acid is now given, so thatboth glycogen and FFA utilization are blocked, theblood glucose is utilized until hypoglycemia re-sults, and exercise stops in only half the time ascompared with the FFA-supported effort.';3
At the time of glycogen depletion then, pacemust slow down to that effort which can be sup-ported by FFA metabolism. Protein is also usedincreasingly as substrate when glycogen stores aredepleted.' As previously noted, the decrement inpace occurs when glycogen content drops below3 to 5 grams per kg of muscle.30
The Fat-Carbohydrate ConnectionIt is evident that FFA spare glycogen as shown
by increased glycogen depletion when FFA mobi-lization is blocked with nicotinic acid. How doesFFA spare glycogen metabolism? Understandingthis requires a brief look at biochemistry.34'66 67
Glucose'1s'''\\ \ \Cell membrane
Glucose G6P=- F6P-'FDP- -* -'Pyruvate
C ytoplasm
_-Citrote.-Acetyl CoAP-Pyruvate
t 8Fatty ocy l-CoA (or ketone bod es)
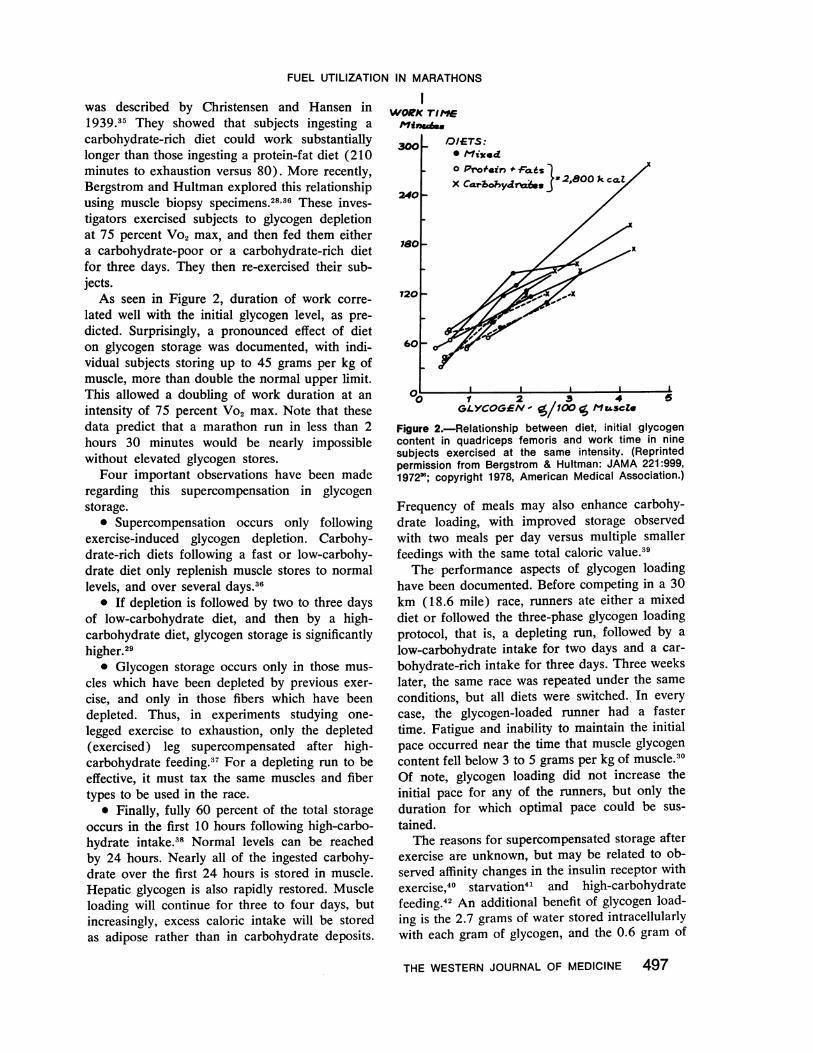
Figure 3.-Mechanism of control of glycolysis by fattyacid oxidation in muscle. The reactions are as follows:(1) membrane transport of glucose; (2) hexokinase;(3) phosphofructokinase; (4) pyruvate transport into themitochondrion; (5) pyruvate dehydrogenase; (6) citratesynthase; (7) further reactions of the citric acid cycle;and (8) beta-oxidation system (or pathway of ketonebody utilization). The dotted lines indicate allostericregulatory mechanism by which some nonequilibriumreactions of glycolysis are controlled via the rate offatty acid (and/or ketone body) oxidation in muscle.The allosteric factor regulating glucose transport is un-known. (FDP=fructose-1,6-diphosphate; F6P=fructose-6-phosphate; G6P = glucose-6-phosphate.) (Reprintedby permission from Newsholme,"4 p 81.)
Figure 3 illustrates cellular metabolism, includ-ing the glycolytic pathway by which glucose in thecytoplasm is metabolized to pyruvate, which thencrosses into the mitochondria to enter the citr-icacid cycle. Fatty acid uptake and transport inmuscle cells are not rate-limiting, ais wNe ha\ve sccn.Fatty acids are always available in excess withexercise, and they will be metabolized to intro-duce acetyl-CoA (acetylcoenzyme A) to the cycle.The increase in the acetyl-CoA to CoA ratio in-
hibits pyruvate dehydrogenase by converting it to
its inactive form. Furthermore, rising citrate levelsinhibit phosphofructokinase (PFKase). Fructose-6-phosphate levels build up in the cell causiniaglucose-6-phosphate levels to rise, and inhllibithexokinase. Thus, the glycolytic pathway is slowvedat several steps when fatty substrates are available.Furthermore, FFA have a direct depressant actionon membrane glucose transport through unknownmechanisms.As previously described, however, increasing
intensity of exercise obligates glycogen breakdownregardless of FFA levels. As pace increases, theATP to ADP ratio in the cell falls, as ATP iS dC-graded more rapidly by the contractile proteins(Figure 4). Creatine phosphate levels fall as hith-energy phosphate is transferred to ADP. A smalldecrease in the ATP to ADP ratio causes large in-creases in several metabolites, includinc adenosinemonophosphate (ANIP), inorganic phosphatc and
THE WESTERN JOURNAL OF MEDICINE 499
Mitochondrla
FUEL UTILIZATION IN MARATHONS
Glose- _Glucose I"*.!-6.-P =F6 FDP
MUSCLE CEL ,--Blood NH4- - ,,--
AMP'-~ATPP---*ADP.-- C P Pyruvate
C°2 T CAcycle ond electron ocetyl CoAPyruvoteLH20 - ronsport chain cylCAPrve
MITOCHONDRION
Figure 4.-Regulation of glycolysis in muscle at theglucose transport, hexokinase and phosphofructokinasereactions. (ADP=adenosine diphosphate; AMP=aden-osine monophosphate; ATP = adenosine triphosphate;CP = creatine phosphate; FDP = fructose-1 ,6-diphos-phate; F6P = fructose-6-phosphate; G6P = glucose-6-phosphate; TCA=tricarboxylic acid.) (Reprinted by per-mission from Newsholme,34 p 81.)
ammonium ion. These regulator molecules allfeedback on PFKase to relieve the citrate blockand accelerate glycolysis. Furthermore, pyruvatewill be preferentially utilized over fatty acetyl-CoA at entry into the electron transport chain asthe ADP concentration rises. Therefore, the ratioof ATP to ADP, which reflects the intensity of effort,or pace, precisely determines the ratio of carbohy-drate to fat metabolism. At lower intensities, fatmetabolism spares glycogen, but as intensity in-creases, more efficient high-energy phosphate gen-eration is required, and fat metabolism is replacedby glycogen.
Thus, we have seen that (1) free fatty acidsare mobilized more rapidly than they are metabo-lized by the working muscle-this accounts forthe rising FFA levels as exercise continues; (2)FFA uptake and utilization are not rate-limiting,and (3) the pace, by determining the cellular ATPto ADP ratio, will determine to what extent FFAcan be used to spare glycogen. Therefore, withexercise, FFA are always available in excess of theneeds of the working muscle, and raising levelsfurther would not be expected to enhance per-formance.
There is only one point during exercise whenFFA availability to muscle may not be optimal.This occurs at the beginning of exercise, when FFAlevels fall slightly while mobilizing mechanismslag behind muscle uptake. This occurs at the sametime as the glycogen burst, and because FFAspares glycogen breakdown, it follows that raisingthe FFA levels at the onset of exercise may spareglycogen for later use. Because the glycogen burst
can consume up to 20 percent of the total glyco-gen stored, this may make a significant contribu-tion to performance.
In rats given a fatty meal and heparin to maxi-mize FFA levels, and then run on a treadmill for30 minutes, glycogen depletion was substantiallyless than in control animals. Both skeletal muscle(type I fibers) and hepatic glycogen were spared,with total glycogen depletion decreased by 40 per-cent. Increased citrate levels were observed in bothmuscle and liver, consistent with increased FAAmetabolism.68 As predicted, time to exhaustionwas consistently prolonged in the animals withinitially elevated FFA.69
Results were similar in human subjects given afatty meal five hours before exercise and heparinintravenously just before a 70 percent Vo2 max,30-minute treadmill run.70 Muscle biopsy speci-mens showed a 40 percent decrease in glycogenconsumption. Total carbohydrate consumptiondecreased 17 percent and fat consumption rose32 percent more than in controls. Blood glucoselevels were higher also, presumably reflecting FFAinhibition of glucose uptake by muscle.
Caffeine will also raise FFA levels through itsphosphodiesterase and catecholamine-releasingactivity.71 Runners were given 330 mg of caffeineor placebo orally in a single-blind study, 60minutes before an 80 percent Vo2 max run toexhaustion. In all runners, caffeine increased timeto exhaustion by an average of 19.5 percent, from75 to 90 minutes.72 The total carbohydrate con-sumed (240 grams) was identical with and with-out caffeine as expected, but fat metabolism rosefrom 57 to 118 grams after caffeine. This permitsglycogen spared from the original glycogen burstto be utilized later, thereby increasing duration ofexercise.
Summary and ConclusionsThe following data summarize studies of fuel
metabolism as applied to the marathon.* The pre-exercise muscle glycogen content de-
termines the duration of performance.* There is obligate glycogen utilization at the
pace of a competitive marathon, approaching 0.5gram per kg of muscle per km.
* When muscle glycogen falls below 3 to 5grams per kg of muscle, pace must decrease, aseffort is sustained by fat metabolism.
* The glycogen burst at the initiation of exer-
500 DECEMBER 1980 * 133 * 6
FUEL UTILIZATION IN MARATHONS
cise contributes a significant portion (approxi-mately 20 percent) of total glycogen utilization.
* Dietary manipulation can double muscle gly-cogen levels.
* Free fatty acids spare glycogen to that re-quired for the obligate carbohydrate substrate asdetermined by the ATP to ADP ratio (pace).
* The rate-limiting step in FFA metabolism ismobilization from adipose tissue.
* Mobilization lags behind utilization only atthe onset of exercise, coincident with the glycogenburst. Otherwise, FFA are available in excess ofneed.
* Raising levels of FFA at the onset of exercisecan substantially spare glycogen that would other-wise be consumed in the glycogen burst and,hence, enhance performance.
In practical terms, the following guidelinesshould be useful to marathon athletes.
* Optimal glycogen loading involves the threephases of depletion (exercise), a carbohydrate-poor diet followed by a carbohydrate-rich diet.Depletion exercise must use the same muscles aswill be used in the race. A carbohydrate-poor dietfor two days will optimize subsequent glycogenstorage. A carbohydrate-rich (greater than 70percent carbohydrate) diet should be consumedfor three days before competition, with only lighttraining, to preserve muscle glycogen stores. Therunner should maximize caloric intake (as carbo-hydrates) for the first 10 to 24 hours of carbohy-drate loading, and then eat only isocaloric, high-carbohydrate diet for the remaining 2 to 21/2 daysbefore competition. This takes advantage of thehigh specific storage immediately after depletion,and minimizes storage in adipose tissue, with itspotentially disadvantageous weight gain.73 Twomeals per day may achieve higher glycogen levelsthan many small feedings.
* The last sugar intake should be four to sixhours before competition to assure high hepaticglycogen stores while allowing insulin levels todecrease to postabsorptive levels; 100 grams ofcarbohydrate (two candy bars) is adequate.
* Caffeine (5 mg per kg of body weight; oneNoDoz tablet contains 100 mg of caffeine) takenorally an hour before competition will raise FFAlevels and spare glycogen that would otherwisebe consumed in the glycogen burst at the begin-ning of the race.
Although these metabolic considerations canbenefit runners at any level of ability, it is im-portant to realize that metabolic manipulations
can in no way approach the beneficial effects ofconsistent training on performance.
REFERENCES1. Williams RS, Logue EF, Lewis JL, et al: Physical condition-
ing augments the fibrinolytic response to venous occlusion inhealthy adults. N Engl J Med 302:987-991, 1980
2. Wood PD, Haskell WL: The effect of exercise on plasma highdensity lipoproteins. Lipids 14:417-427, 1979
3. Costill DL, Fox EL: Energetics of marathon running. MedSci Sports 1:81-86, 1969
4. Margaria R, Cerretelli P, Aghems P: Energy cost of running.J Appl Physiol 18:367-370, 1963
5. Felig P: Amino acid metabolism in exercise. Ann NY AcadSci 301:56-63, 1977
6. Essen B, Hagenfeldt L, Kaijser L: Utilization of blood-borneand intramuscular substrates during continuous and intermittentexercise in man. J Physiol 265:489-506, 1977
7. Havel RJ, Pernow B, Jones NL: Uptake and release of freefatty acids and other metabolites in the legs of exercising men. JAppl Physiol 23:90-99, 1967
8. Saltin B, Henriksson J, Nygaard E, et al: Fiber types andmetabolic potentials of skeletal muscles in sedentary man andendurance runners. Ann NY Acad Sci 301:3-29, 1977
9. Komi PV, Viitasalo JHT, Havu M, et al: Skeletal musclefibres and muscle enzyme activities in monozygous and dizygoustwins of both sexes. Acta Physiol Scand 100:385-392, 1977
10. Essen B, Janssen E, Henriksson J, et al: Metabolic char-acteristics of fibre types in human skeletal muscle. Acta PhysiolScand 95:153-165, 1975
11. Gollnick PD, Piehl K, Saltin B: Selective glycogen depletionin human muscle fibres after exercise of varying intensity and atvarying pedalling rates. J Physiol 241:45-57, 1977
12. Gollnick PD, Karlson J, Piehl K, et al: Selective glycogendepletion in skeletal muscle fibres of man following sustainedcontractions. J Physiol 241:59-67, 1977
13. Bergh U, Thorstensson A, Sjodin B. et al: Maximal oxygenuptake and muscle fiber types in trained and untrained humans.Med Sci Sports 10:151-154, 1974
14. Thorstensson A, Karlsson J: Fatiguability and fibre com-position of human skeletal muscle. Acta Physiol Scand 98:318-322, 1976
15. Costill DL, Fink WJ, Pollock ML: Muscle fiber composi-tion and enzyme activities of elite distance runners. Med SciSports 8:96-100, 1976
16. Saltin B, Nazar K, Costill DL, et al: The nature of thetraining respQnse: Peripheral and central adaptations to one-legged exercise. Acta Physiol Scand 96:289-305, 1976
17. Andersen P, Henriksson J: Training induced changes in thesubgroups of human type II skeletal muscle fibres. Acta PhysiolScand 99:123-125, 1977
18. Kiessling K, Piehl K, Lundquist C: Effect of physical train-ing on ultrastructural features in human skeletal muscle, InPernow B, Saltin B (Eds): Muscle Metabolism During Exercise.NY-London, Plenum Press, 1971, pp 97-102
19. Mole PA, Oscai LB, Holloszy JO: Adaptation of muscle toexercise-Increase in levels of palmityl CoA synthetase, carnitinepalmityl-transferase, and palmityl CoA dehydrogenase, and in thecapacity to oxidize fatty acids. J Clin Invest 50:2323-2330, 1971
20. Bjorntorp P, Fahlen M, Grimby G, et al: Carbohydrate andlipid metabolism in middle-aged, physically well-trained men.Metabolism 21:1037-1044, 1972
21. Nikkila E, Taskinen M, Rehenen S, et al: Lipoproteinlipase activity in adipose tissue and skeletal muscle of runners:Relation to serum lipoprotein. Metabolism 27:1661-1671, 1978
22. Costill DL, Fink WJ, Getchell LH, et al: Lipid metabolismin skeletal muscle of endurance-trained males and females. JAppl Physiol 47:787-791, 1979
23. Askew EW, Huston RL, Dohm GL: Effect of physicaltraining on esterification of glycerol-3-phosphate by homogenatesof liver, skeletal muscle, heart and adipose tissue of rats. Metabo-lism 22:473-480, 1973
24. Froberg SO, Ostman I, Sjostrand NO: Effect of training onesterified fatty acids and carnitine in muscle and on lipolysis inadipose tissue in vitro. Acta Physiol Scand.86:166-174, 1972
25. Galbo H, Richter EA, Holst JJ, et al: Diminished hormonalresponses to exercise in trained rats. J Appl Physiol 43:953-958,1977
26. Essen B: Intramuscular substrate utilization during pro-longed exercise. Ann NY Acad Sci 301:30-44, 1977
27. Golinick PD: Free fatty acid turnover and the availabilityof substrates as a limiting facor in prolonged exercise. Ann NYAcad Sci 301:64-71, 1977
28. Hermansen L, Hultman E, Saltin B: Muscle glycogen duringprolonged severe exercise. Acta Physiol Scand 71:129-139, 1967
29. Bergstrom J, Hermansen L, Hultman E, et al: Diet, muscleglycogen and physical performance. Acta Physiol Scand 71:140-150, 1967
37. Bergstrom J, Hultman E: Muscle glycogen synthesis afterexercise: An enhancing factor localized to the muscle cells inman. Nature 210:309-31(0, 1966
38. Piehl K: Time course for refilling of glycogen stores inhuman muscle fibres following exercise-induced glycogen deple-tion. Acta Physiol Scand 9(0:297-3(02, 1974
39. Sherman WM, Costill DL, Fink WJ, et al: Dietary influencecn 24-hour muscle glycogen restoration following depletion (Abstr).Med Sci Sport Exercise 12:127, 198(1
40. Soman VR, Koivisto VA, Deibert D, et al: Increased insuilinsensitivity and insulin binding after physical training. N EngI JMcd 301:12(00-120)4, 1979
41. DeFronz.o RA, Soman VR, Sherwin RS, et al: Inslin bind-ing to nmonocytes and insulin action in human obesity, starvationand refeeding. J Clin Invest 62:204-213, 1978
42. Kolterman OG, Greenfield M, Reaven GM, et al: Effect ofa high carbohydrate diet on insulin binding to adipocytes and oninsulin action in vivo in man. Diabetes 28:731-736, 1979
43. Wahren J. Felig P. Ahlborg G, et al: Glucose metabolismduLring leg exercise in man. J Clin Invest 50:2715-2725, 1971
44. Vranic M, Kawamori R, Pek S, et al: The essentiality ofinsulin and the role of glucagon in regulating glucose utilizationand production during strenuous exercise in dogs. J Clin Invest57:245-255, 1976
45. Wahren J: Glucose turnover during exercise in man. AnnNY Acad Sci 301:45-55, 1977
46. Ahlborg G, Felig P, Hagenfeldt L, et al: Substrate tturn-over during prolonged exercise in man: Splanchnic and leg me-tabolism of glucose, free fatty acids and amino acids. J ClinInvest 53:108(0-1090, 1974
48. Rizza RA, Cryer PE, Haymond MW, et al: Adrenergicmechanisms for the effects of epinephrine on glucose productionand clearance in man. J Clin Invest 65:682-689, 1980
49. Soman VR, Shamoon H, Sherwin RS: Effects of physiologicinfuLsi(on of epinephrine in normal humans: Relationship betweenthe metabolic response and beta-adrenergic binding. J Clin EndocrMetab 50:294-297, 1980
50. Sacca L, Morrone G, Cicala M, et al: Influence of epi-nephrine, norepinephrine and isoproterenol on glucose homeo-stasis in normal man. J Clin Endocr Metab 50:680-684, 1981)
51. Galbo H, Richter EA, Hilsted J, et al: Hormonal regulationduring prolonged exercise. Ann NY Acad Sci 301:72-80, 1977
52. Issekutz B: Energy mobilization in exercising dogs. Diabetes28 (Suppl 1):39-44, 1979
53. Hultman E, Nilsson LH: Liver glycogen in man-Effect ofdifferent diets and musctular exercise, In Pernow B, Saltin B(Eds): MuLscle Metabolism During Exercise. NY-London. PlenumPress, 1971, pp 143-152
54. Ahlborg G. Felig P: Influence of glucose ingestion on thefuel-hormone response during prolonged exercise. J Appl Physiol41:683-688, 1976
55. HLiltman E: Studies on muscle metabolism of glycogen andactive phosphate in man with special reference to exercise anddiet. Scan J Clin Lab Invest 19 (Suppl 94):63-73, 1967
56. Foster C, Costill DL, Fink WJ: Effects of preexercise feed-ings on endurance performance. Med Sci Sports 11:1-5, 1979
57. Hagenfeldt L: Turnover of individual free fatty acids. FedProc 34:2246-2249, 197558. Steinberg D: The fate of plasma free fatty acids and their
effects on tissue metabolism. Metabolism 13:1264-1272, 196459. Friedberg SJ, Sher PB, Bogdonoff MD, et al: The dynamics
of plasma free fatty acid metabolism during exercise. J LipidResearch 4:34-38, 1963
61). Havel RJ, Naimark A, Borchgrevink CF: TuLrnover rateand oxidaticn of free fatty acids of blood plasma in man duLringexercise: Studies during continuous infuLsion of palmitate-l-C'4.J Clin Invest 42: 1054-1063, 1963
61. Hagenfeldt L: Metabolism of free fatty acids and ketonebodies during exercise in normal and diabetic man. Diabetes 28(Suppl 1):66-70, 1979
62. Holloszy JO, Rennie MJ, Hickson RC, et al: Physiologicalconsequences of the biochemical adaptations to endurance exer-cise. Ann NY Acad Sci 301:440-454, 1977
63. Pernow B, Saltin B: Availability of substrates and capacitvfor prolonged heavy exercise in man. J Appl Physiol 31:416-422,197164. Bergstrom J, HUltman E, Jorfeldt L, et al: Effect of nicotinic
acid on physical working capacity and on metabolism of muscleglycogen in man. J Appl Physiol 26:170-176, 1969
65. Lemon PWR, Mullin JP: Effect of initial muscle glycogenlevJls on protein catabolism during exercise. J Appl Physiol 48:624-635, 1980
67. Newsholme EA: Carbohlydrate metabolism in vivo: Regula-tion of the blood glucose level. Clin Endocr Metab 5:543-578. 197668. Rennie MJ. Winder WW, Holloszy JO: A sparing effect of
increased plasma free fatty acids on muscle and liver glycogencontent in the exercising rat. Biochem J 156:647-655, 1976
70. Costill DL, Coyle E, Dalsky GP, et al: Effects of elevatedplasma FFA and insulin on muscle glycogen usage during exer-cise. J Appl Physiol 43:695-699, 1977
71. Bellet S, Kershbaum A, Finek EM: Response of free fattyacids to coffee and caffeine. Metabolism 17:702-707, 196872. Costill DL, Dalsky GP, Fink WJ: Effects of caffeine in-
gestion on metabolism and exercise performance. Med Sci Sports10:155-158, 197873. Cureton KJ, Sparling PB, Evans BW, et al: Effect of experi-mental alterations in excess weight oni aerobic capacity and dis-