387 N G-Nitro L-Arginine Methyl Ester and Other Alkyl Esters of Arginine are Muscarinic Receptor Antagonists lain L. 0. Buxton, Dennis J. Cheek, Delrae Eckman, David P. Westfall, Kenton M. Sanders, and Kathleen D. Keef Analogues of L-arginine with modifications at the terminal guanidino nitrogen and/or the carboxyl terminus of the molecule have been widely used for their ability to inhibit the production of nitric oxide and are thought to be competitive antagonists of nitric oxide synthase. The present studies were designed to test the possibility that these agents are also muscarinic receptor antagonists. Acetylcholine produced concentration-dependent contraction of endothelium-denuded rabbit coronary artery as well as isolated strips of canine colonic smooth muscle. The arginine analogue NG-nitro L-arginine methyl ester (L-NAME, 100 p,M) but not NG-monomethyl L-arginine (L-NMMA, 100 ,uM) significantly shifted these contractile relations to the right, an effect that was not reversed by addition of 1 mM L-arginine. In radioligand binding studies using the muscarinic radioligand [3H]quinuclidinyl benzilate and tissues known to contain differing contributions of Ml, M2, and M3 muscarinic receptors, addition of increasing concentrations of L-NAME resulted in a monophasic competition of binding with affinities (K1) ranging from 68 juM in endothelium to 317 ,M in whole aorta. Addition of the hydrolysis-resistant guanosine 5'-triphosphate analogue GTPyS (100 gM) had no effect on L-NAME competition of [3H]quinuclidinyl benzilate binding. Addition of L-NAME in radioligand binding competition studies using the agonist carbachol did not result in an alteration of the receptor's affinity for agonist, confirming the competitive nature of the interaction of L-NAME with the muscarinic receptor. Several L-arginine analogues with alkyl ester modifications at the carboxyl end of the molecule as well as those without this modification were evaluated as muscarinic antagonists in radioligand binding experiments. Only those arginine compounds with a modified carboxyl group were able to compete for radioligand binding to the muscarinic receptor. Our results indicate that alkyl esters of L-arginine are muscarinic antagonists and suggest that these compounds are poor choices as nitric oxide synthase inhibitors in studies in which muscarinic receptors are not blocked. (Circulation Research 1993;72:387-395) KEY WoRDs * nitric oxide * nitric oxide synthase inhibitors * NG-nitro L-arginine methyl ester L muscarinic receptors * smooth muscle * radioligand binding In 1988, Palmer et a1l demonstrated that L-arginine was the precursor for nitric oxide (NO) synthesis in endothelial cells. Since that time, significant interest has been focused on the study of the synthesis, release, and subsequent actions of NO (for review see Moncada et a12). The effects of NO that have been studied to date include relaxation of smooth muscles. In blood vessels, NO is known to dilate vascular smooth muscle3 and may contribute to the moment-to-moment maintenance of blood vessel tone. In the gastrointesti- nal tract, NO is thought to be a nonadrenergic non- cholinergic neurotransmitter responsible for inhibition of colonic motility.4 NO is also thought to be released by From the Departments of Pharmacology (I.L.O.B., D.J.C., D.P.W.) and Physiology (D.E., K.M.S., K.D.K.), University of Nevada School of Medicine, Reno. Supported by National Institutes of Health grants P01- DK41315, HL-40399 (K.D.K.), and HL-35416 (I.L.O.B.), and a grant from the American Heart Association, Nevada Affiliate, Inc. Address for correspondence: Dr. lain L. 0. Buxton, University of Nevada, Reno, School of Medicine, Department of Pharmacol- ogy/318, Reno, NV 89557-0046. Received May 26, 1992; accepted October 22, 1992. macrophages and may play a role in signaling in the immune system.2 Numerous congeners of arginine that block the action of nitric oxide synthetase (NOS) in several tissues including endothelium, nerve, and smooth muscle have been described.2 The simplest of these compounds, N0-nitro L-arginine (L-NA), is an inhibitor of NOS both in vivo and in vitro5 and is widely used as its methyl ester analogue NG-nitro L-arginine methyl ester (L-NAME), which is more lipophilic and therefore useful for exper- iments in vivo.6 In studies designed to elucidate the role of NO in the gastrointestinal tract, an inhibitory effect of L-NAME on cholinergic neural responses was some- times observed. This inhibitory effect would be consis- tent with an action of L-NAME at the muscarinic receptor. Indeed, although differences in the actions of NOS inhibitors such as L-NAME in vivo have been attributed to differences in metabolism and tissue dis- tribution,5'6 we have hypothesized that some of these effects could be due to muscarinic receptor blockade. Thus, we report here for the first time the ability of L-NAME, L-arginine methyl ester (L-AME), and other L-arginine alkyl esters to act as muscarinic antagonists. by guest on August 30, 2017 http://circres.ahajournals.org/ Downloaded from
Transcript
387
N G-Nitro L-Arginine Methyl Ester and OtherAlkyl Esters of Arginine are Muscarinic
Receptor Antagonistslain L. 0. Buxton, Dennis J. Cheek, Delrae Eckman, David P. Westfall,
Kenton M. Sanders, and Kathleen D. Keef
Analogues of L-arginine with modifications at the terminal guanidino nitrogen and/or the carboxylterminus of the molecule have been widely used for their ability to inhibit the production of nitric oxideand are thought to be competitive antagonists of nitric oxide synthase. The present studies were designedto test the possibility that these agents are also muscarinic receptor antagonists. Acetylcholine producedconcentration-dependent contraction of endothelium-denuded rabbit coronary artery as well as isolatedstrips of canine colonic smooth muscle. The arginine analogue NG-nitro L-arginine methyl ester(L-NAME, 100 p,M) but not NG-monomethyl L-arginine (L-NMMA, 100 ,uM) significantly shifted thesecontractile relations to the right, an effect that was not reversed by addition of 1 mM L-arginine. Inradioligand binding studies using the muscarinic radioligand [3H]quinuclidinyl benzilate and tissuesknown to contain differing contributions of Ml, M2, and M3 muscarinic receptors, addition of increasingconcentrations of L-NAME resulted in a monophasic competition of binding with affinities (K1) ranging
from 68 juM in endothelium to 317 ,M in whole aorta. Addition of the hydrolysis-resistant guanosine5'-triphosphate analogue GTPyS (100 gM) had no effect on L-NAME competition of [3H]quinuclidinylbenzilate binding. Addition of L-NAME in radioligand binding competition studies using the agonistcarbachol did not result in an alteration of the receptor's affinity for agonist, confirming the competitivenature of the interaction ofL-NAME with the muscarinic receptor. Several L-arginine analogues with alkylester modifications at the carboxyl end of the molecule as well as those without this modification were
evaluated as muscarinic antagonists in radioligand binding experiments. Only those arginine compoundswith a modified carboxyl group were able to compete for radioligand binding to the muscarinic receptor.Our results indicate that alkyl esters of L-arginine are muscarinic antagonists and suggest that thesecompounds are poor choices as nitric oxide synthase inhibitors in studies in which muscarinic receptorsare not blocked. (Circulation Research 1993;72:387-395)KEY WoRDs * nitric oxide * nitric oxide synthase inhibitors * NG-nitro L-arginine methyl ester L
In 1988, Palmer et a1l demonstrated that L-argininewas the precursor for nitric oxide (NO) synthesisin endothelial cells. Since that time, significant
interest has been focused on the study of the synthesis,release, and subsequent actions of NO (for review seeMoncada et a12). The effects of NO that have beenstudied to date include relaxation of smooth muscles. Inblood vessels, NO is known to dilate vascular smoothmuscle3 and may contribute to the moment-to-momentmaintenance of blood vessel tone. In the gastrointesti-nal tract, NO is thought to be a nonadrenergic non-cholinergic neurotransmitter responsible for inhibitionof colonic motility.4 NO is also thought to be released by
From the Departments of Pharmacology (I.L.O.B., D.J.C.,D.P.W.) and Physiology (D.E., K.M.S., K.D.K.), University ofNevada School of Medicine, Reno.Supported by National Institutes of Health grants P01-
DK41315, HL-40399 (K.D.K.), and HL-35416 (I.L.O.B.), and agrant from the American Heart Association, Nevada Affiliate, Inc.
Address for correspondence: Dr. lain L. 0. Buxton, Universityof Nevada, Reno, School of Medicine, Department of Pharmacol-ogy/318, Reno, NV 89557-0046.
Received May 26, 1992; accepted October 22, 1992.
macrophages and may play a role in signaling in theimmune system.2Numerous congeners of arginine that block the action
of nitric oxide synthetase (NOS) in several tissuesincluding endothelium, nerve, and smooth muscle havebeen described.2 The simplest of these compounds,N0-nitro L-arginine (L-NA), is an inhibitor ofNOS bothin vivo and in vitro5 and is widely used as its methyl esteranalogue NG-nitro L-arginine methyl ester (L-NAME),which is more lipophilic and therefore useful for exper-iments in vivo.6 In studies designed to elucidate the roleof NO in the gastrointestinal tract, an inhibitory effectof L-NAME on cholinergic neural responses was some-times observed. This inhibitory effect would be consis-tent with an action of L-NAME at the muscarinicreceptor. Indeed, although differences in the actions ofNOS inhibitors such as L-NAME in vivo have beenattributed to differences in metabolism and tissue dis-tribution,5'6 we have hypothesized that some of theseeffects could be due to muscarinic receptor blockade.Thus, we report here for the first time the ability ofL-NAME, L-arginine methyl ester (L-AME), and otherL-arginine alkyl esters to act as muscarinic antagonists.
388 Circulation Research Vol 72, No 2 February 1993
The affinity of muscarinic receptors for these com-pounds suggests that experiments using them may yieldboth muscarinic receptor antagonism and inhibition ofNOS activity in cells, tissues, or whole animals.
Materials and MethodsTissue PreparationMongrel dogs or guinea pigs of either sex were
anesthetized with pentobarbital sodium (30 mg/kg). Indogs, a segment of proximal colon (6-14 cm from theileocecal sphincter), the submandibular glands, rightand left atria, coronary vessels, and aorta (15-20 cmincluding the aortic arch) were removed and kept inoxygenated (97% 02-3% C02) Krebs-Ringer bicarbon-ate solution (KRB) containing (mM) Na+ 137.4, K' 5.9,Ca2+ 2.5, Cl- 134, HCO3- 15.5, H2P04- 1.2, and dex-trose 11.5. In guinea pigs, the brain was removed andkept on ice in the same solution. Atria, brain, bloodvessels, and submandibular gland were trimmed toremove adherent fat or connective tissue and minced inKRB to remove blood cells. In aorta from three dogs,the vessels were opened longitudinally, and the endo-thelium was removed by scraping and collected bycentrifugation. After washing out fecal material, thecolon was cut into 2x3-cm pieces and pinned out in adissecting dish bathed with fresh KRB. The longitudinalsmooth muscle was carefully removed, and the circularsmooth muscle was separated from the submucosa bythe method of Smith et al.7
Contractile ExperimentsMale rabbits (2-3 kg) were anesthetized with an
overdose of pentobarbital sodium (30 g/kg) injectedinto the ear vein. The heart was removed and placed incold (10°C) oxygenated KRB. A 1-1.5-cm segment ofthe left ventricular coronary artery was carefully dis-sected, cut into ring segments (3 mm long), and de-nuded of endothelium by gentle rubbing with a stain-less-steel pin. Denuded segments were mounted on twotriangular tungsten supports (89-pim diameter). Theupper triangle was attached to a Gould strain gauge,and the lower was attached to a stable mount. A restingforce of 0.6 g was applied to vessel segments in a tissuebath maintained at 37°C filled with oxygenated KRB(pH 7.4). Tissues were equilibrated for at least 1 hourand contracted at 10-15-minute intervals with 60 mMKCl before beginning the experiments. The absence ofendothelium was assessed during the experiment bymeasuring the response to acetylcholine (ACh, 0.1-10,uM) in segments contracted with the histamine Hl-receptor agonist 2-(2-aminoethyl)pyridine.89 In endo-thelium-intact vessels, ACh (10 ,M) produced nearmaximal relaxation, whereas in denuded segments, AChproduced contraction (see Keef et a19).
Colonic smooth muscle strips (10 mmxl mm) weremade by cutting across the entire muscularis parallel tothe circular muscle fibers. Muscles were then attachedwith suture to a stable mount and to a Gould straingauge and immersed in a tissue bath containing KRBmaintained at 37°C. A resting tension of 1 g was applied,and tissues were equilibrated for 1 hour, during whichtime the KRB was exchanged every 15 minutes. Con-tractile experiments in canine colon were performed in
ence from enteric nerves. Before the start of the exper-iments, tissues were repeatedly contracted with 1 ,uMACh, followed by 10-minute rest periods until a stableamplitude of contraction was obtained. Peak contractiletension was then determined in the presence of 100 ,uMACh. Tissues were then reequilibrated with repeatedapplications of 1 ,uM ACh as before. The response tocumulative additions of ACh was then measured in theabsence or presence of a 100-,tM concentration of theL-arginine analogues L-NAME or NG-monomethyl L-ar-ginine (L-NMMA). Responses from each tissue stripwere normalized to the peak ACh-induced contractionobtained with the first addition of 100 ,uM ACh.
Membrane PreparationMembranes from all tissues were prepared in a
similar fashion; tissues were washed three times withice-cold hypotonic buffer A containing (mM) Tris base50, MgCl2 10, and EGTA 1, pH 7.4, and homogenized in10 vol of the same ice-cold buffer using a Polytron (threetimes for 15 seconds at setting 8). The homogenate wasfiltered through a nylon cloth (500 ,um) and centrifugedat 1,000g for 10 minutes (4°C) to remove unwantedmaterial. The resulting pellet was discarded, and thesupernatant was centrifuged at 80,000g for 60 minutes at4°C. The resulting pellet containing plasma membranewas quickly frozen in liquid nitrogen and stored as afrozen powder at -80C for 1-2 weeks until used inradioligand binding studies. Membrane pellets fromthree animals were pooled.
Radioligand Binding StudiesBinding of the nonselective muscarinic receptor an-
tagonist ['Hlquinuclidinyl benzilate (['HIQNB) wasmeasured by a rapid filtration method similar to thatdescribed previously.10 Membrane pellets were resus-pended in buffer A to yield approximately 0.4 mg/mlprotein as determined by the method of Bradford.'1Radioligand binding assays were carried out in a volumeof 500 ,p1 consisting of 440 gl membrane suspension, 50,ul radioligand, and 10 ,l buffer or drug. Saturationbinding experiments used concentrations of ['H]QNBfrom 20 to 5,000 pM, and nonspecific binding wasdetermined by the addition of 1 ,uM atropine. Equilib-rium binding, carried out at 30°C for 75 minutes, wasstable for 120 minutes and completely reversible. Boundand free radioligand were separated by rapid vacuumfiltration over GF/C filters (Whatman Inc., Clifton, N.J.)that had been pretreated with 0.3% polyethylenimine asdescribed by Bruns et al.'2 The filters were washed withtwo 5 -ml aliquots of ice-cold buffer containing (mM)Tris-HCl 5, MgCl2 1, and EGTA 0.1 and counted at 48%efficiency in a liquid scintillation counter (model LS60001C, Beckman Instruments, Inc., Fullerton, Calif.).All determinations were made in triplicate, and specificbinding of ['H]QNB at 0.3 nM was examined in compe-tition studies using increasing concentrations of unla-beled agonists and antagonists.
Data AnalysisSaturation and competition binding curves were an-
alyzed using the computer program GRAPHPAD INPLOT(version 3.0, GraphPad Software, San Diego, Calif.).This program uses nonlinear least-squares regression to
the presence of 1 gM tetrodotoxin to reduce interfer- fit binding data to equations that adhere to the laws of
Buxton et al L-NAME Is a Muscarinic Antagonist 389
A. Control
ACh
B.
z0
I-
z0
:2
0e
100pM L-NAME
0.5 9
AC 2 minACh
C. Control
jVW n4Iv4wACh
D.
100 pM L-NAME
ACh
AC 2 min
C-
c
0z
0H0z
-LOG [ACETYLCHOLINE]FIGURE 1. Effects of NG-nitro L-arginine methyl ester (L-NAME) and NQ-monomethyl L-arginine (L-NMMA) on acetylcholine(ACh)-induced contractions of the rabbit coronary artery and the canine proximal colon. Panels A and B show the effects ofarginine analogues in the rabbit coronary artery denuded of endothelium. Panel A: Tracings showing examples of the contractileresponse obtained on addition of 1 ,iM ACh in the presence and absence of 100 ,M L-NAME. Panel B: Graph showingconcentration-response relation forACh in the absence (0) of arginine analogue or in the presence of 100 ,uM L-NAME (o) or100 ,uML-NMMA (D). Values are mean±SEM ofexperimentsperformed in five to 13 animals. Panels C and D show the effectsof arginine analogues in the canine colon. Panel C: Tracings showing examples of the contractile response obtained on additionof 1 ,uMACh in thepresence and absence of100 ,iML-NAME. Panel D: Graph showing concentration-response relationforAChin the absence (a) of arginine analogue or in the presence of 100 ,uM L-NAME (o) or 100 pM L-NMMA (E). Values aremean±SEM of experiments performed in five to eight animals. *Significantly different from control (p<O.05).
mass action. For saturation binding data, nonlinearcurve fitting was used to generate both the Kd and Bm1,values in preference to the linear transform. Bothmethods yield similar results.13 For competition bindingdata, the nonlinear least-squares approach fits the datato either one or two classes of binding sites and assists indetermining if the two-site model is significantly betterthan the one-site fit of the data (F test). The affinity ofreceptors for L-arginine analogues in competition forradioligand binding (K1) was determined by the methodof Cheng and Prusoff.14 Data are expressed as themean±+SEM. Statistical significance was determined byStudent's t test, where a value ofp<0.05 was consideredsignificant.
DrugsACh, the stable muscarinic agonist carbachol, and the
muscarinic antagonist atropine, as well as GTP ana-logue GTPyS, EGTA, Tris base, and the p-dtative NOSinhibitors L-NAME, L-NMMA, Na-benzoyl L-arginineethyl ester (BAEE), L-AME, L-arginine ethyl ester(L-AEE), and L-NA were purchased from SigmaChemical Co., St. Louis, Mo. [3H]QNB (45.7 Ci/mmol)was obtained from Dupont-NEN, Boston. All othermaterials were obtained from standard sources. Argi-nine analogues (except L-NA) were prepared freshimmediately before use by dissolution in buffer contain-ing (mM) Tris-HCI 5, MgCl2 1, and EGTA 0.1, pH 7.4,with sonication. L-NA was dissolved at a stock concen-
tration of 100 mM in 0.05N NaOH in water andimmediately diluted to the desired concentration usingthe same Tris/MgCl2/EGTA buffer shown above. Solu-tions of L-NAME, L-NMMA, L-AME, and L-AEEwere significantly less effective or ineffective as musca-rinic antagonists if stored frozen at -20°C and reused.
ResultsConcentration-response relations for ACh-induced
contraction in both rabbit coronary artery and caninecolonic smooth muscle were determined by cumulativeaddition of ACh (0.01-100 ,gM). The concentration ofACh that produced half-maximal contraction in coronaryrings (Figure 1B) was 1.5 ,uM (log ED50=-5.81+0.14),whereas in colonic strips 2.1 ,uM ACh (log ED50=-5.67±0.10) was required to produce half-maximal con-traction (Figure 1D).To determine whether the contractile responses to
ACh in coronary artery and colon were modified byarginine analogues, cumulative additions of ACh werealso obtained after preexposure of tissues to eitherL-NAME or L-NMMA. L-NAME (100 pM), but notL-NMMA (100 ,tM), produced a significant shift to theright of the concentration-response relation for ACh inboth coronary artery (log EC50=-4.42+0.31, Figure1B) and colon (log EC50=-5.15±+0.12, Figure 1D).Examples of the effect of 100 ,uM L-NAME on contrac-tions produced by 1 ,tgM ACh in coronary artery andcolon are shown in Figures 1A and 1C, respectively. In
390 Circulation Research Vol 72, No 2 February 1993
CH 0
ACh 3 C
CH3 H3
NH+ 0
ARG H2N-KN OH
H NH3
NH2 0
L-NMMA 3 N N OH
H H NH3
L-NA
L-AME
L-AEE
NH2 0
02N. N N OHH H NH2
NH2 0
H1 NXN 0,1C3H NH2
NH+ 0
H2N N OH3CHH NH2
NH+ 0
L-NAME 02N.N N 3
H H NH2
NH4
BAEE H2N N N,0.'0H
K0
CH3
FIGURE 2. Structures of acetylcholine (ACh), arginine(ARG), andARG analogues used as inhibitors of nitric oxidesynthase. L-NMMA, NG monomethyl L-arginine; L-NA, N0-nitro L-arginine; L-AME, L-argznine methyl ester; L-AEE,L-arginine ethyl ester; L-NAME, NG-nitro L-arginine methylester; BAEE, Na-benzoyl L-arginine ethyl ester.
several experiments cumulative additions of ACh wererepeated once more in the presence of both L-NAMEand L-arginine (1 ,uM). There was no significant differ-ence in the response to ACh plus L-NAME in thepresence or absence of L-arginine in either tissue type(n=3; data not shown).The effect of L-NAME to lower the potency of ACh
in contractile studies taken together with the generalstructural similarity of ACh and the arginine analoguesthat inhibit NOS (Figure 2) suggested to us that thesenitroarginine analogues might bind to muscarinic recep-tors. Since the results in both rabbit coronary artery andcolon suggested that L-NAME was interacting with amuscarinic receptor of the M2 or M3 subtype,10"15 it wasunclear whether the effect of L-NAME might be recep-tor-subtype specific. To test this possibility, we pre-pared membranes from different tissues representative
TABLE 1. Characteristics of Muscarinic Receptors in DifferentTissues: Effects of NG-Nitro-L-Arginine Methyl Ester
Values are mean±SEM of three to five experiments.Specific binding of ['H]quinuclidinyl benzilate (5-5,000 pM) to
membranes prepared from each of the tissues shown was deter-mined by filtration as described in the text. Kd and Bm, values werecalculated from nonlinear fit of the saturation binding data.
of each of the muscarinic receptor subtypes (M1, M2,and M3) well known to be expressed in mammaliantissues.16 Saturation isotherms were performed as wehave described elsewhere'0 to determine the affinity anddensity of muscarinic receptors in several tissues for themuscarinic antagonist radioligand [3H]QNB (Table 1).As demonstrated by the work of Doods et al,17 canineatria were used as a source of M2 receptors, subman-dibular gland was used as a source of M3 receptors, andwhole brain was used as a source of all three receptortypes. Our own data confirm the presence of M2 andM3 receptors in colonic smooth muscle.10 Choices oftissues representing each of the muscarinic receptorsubtypes thought to exist in the cardiovascular systemand elsewhere were made to ensure the general appli-cability of our data. Direct assessment of the effects ofL-NAME was determined both in blood vessels andendothelial cells.
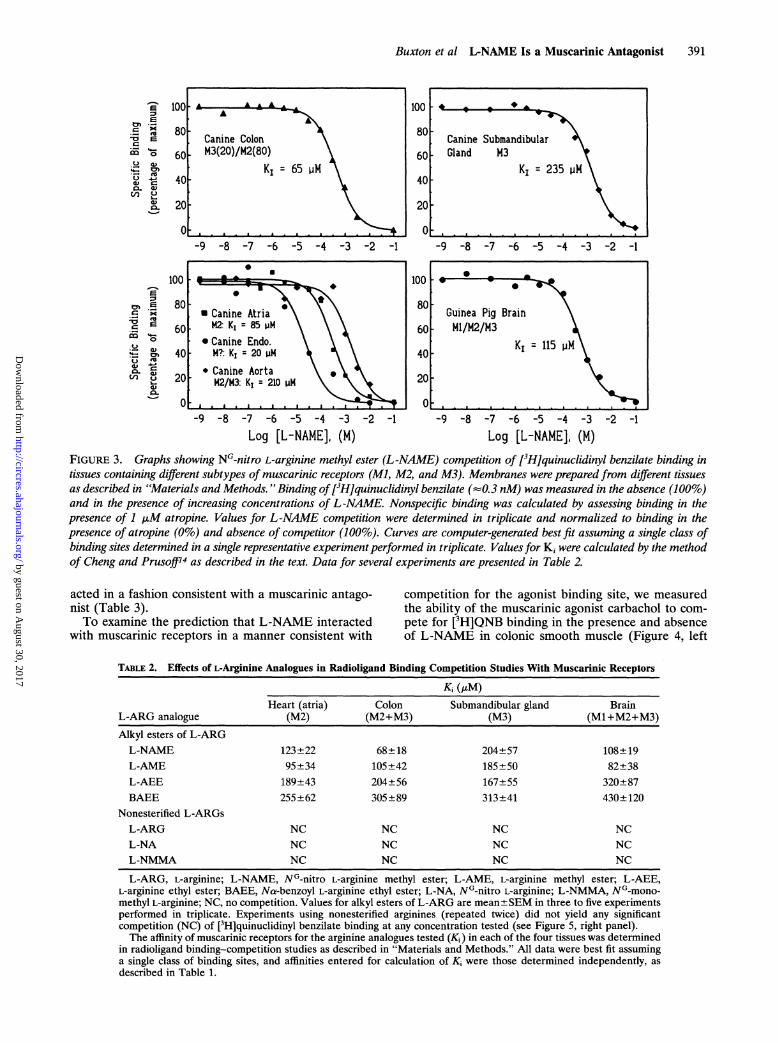
Addition of increasing concentrations of L-NAME inequilibrium binding-competition experiments usingmembranes from each of the five tissue sources and thenonspecific radioligand [3H]QNB (0.3 nM) resulted in acomplete monophasic competition of binding in eachcase, with affinities of the receptor for L-NAME thatvaried from 65 ,uM in colonic smooth muscle to 235 gMin canine submandibular gland (Figure 3, Table 2). Inwhole canine aorta, competition of [3H]QNB binding byL-NAME (Figure 3) resulted in a mean inhibitoryaffinity (K,) of 317±+68 ,uM (n=6), whereas in endothe-lium scraped from aorta, L-NAME competition of[3H]QNB binding yielded a Ki of 86+51 ,uM (n=3).Similar experiments undertaken in canine coronaryarteries (n=17) also demonstrated that L-NAME com-peted for specific [3H]QNB binding, although the bind-ing in the absence of L-NAME was unimpressive (11.4fmol/mg at 450 pM [3H]QNB). The affinity of musca-rinic receptors for L-NAME determined in severaltissues agrees with the ability of L-NAME to interferewith agonist-induced contraction when added to thecontractile experiments at a concentration of 100 ,uM(Table 2). Addition of the hydrolysis-resistant guaninenucleotide GTPyS (100 uM) did not shift the competi-tion curve for L-NAME, suggesting that L-NAME
Log [L-NAME], (M) Log [L-NAME], (M)FIGURE 3. Graphs showing NG-nitro L-arginine methyl ester (L-NAME) competition of [3H]quinuclidinyl benzilate binding intissues containing different subtypes of muscarinic receptors (M], M2, and M3). Membranes were prepared from different tissuesas described in 'Materials and Methods. " Binding of[3H]quinuclidinyl benzilate (--0.3 nM) was measured in the absence (100%)and in the presence of increasing concentrations of L -NAME. Nonspecific binding was calculated by assessing binding in thepresence of 1 ttM atropine. Values for L-NAME competition were determined in triplicate and normalized to binding in thepresence of atropine (0%) and absence of competitor (100%). Curves are computer-generated best fit assuming a single class ofbinding sites determined in a single representative experiment performed in triplicate. Values for K' were calculated by the methodof Cheng and Prusoffi4 as described in the text. Data for several experiments are presented in Table 2
acted in a fashion consistent with a muscarinic antago-nist (Table 3).To examine the prediction that L-NAME interacted
with muscarinic receptors in a manner consistent with
competition for the agonist binding site, we measuredthe ability of the muscarinic agonist carbachol to com-pete for [3H]QNB binding in the presence and absenceof L-NAME in colonic smooth muscle (Figure 4, left
TABLE 2. Effects of L-Arginine Analogues in Radioligand Binding Competition Studies With Muscarinic Receptors
Ki (/M)Heart (atria) Colon Submandibular gland Brain
L-ARG, L-arginine; L-NAME, NG-nitro L-arginine methyl ester; L-AME, L-arginine methyl ester; L-AEE,L-arginine ethyl ester; BAEE, Na-benzoyl L-arginine ethyl ester; L-NA, NG-nitro L-arginine; L-NMMA, NG-mono-methyl L-arginine; NC, no competition. Values for alkyl esters of L-ARG are mean±SEM in three to five experimentsperformed in triplicate. Experiments using nonesterified arginines (repeated twice) did not yield any significantcompetition (NC) of [3H]quinuclidinyl benzilate binding at any concentration tested (see Figure 5, right panel).The affinity of muscarinic receptors for the arginine analogues tested (Ki) in each of the four tissues was determined
in radioligand binding-competition studies as described in "Materials and Methods." All data were best fit assuminga single class of binding sites, and affinities entered for calculation of Ki were those determined independently, asdescribed in Table 1.
392 Circulation Research Vol 72, No 2 February 1993
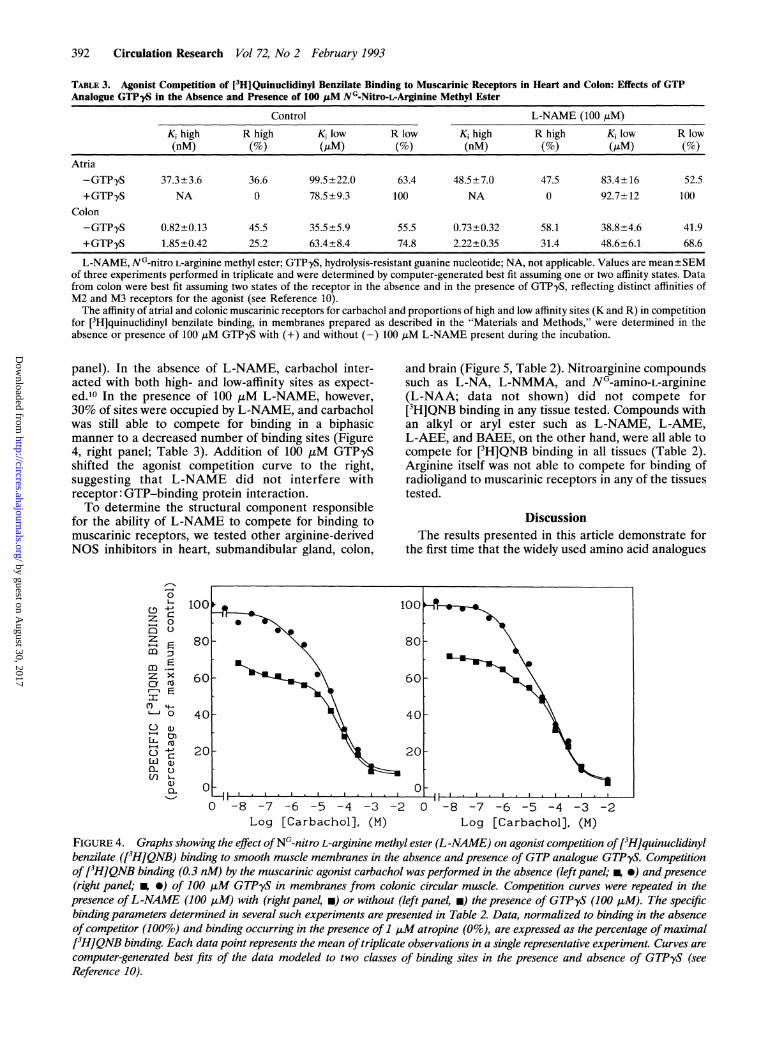
TABLE 3. Agonist Competition of [3H]Quinuclidinyl Benzilate Binding to Muscarinic Receptors in Heart and Colon: Effects of GTPAnalogue GTPyS in the Absence and Presence of 100 ,uM NG-Nitro->LArginine Methyl Ester
Control L-NAME (100 ,uM)
Ki high R high Ki low R low K, high R high K, low R low(nM) (o M)6 (nM)(S)(M
Atria-GTPyS 37.3+3.6 36.6 99.5±22.0 63.4 48.5±7.0 47.5 83.4±16 52.5+GTPyS NA 0 78.5±9.3 100 NA 0 92.7±12 100
L-NAME, NG-nitro L-arginine methyl ester; GTPyS, hydrolysis-resistant guanine nucleotide; NA, not applicable. Values are mean±SEMof three experiments performed in triplicate and were determined by computer-generated best fit assuming one or two affinity states. Datafrom colon were best fit assuming two states of the receptor in the absence and in the presence of GTPyS, reflecting distinct affinities ofM2 and M3 receptors for the agonist (see Reference 10).The affinity of atrial and colonic muscarinic receptors for carbachol and proportions of high and low affinity sites (K and R) in competition
for ['H]quinuclidinyl benzilate binding, in membranes prepared as described in the "Materials and Methods," were determined in theabsence or presence of 100 ,uM GTPyS with (+) and without (-) 100 juM L-NAME present during the incubation.
panel). In the absence of L-NAME, carbachol inter-acted with both high- and low-affinity sites as expect-ed.'0 In the presence of 100 ,uM L-NAME, however,30% of sites were occupied by L-NAME, and carbacholwas still able to compete for binding in a biphasicmanner to a decreased number of binding sites (Figure4, right panel; Table 3). Addition of 100 ,uM GTPySshifted the agonist competition curve to the right,suggesting that L-NAME did not interfere withreceptor: GTP-binding protein interaction.To determine the structural component responsible
for the ability of L-NAME to compete for binding tomuscarinic receptors, we tested other arginine-derivedNOS inhibitors in heart, submandibular gland, colon,
and brain (Figure 5, Table 2). Nitroarginine compoundssuch as L-NA, L-NMMA, and N0-amino-L-arginine(L-NAA; data not shown) did not compete for[3H]QNB binding in any tissue tested. Compounds withan alkyl or aryl ester such as L-NAME, L-AME,L-AEE, and BAEE, on the other hand, were all able tocompete for [3H]QNB binding in all tissues (Table 2).Arginine itself was not able to compete for binding ofradioligand to muscarinic receptors in any of the tissuestested.
DiscussionThe results presented in this article demonstrate for
the first time that the widely used amino acid analogues
FIGURE 4. Graphs showing the effect ofNG-nitro L-arginine methyl ester (L-NAME) on agonist competition of[3H]quinuclidinylbenzilate ([3H]QNB) binding to smooth muscle membranes in the absence and presence of GTP analogue GTPyS. Competitionof[3H]QNB binding (0.3 nM) by the muscarinic agonist carbachol was performed in the absence (left panel; ig 0) and presence(right panel; g *) of 100 ,uM GTPyS in membranes from colonic circular muscle. Competition curves were repeated in thepresence ofL-NAME (100 ,M) with (right panel, m) or without (left panel, m) the presence of GTPyS (100 LM). The specificbindingparameters determined in several such experiments are presented in Table 2. Data, normalized to binding in the absenceof competitor (100%) and binding occurring in the presence of 1 ,uM atropine (0%), are expressed as the percentage ofmaximal[3H]QNB binding. Each data point represents the mean of triplicate observations in a single representative experiment. Curves arecomputer-generated best fits of the data modeled to two classes of binding sites in the presence and absence of GTPyS (seeReference 10).
Buxton et al L-NAME Is a Muscarinic Antagonist 393
-7 -6 -5 -4 -3 -2 -7 -6 -5 -4 -3 -2
Log [arginine analog], (M)FIGURE 5. Graphs showing the effects of nitric oxide synthase inhibitors on competition of [3H]quinuclidinyl benzilate([3H]QNB) binding in smooth muscle membranes. L-NAME, NG-nitro L-arginine methyl ester; L-AME, L-arginine methyl ester;L-AEE, L-arginine ethyl ester; BAEE, Na-benzoyl L-arginine ethyl ester; L-ARG, L-arginine; L-NA, NG-nitro L-arginine;L-NMMA, N0-monomethyl L-arginine. Leftpanel: Arginine analogues possessing an alkyl ester modification at the carboxyl endof the amino acid were used in competition for [3H]QNB binding in membranes prepared from colonic smooth muscle. Data,normalized to binding in the absence of competitor (100%) and binding occurring in the presence of 1 ,uM atropine (0%), are
expressed as the percentage ofmaximal [3H]QNB binding (0.3 nM). Curves are computer-generated best fit assuming the presenceof a single class of binding sites. Data are the mean of triplicate observations from three representative experiments. Right panel:L-ARG and L-ARG analogues without an alkyl ester modification at the carboxyl end of the amino acid were used in competitionfor [3H]QNB binding in colonic smooth muscle membranes and found to be without effect. The results of additional experimentsare presented in Table 2.
of arginine with alkyl or aryl esterification of thecarboxyl group are antagonists of muscarinic receptors.This conclusion is based on observations in intactsegments of vascular and visceral smooth muscle as wellas the results of radioligand binding studies with each ofthe muscarinic receptors commonly expressed in mam-malian cardiovascular and other tissues.The results of experiments with L-NAME and
L-NMMA in the rabbit coronary artery revealed thatL-NAME significantly reduced the amplitude of ACh-induced contractions but L-NMMA did not. The inhi-bition of contraction by L-NAME was not reversed withL-arginine. These observations support the hypothesisthat L-NAME antagonizes muscarinic receptors.NO may be spontaneously generated from several
different sources in isolated segments of canine colon.These sources include enteric neurons, vascular endo-thelial cells, and possibly an inducible form of NOS insmooth muscle cells.4'18 Arginine analogues wouldtherefore be predicted to have an excitatory effect on
the response to ACh in the canine colon because of theremoval of an inhibitory substance (i.e., NO). In con-
trast to this predicted effect of arginine analogues, we
observed instead that the contractile response to AChwas reduced in the presence of 100 gM L-NAME andwas unchanged in the presence of 100 gM L-NMMA.The inhibition of ACh-induced contractions in thepresence of L-NAME is compatible with the hypothesisthat this analogue antagonizes muscarinic receptors,whereas the lack of an effect of L-NMMA could be
interpreted in a number of different ways. To providedirect evidence for the effect of these analogues on
muscarinic receptors, we therefore undertook a series ofradioligand binding studies.Our radioligand binding studies demonstrate that
each of the alkyl esters of L-arginine tested, L-NAME,L-AME, and L-BAEE, were effective antagonists ofmuscarinic receptors regardless of the complement ofreceptor subtypes present in the tissue tested. In con-
trast, the nitroarginine compounds without altered car-
boxyl groups, L-NA, L-NMMA, and L-NAA, did notcompete for [3H]QNB binding in any tissue tested.These results correlate well with the L-NAME-inducedreduction in ACh-induced contractions observed inboth the rabbit coronary artery and the canine colon, an
effect not produced by L-NMMA.The notion that the alkyl esters of L-arginine compete
for muscarinic receptor binding as antagonists acting atthe known binding site for agonist is supported by our
data showing that 1) guanine nucleotide does not alterthe ability of alkyl esters of L-arginine to compete for[3H]QNB binding and 2) these esters compete foragonist binding directly and reversibly, without interfer-ing with GTP-dependent agonist binding. The data ofTable 3 demonstrate that in atria, where all M2 musca-
rinic receptors are shifted to low affinity in the presenceof GTPyS, addition of L-NAME does not alter thiseffect. Furthermore, L-NAME does not alter the ago-nist competition in canine colon, in which it is known10that addition of GTPyS shifts the agonist binding com-
394 Circulation Research Vol 72, No 2 February 1993
petition curve to a low-affinity curve best fit assumingtwo agonist:receptor complexes (M2 and M3). Theassignment of L-NAME and the other alkyl esters ofL-arginine as nonselective muscarinic antagonists alsofollows from our data. In competition studies usingtissues known to possess different forms of the musca-rinic receptor, the competition curve is monophasic. Ifthe alkyl esters of L-arginine were selective for one formof the muscarinic receptor, one might expect that curveswould be shallow and might be best fit assuming thepresence of more than one receptor:antagonist com-plex. This was not the case. The simplest conclusionfrom these studies then is that the alkyl esters ofL-arginine compete at the agonist binding site of severalforms of the muscarinic receptor nonselectively andwith moderate affinity, a proposal that is supported bythe similarity of the structure of these compounds whencompared against ACh (Figure 2).The effects of the alkyl esters L-NAME, L-AME, and
L-BAEE on muscarinic receptors have important impli-cations in the interpretation of a variety of experiments.ACh was the first agonist shown to release a substancefrom blood vessels that was later identified as NO,19 andit is still frequently used as a method for releasing NOfrom endothelial cells (for review see Reference 20).Our results clearly suggest that a portion of the inhibi-tory effects of L-NAME and related analogues onACh-induced relaxation could be due to antagonism atthe muscarinic receptor. We provide functional andradioligand binding evidence in blood vessels, suggest-ing that the addition of L-NAME and related com-pounds will block both smooth muscle and endothelialcell receptors. Direct measurement of muscarinic recep-tors in blood vessels and their subtype assignment isdifficult because of the small numbers of receptorspresent.2'-23 Indeed, muscarinic receptors on endothe-lial cells were determined by Tracey and Peach24 to belost during isolation and culture, but although theseauthors were able to determine that freshly preparedendothelial cells possess mRNAs for each of the com-monly expressed muscarinic receptor subtypes (ml, m2,and m3), they could not determine which were trans-lated. Thus, conclusions as to the muscarinic receptorsubtype(s) present on the endothelial cell must be madewith caution. Functional studies have suggested thatendothelial cells probably possess both M2 and M3receptors.2526 Although the muscarinic receptor com-plement on endothelium is not clear, we provide evi-dence that L-NAME is an antagonist at these as well asother muscarinic receptor subtypes.
In studies in which cholinergic excitatory nerves arestimulated to release ACh, alkyl esters of arginine mayalso produce complicated effects on the response, in-volving both the excitatory effect of removing NO that isdue to NOS inhibition and the inhibitory effect ofantagonizing muscarinic receptors. The overall changein contractile amplitude obtained on addition ofL-NAME to a tissue is therefore likely to differ depend-ing on how the two opposing effects sum together in aparticular tissue. Indeed, it was our initial observationthat the nerve-stimulated cholinergic contraction incanine colon was sometimes reduced on addition ofL-NAME, and it was this observation that initially ledus to explore the hypothesis that arginine analoguesmight also antagonize muscarinic receptors.
The ability of alkyl esters of arginine to antagonizemuscarinic receptors also has important implications ininterpreting in vivo studies. For example, in thesestudies various physiological parameters have beenmonitored during exposure to L-NAME.2728 If theintroduced analogue not only antagonizes NOS but alsoinhibits muscarinic receptors, then the physiologicaleffects observed are likely to be the sum of these twoactions. A key observation in determining whether theeffects of L-NAME are due to antagonism of NOS or toantagonism of muscarinic receptors is the ability ofL-arginine to reverse the effects of L-NAME. We havefound that the antagonism of muscarinic receptors byL-NAME cannot be overcome by L-arginine, whereasreversal of NOS antagonism by L-arginine has beenwidely reported and recently reviewed.2We suggest that the results presented here, that alkyl
and aryl esters of L-arginine are muscarinic antagonists,can be extended from the specific tissues in which theywere obtained (rabbit coronary artery, canine heart,coronary artery, aorta, endothelium, colon, submandib-ular gland, and guinea pig brain) to any tissue contain-ing Ml, M2, or M3 receptors. Furthermore, studiesusing these amino acid analogues should take intoaccount the possibility that their actions are, at least inpart, due to antagonism of muscarinic receptors.
pathophysiology, and pharmacology. Pharmacol Rev 1991;43:109-142
3. Palmer RMJ, Ferrige AG, Moncada S: Nitric oxide releaseaccounts for the biological activity of endothelium-derived relaxingfactor. Nature 1987;327:524-526
4. Ward SM, Dalziel HH, Thornbury KD, Westfall DP, Sanders KM:Nonadrenergic, noncholinergic inhibition and rebound excitationin canine colon depend on nitric oxide. Am J Physiol 1992;262(Gastrointest Liver Physiol):G237-G243
5. Rees DD, Palmer RMJ, Schulz R, Hodson HF, Moncada S: Char-acterization of three inhibitors of endothelial nitric oxide synthasein vitro and in vivo. Br J Pharmacol 1990;101:746-752
6. Rees DD, Schulz R, Hodson HF, Palmer RMJ, Moncada S: Iden-tification of some novel inhibitors of the vascular nitric oxidesynthase in vivo and in vitro, in Moncada S, Higgs EA (eds): NitricOxide from L-Arginine: A Bioregulatory System. Amsterdam, Else-vier Science Publishing Co, Inc, 1990, pp 485-487
7. Smith TK, Reed JB, Sanders KM: Origin and propagation ofelectrical slow waves in circular muscle of canine proximal colon.Am J Physiol 1987;252(Cell Physiol 21):C215-C224
8. Durant GJ, Ganellin CR, Parsons ME: Chemical differentiation ofhistamine H1- and H2-receptor agonists.JMed Chem 1975;18:905-909
9. Keef KD, Pasco JS, Eckman DM: Purinergic relaxation and hyper-polarization in guinea pig and rabbit coronary artery: Role of theendothelium. J Pharmacol Exp Ther 1992;260:592-600
10. Zhang L, Horowitz B, Buxton ILO: Muscarinic receptors in caninecolonic circular smooth muscle: I. Coexistence of M2 and M3 sub-types. Mol Pharmacol 1991;40:943-951
11. Bradford MM: A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle of pro-tein-dye binding. Anal Biochem 1976;72:248-254
12. Bruns RF, Lawson-Wendling K, Pugsley TA: A rapid filtrationassay for soluble receptors using polyethylenimine-treated filters.Anal Biochem 1983;132:74-78
13. Schiemann WP, Walther JM, Buxton ILO: On the ability of endog-enous adenosine to regulate purine nucleoside receptor binding ofantagonists in smooth muscle membranes. J Pharmacol Exp Ther1990;255:886-892
14. Cheng Y-C, Prusoff WH: Relationship between the inhibition con-stant (K1) and the concentration of inhibitor which causes 50 percent inhibition (I50) of an enzymatic reaction. Biochem Pharmacol1973;22:3099-3108
17. Doods HN, Mathy M, Davidesko D, van Charldorp KJ, de JongeA, van Zwieten PA: Selectivity of muscarinic antagonists in radio-ligand and in vivo experiments for the putative Ml, M2, and M3receptors. J Pharmacol Exp Ther 1987;242:257-262
18. Rees DD, Cellek S, Palmer RMJ, Moncada S: Dexamethasoneprevents the induction by endotoxin of a nitric oxide synthase andthe associated effects on vascular tone: An insight into endotoxinshock. Biochem Biophys Res Commun 1990;173:541-547
19. Satchell D: Use of purine nucleotide and nucleoside metabolisingenzymes as tools to determine the presence of purinergic nerve
transmission in smooth muscle. Adv Exp Med Biol 1990;253B:435-440
26. Brunner F, Ktihberger E, Groschner K, Poch G, Kukovetz WR:Characterization of muscarinic receptors mediating endothelium-dependent relaxation of bovine coronary artery. Eur J Pharmacol1991;200:25-33
27. Persson MG, Gustafsson LE, Wiklund NP, Moncada S, HedqvistP: Endogenous nitric oxide as a probable modulator of pulmonarycirculation and hypoxic pressor response in vivo. Acta PhysiolScand 1990;140:449-457
28. Gardiner SM, Compton AM, Kemp PA, Bennett T: Effects ofN0-nitro-L-arginine methyl ester or indomethacin on differentialregional and cardiac haemodynamic actions of arginine vasopressinand lysine vasopressin in conscious rats. Br J Pharmacol 1991;102:65-72
http://circres.ahajournals.org/content/72/2/387World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions: