Functional Characterization of Arabidopsis Calreticulin1a: A Key Alleviator of Endoplasmic Reticulum Stress Anna Christensen 1 , Karin Svensson 2 , Staffan Persson 3, *, Joanna Jung 4 , Marek Michalak 4 , Susanne Widell 2 and Marianne Sommarin 1, 5 1 Department of Biochemistry, Center for Chemistry and Chemical Engineering, Lund University, PO Box 124, SE-22100 Lund, Sweden 2 Department of Cell and Organism Biology, Biology Building, Lund University, So ¨lvegatan 35, SE-22362 Lund, Sweden 3 Max-Planck-Institute of Molecular Plant Physiology, Am Muehlenberg 1, DE-14476 Potsdam, Germany 4 Department of Biochemistry, University of Alberta, Edmonton, Alberta T6G2H7, Canada 5 Department of Plant Physiology, Umea ˚ Plant Science Centre, Umea ˚ University, SE-90187, Umea ˚, Sweden The chaperone calreticulin plays important roles in a variety of processes in the endoplasmic reticulum (ER) of animal cells, such as Ca 2þ signaling and protein folding. Although the functions of calreticulin are well characterized in animals, only indirect evidence is available for plants. To increase our understanding of plant calreticulins we introduced one of the Arabidopsis isoforms, AtCRT1a, into calreticulin-deficient (crt /) mouse embryonic fibroblasts. As a result of calreticulin deficiency, the mouse crt /fibro- blasts have decreased levels of Ca 2þ in the ER and impaired protein folding abilities. Expression of the AtCRT1a in mouse crt /fibroblasts rescued these phenotypes, i.e. AtCRT1a restored the Ca 2þ -holding capacity and chaperone functions in the ER of the mouse crt /fibroblasts, demonstrating that the animal sorting machinery was also functional for a plant protein, and that basic calreticulin functions are conserved across the Kingdoms. Expression analyses using a b-glucuronidase (GUS)–AtCRT1a promoter construct revealed high expression of CRT1a in root tips, floral tissues and in association with vascular bundles. To assess the impact of AtCRT1a in planta, we generated Atcrt1a mutant plants. The Atcrt1a mutants exhibited increased sensitivity to the drug tunicamycin, an inducer of the unfolded protein response. We therefore conclude that AtCRT1a is an alleviator of the tunicamycin-induced unfolded protein response, and propose that the use of the mouse crt /fibroblasts as a calreticulin expression system may prove useful to assess functionalities of calreticulins from different species. Keywords: Endoplasmic Reticulum — Plant — Arabidopsis — Tunicamycin — Bradykinin — Calcium. Abbreviations: BSA, bovine serum albumin; CNX, calnexin; CRT, calreticulin; DTT, dithiothreitol; ER, endoplasmic reticulum; GUS, b-glucuronidase; HA, hemagglutinin; InsP3, inositol 1,4,5-trisphosphate; PBS, phosphate-buffered saline; PEPC, phosphoenolpyruvate carboxylase; PMSF, phenylmethyl- sulfonyl fluoride; PVDF, polyvinylidene difluoride; RT–PCR, reverse transcription–PCR; SHD, SHEPHERD; UPR, unfolded protein response. Introduction The endoplasmic reticulum (ER) plays a vital role in a variety of cellular processes including Ca 2þ storage and release, lipid and protein synthesis, protein folding and post-translational modifications (Trewavas and Malho 1998, Michalak et al. 1999, Baumann and Walz, 2001, Persson and Harper, 2006). The ubiquitous ER protein calreticulin (CRT) is important for several of these processes in animal cells, including Ca 2þ homeostasis (Camacho and Lechleiter 1995, Mery et al. 1996, Arnaudeau et al. 2002) and protein folding (Peterson et al. 1995, Culina et al. 2004). In addition, animal CRTs are suggested to be involved in cell adhesion (Coppolino et al. 1997), apoptosis (Groenendyk and Michalak 2005) and regulation of gene expression (Burns et al. 1994, Dedhar et al. 1994). More specifically, CRT is essential for cardiac development in mice (Mus musculus), and CRT deficiency (crt /) is embryonic lethal (Nakamura et al. 2001a). Cells derived from CRT-deficient embryos have impaired Ca 2þ homeostasis and compromised protein folding (Nakamura et al. 2001b). The latter became apparent through reduced binding of the agonist bradykinin to the cell surface receptor in CRT-deficient cells. It appeared that the bradykinin-binding receptor, coupled to inositol 1,4,5-trisphosphate (InsP 3 )-dependent Ca 2þ release from the ER, is impaired in the crt /cells, suggesting that CRT affects the folding of the bradykinin receptor (Nakamura et al. 2001b). Expression of mammalian recom- binant CRT rescues this phenotype, indicating that the crt /mouse cells provide an excellent tool for exploring functional properties of CRTs. Although CRT is well characterized in mammals, the functional roles of plant CRTs are much less well under- stood, and have only indirectly been associated with protein folding (Denecke et al. 1995) and Ca 2þ homeostasis (Persson et al. 2001, Wyatt et al. 2002, A ˚ kesson et al. 2005). CRT is expressed in most plant tissues (Crofts and Denecke 1998), with high abundance in flowers *Corresponding author: E-mail, [email protected]; Fax, þ49-331-567-898149. Plant Cell Physiol. 49(6): 912–924 (2008) doi:10.1093/pcp/pcn065, available online at www.pcp.oxfordjournals.org ß The Author 2008. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: [email protected]912 Downloaded from https://academic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
Transcript
Functional Characterization of Arabidopsis Calreticulin1a: A Key Alleviator
of Endoplasmic Reticulum Stress
Anna Christensen1, Karin Svensson
2, Staffan Persson
3,*, Joanna Jung
4, Marek Michalak
4,
Susanne Widell2and Marianne Sommarin
1, 5
1 Department of Biochemistry, Center for Chemistry and Chemical Engineering, Lund University, PO Box 124, SE-22100 Lund, Sweden2 Department of Cell and Organism Biology, Biology Building, Lund University, Solvegatan 35, SE-22362 Lund, Sweden3 Max-Planck-Institute of Molecular Plant Physiology, Am Muehlenberg 1, DE-14476 Potsdam, Germany4 Department of Biochemistry, University of Alberta, Edmonton, Alberta T6G2H7, Canada5 Department of Plant Physiology, Umea Plant Science Centre, Umea University, SE-90187, Umea, Sweden
The chaperone calreticulin plays important roles in
a variety of processes in the endoplasmic reticulum (ER) of
animal cells, such as Ca2þ signaling and protein folding.
Although the functions of calreticulin are well characterized
in animals, only indirect evidence is available for plants.
To increase our understanding of plant calreticulins we
introduced one of the Arabidopsis isoforms, AtCRT1a, into
Plant Cell Physiol. 49(6): 912–924 (2008)doi:10.1093/pcp/pcn065, available online at www.pcp.oxfordjournals.org� The Author 2008. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
912
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
(Nelson et al. 1997, Borisjuk et al. 1998). In the plant cell,
CRT is mainly localized to the ER (Opas et al. 1996,
Borisjuk et al. 1998), but has also been shown to be enriched
in the Golgi apparatus in Liriodendron tulipifera, possibly
during maturation of the CRT protein (Navazio et al.
2002), and in the nuclear envelope in maize (Zea mays)
and tobacco (Nicotiana tabacum; Denecke et al. 1995,
Napier et al. 1995). Furthermore, CRT may be unevenly
distributed within the ER, with possible enrichment
associated with plasmodesmata in several tissues (Baluska
et al. 1999, Chen et al. 2005). The apparent complexity in
expression and localization of plant CRTs may be explained
by developmental and tissue-dependent variations, but
may also be due to different homologs of the protein
(Persson et al. 2002, Persson et al. 2003). Two or more
CRT genes are present both in plants and in mammals
(Nelson et al. 1997, Persson et al. 2002, Persson et al. 2003).
Phylogenetic studies and expression analyses of Arabidopsis
(Arabidopsis thaliana) as well as rice (Oryza sativa) have
shown that CRTs in higher plants form two distinct groups;
CRT1a/CRT1b and CRT3 (Persson et al. 2003), suggest-
ing that the observed complexity in CRT expression and
distribution in plants reflect different isoforms of the
protein.
In this study, we focused on functional characterization
of the Arabidopsis CRT isoform AtCRT1a. We used crt�/�
mouse embryonic fibroblasts as the model system to
investigate AtCRT1a functions as a chaperone and Ca2þ
storage protein. Expression of AtCRT1a in crt�/� mouse
cells rescued the mutant phenotypes, which provide the first
direct evidence that the essential functional properties
of CRTs are conserved across the two Kingdoms.
Furthermore, Arabidopsis mutant analyses established
that AtCRT1a has a key role in alleviating the tunicamy-
cin-induced unfolded protein response (UPR) in higher
plants.
Results
Expression of AtCRT1a
From a comparison of the two fully sequenced
genomes of Arabidopsis and rice, it is clear that the
CRT1a/1b and CRT3 groups evolved before the split
between monocots and eudicots (Fig. 1A). To evaluate the
functional properties of specific CRT isoforms, we here
focus on one of the CRTs in the CRT1a/b cluster, CRT1a,
in Arabidopsis. We chose to focus on AtCRT1a because of
its relatively high abundance and because of its rapid
induction in response to tunicamycin, a blocker of N-linked
glycosylation and inducer of the UPR (Persson et al. 2003).
First the relative expression levels were assessed using
(Fig. 1B). AtCRT1a transcripts were present in flowers,
siliques, rosette leaves, roots, pedicels and cauline leaves at
similar expression levels (Fig. 1B). This prompted a more
detailed investigation of the expression within the respective
plant organs. We generated transgenic plants where a
b-glucuronidase (GUS) reporter gene was placed under the
control of the 1.5 kb promoter region upstream of the
AtCRT1a open reading frame. In roots, GUS activity was
strongest in the root tips, suggesting a role for CRT in
growing tissues (Fig. 1C). GUS activity was also detected in
floral tissues, specifically associated with the ovules, and
in and around vascular tissues in rosette leaves (Fig. 1C).
A
C
B
Fig. 1 Phylogenetic analysis of CRTprotein sequences in plants and expressionanalysis of AtCRT1a. (A) Phylogeneticanalysis of CRT amino acid sequencesfrom Arabidopsis and rice. Human CRT1was used as the outgroup. (B) Expressionanalysis of AtCRT1a in different tissues inArabidopsis assessed by semi-quantitativeRT–PCR. Upper panel: AtCRT1a (tissuesfrom 4-week-old plants). Amplification ofACTIN RNA was used as control (lowerpanel). (C) Expression of AtCRT1a assessedby a promoter: GUS reporter gene fusionin transgenic Arabidopsis. Arrows indicatemost prominent expression in the root tip of14-day-old seedlings (left panel), ovules infloral tissue (middle panel) and vasculartissue in rosette leaves of 14-day-oldseedlings (right panel).
Functions of calreticulins in higher plants 913
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
The expression patterns correlated well with microarray-
based databases, such as Genevestigator (https://www.
genevestigator.ethz.ch; Zimmermann et al. 2004).
Ca2þ binding properties of AtCRT1a
Plant CRTs seem to have a large Ca2þ holding capacity
(Borisjuk et al. 1998, Persson et al. 2001). These results have
mainly been obtained from plant extracts where multiple
CRT isoforms could not be distinguished, and only limited
information is therefore available for individual CRT
isoforms in plants. Analogous to CRT1 from human and
mouse, AtCRT1a has high levels of the negatively charged
aspartic and glutamic acids in the C-terminal region
(Fig. 2A). The negatively charged amino acids in this
region are believed to be responsible for the Ca2þ-holding
capacity in mammalian CRT proteins (Baksh and Michalak
1991). To test whether AtCRT1a may bind Ca2þ in vitro,
we generated a 6�histidine (6�His)-tagged AtCRT1a
construct, expressed it in Escherichia coli and purified the
recombinant protein (Fig. 2B, left panel). Ca2þ binding of
the recombinant AtCRT1a was investigated using 45Ca2þ
overlay of protein blots with a 6�His-tagged human (Homo
sapiens) CRT1, HsCRT1, as positive control. Distinct
radiolabeled bands were detected at apparent molecular
masses in SDS-PAGE of 60 kDa for AtCRT1a and at
56 kDa for HsCRT1 (Fig. 2B, right panel). These bands
corresponded to two protein bands detected with an anti-
6�His antibody by Western blot analysis (data not shown)
and by Coomassie Brilliant Blue staining (compare panels,
Fig. 2B). No labeling was detected for a 6�His-tagged
phosphoenolpyruvate carboxylase (PEPC), used as negative
control, confirming that under these conditions the His tag
itself did not bind Ca2þ (Fig. 2B, right panel). These data
show that both HsCrt1 and AtCrt1a are able to bind Ca2þ
in vitro.
Expression of AtCRT1a in crt�/� mouse embryonic
fibroblasts
Mutant analysis of rabbit (Oryctolagus cuniculus)
CRT1 revealed that several amino acids are essential for
protein folding and Ca2þ storage capacity in animal CRTs
(Martin et al. 2006). These amino acids are conserved in
HsCRT1, mouse (M. musculus) CRT1 and AtCRT1a
(Fig. 2A). In addition, both AtCRT1a and HsCRT1 bind
Ca2þ in vitro (Fig. 2B, right panel), suggesting that the basic
functional CRT properties may be conserved between
plants and animals. To test whether AtCRT1a has similar
function to its animal counterpart, we generated a construct
where an AtCRT1a cDNA was fused to a hemagglutinin
(HA) tag (Fig. 3A) and transformed it into crt�/� mouse
embryonic fibroblasts. Six stable transformants were
isolated using zeocin as a selectable marker, and were
assessed using immunoblotting with anti-HA antibodies.
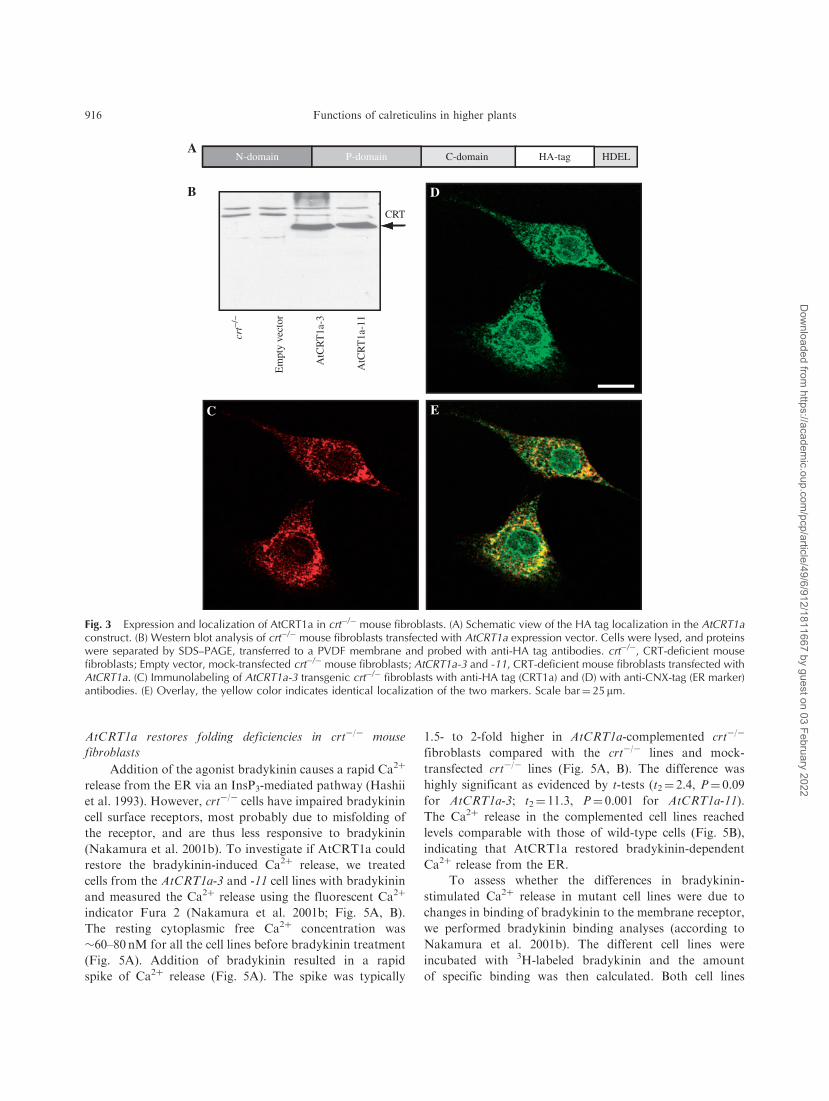
A typical result is shown in Fig. 3B for two of these
AtCRT1a transgenic crt�/� cell lines (AtCRT1a-3 and -11).
A single 60-kDa protein band was observed in both cell
lines, while no signal was detected in the crt�/� cell line, or
in mock-transfected cell lines, confirming that the observed
protein bands corresponded to the recombinant AtCRT1a.
Immunofluorescence studies of crt�/� cells expressing
AtCRT1a using the anti-HA antibodies revealed a reticu-
lar-like network reminiscent of the ER (Fig. 3C).
To confirm that this network indeed was ER, we also
immunolabeled the cells with antibodies against calnexin
(CNX), an ER-localized chaperone (Fig. 3D). The CNX
pattern of staining in a reticular-like network overlapped
with that of the HA tag antibodies confirming that the
recombinant AtCRT1a was targeted to the ER in the crt�/�
mouse fibroblasts (Fig. 3E).
AtCRT1a restores Ca2þ deficiencies in crt�/� mouse
fibroblasts
The mouse crt�/� fibroblasts have reduced ER Ca2þ
storage capacity (Fig. 4A; Nakamura et al. 2001b).
To assess whether the introduction of the recombinant
AtCRT1a could restore this deficiency, the AtCRT1a-3 and
AtCRT1a-11 cell lines were cultured for 50 h with 45Ca2þ
and then resuspended in a Ca2þ-free buffer. The cells were
pre-incubated at 378C followed by addition of thapsigargin,
an inhibitor of the SERCA pump (Thastrup et al. 1990),
to measure Ca2þ in rapidly exchangeable stores (Nakamura
et al. 2001b), or by addition of the Ca2þ ionophore
ionomycin, to measure the total amount of Ca2þ in the
cell. Released Ca2þ was determined in the supernatant after
pelleting the cells (background values collected in the
absence of thapsigargin or ionomycin accounted for
approximately 40% and are subtracted from data presented
in Fig. 4). Addition of thapsigargin caused a 2-fold increase
in Ca2þ release in AtCRT1a-3 and -11 compared with the
crt�/� and mock-transfected cells (Fig. 4A). The increase
of Ca2þ in thapsigargin-sensitive stores for AtCRT1a-3
and -11 was highly significant as measured by t-tests
(t2¼ 13.7, P¼ 0.000164 for AtCRT1a-3; t2¼ 1.1,
P¼ 0.322455 for AtCRT1a-11) comparing stably trans-
fected clones with the crt�/� fibroblasts. The thapsigargin-
triggered Ca2þ release from the AtCRT1a transgenic lines
was, furthermore, of similar size to those of the wild-type
control line. Addition of ionomycin caused a larger Ca2þ
release compared with thapsigargin (Fig. 4B). The extra
Ca2þ released by ionomycin is most probably emanating
from intracellular compartments other than the ER, such as
the mitochondria. Thus, our data show that AtCRT1a
restored thapsigargin-sensitive Ca2þ stores in the crt�/�
mouse fibroblasts.
914 Functions of calreticulins in higher plants
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
M L L S V P L L L G L L G L A V A E P A V Y F K E Q F L D G D G W T S R W I E S K H K S D - -M L L S V P L L L G L L G L A A A D P A I Y F K E Q F L D G D A W T N R W V E S K H K S D - -
M A K L N P K F I S L I L F A L V V I V S A E V I F E E K F E D G - - W E K R W V K S D W K K D D N
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
- F G K F V L S S G K F Y G D E E K D K G L Q T S Q D A R F Y A L S A S F E P F S N K G Q T L V V Q- F G K F V L S S G K F Y G D L E K D K G L Q T S Q D A R F Y A L S A K F E P F S N K G Q T L V V QT A G E W K H T A G N W S G D A N - D K G I Q T S E D Y R F Y A I S A E F P E F S N K D K T L V F Q
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
F T V K H E Q N I D C G G G Y V K L F P N S L D Q T D M H G D S E Y N I M F G P D I C G P G T K K VF T V K H E Q N I D C G G G Y V K L F P S G L D Q K D M H G D S E Y N I M F G P D I C G P G T K K VF S V K H E Q K L D C G G G Y M K L L S D D V D Q T K F G G D T P Y S I M F G P D I C G Y S T K K V
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
H V I F N Y K G K N V L I N K D I R C K D D E F T H L Y T L I V R P D N T Y E V K I D N S Q V E S GH V I F N Y K G K N V L I N K D I R C K D D E F T H L Y T L I V R P D N T Y E V K I D N S Q V E S GH A I L T Y N G T N H L I K K E V P C E T D Q L T H V Y T F V L R P D A T Y S I L I D N V E K Q T G
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
S L E D D W D F L P P K K I K D P D A S K P E D W D E R A K I D D P T D S K P E D W D - K P E H I PS L E D D W D F L P P K K I K D P D A A K P E D W D E R A K I D D P T D S K P E D W D - K P E H I PS L Y S D W D L L P A K K I K D P S A K K P E D W D D K E Y I P D P E D T K P A G Y D D I P K E I P
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
D P D A K K P E D W D E E M D G E W E P P V I Q N P E Y K G E W K P R Q I D N P D Y K G T W I H P ED P D A K K P E D W D E E M D G E W E P P V I Q N P E Y K G E W K P R Q I D N P D Y K G T W I H P ED T D A K K P E D W D D E E D G E W T A P T I P N P E Y N G E W K P K K I K N P A Y K G K W K A P M
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
I D N P E Y S P D P S I Y A Y D N F G V L G L D L W Q V K S G T I F D N F L I T N D E A Y A E E F GI D N P E Y S P D A N I Y A Y D S F A V L G L D L W Q V K S G T I F D N F L I T N D E A Y A E E F GI D N P E F K D D P E L Y V F P K L K Y V G V E L W Q V K S G S L F D N V L V S D D P E Y A K K L A
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
N E T W G V T K A A E K Q M K D K Q D E E Q R L K E E E E D K - - - - K R K E E E E A E D K E - - -N E T W G V T K A A E K Q M K D K Q D E E Q R L K E E E E D K - - - - K R K E E E E A E D K E - - -E E T W G K H K D A E K A A F D E A E K K - - - R E E E E S K D A P A E S D A E E E A E D D D N E G
H. sapiens Crt1M. musculus Crt1A. thalina Crt1a
D D E D K D E D E E D E E D K E E D E E E D V P G Q A K D E LD D D D R D E D E D E E D E K E E D E E E - S P G Q A K D E LD D S D N E S K S E E T K E A E E T K E A E E T D A A H D E L
-
C domain
-
SS
N domain P domain
ER-R
-
*
kD
60
56
MW
PEPC
AtC
RT
1a
HsC
RT
1
PEPC
AtC
RT
1a
HsC
RT
1
B
Fig. 2 Comparison of CRT protein sequences and Ca2þ binding of recombinant AtCRT1a. (A) Comparison of amino acid sequences ofhuman CRT1, mouse CRT1 and Arabidopsis CRT1a (GenBank accession Nos. AAA51916, NP_031617 and AAC49695, respectively).Identical and similar amino acids are highlighted in gray. The approximate positions of the histidine-rich N-terminus (N domain), theproline-rich internal region (P domain) and the negatively charged C-terminus (C domain) are indicated with arrows. The putative ER signalsequence (SS) and the ER-retrieval signal (ER-R) are also indicated. The vertical black arrows indicate three highly conserved cysteineresidues. The asterisk indicates a conserved histidine residue in the N domain. Four amino acids (Glu239, Asp241, Glu243 and Trp244,numbering referring to the HsCRT1sequence) that are highlighted in black boxes have been implicated in interactions between ERp57 andanimal CRT1s. (B) Purified recombinant proteins (5 mg per lane) were separated by SDS–PAGE and either stained with Coomassie BrilliantBlue R250 (left panel) or transferred to a PVDF membrane and probed with 45Ca2þ (right panel). His-tagged PEPC was used as a negativecontrol for 45Ca2þ binding. MW, lane with molecular weight standards.
Functions of calreticulins in higher plants 915
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
AtCRT1a restores folding deficiencies in crt�/� mouse
fibroblasts
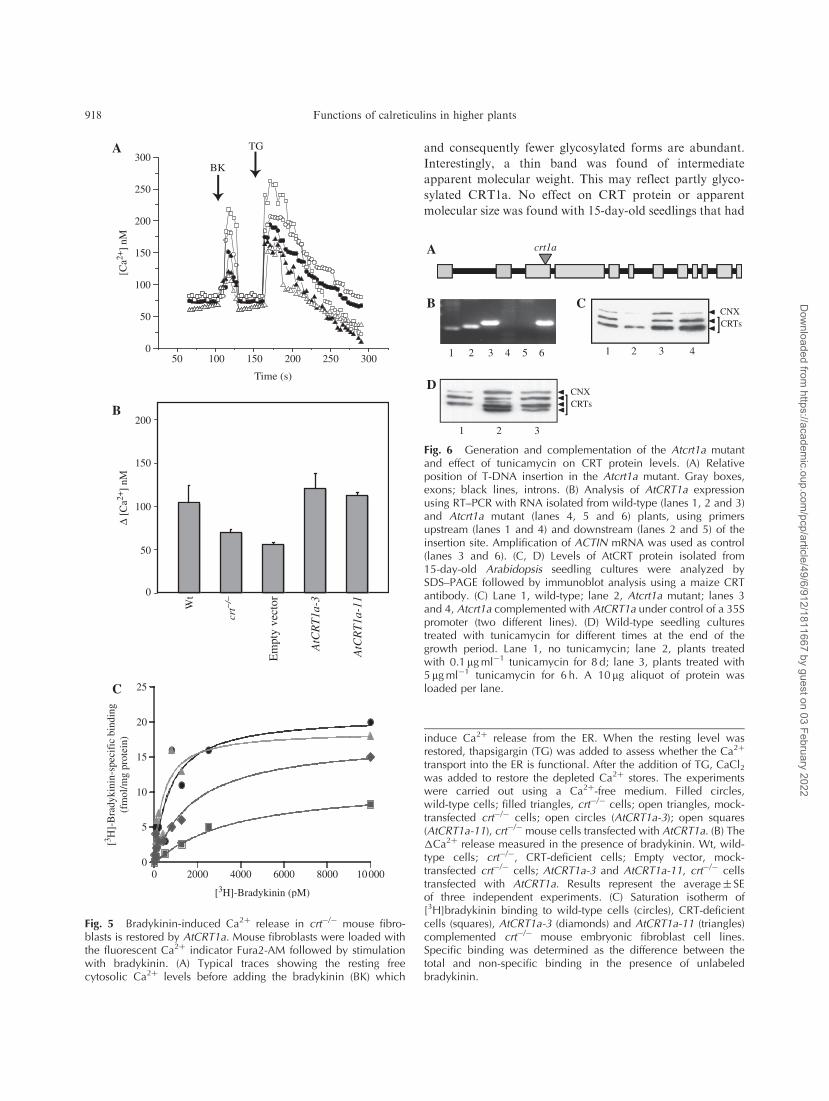
Addition of the agonist bradykinin causes a rapid Ca2þ
release from the ER via an InsP3-mediated pathway (Hashii
et al. 1993). However, crt�/� cells have impaired bradykinin
cell surface receptors, most probably due to misfolding of
the receptor, and are thus less responsive to bradykinin
(Nakamura et al. 2001b). To investigate if AtCRT1a could
restore the bradykinin-induced Ca2þ release, we treated
cells from the AtCRT1a-3 and -11 cell lines with bradykinin
and measured the Ca2þ release using the fluorescent Ca2þ
indicator Fura 2 (Nakamura et al. 2001b; Fig. 5A, B).
The resting cytoplasmic free Ca2þ concentration was
�60–80 nM for all the cell lines before bradykinin treatment
(Fig. 5A). Addition of bradykinin resulted in a rapid
spike of Ca2þ release (Fig. 5A). The spike was typically
1.5- to 2-fold higher in AtCRT1a-complemented crt�/�
fibroblasts compared with the crt�/� lines and mock-
transfected crt�/� lines (Fig. 5A, B). The difference was
highly significant as evidenced by t-tests (t2¼ 2.4, P¼ 0.09
for AtCRT1a-3; t2¼ 11.3, P¼ 0.001 for AtCRT1a-11).
The Ca2þ release in the complemented cell lines reached
levels comparable with those of wild-type cells (Fig. 5B),
indicating that AtCRT1a restored bradykinin-dependent
Ca2þ release from the ER.
To assess whether the differences in bradykinin-
stimulated Ca2þ release in mutant cell lines were due to
changes in binding of bradykinin to the membrane receptor,
we performed bradykinin binding analyses (according to
Nakamura et al. 2001b). The different cell lines were
incubated with 3H-labeled bradykinin and the amount
of specific binding was then calculated. Both cell lines
B
crt−/
−
Em
pty
vect
or
AtC
RT
1a-3
AtC
RT
1a-1
1
D
C E
CRT
N-domain P-domain C-domain HA-tag HDELA
Fig. 3 Expression and localization of AtCRT1a in crt�/� mouse fibroblasts. (A) Schematic view of the HA tag localization in the AtCRT1aconstruct. (B) Western blot analysis of crt�/� mouse fibroblasts transfected with AtCRT1a expression vector. Cells were lysed, and proteinswere separated by SDS–PAGE, transferred to a PVDF membrane and probed with anti-HA tag antibodies. crt�/�, CRT-deficient mousefibroblasts; Empty vector, mock-transfected crt�/� mouse fibroblasts; AtCRT1a-3 and -11, CRT-deficient mouse fibroblasts transfected withAtCRT1a. (C) Immunolabeling of AtCRT1a-3 transgenic crt�/� fibroblasts with anti-HA tag (CRT1a) and (D) with anti-CNX-tag (ER marker)antibodies. (E) Overlay, the yellow color indicates identical localization of the two markers. Scale bar¼ 25 mm.
916 Functions of calreticulins in higher plants
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
expressing AtCRT1a (AtCRT1a-3 and AtCRTa-11) showed
inreased binding of 3H-labeled bradykinin compared with
crt�/�, and for AtCRTa-11 reaching levels similar to the
wild-type cell line (Fig. 5C), indicating that binding abilities
of bradykinin were restored in crt�/� cells expressing
AtCRT1a. Earlier studies demonstrated that the amount
of bradykinin receptors was reduced by 50% in crt�/� lines
(Nakamura et al. 2001b). This fits well with the restoration
of Ca2þ release in complemented cell lines found here,
supporting that AtCRT1a was important for correct folding
of the bradykinin receptor.
AtCRT1a alleviates the tunicamycin-induced unfolded protein
response in Arabidopsis
Expression of AtCRT1a in mouse crt�/� fibroblasts
shows that AtCRT1a is active both in controlling Ca2þ
homeostasis and presumably in protein folding. To assess
its role in planta, we isolated a homozygous Atcrt1a T-DNA
mutant from the SALK collection (SALK_055452;
Fig. 6A). This line showed no detectable transcript levels
(Fig. 6B; lanes 4 and 5), confirming that the T-DNA
insert disrupted expression of the gene. In agreement with
this, one protein band is missing in the microsomal
fraction from the mutant AtCrt1a cell line using maize
CRT antibodies (Fig. 6C; compare lanes 1 and 2). This
antibody recognizes all CRTs as well as a protein band
of an apparent molecular mass of 66 kDa presumably
corresponding to CNX (Persson et al. 2003). It has been
suggested that AtCRT1a harbors more potential N-linked
glycosylation sites compared with AtCRT1b and AtCRT3,
experimentally shown by treatment with N-glycosidase F
(Persson et al. 2003). Thus, AtCRT1a should band at a
higher molecular size than expected based on the number
of amino acids. The disappearance of the CRT band with
higher molecular size in the Atcrt1a background supports
this hypothesis. We conclude that the missing band in
Fig. 6C, lane 2, corresponds to AtCRT1a, and, as predicted
from the number of available N-linked glycosylation
sites, AtCRT1a is the more heavily glycosylated CRT.
No visible effects on growth and morphology were
evident in Atcrt1a plants when grown under normal
conditions (Fig. 7A). Previous studies have shown that
short-term treatment with high concentrations of tunica-
mycin, an inhibitor of N-linked glycosylation and inducer
of the UPR, results in increased CRT expression in
Arabidopsis (Persson et al. 2003), possibly to alleviate the
cell stress. Short-term treatment of 15-day-old plants with
high concentrations of tunicamycin (5 mgml�1 for 6 h) also
resulted in increases at the protein level (Fig. 6D, lane 3).
To test whether longer exposure of tunicamycin sustains the
up-regulation of CRTs, we treated 15-day-old seedlings
with low levels of tunicamycin (0.1 mgml�1) given for the
last 8 d of culture (Fig. 6D, lane 2). The protein levels
corresponding to CRTs were indeed higher compared with
untreated seedlings also with this exposure. The increase in
CRT levels in the seedlings treated for 8 d corresponded to
lower apparent molecular weight bands compared with
control seedlings, as well as to the to 6 h treated seedlings
(compare lane 1 with lanes 2 and 3 in Fig. 6D). With the
prolonged treatment in contrast to the shorter one, all CRT
present had been synthesized during tunicamycin exposure
0
5
10
15
20
25A
AtC
RT
1a-1
1
AtC
RT
1a-3
Em
pty
vect
or
Wt
Ca2+
rel
ease
d (p
mol
es C
a2+/1
06 cel
ls)
0
5
10
15
20
25B
Ca2+
rel
ease
d (p
mol
es C
a2+/1
06 cel
ls)
crt−/
−
AtC
RT
1a-1
1
AtC
RT
1a-3
Em
pty
vect
or
Wt
crt−/
−
Fig. 4 ER Ca2þ content in crt�/� mouse fibroblasts is restoredby expression of AtCRT1a. Cellular Ca2þ content was determinedby incubation with 55mCi of 45Ca2þ followed by addition ofthapsigargin (A) or ionomycin (B). Wt, wild-type mouse embryonicfibroblasts; crt�/�, CRT-deficient mouse fibroblasts; Empty vector,mock-transfected crt�/� mouse fibroblasts; AtCRT1a-3 and -11,CRT-deficient mouse fibroblasts transfected with AtCRT1a. Thebackground values (counts min�1 in supernatant before additionof thapsigargin or ionomycin) were subtracted. Results representthe average� SE of three independently performed experiments.
Functions of calreticulins in higher plants 917
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
and consequently fewer glycosylated forms are abundant.
Interestingly, a thin band was found of intermediate
apparent molecular weight. This may reflect partly glyco-
sylated CRT1a. No effect on CRT protein or apparent
molecular size was found with 15-day-old seedlings that had
ABK
TG
Time (s)
B
0
50
100
150
200
C
250
300
150
200
50
50 100 150 200 250 300
100
0
0 2000 4000 6000 8000 100000
5
10
15
20
25
[Ca2+
] nM
∆ [C
a2+]
nM
AtC
RT
1a-1
1
AtC
RT
1a-3
Em
pty
vect
or
Wt
crt−/
−
[3 H]-
Bra
dyki
nin-
spec
ific
bin
ding
(fm
ol/m
g pr
otei
n)
[3H]-Bradykinin (pM)
Fig. 5 Bradykinin-induced Ca2þ release in crt�/� mouse fibro-blasts is restored by AtCRT1a. Mouse fibroblasts were loaded withthe fluorescent Ca2þ indicator Fura2-AM followed by stimulationwith bradykinin. (A) Typical traces showing the resting freecytosolic Ca2þ levels before adding the bradykinin (BK) which
induce Ca2þ release from the ER. When the resting level wasrestored, thapsigargin (TG) was added to assess whether the Ca2þ
transport into the ER is functional. After the addition of TG, CaCl2was added to restore the depleted Ca2þ stores. The experimentswere carried out using a Ca2þ-free medium. Filled circles,wild-type cells; filled triangles, crt�/� cells; open triangles, mock-transfected crt�/� cells; open circles (AtCRT1a-3); open squares(AtCRT1a-11), crt�/� mouse cells transfected with AtCRT1a. (B) The�Ca2þ release measured in the presence of bradykinin. Wt, wild-type cells; crt�/�, CRT-deficient cells; Empty vector, mock-transfected crt�/� cells; AtCRT1a-3 and AtCRT1a-11, crt�/� cellstransfected with AtCRT1a. Results represent the average� SEof three independent experiments. (C) Saturation isotherm of[3H]bradykinin binding to wild-type cells (circles), CRT-deficientcells (squares), AtCRT1a-3 (diamonds) and AtCRT1a-11 (triangles)complemented crt�/� mouse embryonic fibroblast cell lines.Specific binding was determined as the difference between thetotal and non-specific binding in the presence of unlabeledbradykinin.
A
B C
crt1a
D
1 2 3 4 1 2 3 45 6
CRTsCNX
1 2 3
CNXCRTs
Fig. 6 Generation and complementation of the Atcrt1a mutantand effect of tunicamycin on CRT protein levels. (A) Relativeposition of T-DNA insertion in the Atcrt1a mutant. Gray boxes,exons; black lines, introns. (B) Analysis of AtCRT1a expressionusing RT–PCR with RNA isolated from wild-type (lanes 1, 2 and 3)and Atcrt1a mutant (lanes 4, 5 and 6) plants, using primersupstream (lanes 1 and 4) and downstream (lanes 2 and 5) of theinsertion site. Amplification of ACTIN mRNA was used as control(lanes 3 and 6). (C, D) Levels of AtCRT protein isolated from15-day-old Arabidopsis seedling cultures were analyzed bySDS–PAGE followed by immunoblot analysis using a maize CRTantibody. (C) Lane 1, wild-type; lane 2, Atcrt1a mutant; lanes 3and 4, Atcrt1a complemented with AtCRT1a under control of a 35Spromoter (two different lines). (D) Wild-type seedling culturestreated with tunicamycin for different times at the end of thegrowth period. Lane 1, no tunicamycin; lane 2, plants treatedwith 0.1 mgml�1 tunicamycin for 8 d; lane 3, plants treated with5mgml�1 tunicamycin for 6 h. A 10 mg aliquot of protein wasloaded per lane.
918 Functions of calreticulins in higher plants
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
been treated with dithiothreitol (DTT), either at a low
concentration (0.2mM) for 8 d (fresh DTT was added every
second day) or at a high concentration (10mM) for 6 h,
supporting that the response to tunicamycin was not a
general stress response, but indeed related to the tunicamy-
cin-induced UPR (data not shown). To test whether the
deletion of AtCRT1a affected the tunicamycin sensitivity,
we treated 1-week old Atcrt1a and wild-type seedlings
with low tunicamycin concentrations (0.1mgml�1) for 8 d.
In agreement with the above, the tunicamycin treatment
induced CRT expression in the treated seedlings compared
with untreated seedlings (data not shown), probably
reflecting expression of CRTs other than AtCRT1a.
After 8 d, the tunicamycin-treated (0.1mgml�1) Atcrt1a
seedlings were smaller compared with wild-type seedlings
(Fig. 7B). To assess which tissues were most affected
in Atcrt1a in response to the tunicamycin treatment, we
measured leaf size, and fresh and dry weights of treated and
control seedlings (Fig. 7C). t-tests revealed that all differ-
ences between values of Atcrt1a mutant seedlings compared
with wild-type seedlings were significant (Fig. 7). To ensure
that the tunicamycin-sensitive phenotype (Fig. 7) was due to
the disruption of the AtCRT1a gene, we complemented
the mutant with an AtCRT1a cDNA under the control
of a 35S promoter (Figs. 6C, lanes 3 and 4). The comple-
mented mutant was indistinguishable from the wild-type
seedlings after tunicamycin treatment (Fig. 7B, C), con-
firming that the drug-sensitive phenotype was indeed due
to disruption of the AtCRT1a gene. Thus, AtCRT1a is
protecting the plant against tunicamycin stress and may
therefore be viewed as an alleviator of the tunicamycin-
induced UPR in Arabidopsis.
Discussion
Plant CRTs are generally assumed to have retained
functions attributed to mammalian homologs. However,
no functional characterization of plant CRTs has been
presented. We show here that the Arabidopsis CRT isoform,
AtCRT1a, can substitute for animal CRTs with regard to
Ca2þ binding and protein folding abilities. Consistent with
this, the amino acid residues that are necessary for these
functions in mammalian CRTs are conserved in AtCRT1a
(Fig. 2A). These amino acid residues are also conserved in
AtCRT1b, suggesting that AtCRT1b may be functionally
Fig. 7 The Atcrt1a mutant is sensitive to tunicamycin treatment.Seeds from wild-type, Atcrt1a, and Atcrt1a complemented withan AtCRT1a cDNA were germinated and grown on normal MSmedium for 1 week, after which they were transferred to the samemedium� tunicamycin and grown for another 8 d. (A) Control,no tunicamycin. (B) Tunicamycin-treated plants. Tunicamycin(0.1 mgml�1) was added for 8 d at the end of the growth period.Data shown in A and B are from one experiment. All experimentshave been repeated at least three times with similar phenotypictraits, and the size of the scale bars is 10mm. Wt, wild-type;Atcrt1a, Atcrt1a mutant; Atcrt1a:AtCRT1a, Atcrt1a mutant com-plemented with AtCRT1a. (C) Leaf size, fresh weight and dry weight
of seedlings exposed to tunicamycin are shown as a percentage,with wild-type levels as 100%. Black bars, wild-type; white bars,Atcrt1a mutant; gray bars, Atcrt1a complemented with AtCRT1a.Results represent the average of three independent experiments.The differences between the tunicamycin-treated wild-type andmutant were significant (t19¼ 4.2; P¼ 0.0001 for leaf size, t9¼ 7.4,P¼ 7.74E-05 for fresh weight and t9¼ 3.4, P¼ 0.009 for dry weightmeasurements).
Functions of calreticulins in higher plants 919
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
redundant with AtCRT1a. Interestingly, the plant CRT1a
protein was found to be located in the ER in the mouse cell,
despite a slightly different retrieval signal (HDEL compared
with KDEL), suggesting that the animal cell sorting
machinery also worked for the plant protein in this case.
Generation of an Atcrt1a mutant only displayed
a conditional phenotype in response to tunicamycin
(Fig. 7), a blocker of N-linked glycosylation and therefore
an inducer of the UPR in the ER (Liu et al. 2007). CRT
deficiencies in other species have resulted in more severe
phenotypes (Nakamura et al. 2001a). For example, CRT
deficiency in mouse causes cardiac arrest and is therefore
embryonic lethal (Nakamura et al. 2001a). We therefore
anticipated a more severe phenotype in CRT-deficient
Arabidopsis plants. The weak phenotype of Atcrt1a may
consequently suggest that other CRT isoforms, most
probably AtCRT1b, may substitute for AtCRT1a in the
Atcrt1a mutant. Should AtCRT1b functionally compensate
for AtCRT1a in the Atcrt1a background we would antici-
pate that the proteins, and possibly genes, are produced and
expressed, respectively, at similar developmental stages
and tissue types. In agreement with this, AtCRT1a and
AtCRT1b are transcriptionally tightly coordinated over
approximately 1,200 microarray data sets (Obayashi et al.
2006, Thelin et al. unpublished). Generation of double
mutants between Atcrt1a and 1b may therefore be infor-
mative for deciphering the effect of removal of CRT activity
in Arabidopsis. The expression pattern of the third CRT
isoform in Arabidopsis, AtCRT3, on the other hand, differs
substantially from that of AtCRT1a and 1b (Persson et al.
2003, Obayashi et al. 2006, Thelin et al. unpublished),
suggesting that AtCRT3 may have a role complementary
to that of the other CRT isoforms. Interestingly, several of
the well conserved amino acids that have been shown to be
essential for basic CRT functions, such as protein folding
and Ca2þ binding abilities, are not conserved in AtCRT3
(data not shown). A comparison using the mouse crt�/�
fibroblasts for AtCRT1a and AtCRT3 may therefore
reveal functional divergence among the CRT isoforms in
Arabidopsis. A more general approach comparing CRT
amino acid sequences from various species may similarly
prove suitable to identify essential amino acids, and may
allow inference of functional roles for specific CRTs.
Several other chaperones, such as CNX1, CNX2 and
SHEPHERD (SHD), are also highly co-expressed with
AtCRT1a and 1b (Obayashi et al. 2006, Thelin et al.
unpublished), indicating that a tightly knit network of
chaperone-like proteins may work together in higher plants.
Considering that CRTs and CNXs may mimic each other
during interactions with maturing proteins (Danilczyk et al.
2000), CNX functions may also partly suppress mutant
phenotypes in Atcrt1a. It may therefore be necessary also
to mutate the CNXs, in addition to the CRTs, to abolish
the ER quality control cycle completely in Arabidopsis.
Mutation in SHD, encoding a GRP94 homolog, causes
enlarged shoot and root apical meristems, and pistil
swelling (Ishiguro et al. 2002). As mentioned above, SHD
is highly co-expressed with AtCRT1a, possibly reflecting
functional relatedness between SHD and AtCRT1a.
Interestingly, AtCRT1a showed very specific expression in
the expansion zone of roots and in floral tissues, specifically
in the ovule (Fig. 1), suggesting that AtCRT1a indeed
may be important for the pistil and root development
similarly to SHD.
CRTs are believed to be the main source of Ca2þ
sequestration in plant cells (Navazio et al. 1995, Persson
and Harper 2006), and directly influence ER Ca2þ levels
in vitro (Persson et al. 2001) and in vivo (Wyatt et al. 2002).
Consistent with this, AtCRT1a restored deficiencies in Ca2þ
homeostasis in crt�/� mouse embryonic fibroblasts (Fig. 4).
As apparent in animals, CRT is a key connector of Ca2þ
signaling and ER protein folding, i.e. the disruption of
CRT in mouse impairs ER Ca2þ release most probably by
misfolding of the bradykinin receptor (Nakamura et al.
2001a). In addition, the ER Ca2þ levels are important for
ER chaperone interactions and therefore protein folding
efficiency in animal cells (Corbett et al. 1999). This implies
that CRT deficiency reduces both ER Ca2þ and the ER
protein folding efficiency. No data linking ER Ca2þ
homeostasis and ER chaperone efficiency are available for
plants. However, the Ca2þ holding ability of AtCRT1a and
the reduced resistance to the UPR, i.e. increased sensitivity
to tunicamycin, in Atcrt1a suggest that the ER Ca2þ
holding potential and the folding efficiency may also be
linked by CRTs in higher plants. Considering that the
amino acid residues necessary for interactions between
CRTs and ERp57, important for disulfide bond rearrange-
ments, are conserved in AtCRT1a, it appears likely that
these Ca2þ-dependent interactions may also be present in
plants. This hypothesis is further corroborated by restora-
tion of the putative folding deficiency in crt�/� mouse
fibroblasts by AtCRT1a complementation (Fig. 5).
Taken together, we show that the two main CRT-
associated functions, the Ca2þ-regulating and chaperone-
like functions, are conserved between plants and animals.
We also show that disruption of one of the CRTs,
AtCRT1a, causes lower stress tolerance to the UPR in
Arabidopsis. Furthermore, we believe that functional
redundancies for a multitude of CRT homologs from
different species may be explored using the mouse crt�/�
fibroblasts as described here.
Materials and Methods
Computational analysis of CRT protein sequences
Protein sequences for human CRT1, mouse CRT1 andArabidopsis CRT1a (accession Nos. AAA51916, NP_031617 and
920 Functions of calreticulins in higher plants
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
AAC49695, respectively) were obtained from the National Centerfor Biotechnology Information (NCBI; www.ncbi.nlm.nih.gov).Alignment was done using ClustalW, MacVector 8.0 software(Oxford Molecular Group plc, UK). A heuristic search usinga maximum parsimony method was performed on the alignedsequences of seven CRT isoforms from Arabidopsis, rice andhuman using the PAUP v.4b10 software (Sinauer Associates,Sunderland, MA, USA), and bootstrap values based on 1,000replicates were used.
Isolation of Arabidopsis RNA and RT–PCR
Total RNA was isolated from various Arabidopsis tissues(4 weeks old) using a Qiagen RNeasy Plant Mini Kit (Qiagen Inc.,Valencia, CA, USA). RNA (1mg) was digested with DNase(Promega, Madison, WI, USA) followed by cDNA synthesiswith an iScript cDNA Synthesis Kit (Bio-Rad, Hercules,CA, USA). Reverse transcription–PCR (RT–PCR) primers forAtCRT1a were as follows: forward primer 50AGCACAAGGATGCGGAGA-30 and reverse primer 50-TCAGAGTCGGATTCAGC-30, and were designed over exon–exon junctions.ACTIN was used as control.
Expression analysis of AtCRT1a
A 1.5 kb long nucleotide sequence upstream of the ATGstart codon comprising the promoter and 50 untranslated regionof AtCRT1a was amplified from genomic Arabidopsis DNA. Theproduct was recombined into the pENTR D-TOPO vector usingthe Gateway system (Invitrogen, Carlsbad, CA, USA) and theninserted into the pKGWFS7 destination vector (Plant SystemsBiology VIB-Ghent University, Belgium) using the LR recombina-tion mix (Invitrogen). The vector was transformed into electro-competent Agrobacterium tumefaciens (strain GV3101) andtransformed into wild-type (col-0) Arabidopsis using the floraldip method (Clough and Bent 1998). Transgenic seeds wereselected on 1/2 Murashige and Skoog (MS) agar plates with50 mgml�1 BASTA (glufosinate ammonium, Fluka, Switzerland),and third-generation seeds were analysed. To visualize GUSexpression patterns, the plant materials were incubated in GUSsubstrate solution [100mM potassium phosphate buffer, pH 7.0,1mM EDTA, 0.1% (w/v) Triton X-100, 12mg ml�1 5-bromo-4chloro-3-indolyl b-D-glucuronic acid (Duchefa Biochemie,Haarlem, The Netherlands)], vacuum infiltrated for 30min,incubated at 378C overnight and destained in 70% (v/v) ethanol.The results shown are representative for three homozygous lineswith similar expression patterns.
Expression and purification of recombinant CRT
For expression of human CRT1 and Arabidopsis CRT1a inE. coli, the full-length CRT genes were amplified and cloned intoa pET15b plasmid (Novagen, Madison, WI, USA). The 6�His-tagged proteins were expressed in BL21 (E. coli) cells in LBmedium containing 100 mg ml�1 ampicillin. Cultures were grown tomid-log phase followed by the induction of expression by 1mMisopropyl-b-D-thiogalactopyranoside (IPTG) for 4 h. Cells werecentrifuged at 4,600� g for 15min and resuspended in 50mMTris–HCl, pH 8.0, 300mM NaCl, 2mM phenylmethylsulfonylfluoride (PMSF) and protease inhibitor cocktail (10 ml ml�1)followed by sonication at 22W for 240 s, and then centrifuged at52,000� g for 20min. The lysates were mixed and incubatedovernight with Ni2þ-nitrilotriacetic acid–agarose beads (QiagenInc.) and 20mM imidazole. After incubation the beads werewashed with 50mM Tris–HCl, pH 8.0, 300mM NaCl and 50mMimidazole, and the His-tagged proteins were eluted with 50mM
Tris–HCl, pH 8.0, 300mM NaCl and 300mM imidazole. Proteindetermination was according to Bearden (1978) using a dye-binding assay with bovine serum albumin (BSA) as standard.
45Ca2þ overlays of protein blots
Purified proteins were separated by SDS–PAGE and trans-ferred to an Immobilon polyvinylidene difluoride (PVDF) transfermembrane (Millipore, Bedford, MA, USA). The membrane waswashed three times for 10min with 60mM KCl, 5mM MgCl2and 10mM imidazole-HCl, pH 6.8, and then incubated overnightwith 60mM KCl, 5mM MgCl2, 10mCi ml�1 45Ca2þ and 10mMimidazole-HCl, pH 6.8 (Maruyama et al. 1984). The membranewas then washed for 10min with deionized water and dried.Radiolabeled proteins were detected with PhosphorImager(GE Healthcare, Buckinghamshire, UK).
Generation of transgenic AtCrt1a mouse embryonic fibroblasts
A full-length AtCRT1a cDNA (GenBank accessionNo. U66343) was cloned into a pcDNA3.1/Zeo vector containingan HA tag in the C-terminus, upstream of the ER retention signal(HDEL) of AtCRT1a to generate pcDNA-CRT1a-HA (Fig. 3A).Wild-type and crt�/� mouse embryonic fibroblasts were grown at378C in a 5% CO2 environment in Dulbecco’s modified Eagle’smedium containing 10% (w/v) fetal bovine serum and 1% (w/v)pencillin streptomycin-glutamine (Nakamura et al. 2001a). crt�/�
cells were transfected with pcDNA CRT1a-HA using EffecteneTransfection Reagent (Qiagen Inc.), and stably transfected celllines were selected in the presence of 400 mg ml�1 zeocin. Six ofthese cell lines were analyzed in more detail with respect toAtCRT1a protein expression, subcellular localization, and effectson Ca2þ homeostasis and protein folding. Typical results shownare from cell lines AtCRT1a-3 and AtCRT1a-11.
SDS–PAGE and immunoblotting
SDS–PAGE was carried out essentially according to Laemmli(1970) with a Bio-Rad Mini-Protean II electrophoresis system.For mouse embryonic fibroblasts, the cells were lysed with RIPAbuffer [50mM Tris–HCl, pH 7.5, 150mM NaCl, 1mM EGTA,1mM EDTA, 1% (w/v) Triton X-100, 0.5% (w/v) deoxycholicacid, 0.1% (w/v) SDS, 1mM benzamidine, 1mM PMSF,0.025mg ml�1 aprotinin, 0.01mg ml�1 pepstatin, 0.05mg ml�1
E-64, 0.025mg ml�1 leupeptin and 0.1mg ml�1 tosyl phenylalanylchloromethyl ketone (TPCK)] and proteins were solubilized byaddition of sample buffer [125mM Tris–HCl, pH 6.8, 4%(w/v) SDS, 20% (v/v) glycerol, 5% b-mercaptoethanol and0.02% bromphenol blue], and separated using a 12% SDS–polyacrylamide gel. For immunoblotting analysis, proteins wereelectrophoretically transferred to a PVDF membrane (Millipore)for 1 h at 100V. After transfer, the membrane was blocked with5% blocking solution [5% (w/v) skimmed milk powder in 10mMTris–HCl, pH 8.0, 150mM NaCl], and washed in 10mMTris–HCl, pH 8.0, 150mM NaCl and 0.05% (w/v) Tween-20.For mouse embryonic fibroblasts, the blotting membranes wereprobed with antibodies against the HA tag (1 : 1,000). The HA tagantibodies were detected with anti-mouse IgG–horseradishperoxidase secondary antibodies (1 : 10,000). For plants, themembrane was probed with maize CRT antibodies (1 : 10,000) toscreen for Atcrt1a mutants together with secondary antibodiesanti-rabbit IgG coupled to horseradish peroxidase (1 : 10,000).The immunodecorated bands were visualized using the enhancedchemiluminescence detection method.
Functions of calreticulins in higher plants 921
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
Immunohistochemistry of mouse embryonic fibroblasts expressingAtCRT1a
For intracellular localization of AtCRT1a in mouse embryo-nic fibroblasts, cells were cultured on coverslips, washed withphosphate-buffered saline (PBS; 5mM potassium phosphatebuffer, pH 7.5, 150mM NaCl), fixed with 4% (w/v) formaldehyde,extracted with 0.1% (w/v) saponin, 2% (w/v) milk powder in PBS,and incubated with HA tag monoclonal antibodies (1 : 50) andCNX polyclonal antibodies (Groenendyk et al. 2006; 1 : 800).The HA tag antibodies were detected with fluorescence AlexaFluor anti-mouse secondary antibodies at �em¼ 680 nm and theCNX antibodies with fluorescence Alexa Fluor anti-rabbitsecondary antibodies at �em¼ 546 nm.
Measurements of ER Ca2þ capacity
Mouse embryonic fibroblasts were cultured for 50 h in thepresence of 55 mCi of 45Ca2þ (Nakamura et al. 2001a). Cells werewashed twice with 5mM EDTA in PBS, and then once with 5mMEDTA in culture medium (without fetal bovine serum), detachedfrom Petri dishes by trypsinization [0.25% (w/v) trypsin and 0.02%(w/v) EDTA in Ca2þ/Mg2þ-free PBS], and finally resuspended inCa2þ-free buffer [143mMNaCl, 6mM KCl, 1mM MgSO4, 20mMHEPES-NaOH, pH 7.4, 0.1% (w/v) glucose and 0.1mg ml�1
sulfinpyrazone]. For 45Ca2þ release experiments, aliquots of5� 106 cells ml�1 were pre-incubated for 3min at 378C followedby treatment with thapsigargin or ionomycin for 4min at 378C.Cells were then pelleted by centrifugation (9,600� g for 3min) andthe radioactivity in the supernatant was measured in a BeckmanLS 7800 scintillation counter. Background values, i.e. radioactivityrecovered in the supernatant in control incubations withoutadditions of thapsigargin or ionomycin, were subtracted prior topresentations in the figures.
Fluorescence Ca2þ measurements of mouse embryonic fibroblasts
Cells were incubated with 2 ml ml�1 Fura 2-AM (Sigma-Aldrich) essentially as described in Nakamura et al. (2001a),washed with PBS, trypsinized [0.25% (w/v) trypsin and 0.02%(w/v) EDTA in Ca2þ/Mg2þ-free PBS], centrifuged, resuspended inCa2þ buffer [143mM NaCl, 6mM KCl, 1mM MgSO4, 20mMHEPES-NaOH, pH 7.4, 0.1% (w/v) glucose, 1mM CaCl2and 0.1mg ml�1 sulfinpyrazone], centrifuged and resuspended inCa2þ-free buffer [143mMNaCl, 6mM KCl, 1mM MgSO4, 20mMHEPES, pH 7.4, 0.1% (w/v) glucose and 0.1mg ml�1 sulfinpyr-azone]. The cells were transferred to a cuvette and 2mM EGTAwas added. Fluorescence was measured at �ex¼ 340 nm. When theresting free cytoplasmic Ca2þ level of the cells (basal Ca2þ level)was reached, the cells were stimulated with 600 nM bradykinin(Fig. 5A, B). Thapsigargin (300 nM), a SERCA (ER Ca2þ-ATPase)inhibitor, was later added to the cells to measure the Ca2þ store inthe ER and to control that no Ca2þ release of bradykinin-inducedcells was due to incomplete Ca2þ storage functions in the ER(Fig. 5A). To measure the store-operated Ca2þ influx, 2mMCaCl2 was added to the cells. Ionomycin (7.5mM) together withCaCl2 (4mM) was added to the cells to obtain the maximum value,and to obtain the minimum value 32mM EGTA, 24mMTris–HCl, pH 7.4 and 0.4% (w/v) Triton X-100 were added tothe cells.
[3H]Bradykinin binding assay
Cells were plated at 2� 104 cells ml�1 in 24-well plates for24 h prior to the binding experiments. The culture medium wasremoved and cells were equilibrated on ice for 10min with bindingbuffer (20mMHEPES-NaOH, pH 7.4, 17mMNaCl, 5.4mMKCl,
0.44mM KH2PO4, 0.63mM CaCl2, 0.21mM MgSO4, 0.34mMNa2HPO4, 110mM N-methylglucamine, 0.1% (w/v) BSA and2mM bacitracin; Yang et al. 1999). Cells were then incubatedin the presence of different concentrations of [3H]bradykinin(0.1–20 nM) in the presence or absence of 10 mM unlabeledbradykinin. After 4 h incubation, cells were washed three timeswith a binding buffer, re-suspended in 250 ml of 100mMNaOH perwell, harvested and counted by scintillation counting. The proteinconcentration was measured by DC Protein Assay Bio-Rad.The amount of specific binding was calculated as the total bindingminus the binding in the presence of 10mM unlabeled bradykinin;binding curves were fitted using a Prism program (GraphPad,San Diego, CA, USA).
Screening of the Atcrt1a mutant
Arabidopsis seeds with a T-DNA insertion in the CRT1a genewere obtained from the SALK collection (SALK_055452; Alonsoet al. 2003). To screen for homozygous plants, DNA was extractedusing the CTAB/chloroform-method (CTAB; 2% N-cetyl-NNN-trimethyl-ammonium bromide, 100mM Tris–HCl, pH 8.0, 20mMEDTA and 1.4M NaCl; Murray and Thompson 1980). A T-DNAleft border primer for the insert 50-CGC CCT GAT AGA CGGTTT TTC-30 was used and primers were made approximately600 bp away from and on each side of the insert position: forward50 CCA CCT TTG CTT AGA GCT CGT CC-30, reverse 50-TTGTAA GAT TTT TGT TTTCCCTG-30. For RNA expression, totalRNA was isolated using a Qiagen RNeasy Plant Mini Kit(Qiagen Inc.). RNA (1 mg) was digested with DNase (Promega)followed by cDNA synthesis with an iScript cDNA Synthesis Kit(Bio-Rad). RT–PCR primers for AtCRT1a were as follows:upstream of the T-DNA insert, forward primer 50-TGGCGAAACTAAACCCTAAA-30, and reverse primer 50-ATCTCTTCTCCCATCCATCCATCTT-30, and downstream of the T-DNAinsert, forward primer 50-AGCACAAGGATGCGGAGA-30 andreverse primer 50-TCAGCGTCGGATTCAGC. Primers weredesigned over exon–exon junctions. ACTIN1 was used as control.For protein expression analyses, seedlings were ground with sandin a homogenization buffer (50mM MOPS-KOH, pH 7.5, 0.33Msucrose, 5mM EDTA) with 2.5ml of buffer per g of plant material.The samples were centrifuged for 15min at 5,000�g and thesupernatants were collected for immunoblot analyses.
Complementation of Atcrt1a
A full-length AtCRT1a cDNA (GenBank accession No.U66343) was cloned into a p35SBARN binary vector (Cloughand Bent 1998). The construct was transformed into electrocom-petent A. tumefaciens GV3101, and transformed into the Atcrt1amutant by floral dip (Clough and Bent 1998). Transformed plantswere grown on 1/2 MS medium with 0.8% agar containing25mg ml�1 BASTA (resistance conferred from the AtCRT1atransgene).
Tunicamycin treatment of Arabidopsis seedlings
Growth experiment. Arabidopsis plants were germinated andgrown for 1 week on 1/2 MS medium with 0.8% (w/v) agar at 238Cwith 16 h of light (100mEm�2 s�1), then transferred to 1/2 MSmedium with 0.8% agar� 0.1mg ml�1 tunicamycin and grownfor another 8 d. Control plants (15 d old; no tunicamycin) andtunicamycin-treated plants were photographed, and leaf length,fresh weight and dry weight were measured.
Western blotting experiment. Arabidopsis seedling cultureswere germinated and grown in 250ml flasks [100ml per flaskof Gamborg B5 salt, 1.5% (w/v) sucrose and 0.5mg ml�1
922 Functions of calreticulins in higher plants
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
MES-KOH, pH 5.7]. The cultures were kept on a rotary shaker(125 rpm) for 15 d at 238C, with 16 h of light. At the end ofthe growth period, the medium was supplemented with either0.1 mg ml�1 tunicamycin for 8 d or 5mg ml�1 tunicamycin for 6 h.Seedlings (50mg) from the different treatments and untreatedcontrol were homogenized in 2� SDS–PAGE loading bufferwith 1 ml of protease inhibitor cocktail per 100 ml of buffer. Thesamples were then centrifuged for 2min at 3,000 rpm and 10 mlof the sample supernatants were subjected to SDS–PAGE andimmunoblot analysis.
Statistics
The statistical significance of differences between treatmentsor measurements of different cell lines was assessed by Student’st-test (Microsoft Office Excel, Microsoft Corporation, Redmond,WA, USA). The number of replicates and the level of significanceis given in the text or figure legends.
Funding
The Swedish Research Council (to M.S.); Swedish
Foundation for Strategic Research (to M.S.), The Swedish
Research Council Formas (to M.S.); Carl Tryggers
Foundation (to S.W.); Canadian Institutes of Health
Research (Grant MOP-1529 to M.M.); Alberta Heritage
Foundation for Medical Research (M.M.). J.J. is a recipient
of a studentship award from the Alberta Heritage
Foundation for Medical Research.
Acknowledgments
We are grateful to Monika Dabrowska for superb technicalhelp, Dr. Magnus Alsterfjord for help with the AtCRT1a promoter:GUSconstruct, Lisa Thelin for help with the phylogenetic tree, andDr. Peter Olsson for the gift of the recombinant PEPC protein.
References
Akesson, A., Persson, S., Love, J., Boss, W.F., Widell, S. andSommarin, M. (2005) Overexpression of the Ca2þ-binding proteincalreticulin in the endoplasmic reticulum improves growth of tobaccocell suspensions (Nicotiana tabacum, L.) in high Ca2þ-medium. Physiol.Plant. 123: 92–99.
Alonso, J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H., et al.(2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana.Science 301: 653–657.
Arnaudeau, S., Frieden, M., Nakamura, K., Castelbou, C., Michalak, M.and Demaurex, N. (2002) Calreticulin differentially modulates calciumuptake and release in the endoplasmic reticulum and mitochondria.J. Biol. Chem. 277: 46696–46705.
Baksh, S. and Michalak, M. (1991) Expression of calreticulin in Escherichiacoli and identification of its Ca2þ binding domains. J. Biol. Chem. 266:21458–21465.
Baluska, F., Samaj, J., Napier, R. and Volkmann, D. (1999) Maizecalreticulin localizes preferentially to plasmodesmata in root apex.Plant J. 19: 481–488.
Baumann, O. and Walz, B. (2001) Endoplasmic reticulum of animal cellsand its organization into structural and functional domains. Int. Rev.Cytol. 205: 149–214.
Bearden, J.Jr (1978) Quantification of submicrogram quantities of proteinby an improved protein–dye binding assay. Biochim. Biophys. Acta 533:525–529.
Borisjuk, N., Sitailo, L., Adler, K., Malysheva, L., Tewes, A., Borisjuk, L.and Manteuffel, R. (1998) Calreticulin expression in plant cells:developmental regulation, tissue specificity and intracellular distribution.Planta 206: 504–514.
Burns, K., Duggan, B., Atkinson, E.A., Famulski, K.S., Nemer, M.,Bleackley, R.C. and Michalak, M. (1994) Modulation of gene expressionby calreticulin binding to the glucocorticoid receptor. Nature 367:476–480.
Camacho, P. and Lechleiter, J.D. (1995) Calreticulin inhibits repetitiveintracellular Ca2þ waves. Cell 82: 765–771.
Chen, M.-H., Tian, G.-W., Gafni, Y. and Citosvsky, V. (2005) Effectsof calreticulin on viral cell-to-cell movement. Plant Physiol. 138:1866–1876.
Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified methodfor Agrobacterium-mediated transformation of Arabidopsis thaliana.Plant J. 16: 735–743.
Coppolino, M.G., Woodside, M.J., Demaurex, N., Grinstein, S.,St-Arnaud, R. and Dedhar, S. (1997) Calreticulin is essential forintegrin-mediated calcium signalling and cell adhesion. Nature 386:843–847.
Corbett, E.F., Oikawa, K., Francois, P., Tessier, D.C., Kay, C.,Bergeron, J.J., Thomas, D.Y., Krause, K.H. and Michalak, M. (1999)Ca2þ regulation of interactions between endoplasmic reticulumchaperones. J. Biol. Chem. 274: 6203–6211.
Crofts, A.J. and Denecke, J. (1998) Calreticulin and calnexin in plants.Trends Plant Sci. 3: 396–399.
Culina, S., Lauvau, G., Gubler, B. and van Endert, P.M. (2004) Calreticulinpromotes folding of functional human leukocyte antigen class I moleculesin vitro. J. Biol. Chem. 279: 54210–54215.
Danilczyk, U.G., Cohen-Doyle, M.F. and Williams, D.B. (2000) Functionalrelationship between calreticulin, calnexin, and the ER luminal domain ofcalnexin. J. Biol. Chem. 275: 13089–13097.
Dedhar, S., Rennie, P.S., Shago, M., Hagesteijn, C.Y., Yang, H., Filmus, J.,Hawley, R.G., Bruchovsky, N., Cheng, H. and Matusik, R.J. (1994)Inhibition of nuclear hormone receptor activity by calreticulin. Nature
Zeijl, M.J., Sinjorgo, K.M. and Palva, E.T. (1995) The tobacco homologof mammalian calreticulin is present in protein complexes in vivo.Plant Cell 7: 391–406.
Groenendyk, J. and Michalak, M. (2005) Endoplasmic reticulum qualitycontrol and apoptosis. Acta Biochim. Pol. 52: 381–395.
Groenendyk, J., Zuppini, A., Shore, G., Opas, M., Bleackley, R.C. andMichalak, M. (2006) Caspase 12 in calnexin-deficient cells. Biochemistry45: 13219–13226.
Hashii, M., Nozawa, Y. and Higashida, H. (1993) Bradykinin-inducedcytosolic Ca2þ oscillations and inositol tetrakisphosphate-induced Ca2þ
influx in voltage-clamped ras-transformed NIH/3T3 fibroblasts. J. Biol.Chem. 268: 19403–19410.
Ishiguro, S., Watanabe, Y., Ito, N., Nonaka, H., Takeda, N., Sakai, T.,Kanaya, H. and Okada, K. (2002) SHEPHERD is the ArabidopsisGRP94 responsible for the formation of functional CLAVATA proteins.EMBO J. 21: 898–908.
Laemmli, U.K. (1970) Cleavage of structural proteins during assembly ofthe head of bacteriophage T4. Nature 227: 680–685.
LeClere, S. and Bartel, B. (2001) A library of Arabidopsis 35S-cDNA linesfor identifying novel mutants. Plant Mol. Biol. 46: 695–703.
Liu, J.X., Srivastava, R., Che, P and Howell, S.H. (2007) An endoplasmicreticulum stress response in Arabidopsis is mediated by proteolyticprocessing and nuclear relocation of a membrane-associated transcrip-tion factor, bZIP28. Plant Cell 19: 4111–4119.
Martin, V., Groenendyk, J., Steiner, S.S., Guo, L., Dabrowska, M.,Parker, J.M., Muller-Esterl, W., Opas, M. and Michalak, M. (2006)Identification by mutational analysis of amino acid residues essential inthe chaperone function of calreticulin. J. Biol. Chem. 281: 2338–2346.
Maruyama, K., Mikawa, T. and Ebashi, S. (1984) Detection of calciumbinding proteins by 45Ca autoradiography on nitrocellulose membraneafter sodium dodecyl sulfate gel electrophoresis. J. Biochem. 95: 511–519.
Mery, L., Mesaeli, N., Michalak, M., Opas, M., Lew, D.P. andKrause, K.H. (1996) Overexpression of calreticulin increases
Functions of calreticulins in higher plants 923
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022
Michalak, M., Corbett, E.F., Mesaeli, N., Nakamura, K. and Opas, M.(1999) Calreticulin: one protein, one gene, many functions. Biochem. J.344: 281–292.
Murray, M.G. and Thompson, W.F. (1980) Rapid isolation of highmolecular weight plant DNA. Nucleic Acids Res. 8: 4321–4325.
Nakamura, K., Robertson, M., Liu, G., Dickie, P., Nakamura, K.,Guo, J.Q., Duff, H.J., Opas, M., Kavanagh, K. and Michalak, M.(2001a) Complete heart block and sudden death in mice overexpressingcalreticulin. J. Clin. Invest. 107: 1245–1253.
Nakamura, K., Zuppini, A., Arnaudeau, S., Lynch, J., Ahsan, I., et al.(2001b) Functional specialization of calreticulin domains. J. Cell Biol.154: 961–972.
Napier, R.M., Treman, S., Henderson, J., Boyce, J.M., Hawes, C.,Fricker, M.D. and Venis, M.A. (1995) Purification, sequencing andfunctions of calreticulin from maize. J. Exp. Bot. 46: 1603–1613.
Navazio, L., Baldan, B., Dainese, P., James, P., Damiani, E., Margreth, A.and Mariani, P. (1995) Evidence that spinach leaves express calreticulinbut not calsequestrin. Plant Physiol. 109: 983–990.
Navazio, L., Miuzzo, M., Royle, L., Baldan, B., Varotto, S., Merry, A.H.,Harvey, D.J., Dwek, R.A., Rudd, P.M. and Mariani, P. (2002)Monitoring endoplasmic reticulum-to-Golgi traffic of a plant calreticulinby protein glycosylation analysis. Biochemistry 41: 14141–14149.
Nelson, D.E., Glaunsinger, B. and Bohnert, H.J. (1997) Abundantaccumulation of the calcium-binding molecular chaperone calreticulinin specific floral tissues of Arabidopsis thaliana. Plant Physiol. 114: 29–37.
Obayashi, T., Kinoshita, K., Nakai, K., Shibaoka, M., Hayashi, S.,Saeki, M., Shibata, D., Saito, K. and Ohta, H. (2006) ATTED-II:a database of co-expressed genes and cis elements for identifyingcoregulated gene groups in Arabidopsis. Nucleic Acids Res. 35: 863–869.
Opas, M., Tharin, S., Milner, R.E. and Michalak, M. (1996) Identificationand localization of calreticulin in plant cells. Protoplasma 191: 164–171.
Persson, S. and Harper, J. (2006) The ER and cell calcium. In ThePlant Endoplasmic Reticulum. Edited by Robinson, D.G. Plant CellMonographs. Springer Verlag, Heidelberg.
Persson, S., Rosenquist, M. and Sommarin, M. (2002) Identification ofa novel calreticulin isoform (Crt2) in human and mouse. Gene 297:151–158.
Persson, S., Rosenquist, M., Svensson, K., Galvao, R., Boss, W.F. andSommarin, M. (2003) Phylogenetic analyses and expression studies revealtwo distinct groups of calreticulin isoforms in higher plants. PlantPhysiol. 133: 1385–1396.
Persson, S., Wyatt, S.E., Love, J., Thompson, W.F., Robertson, D. andBoss, W.F. (2001) The Ca(2þ) status of the endoplasmic reticulumis altered by induction of calreticulin expression in transgenic plants.Plant Physiol. 126: 1092–1104.
Peterson, R., Ora, A., Van, P.N. and Helenius, A. (1995) Transient, lectin-like association of calreticulin with folding intermediates of cellular andviral glycoproteins. Mol. Biol. Cell 6: 1173–1184.
Thastrup, O., Cullen, P.J., Drobak, B.K., Hanley, M.R. and Dawson, A.P.(1990) Thapsigargin, a tumor promoter, discharges intracellular Ca2þ
stores by specific inhibition of the endoplasmic reticulum Ca2-ATPase.Proc. Natl Acad. Sci. USA 87: 2466–2470.
Trewavas, A.J. and Malho, R. (1998) Ca2þ signalling in plant cells: the bignetwork! Curr. Opin. Plant Biol. 1: 428–433.
Wyatt, S.E, Tsou, P.L. and Robertson, D. (2002) Expression of the highcapacity calcium-binding domain of calreticulin increases bioavailablecalcium stores in plants. Transgenic Res. 11: 1–10.
Yang, C.M., Tsai, Y.J., Pan, S.L., Wu, W.B., Wang, C.C., Lee, Y.S.,Lin, C.C., Huang, S. and Chiu, C.T. (1999) Pharmacological andfunctional characterization of bradykinin receptors in rat culuredvascular smooth muscle cells. Cell Signal. 11: 853–862.
Zimmermann, P., Hirsch-Hoffmann, M., Hennig, L. and Gruissem, W.(2004) GENEVESTIGATOR. Arabidopsis microarray database andanalysis toolbox. Plant Physiol. 136: 2621–2632.
(Received January 23, 2008; Accepted April 15, 2008)
924 Functions of calreticulins in higher plants
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/6/912/1811667 by guest on 03 February 2022