Journal of Experimental Marine Biology and Ecology 184 (1994) 143-158 JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY Gametogenesis in deep- and surface-dwelling oceanic stalked barnacles from the NE Atlantic Ocean A. Green a, P. A. Tyler +*, M. V. Angel b, J. D. Gage” “Department of Oceanography, University of Southampton, Southampton SO9 5NH, UK bInstitute of Oceanographic Sciences, Deacon Laboratory, Wormley. Godalming, Surrey GU8 5VB, UK ‘Scottish Association for Marine Science, P.O. Box 3, Oban, Argyll PA34 4AD. UK Received 28 March 1994; revision received 8 July 1994; accepted 28 July 1994 Abstract The gametogenic development of the lower bathyal stalked barnacle, Poecilusma kaempferi Darwin, found attached to the exoskeleton of the large lithodid crab Neolithodes grimaldi, and the surface-living oceanic stalked barnacle Lepas anatiferu L., living on an Argos buoy trapped in a mesoscale eddy in the NE Atlantic, has been examined. In P. kuempferi initiation of both oogenesis and spermatogenesis occurs at about -day 70; the first ripe testis is coincident with penis development at -day 145 and the first vitellogenic oocytes at -day 155; egg lamellae were observed in individuals - 170 days old. In L. anatifera testis development started at -day 43 and male organs were mature by - day 13 1; oocytes were first observed in individuals estimated to be - 26 days old and large oocytes were first found in animals after - 114 days; no egg lamellae were found in the oldest animals examined. These rates are slower than previously observed in L. anatiferu but indicate that this species will undergo gametogenesis outside its normal geo- graphic range although the temperature may be too low for successful completion of reproduc- tion. Keywords: Deep-sea; Gametogenesis; Reproduction; Stalked barnacle 1. Introduction In the perceived constancy in the physico-chemical environment of the deep-sea, reproduction in deep-sea invertebrates was predicted as occurring quasi-continuously throughout the year (Orton, 1920). As a result of time-series sampling at selected stations in deep-sea environments predictable seasonal reproduction is now known to * Corresponding author. 0022-0981/94/$7.00 0 1994 Elsevier Science B.V. All rights reserved SSDI 0022-098 l(94)OO 116-2
Transcript
Journal of Experimental Marine Biology and Ecology 184 (1994) 143-158
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
Gametogenesis in deep- and surface-dwelling oceanic stalked barnacles from the NE Atlantic Ocean
A. Green a, P. A. Tyler +*, M. V. Angel b, J. D. Gage”
“Department of Oceanography, University of Southampton, Southampton SO9 5NH, UK bInstitute of Oceanographic Sciences, Deacon Laboratory, Wormley. Godalming, Surrey GU8 5VB, UK
‘Scottish Association for Marine Science, P.O. Box 3, Oban, Argyll PA34 4AD. UK
Received 28 March 1994; revision received 8 July 1994; accepted 28 July 1994
Abstract
The gametogenic development of the lower bathyal stalked barnacle, Poecilusma kaempferi
Darwin, found attached to the exoskeleton of the large lithodid crab Neolithodes grimaldi, and the surface-living oceanic stalked barnacle Lepas anatiferu L., living on an Argos buoy trapped in a mesoscale eddy in the NE Atlantic, has been examined. In P. kuempferi initiation of both oogenesis and spermatogenesis occurs at about -day 70; the first ripe testis is coincident with penis development at -day 145 and the first vitellogenic oocytes at -day 155; egg lamellae were observed in individuals - 170 days old. In L. anatifera testis development started at -day 43 and male organs were mature by - day 13 1; oocytes were first observed in individuals estimated to be - 26 days old and large oocytes were first found in animals after - 114 days; no egg lamellae were found in the oldest animals examined. These rates are slower than previously observed in L. anatiferu but indicate that this species will undergo gametogenesis outside its normal geo- graphic range although the temperature may be too low for successful completion of reproduc- tion.
In the perceived constancy in the physico-chemical environment of the deep-sea, reproduction in deep-sea invertebrates was predicted as occurring quasi-continuously throughout the year (Orton, 1920). As a result of time-series sampling at selected stations in deep-sea environments predictable seasonal reproduction is now known to
* Corresponding author.
0022-0981/94/$7.00 0 1994 Elsevier Science B.V. All rights reserved
SSDI 0022-098 l(94)OO 116-2
144 A. Green et al. /J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158
occw in some bathyal and abyssal species (see Gage & Tyler, 1991). In addition to these two reproductive patterns there is very limited evidence that, in a few species, repro- duction may occur very rapidly when a suitable substratum and/or a food source becomes available (Turner, 1973; Tyler et al., 1992).
Most deep-sea invertebrates, with the exception of those species found at hydro- thermal vents, (see Tunnicliffe, 1991) were believed to grow slowly and have long life spans (Turekian et al., 1975). Micro-, meio-, and small macrofaunal species in the deep-sea are known to grow and reproduce rapidly (Gooday, 1988, Lochte & Turley, 1988; Scheltema, 1987) whilst Gage (1990) has shown that some deep-sea megafaunal species, notabiy echinoderms, can grow to adult size in a relatively short time (N 2- 3 yr) and have a life span of - 10 s of years. More dramatically, Lampitt (1990) demonstrated that the deep-sea stalked barnacle Poecilasma kaempferi can grow to adult size in less than 228 days at an ambient temperature of 3 “C.
In surface waters many species of stalked barnacles, particularly from the tropics, are known for their rapid growth (Darwin, 1851; Evans, 1958; MacIntyre, 1966) al- though temperate species grow more slowly and have delayed reproduction (Batham, 1945: Barnes & Reese, 1960). Little is known, however, of the rate at which gameto- genesis occurs. In shallow water the pattern and rate of reproduction in balanomorph barnacles has been described and debated (see Barnes 1989, 1992 for reviews) but reproduction in lepadomorph barnacles is less well described (Batham, 1945; Evans, 1958; Patel, 1959; Barnes & Reese, 1960; Hilgard, 1960; Zann & Harker, 1978; Lewis & Chia, 1981; Cimberg, 1981; Eckert & Eckert, 1987; Barnes, 1989, 1992).
Samples of the bathyal stalked barnacle P. kaempferi were collected from the cara- pace of the deep-sea stone crab ~eo~jt~odesgrim~~dj. In addition, the tropic~/subtropical surface-dwelling stalked barnacle, Lepas an~t~r~, which colonised a surface buoy fitted with an Argos transmitter and drogued at a depth of 70 m has been investigated. The buoy was deployed on 8th April 1989 and retrieved on 13 May 1990 during RRS “Discovery” cruise 191. Pingree and Le Cann (1991) have described the track of the buoy during a year of its deployment during which time it spent 66 days in a cyclonic eddy and 130 days in an anticyclonic eddy undergoing 21 rotations around the eddy centre. For the remaining period the buoy was not in an eddy. Throughout its whole deployment the buoy moved at a rate of 1.4 cm.s -i to the northwest, and remained close to 51.5” N: 22” W for most of that time. The sea surface temperature (SST) measured at I m throughout the year showed a strong seasonal signal, and a strong high-frequency signal in summer as a result of diurnal heating (see Fig. 2 in Pingree & Le Cann, 1991).
Poecilusma kaempferi was believed to colonise, exclusively, natural substrata such as the exoskeletons of deep-sea crabs (Williams & Moyse, 1988) and to have a cosmo- politan distribution. The observations of Lampitt (1990) were the first records of P. k~empfe~ occurring on an artificial substratum. Lepas anfft~ra is found colonizing a variety of natural and anthropogenic material in the tropical/subtropic~ surface waters of the world ocean (Tsikhon-Lukanina et al. 1976; Zann & Harker, 1978).
By using the size/age relationship for P. kaempferi described by Lampitt (1990) and the known deployment time of the buoy for L. anatifera, an attempt has been made to determine the the timing of the main reproductive events in these two species.
A. Green et al. /J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158 145
2. Materials and methods
P~eci~~~~a k~e~pferi was taken from the carapaces of the stone crab iV. ~~~~l~i collected by Semi-Balloon Otter Trawl (Merrett & Marshall, 1981) at 2000 m depth in the Rockall Trough, NE Atlantic Ocean. All samples were fixed in 10% seawater formalin and transferred to 70% alcohol for long-term storage.
Lepas anatifera was collected from an Argos buoy deployed in the NE Atlantic in the region of the East Thulean Rise (N 5 1.5 “N: 22” W) and had spent part of the 320 day deployment circulating in two mesoscale eddies. Both eddies although rotating remained in the same geographical position throughout the deployment. The body of the float had been treated with antifouling paint and the settlement of Lepas was therefore restricted to the bottom of the float, to small areas where the paint had been scratched, and to patches of “Velcro” on the side of the buoy. The largest barnacles were on the bottom of the buoy at a depth of 1 m. A few cyprids were found amongst the clumps, so further settlement was taking place at the time of recovery. All the undamaged Lepas (n = 693) were removed and preserved in 5% seawater formalin. On board ship the carina length of all the animals was measured to the nearest mm using a Wild M5 stereomicroscope fitted with scaled eyepiece.
In the faboratory the scutuml tergum and carina length of 50 individuals of each species of different sizes were measured with vernier calipers. Capitulum length is the traditional independent size variable used in population studies of stalked barnacles although other measurements have been used (Evans, 1958; Hilgard, 1960). In larger specimens, particularly of L. anatiferu there was variation in capitular length as a re- sult of fixation artefacts in the tissue connecting the scutum and tergum. As a result it was decided that a more rigorous independent variable was scutum length and this measurement has been used in this study. The relationship between scutum and capitular length is:
Capitulum length = 0.39 + 1.079 x Scutum length for P. kaempferi
and
Capitulum length = 2.59 + 1.206 x Scutum length for L. anat$m
Each individual (soft parts, shell and stalk) was wet-weighed. The shell was removed from each individual to expose the internal organs. Penis length (preserved condition) was measured under a stereomicroscope using a calibrated eye-piece graticule. The body was cut into symmetrical halves to expose the testis. In P. k~e~pfe~ the ovary was very small and the ovary and peduncle were processed intact. In L. unatifera the peduncle was removed and cut into symmetrical halves to expose the ovary which was removed easily.
Testis and ovary were processed through graded alcohols, cleared in histoclear and embedded in paraffin wax. Sections were cut at 7 pm and stained with Masson’s trichrome. The stages of testis development were based on the following criteria. Stage 1: no testis found; Stage 2: spermatogonia and spermatocytes only; Stage 3: sperma- tids present; Stage 4: spermatozoa present but sperm tails make up < 50% of cross
146 A. Green et al. /_I. Exp. Mar. Biol. Ecol. 184 (1994) 143-158
section of testis; Stage 5: testis composed almost exclusively of spermatozoa. Inter- mediate stages could be recognised and were scored as half stages. Two fields of each testis were scored and a mean stage calculated. Oocytes were measured using a dig- itising tablet and Sigmascan software. Twenty individuals of different sizes, of both species, were examined for the presence of egg lamellae.
3. Results
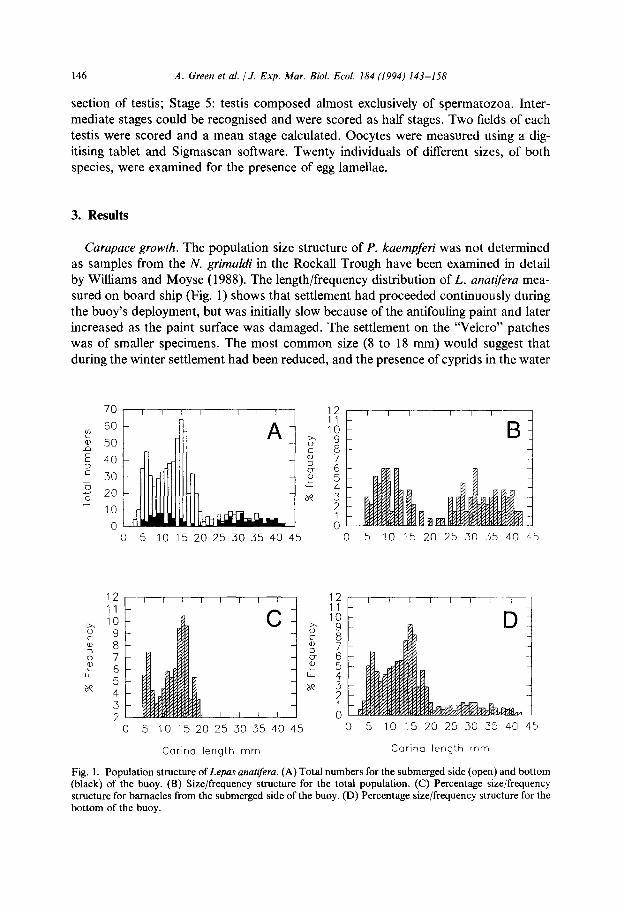
Carapace growth. The population size structure of P. kaempferi was not determined as samples from the N. grimafdi in the Rockall Trough have been examined in detail by Williams and Moyse (1988). The length/frequency distribution of L. anatifra mea- sured on board ship (Fig. 1) shows that settlement had proceeded continuously during the buoy’s deployment, but was initially slow because of the antifouling paint and later increased as the paint surface was damaged. The settlement on the “Velcro” patches was of smaller specimens. The most common size (8 to 18 mm) would suggest that
during the winter settlement had been reduced, and the presence of cyprids in the water
Fig. 1. Population structure of Lepas anatifera. (A) Total numbers for the submerged side (open) and bottom (black) of the buoy. (B) Size/frequency structure for the total population. (C) Percentage size/frequency structure for barnacles from the submerged side of the buoy. (D) Percentage size/frequency structure for the bottom of the buoy.
A. Green et ui.,l.?. Exp..Mar. Bid. Ed 18411994j 143-158 147
column indicated that the warming of the surface waters and the onset of the spring bloom [which was at a peak at the time (Savidge et al. 1992)J had stimulated the ini-
tiation of further settlement. The possibility cannot be ruled out that growth varied with position on the buoy and this has smoothed the length/frequency distribution.
The five plates (2 scuta, 2 terga and a carina) that form the carapace showed iso- metric growth in both species. All regression values were significant (Fig. 2). Compari- son of the length of the scutum with wet weight (Fig. 3) showed an inflexion. Linear
regressions were fitted to the data until an optimal tit was achieved. These regressions
Salurn Icngth (mnj SC”:“;” lenq:h (ml)
2 3 4 5 6 7 8 C 5 10 15 20 25
Tergum length (mm) Tergum length (mm)
a / 7 ,/=
'? 2"
5" _"
4
E 3
$2
2 1 1
0 C, 2 4 6 8 10
10 15 20 25 3c
Conno length (mm) iGlrirn lengtn (mm)
Fig. 2. Relationship between the plates ofthe capitulum of (A)to (C) P. kam~fki and (D) to (F). L. amrifEra. Fitted linear regressions significant p= eO.001.
148 A. Green et al. / J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158
Scutum length mm
0 0 5 10 15 20 25 30
Scutum length mm
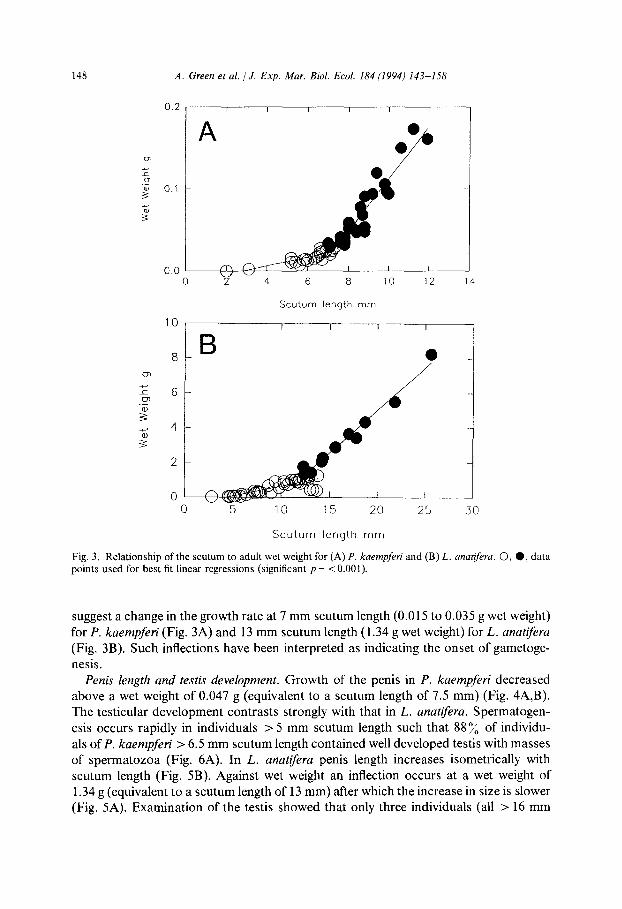
Fig. 3. Relationship of the scutum to adult wet weight for (A) P. kaempferi and (B) L. anarifera. 0, 0, data points used for best fit linear regressions (significant p = < 0.00 1).
suggest a change in the growth rate at 7 mm scutum length (0.015 to 0.035 g wet weight) for P. kaempferi (Fig. 3A) and 13 mm scutum length (1.34 g wet weight) for L. anatiferu
(Fig. 3B). Such inflections have been interpreted as indicating the onset of gametoge-
nesis. Penis length and testis development. Growth of the penis in P. kaempferi decreased
above a wet weight of 0.047 g (equivalent to a scutum length of 7.5 mm) (Fig. 4A,B). The testicular development contrasts strongly with that in L. anatifera. Spermatogen-
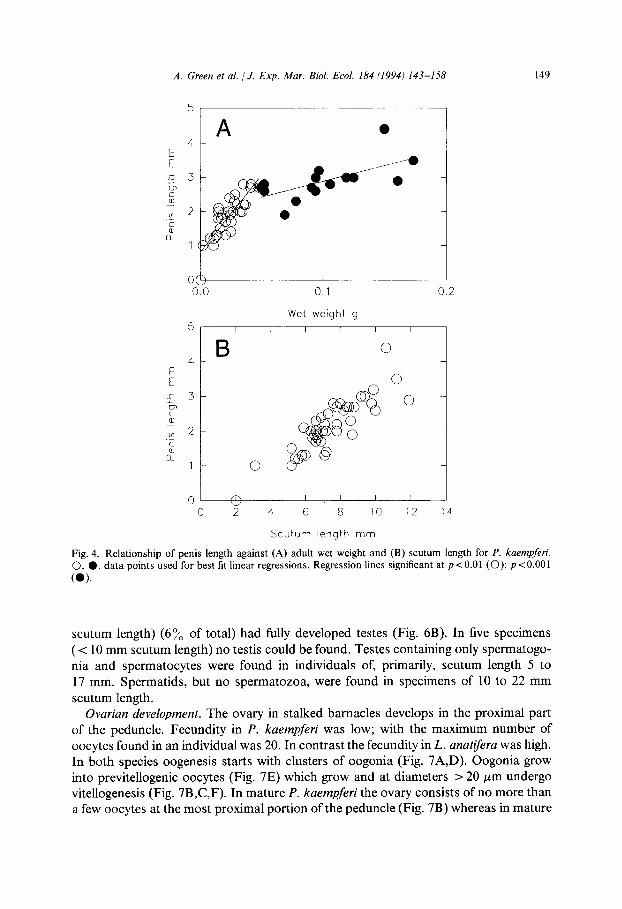
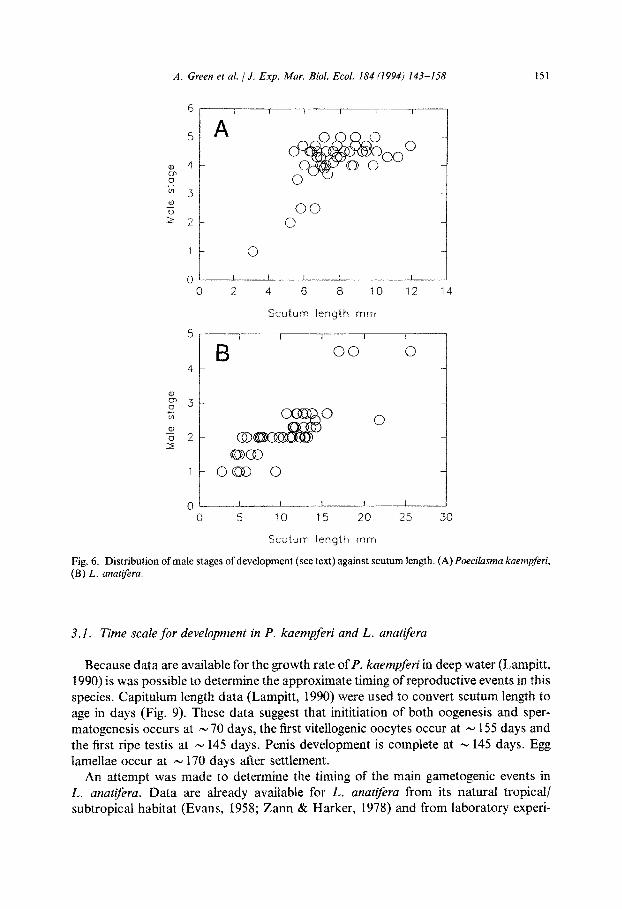
esis occurs rapidly in individuals > 5 mm scutum length such that 88% of individu- als of P. kaempferi > 6.5 mm scutum length contained well developed testis with masses of spermatozoa (Fig. 6A). In L. anatiferu penis length increases isometrically with scutum length (Fig. 5B). Against wet weight an inflection occurs at a wet weight of 1.34 g (equivalent to a scutum length of 13 mm) after which the increase in size is slower (Fig. 5A). Examination of the testis showed that only three individuals (all > 16 mm
A. Green et al. 1 J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158 149
5
4 E E
f. 3
F 2 .‘o 2
6 IL
1
0 0.0 0.1 0.2
Wet weight g
57. ’ I I I
B 0 4- -
f 0
Gj” 0
a ~ 2-
El a
1 - 0
0’ /i\ I I I I / I 0 2 4 6 8 10 12 14
Scutum length mm
Fig. 4. Relationship of penis length against (A) adult wet weight and (B) scutum length for P. kaempferi. 0, 0, data points used for best fit linear regressions. Regression lines significant at p<O.Ol (0); p<O.OOl
(0).
scutum length) (6% of total) had fully developed testes (Fig. 6B). In five specimens
(< 10 mm scutum length) no testis could be found. Testes containing only spermatogo- nia and spermatocytes were found in individuals of, primarily, scutum length 5 to
17 mm. Spermatids, but no spermatozoa, were found in specimens of 10 to 22 mm scutum length.
Ovarian development. The ovary in stalked barnacles develops in the proximal part of the peduncle. Fecundity in P. kaempferi was low; with the maximum number of oocytes found in an individual was 20. In contrast the fecundity in L. anatifera was high. In both species oogenesis starts with clusters of oogonia (Fig. 7A,D). Oogonia grow into previtellogenic oocytes (Fig. 7E) which grow and at diameters > 20 pm undergo vitellogenesis (Fig. 7B,C,F). In mature P. kaempferi the ovary consists of no more than a few oocytes at the most proximal portion of the peduncle (Fig. 7B) whereas in mature
150 A. Green et al. /J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158
0 - 2 4 6 8 IO
16, Wet weight g
I I I / I I
14 B t
0 5 lo- 15 20 25 30
Scutum length mm
Fig. 5. Relationship of penis length against (A) adult wet weight and (B) scutum length for L. anatifera. 0, 0, points used for best fit linear regressions. Regression lines significant p = <O.OOl.
L. anatifru the ovary is a discrete structure in the proximal part of the peduncle
containing many oocytes (Fig. 7F). The oocyte size data were used to determine mean oocyte size for each individual.
Many P. kaempferi -c 9 mm scutum length contained no oocytes (Fig. 8A). The mean
oocyte size of individuals > 6 mm scutum length was very variable (Fig. 8A) and it was not possible to fit a curve to these data. In L. anatifru individuals of - 3 mm scutum length contained oocytes (Fig. SB). Most individuals < 14 mm scutum length had a mean oocyte diameter of < 40 pm although some individuals contained well developed oocytes at a scutum length of 12.5 mm. Mean oocytes size was variable in the larger L. anatifera examined (Fig. 8B). Again it was not possible to fit a curve to these data.
In P. kaempferi 40% of individuals > 8 mm scutum length contained egg lamellae. Degenerating ova, and possibly very early developing embryos, were found in the egg lamella. No egg lamellae were found in L. anatifru.
A. Green et al. /J. Exp. Mar. Biol. EcoL 184 (1994) 143-158 151
0’ 0 2 4 6 8 10 12 14
Scutum length mm
5 T I / I I E3 00 0
4-
0 5 10 15 20 35 30
Saturn length mm
Fig. 6. Distribution of male stages of development (see text) against scutum length. (A) Poecilasma kaempferi, (B) L. anatzfera.
3.1. Time scale for development in P. kaempferi and L. anatifera
Because data are available for the growth rate of P. kaempferi in deep water (Lampitt, 1990) is was possible to determine the approximate timing of reproductive events in this species. Capitulum length data (Lampitt, 1990) were used to convert scutum length to age in days (Fig. 9). These data suggest that inititiation of both oogenesis and sper- matogenesis occurs at N 70 days, the first vitellogenic oocytes occur at - 155 days and the first ripe testis at - 145 days. Penis development is complete at - 145 days. Egg lamellae occur at - 170 days after settlement.
An attempt was made to determine the timing of the main g~etogenic events in L. anat$?ra. Data are already available for L. anatifra from its natural tropical/ subtropical habitat (Evans, 1958; Zann & Harker, 1978) and from laboratory experi-
152 A. Green et al. /J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158
Fig. 7. Oogenic development in P. kaempferi, (A) clusters of oogonia (arrowed); (B) Vitellogenic oocytes (arrowed) at the proximal end of the peduncle; (C) Vitellogenic oocytes; L. anatifera (D) clusters of oogo- nia (arrowed); (E) oogonia and previtellogenic (Pv) oocytes; (F) Vitellogenic oocytes. Scale bars: A, C, D, I3 = 50 pm; B, F = 100 pm.
mentation (Patel, 1959). Under laboratory conditions penis activity and fertilization in
this species occurs only at temperatures above 19 “C (Pate& 1959). These temperatures were reached only between day 190 and 215 of the deployment of the Argos buoy (Pingree & Le Gum, 1991). If cyprids from this reproductive event settled on the buoy they would form the cohort with a maximum scutum size of 20 mm (Fig. 1A). The end of the deployment would be 175 days later after the temperature dropped below 16 “C.
A. Green et al. /J. Exp. Mar. Biol. Ed. 184 (1994) 143-158 153
This would suggest a growth rate of 0.1 mm+day -’ although this rate is unlikely to be constant as the temperature dropped to 10 “C during this period. These data suggest
that testis development started at 5 mm scutum length ( - 43 days after settlement) and mature male organs are found at a scutum length of 15 mm ( - 13 1 days). Developing oocytes were found at a length of 3 mm (N 26 days) and mature oocytes at 13 mm scutum length (- 114 days).
60
40
20
0 2 4 6 8 10 12 14
100
80
60
40
20
0
B 0
0
Scutum length mm
I I I -_.- I’-- I
0
1 0 5 10 15 20 25 30
Scutum length mm
Fig. 8. Average oocyte diameter as a function of scutum length (A) P. k nernpferi. (B) L. nrraf$~u.
154 A. Green et al. 1 J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158
E ,, 1; y = 59.09exp0.12x i II & 10
c” at : .-
E Ls ;
+’ 2 5
v) 4 2 1 0’ I I I I
0 50 100 150 200 250
Julian Day after settlement
Fig. 9. Growth curve of P. kaempferi based on the data of Lampitt (1990). Exponential curve significant at p= <0.001.
4. Discussion
Since Darwin’s observations of lepadomorph barnacles rapid growth has been established as one of the prime characteristics of species living in shallow water or attached to floating objects pa~icularly in tropical and subtropical environments (Darwin, 1851; Evans, 1958; MacIntyre, 1966). Lampitt (1990) demonstrated clearly that the deep-sea species P. kaempferi also shows rapid growth. Data are available for a number of species for the size at which egg lamellae appear (Hilgard, 1960) although the timing of this event has been quoted only rarely (Evans, 1958; Barnes & Reese, 1960). In addition, very little is known of the timing of the main g~etogenic events in any lepadomorph species. Lewis and Chia (198 1) describe reproduction in Pollicipes poZymerus from intertidal waters off the San Juan Islands and showed that this species reached sexual maturity during the first year after settlement. Williams and Moyse (1988) suggest that P. kuempferi becomes sexually mature at 13 mm capitulum length. One of us (M.V.A) observed L. an&f& and Conchoderma virgatum that had settled on the rope droguing a drifting mooring deployed in the vicinity of 10.5” N 20” W (SST = 26 “C) for 1 I days during Discovery stations 6662-6665 in 1968, were already releasing nauplii. Cyprid larvae of both species were collected at the time, settled on corks and kept fed to satiation in jars at laboratory temperatures (N 25 “C). Both species achieved sexual maturity and were releasing nauplii within 13 days. Concho- derma auritum, usually an epibiont on humpback and sperm whales, occasionally presents a serious fouling problem on large bulk carriers. It is also known to grow rapidly although its maturation rate has not been recorded. By examining the game- togenic pattern in individuals of different sizes of P. kaempferi and L. anatifera it has
A. Green et al. /J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158 155
been possible to give a maximum estimate of the onset of gametogenesis, maturity, and
the formation of egg lamelllae.
It may not be surprising, even in a deep-sea species, that the rate of gametogenic development in P. kaempferi is rapid. Growth rate is known to be rapid (Lampitt, 1990) and our estimates suggest that the testis is fully developed at - 145 days, the first large oocytes are found at - 155 days and egg lamellae are found at 170 days. Penis growth appears to be complete at - 145 days. The temperature at the site of collection in the
Rockall Trough is approximately 3 ‘C and varies little throughout the year (Gage et al. 1980). These estimates may be biased by the deposition of phytodetritus that occurred during Lampitt’s study which increased the growth rate of the barnacle post-deposition
(Lampitt, 1990). The results suggest that L. anatifera, in these samples from the NE Atlantic, is un-
dergoing gametogenesis more slowly than that observed by individuals of the same
species in its typical tropical/subtropical habitat. The estimates suggest that L. anatifera
from the Argos buoy reach reproductive maturity at - 114 days for female organs and - 130 days for male organs. This slow rate is a function of the “low” temperature
throughout the deployment of the buoy compared with the subtropical temperatures of - 26 ‘C at sites of previous measurements. Exceptional growth rates have been found in both L. anatifera and L. hillii (Evans, 1958) and in both species at least one indi- vidual contained egg lamellae by day 30. Evans’ (1958) specimens were growing at 24.2 to 26.1 “C whereas the maximum temperature during the time of deployment of the Argos buoy was 10.2 to - 18.4 ‘C perhaps accounting for the observed slower repro-
ductive rate. No egg lamellae are found in even the largest individuals of L. anatifera.
This may be a result of the inactivity of the penis at ambient temperatures. Under laboratory conditions Pate1 (1959) observed penis activity only at > 19 ‘C, a tempera-
ture rarely reached in the oceanic NE Atlantic. Penis activity may occur below this temperature in the open ocean of the N. Atlantic as the population structure on the Argos buoy represents two cohorts, of which the smaller individuals which may have
resulted from reproduction in the larger individuals. The rapid growth and gametogenesis in both species may be a function of their life
style. P. kaempferi lives (naturally) on inter alia the exoskeleton of the deep-sea stone crab N. grimaldi (Williams & Moyse, 1988; P.A. Tyler & J.D. Gage, pers. obs.). The barnacle is found on the carapace, the spines and legs of the crab where it receives protection from predators and a food supply from the feeding of N. grimaldi. Like all
crabs N. grimaldi has to undergo ecdysis to grow; the periodicity at which ecdysis occurs in N. grimaldi is unknown, but it may occur every 2 yr (A.L. Rice, pers. comm). If this
be the case, once ecdysed, the exoskeleton will disintegrate leaving the epizoic P. kaempferi with no substratum. P. kaempferi has been collected attached to the exuviae of crabs but in this condition the barnacle will be subjected to covering by sediment or predation. As a result, selective pressure, has lead to P. kaempferi grow-
ing and reproducing rapidly within the time limit set by the intermoult of the host, to allow dispersal and colonisation to new crabs exoskeltons. N. grimaldi is not common in the NE Atlantic and < 50% of the specimens taken in trawls are covered with P. kaempferi (P.A. Tyler & J.D. Gage, pers. obs.). This suggests that P. kaempferi has not been able to colonise all the N. grimaldi or that newly ecdysed ones are not avail-
1% A. Green et al. 1 J. Exp. Mar. Bid. Ed. I84 (1994) 143-158
able for colonisation. A similar life history has been suggested for Platyiepas ophiophilus that lives on sea snakes that slough every 4 to 6 wk and for Octolasmis warwickii that lives on the lobster ~henus orientates which, under aquarium conditions, moults 2 or 3 times each year (Zann & Harker, 1978)
Lepas anatifera lives on floating objects in the open ocean. Although both inert natural and anthropogenic objects are observed at the sea surface they are by no means common. Once one is located and settled by larvae, growth to maturity needs to be rapid, partly because the competition for space is often intense, and because most natural floating objects (wood, feathers, shells, and floating shells such as Spirula) are ephemeral. Thus, before man introduced more permanent objects such as fishing floats, as well as a greater range of ephemera (wooden crates, buoyancy rope and fuel oil) there would have been strong selective pressure for rapid growth, early maturity and iteropa- rous reproduction to produce the maximum number of larvae as quickly as possible.
The settlement on a satellite-tracked buoy has provided a unique, albeit serendipi- tous, case history of L. anatifera growth and development, and one that would merit a properly designed experiment. The general track of the floating buoys in this part of the northeast Atlantic is to follow the main current drift to the northeast. Buoys which are not circulating in such stationary eddies normally track rapidly across the Faroes/ Iceland Ridge into the Norwegian Sea and in a few weeks will be moving northwards along the coast of Norway. L. anatifera is a species found normally further south. Cyprids occur patchily and seasonally in samples collected by the Continuous Plank- ton Recorder (Roskell, 1975). The vernacular name of the goose barnacle refers to the coincidence of the barnacles washed up on beaches when the geese disappeared on their springtime migrations to their breeding grounds at high latitudes, (which led people, in medieval times, to believe the geese and barnacles were alternating forms of the same beast).
There are a number of other planktonic species that show similar patchy and sea- sonal occurrences off Britain. The neustonic chondrophore Vellela sp. is, occasionally, found in swarms at 50” N off Britain. This, and other ‘“Lusitanian” species (sensu Fraser, 1968) were considered to have arisen as part of the Mediterranean outflow and advected north to the north of Scotland. The observations of L. anat@-a suggest mesoscale eddies play a significant role in the erratic seasonal occurrence of these species in more northern waters. It seems likely that the quasi-stable eddies, such as that tracked by Pingree and Le Cann (1991), allow these species to persist at higher latitudes longer than normal and that the ecology of pelagic communities is strongly influenced by the eddy environment.
The driving force in the evolution of rapid growth and reproduction in stalked bar- nacles, and other deep-sea species, may be the ephemeral nature of the substratum to which they are attached. This reproductive pattern would appear to occur in other tr~sitory env~onments such as wood (Turner, 1973) and hydrothe~al vents (Tunnicliffe, 1991).
A. Green et al. /J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158 157
Acknowledgements
We wish to thank Drs. R.S. Lampitt and A.L. Rice (IOSDL) for discussions and
access to data on growth rates of Poecilusma kaempferi and to Dr. R. Williams (Uni- versity of Wales, Swansea) for discussions. The manuscript benefitted considerably from the attention of Dr. M. Barnes (SAMS, Dunstaffnage Marine Laboratory, Oban).
AG was supported by an NERC Advanced Course studentship which is gratefully acknowledged.
References
Barnes, H. & E.S. Reese, 1960. The behaviour of the stalked intertidal barnacle Pollicipes polymerus J.B. Sowerby, with special reference to its ecology and distribution. J. Anim. Ecol., Vol. 29, pp. 169-
185.
Barnes, M., 1989. Egg production in cirripedes. Oceanogr. Mar. Biol. Annu. Rev., Vol. 27, pp. 91-166.
Barnes, M., 1992. The reproductive periods and condition of the penis in several species of common
cirripedes. Oceanogr. Mar. Biol. Annu. Rev., Vol. 30, pp. 483-525.
Batham, E.J., 1945. Pollicipes spinosus Quoy and Gaimard. I. Notes on the biology and anatomy of the adult
barnacle. Trans. R. Sot. N.Z., Vol. 74, pp. 359-374.
Cimberg, R.L., 1981. Variability in brooding activity in the stalked barnacle Pollicipes polymerus. Biol. Bull., Vol. 160, pp. 31-42.
Darwin, C., 185 1. A monograph on the sub-class Cirripedia, with figures of all the species of the Lepadidae: or, pedunculated cirripedes. The Ray Society, London, 400 pp.
Eckert, K.L. & S.A. Eckert, 1987. Growth rate and reproductive condition of the barnacle Conchoderma virgatum on gravid leatherback sea turtles in Caribbean waters. J. Crust. Biol., Vol., 7, pp. 682-690.
Evans, F., 1958. Growth and maturity of the barnacles Lepas hillii and Lepas anatijkra. Nature, Vol. 182,
pp. 1245-1246.
Fraser, J.H., 1968. The overflow of oceanic plankton to the shelf waters of the north-east Atlantic. Sarsia, Vol. 34, pp. 313-330.
Gage, J.D., 1990. Skeletal growth markers in the deep-sea brittle stars Ophiura ijungmani and Ophiomusium lymani. Mar. Biol., Vol. 104, pp. 427-435.
Gage, J.D. & P.A. Tyler, 1991. Deep-sea biology: a natural history of organisms at the deep-seajoor. Cambridge
University Press, 504 pp.
Gage, J.D., R.H. Lightfoot, M. Pearson & P.A. Tyler, 1980. An introduction to a time series of abyssal
macrobenthos: methods and principal sources of variability. Oceanol. Acta, Vol. 3, pp. 169-176.
Gooday, A.J., 1988. A response by benthic Foraminifera to deposition ofphytodetritus in the deep sea Nature,
Vol. 332, pp. 70-73.
Hilgard, G.H., 1960. A study of reproduction in the intertidal barnacle Mite/la polymerus, in Monterey Bay,
California. Biol. Bull., Vol. 119, pp. 169-188.
Lampitt, R.S., 1990. Directly measured rapid growth of a deep-sea barnacle. Nature, Vol. 345, pp. 805- 807.
Lewis, CA. & F-S. Chia, 1981. Growth, fecundity and reproductive biology in the pedunculate cirripede
Pollicipes polymerus at San Juan Island, Washington. Can. J. Zool., Vol. 59, pp. 893-901.
Lochte, K. & CM. Turley, 1988. Bacteria and cyanobacteria associated with phytodetritus in the deep sea.
Nature , Vol. 333, pp. 67-69.
MacIntyre, R.J., 1966. Rapid growth in stalked barnacles. Nature, Vol. 212, pp. 637-638.
Merrett, N.R. & N.B. Marshall, 1981. Observations on the ecology of deep-sea bottom-living fishes collected
off northwest Africa (08”-27”N). Prog. Oceanogr., Vol. 9, pp. 185-244.
Orton, J.H., 1920. Sea-temperature, breeding and distribution in marine animals. J. Mar. Biol. Assoc. U.K., Vol. 12, pp. 339-366.
158 A. Green et al. /J. Exp. Mar. Biol. Ecol. 184 (1994) 143-158
Patel, B., 1959. The influence of temperature on the reproduction and moulting of Lepus anatifera L. under
laboratory conditions. J. Mar. Biol. Assoc. U.K., Vol. 38, pp. 589-597.
Pingree, R.D. & B. Le Cann, 1991. Drifting Buoy in the field of flow of two eddies on East Thulean Rise
(Northeast Atlantic). J. Geophys, Res., Vol. 96, pp. 16759-16777.
Roskell, J., 1975. Continuous plankton records: A plankton atlas of the North Atlantic and the North Sea.
Supplement 2. The oceanic cirripede larvae 1955-1972. Bull. Mar. Ecol., Vol. 8, pp. 185-199.
1992. The BOFS 1990 Spring Bloom Experiment: Temporal evolution and spatial variability of the
hydrographic field. Prog. Oceanogr., Vol. 29, pp. 235-281.
Scheltema, A.H., 1987. Reproduction and rapid growth in a deep-sea aplacophoran mollusc, Prochaetoderma yongei. Mar. Ecol. Prog. Ser., Vol. 37, pp. 171-180.
Tsikhon-Lukanina, Y.A., I.N. Soldatova, LA. Kuznetsova & 1.1. Ll’in, 1976. Macrofouling community in
the Strait of Tunisia (Sicily). Oceanology, Vol. 16, pp. 519-521.
Tunnicliffe, V., 1991. The biology of hydrothermal vents: ecology and evolution. Oceanogr. Mar. Biol. Annu. Rev., Vol. 29, pp. 319-407.
Turekian, K., J.K. Cochran, D.P. Kharkar, R.M. Cerrato, J.R. Vaisnys, H.L. Sanders, J.F. Grassle & J.A.
Allen, 1975. Slow growth rate of a deep-sea clam determined by “sRa chronology. Proc. Nat. Acad. Sci., Vol. 72, pp. 2829-2832.
Turner, R.D., 1973. Wood boring bivalves, opportunistic species in the deep sea. Science, Vol. 180, pp. 1377-
1379.
Tyler P.A., J.D. Gage & D.S.M. Billett, 1992. Reproduction and recruitment in deep-sea invertebrate
populations in the NE Atlantic Ocean: a review of the options. In, Marine eutrophication and population dynamics, edited by G. Colombo et al., Olsen and Olsen, Fredensborg, pp. 257-262.
Williams, R. & J. Moyse, 1988. Occurrence, distribution and orientation of Poecilasma kaempferi Darwin
(Cirripedia: Pedunculata) epizoic on Neolithodes grimaldi Milne-Edwards and Bouvier (Decapoda: Ano-
mura) in the northeast Atlantic. J. Crust. Biol., Vol. 8, pp. 177-186.