Copyright Ó 2010 by the Genetics Society of America DOI: 10.1534/genetics.110.116756 Gene Duplication, Gene Conversion and the Evolution of the Y Chromosome Tim Connallon 1 and Andrew G. Clark Department of Molecular Biology and Genetics, Cornell University, Ithaca, New York 14853-2703 Manuscript received March 17, 2010 Accepted for publication May 31, 2010 ABSTRACT Nonrecombining chromosomes, such as the Y, are expected to degenerate over time due to reduced efficacy of natural selection compared to chromosomes that recombine. However, gene duplication, coupled with gene conversion between duplicate pairs, can potentially counteract forces of evolutionary decay that accompany asexual reproduction. Using a combination of analytical and computer simulation methods, we explicitly show that, although gene conversion has little impact on the probability that duplicates become fixed within a population, conversion can be effective at maintaining the functionality of Y-linked duplicates that have already become fixed. The coupling of Y-linked gene duplication and gene conversion between paralogs can also prove costly by increasing the rate of nonhomologous crossovers between duplicate pairs. Such crossovers can generate an abnormal Y chromosome, as was recently shown to reduce male fertility in humans. The results represent a step toward explaining some of the more peculiar attributes of the human Y as well as preliminary Y-linked sequence data from other mammals and Drosophila. The results may also be applicable to the recently observed pattern of tetraploidy and gene conversion in asexual, bdelloid rotifers. N ONRECOMBINING chromosomes are often asso- ciated with genetic degradation and a loss of functional genes, and nowhere is this pattern more exaggerated than on the Y chromosome (Charlesworth and Charlesworth 2000; Bachtrog 2006). However, in addition to the more widely recognized pattern of gene loss, genome sequences of mammals and Drosoph- ila are also yielding evidence for Y-linked functional gene gain followed by amplification of duplicate genes (Skaletsky et al. 2003; Koerich et al. 2008; Carvalho et al. 2009; Krsticevic et al. 2009; Hughes et al. 2010). Duplication and retention of functional Y-linked gene copies is somewhat surprising because evolutionary the- ory predicts an opposing pattern. First, to the extent that gene duplicates are fixed via positive selection, they are less likely to become fixed on nonrecombining relative to recombining chromosomes (Otto and Goldstein 1992; Clark 1994; Yong 1998; Otto and Yong 2002; Tanaka and Takahasi 2009). Second, regardless of whether Y-linked duplicates become fixed via genetic drift or by natural selection, the actions of Muller’s ratchet, genetic hitchhiking, and background selection are expected to greatly increase the probabil- ity that Y-linked genes degenerate into nonfunctional pseudogenes (Charlesworth and Charlesworth 2000; Bachtrog 2006; Engelstadter 2008). The issue is more complex when one considers data from the well-characterized human Y chromosome. A majority of functional Y-linked genes are members of duplicate gene pairs residing within large palindromes and are almost exclusively testis expressed (Skaletsky et al. 2003). In contrast to many of the single-copy genes with X-linked homologs, members of Y-linked gene families are apparently not degenerating, but rather have become fixed and maintained over many millions of years (Skaletsky et al. 2003; Yu et al. 2008). Although Y chromosomes are not well characterized in other taxa, currently available data suggest that duplication is a common feature of Y chromosomes in other mammal species as well as Drosophila (Rozen et al. 2003; Verkaar et al. 2004; Murphy et al. 2006; Alfo ¨ ldi 2008; Wilkerson et al. 2008; Krsticevic et al. 2009; Geraldes et al. 2010). Thus, patterns of gene duplication and retention, for at least a subset of Y-linked genes, may be a general rule of Y chromosome evolution. Another attribute of the mammalian Y appears to be relevant for duplicate gene evolution. Comparative analysis between humans and chimpanzees suggests ongoing recombination between the gene duplicate pairs that reside on the same Y chromosome. Such ‘‘intrachromosomal’’ recombination includes both non- reciprocal (gene conversion) and reciprocal exchange (crossing over) between gene duplicate pairs (Rozen et al. 2003; Lange et al. 2009). Gene conversion between Supporting information is available online at http://www.genetics.org/ cgi/content/full/genetics.110.116756/DC1. 1 Corresponding author: Department of Molecular Biology and Genetics, Cornell University, Biotechnology Bldg. (Room 227), Ithaca, NY 14853- 2703. E-mail: [email protected]Genetics 186: 277–286 (September 2010)
Transcript
Copyright � 2010 by the Genetics Society of AmericaDOI: 10.1534/genetics.110.116756
Gene Duplication, Gene Conversion and the Evolutionof the Y Chromosome
Tim Connallon1 and Andrew G. Clark
Department of Molecular Biology and Genetics, Cornell University, Ithaca, New York 14853-2703
Manuscript received March 17, 2010Accepted for publication May 31, 2010
ABSTRACT
Nonrecombining chromosomes, such as the Y, are expected to degenerate over time due to reducedefficacy of natural selection compared to chromosomes that recombine. However, gene duplication,coupled with gene conversion between duplicate pairs, can potentially counteract forces of evolutionarydecay that accompany asexual reproduction. Using a combination of analytical and computer simulationmethods, we explicitly show that, although gene conversion has little impact on the probability thatduplicates become fixed within a population, conversion can be effective at maintaining the functionalityof Y-linked duplicates that have already become fixed. The coupling of Y-linked gene duplication andgene conversion between paralogs can also prove costly by increasing the rate of nonhomologouscrossovers between duplicate pairs. Such crossovers can generate an abnormal Y chromosome, as wasrecently shown to reduce male fertility in humans. The results represent a step toward explaining some ofthe more peculiar attributes of the human Y as well as preliminary Y-linked sequence data from othermammals and Drosophila. The results may also be applicable to the recently observed pattern oftetraploidy and gene conversion in asexual, bdelloid rotifers.
NONRECOMBINING chromosomes are often asso-ciated with genetic degradation and a loss of
functional genes, and nowhere is this pattern moreexaggerated than on the Y chromosome (Charlesworth
and Charlesworth 2000; Bachtrog 2006). However,in addition to the more widely recognized pattern ofgene loss, genome sequences of mammals and Drosoph-ila are also yielding evidence for Y-linked functionalgene gain followed by amplification of duplicate genes(Skaletsky et al. 2003; Koerich et al. 2008; Carvalho
et al. 2009; Krsticevic et al. 2009; Hughes et al. 2010).Duplication and retention of functional Y-linked genecopies is somewhat surprising because evolutionary the-ory predicts an opposing pattern. First, to the extentthat gene duplicates are fixed via positive selection,they are less likely to become fixed on nonrecombiningrelative to recombining chromosomes (Otto andGoldstein 1992; Clark 1994; Yong 1998; Otto andYong 2002; Tanaka and Takahasi 2009). Second,regardless of whether Y-linked duplicates become fixedvia genetic drift or by natural selection, the actions ofMuller’s ratchet, genetic hitchhiking, and backgroundselection are expected to greatly increase the probabil-ity that Y-linked genes degenerate into nonfunctional
pseudogenes (Charlesworth and Charlesworth
2000; Bachtrog 2006; Engelstadter 2008).The issue is more complex when one considers data
from the well-characterized human Y chromosome. Amajority of functional Y-linked genes are members ofduplicate gene pairs residing within large palindromesand are almost exclusively testis expressed (Skaletsky
et al. 2003). In contrast to many of the single-copy geneswith X-linked homologs, members of Y-linked genefamilies are apparently not degenerating, but ratherhave become fixed and maintained over many millionsof years (Skaletsky et al. 2003; Yu et al. 2008). AlthoughY chromosomes are not well characterized in other taxa,currently available data suggest that duplication is acommon feature of Y chromosomes in other mammalspecies as well as Drosophila (Rozen et al. 2003; Verkaar
et al. 2004; Murphy et al. 2006; Alfoldi 2008; Wilkerson
et al. 2008; Krsticevic et al. 2009; Geraldes et al. 2010).Thus, patterns of gene duplication and retention, for atleast a subset of Y-linked genes, may be a general rule ofY chromosome evolution.
Another attribute of the mammalian Y appears to berelevant for duplicate gene evolution. Comparativeanalysis between humans and chimpanzees suggestsongoing recombination between the gene duplicatepairs that reside on the same Y chromosome. Such‘‘intrachromosomal’’ recombination includes both non-reciprocal (gene conversion) and reciprocal exchange(crossing over) between gene duplicate pairs (Rozen
et al. 2003; Lange et al. 2009). Gene conversion between
Supporting information is available online at http://www.genetics.org/cgi/content/full/genetics.110.116756/DC1.
1Corresponding author: Department of Molecular Biology and Genetics,Cornell University, Biotechnology Bldg. (Room 227), Ithaca, NY 14853-2703. E-mail: [email protected]
the duplicates potentially maintains gene function bycounteracting stochastic forces of Y chromosome de-generation (Rozen et al. 2003; Charlesworth 2003;Noordam and Repping 2006). The rationale behindthis hypothesis is subtle. As with other clonally inheritedchromosomes, each evolutionary lineage of the Y is phys-ically coupled to, and its evolutionary fate is influencedby, the presence of deleterious mutations. Mutation-bearing lineages represent evolutionary dead ends unlessthey can somehow remove or compensate for deleteriousmutations. Recombination between duplicates can ‘‘res-cue’’ functionality via gene conversion between func-tional and nonfunctional copies.
On the other hand, double-strand DNA breaks, whichprecede gene conversion events (Marais 2003), alsoprecede crossing over. Crossovers between Y-linkedgenes can generate acentric and dicentric Y chromo-somes, resulting in infertility and disruption of thesex determination pathway (e.g., Repping et al. 2002;Heinritz et al. 2005; Lange et al. 2009). Consideringboth gene conversion and crossing over on the Y, re-combination can be viewed as a factor that either con-strains (via gene conversion) or promotes (via crossingover) Y chromosome degeneration.
These observations concerning Y chromosome genecontent and recombination raise interesting questionsthat have not been formally addressed by evolutionarytheory (but see the recent study by Marais et al. 2010).First, what conditions favor the evolutionary invasion ofY-linked gene duplicates, and does recombination in-fluence the probability that duplicates eventually becomefixed within a population? Second, what affect doesrecombination have on Y-linked fitness and the mainte-nance of functional duplicate genes? To address thesequestions, we develop and analyze a series of population-genetic models of Y chromosome evolution. We showthat, when direct selection on gene duplicates is weak,biased gene conversion can increase, whereas crossingover will decrease, their probability of fixation. For dupli-cates with larger fitness effects, the probability of fixationis largely independent of Y-linked recombination. Finally,gene conversion has a major impact on the retention offunctional Y-linked genes that are already fixed within thepopulation and maintains multiple gene copies with orwithout selection favoring these duplicates.
MODEL AND RESULTS
Gene conversion and the invasion of new geneduplicates: We first consider conditions favoring theevolutionary invasion of new Y-linked duplicate genes atlow initial frequency within the population. Determin-istic invasion dynamics are described for a two-locusmodel, and it is shown separately that the two-locusmodel characterizes duplicate gene invasion conditionson a Y chromosome carrying an arbitrary number ofgenes (see supporting information, File S1). We then
develop and analyze a diffusion approximation andperform stochastic simulations to examine the proba-bility that a rare gene duplicate eventually becomesfixed within a population of small size.
Invasion of a new gene duplicate: Consider a singleY-linked locus with a functional allele, A, and a non-functional allele, a. Mutation from A to a occurs at rate uper generation and there is no back mutation. Byintroducing a duplication of the locus, the population isexpanded to include five genotypic classes: the originalsingle-copy classes (A and a), those with two functionalgene copies (AA), those with one functional and onenonfunctional copy (Aa), and those with two nonfunc-tional copies (aa). As in the single-locus case, transitionsbetween states (AA / Aa or aA; Aa or aA / aa) can occurby mutation, at rate of u per locus; because there are nowtwo loci, the mutation rate per chromosome is 2u.
For Y chromosomes carrying duplicates, recombina-tion (crossing over and gene conversion) can poten-tially occur between loci. Throughout our analysis, weexamine cases where recombination occurs at a rate of dper paralog pair, per generation. The probability that asingle recombination event is a crossover, which gen-erates an abnormal (sterile) Y chromosome (e.g.,Repping et al. 2002; Heinritz et al. 2005; Lange et al.2009), is equal to the constant c . The remainder ofrecombination events (1� c) represent gene conversionevents between duplicate pairs. Gene conversion in-volving Aa or aA individuals yields AA or aa sperm at rateb and 1 � b, respectively. Thus, b can be viewed as abiased gene conversion parameter, where the functionalcopy A preferentially replaces the nonfunctional awhenever b . 0.5 (there is no bias when b ¼ 0.5).
Compared to individuals with two functional genecopies, individuals with zero functional copies suffer afitness reduction of s, while those with one functionalcopy suffer a reduction of sh, where h is equivalent to adominance coefficient. Complete masking of a non-functional allele occurs when h ¼ 0, and there is nodirect fitness benefit of carrying two vs. one functionalgene. Partial masking occurs when 1 . h . 0; in suchcases, there is a fitness benefit of having two functionalcopies. Genotypes, genotypic fitness, and zygotic fre-quencies are described in Table 1.
TABLE 1
Parameterization for the gene duplicate invasion model
Genotype Frequency Fitness
AA x11 1Aa, aA x10 1 � shA x1 1 � shaa x00 1 � sa x0 1 � sAbnormal Y xs 0
For a sequence of events of (i) birth, (ii) selection,(iii) mutation, (iv) recombination, and (v) randommating (and ignoring factors of u2), the frequencychange of each genotype, per generation, is given bythe following six recursions,
x119¼x11
wð1� 2uÞð1� dcÞ
1x11
w2udð1� cÞb 1
x10ð1� shÞw
ð1� uÞdð1� cÞb
x109¼x11
w2uð1� dÞ1 x10ð1� shÞ
wð1� uÞð1� dÞ
x009¼x11
w2udð1� cÞð1� bÞ1 x10ð1� shÞ
wuð1� dcÞ
1x10ð1� shÞ
wð1� uÞdð1� cÞð1� bÞ1 x00ð1� sÞ
wð1� dcÞ
xs9 ¼x11
wdc 1
x10ð1� shÞw
dc 1x00ð1� sÞ
wdc
x19 ¼ x1ð1� shÞw
ð1� uÞ
x09 ¼ x1ð1� shÞw
u 1x0ð1� sÞ
w;
where mean fitness is �w ¼ x11 1 ðx10 1 x1Þð1� shÞ 1
ðx00 1 x0Þð1� sÞ.To describe conditions promoting the invasion of
duplicates, we analyzed the stability of an evolutionaryequilibrium in which duplicated genotypes are absentfrom the population. Under such a condition, the fre-quencies x1 and x0 equilibrate to x1 ¼ 1� uð1� hsÞ=½sð1� hÞ� ¼ 1� x0 and the leading eigenvalue of thestability matrix is
Selection favors the invasion of a duplicate when theleading eigenvalue is greater than one (Otto and Day
2007). The magnitude of the leading eigenvalue alsorepresents the strength of selection acting in favor of a rareduplicate gene [i.e., the probability of fixation is pro-portional to l (Otto and Bourguet 1999; Otto andYong 2002); see below for additional details]. Withoutrecombination (d¼ 0), the leading eigenvalue reduces to
l ¼ 1
21
1� 2u 1 jshð1� uÞ � uj2ð1� shÞð1� uÞ ð1bÞ
and evolutionary invasion of a duplicate-bearing Y isfavored when sh . u/(1 � u). Duplicates are favored
when the direct fitness benefit of additional functionalgene copies outweighs the indirect consequences ofdoubling the deleterious mutation rate, as previouslyreported for both haploid and diploid systems withoutrecombination (Clark 1994; Otto and Yong 2002; alsosee Otto and Goldstein 1992).
How does recombination alter the evolutionary dy-namics of Y chromosomes? When duplicates do notdirectly increase fitness (sh ¼ 0), and there is no re-combination, selection never favors invasion (Equation1b above). We can ask whether gene conversion expandsthe conditions favorable to invasion of a duplicate in away that is similar to previous models of gene duplicationwith crossing over (Otto and Yong 2002). By permittingY-linked recombination between duplicates, and assum-ing that the crossover rate is zero (dc ¼ 0; hence, allrecombination is by gene conversion), the leading ei-genvalue can be approximated for low rates of geneconversion (d � 0, per generation),
l ¼ l
����d¼0
1@l
@d
����d¼0
d 1 Oðd2Þ � 1 1 dð2b � 1Þ; ð1cÞ
which indicates that selection favors duplicates (l . 1)when gene conversion is biased toward transmission offunctional over nonfunctional gene copies (b . 0.5).Numerical evaluation of Equation 1a indicates that,although higher rates of gene conversion can increasethe leading eigenvalue (and hence the probability ofinvasion), this positive relationship quickly saturates.Thus, a little bit of gene conversion has about as muchof an impact on the leading eigenvalue as a high rate ofgene conversion does. Nevertheless, the strength ofsuch positive selection (with magnitude of l � 1) is onthe order of the mutation rate (u) and is thereforeextremely weak. Stochastic simulations (see below) showthat the probability of duplicate fixation is marginallyinfluenced by biased gene conversion alone.
Further analysis of Equation 1a shows that, as with thecase of no recombination (Otto and Yong 2002),selection will favor duplicates if they directly increasefitness (sh . 0). Gene conversion (including unbiasedgene conversion: b ¼ 0.5) can increase the strength ofselection favoring invasion of a duplicate (l � 1; Figure1). However, the relative impact of gene conversion isminor when sh ? u. In other words, when there areweak direct benefits of having multiple gene copies, thestrength of natural selection favoring Y-linked geneduplicates will be enhanced by gene conversion be-tween paralogs. This conclusion holds if the crossoverrate between duplicate pairs (dc) is small (Figure 1). Asthe rate of crossing over increases, the production ofabnormal Y haplotypes can generate purifying selectionagainst Y chromosomes that carry gene duplicates.
Why should gene conversion broaden duplicate in-vasion conditions under weak selection? An intuitiveexplanation can be reached by considering the recursion
Gene Conversion and Y Evolution 279
dynamics for a population fixed for the single-gene hap-lotype. Because this explanation is heuristic, we ignorecrossovers and assume that they do not occur (c¼ 0). Therate of increase for a rare haplotype with two functionalgene copies depends on its relative competitivenessagainst the resident, single-copy haplotype. For initialcondition x11 ¼ 1/N and x10 ¼ x00 ¼ 0, the expectedproportion of functional duplicate haplotypes (x11) withinthe gamete pool is E ½x119� ¼ x11½1� 2uð1� dbÞ�=�w, and
the duplicate is favored when ½1� 2uð1� dbÞ�=�w . 1.Invasion is clearly facilitated by gene conversion (db .
0). Nevertheless, because the term 2u(1� db) is extremelysmall, gene conversion will marginally influence theprobability of fixation whenever sh ? u.
Probability of duplicate fixation: The deterministicmodel presented above can be modified to describethe evolutionary dynamics in finite populations. Follow-ing Otto and Bourguet (1999) and Otto and Yong
(2002), the selection coefficient for a rare gene dupli-cate can be approximated as l � 1, where l is theleading eigenvalue of the stability matrix (Equation 1a,above). Given this selection coefficient, the probabilitythat a rare duplicate is eventually fixed can be estimatedby diffusion approximation (Kimura 1957, 1962), withdrift and diffusion coefficients M¼ (l� 1)x(1� x) andV¼ x(1� x)/N, respectively, where x is the frequency ofa duplicate-bearing Y haplotype and N is the Y chromo-some effective population size. For an initial frequencyof 1/N, the probability that a duplicate is fixed will be
PrðfixationÞ ¼ 1� e2ð1�lÞ
1� e2N ð1�lÞ �2ðl� 1Þ
1� e2N ð1�lÞ: ð2Þ
To assess the validity of Equation 2, we conductedcomputer simulations that incorporate mutation, selec-tion, and genetic drift. Each simulation was initiated atx11¼ 1/N, x0¼ u(1� hs)/(s� sh), and x1¼ 1� x11� x0.To generate genotypic frequencies for the next gener-ation, N genotypes were randomly drawn from amultinomial distribution, after selection, from the sixgenotypes described above. Mutation–selection–driftrecursions were iterated until the duplicate genotypewas either fixed or lost from the population. Equation 2provides a good approximation for the probability ofduplicate fixation over a broad range of parameterspace (Figure 2 and Figure S1). As direct selection on aduplicate approaches zero (sh / 0), the probability offixation approaches 1/N. As direct selection increases instrength (1 ? 1 � l ? 1/N), the probability of fixationapproaches 2(l � 1).
Gene conversion had little impact on the probabilityof duplicate fixation (see Figure S1). As shown above,the leading eigenvalue of the stability matrix is notsubstantially influenced by gene conversion unless sh isof similar order to u. Even though the selection co-efficient approximation (l � 1) can increase with geneconversion, its absolute magnitude under weak directselection (sh � 0) will generally be too small for naturalselection to be effective, unless of course Nu . 1, whichis particularly unlikely for Y-linked loci. Thus, geneconversion is unlikely to significantly enhance the rateof duplicate gene fixation, but can potentially reducethe fixation rate of duplicates if the rate of deleteriouscrossovers between paralogs is high.
Gene conversion and the maintenance of geneduplicates: A major hypothesis inspired by the human Ychromosome is that gene conversion between duplicates
Figure 1.—Gene conversion can enhance the strength ofpositive selection for rare duplicate genes, whereas crossoversselect against duplicates. Selection coefficient approxima-tions (l � 1) are based on the leading eigenvalue (Equation1a), as described and justified in the text, and are presented asa ratio of selection with (d . 0) vs. without recombination(d ¼ 0). Representative results are presented for u ¼ 10�5
and assume that there is no gene conversion bias (i.e., b ¼ 0.5).
may prevent the accumulation of mutations and ulti-mately prevent or slow down Y chromosome degenera-tion due to Muller’s ratchet (Charlesworth 2003;Rozen et al. 2003; Noordam and Repping 2006). To for-mally evaluate this possibility, we considered two modelsfor the maintenance of functional Y-linked genes. We firstconducted simulations of our two-locus model with initialcondition x11 ¼ 1 (a pair of functional duplicates is ini-tially fixed within the population) and analyzed whethergene conversion prevented the loss of one or both of thefunctional gene copies. Gene conversion between Y-linkedparalogs decreased the rate of gene loss under a widerange of fitness conditions, including the extreme casewhere there was no direct benefit of having two, asopposed to one, functional gene copies (Figure S2).Although gene conversion can substantially reduce therate of gene loss, the results indicate that loss of completelyredundant genes (where sh ¼ 0) will persist under geneconversion, albeit at a substantially reduced rate.
Prior models of Muller’s ratchet generally find thatthe rate at which deleterious mutations become fixeddepends upon both the strength of purifying selectionand the number of loci evolving on an asexual chromo-some (Charlesworth and Charlesworth 2000;Bachtrog 2008). To account for selection and geneconversion across many loci, we extended our model todescribe the degeneration of Y chromosomes carryingan arbitrary number of genes. To permit gene conver-sion, we assumed that each Y initially carries n distinctgene types, each with a duplicate copy (for a total of 2nloci). Because the increased number of genes greatlyexpands the number of possible genotypic and fitnessstates (and consequently the matrix of transition prob-
abilities between states), we made a simplifying assump-tion that each of the n gene types represents an essentialmale fertility factor. Males lacking a functional copy ofone or more gene types are sterile and comprise aheterogeneous genotypic class with reproductive suc-cess of zero. Although the essentiality assumption isuseful for modeling purposes, it will often be biologi-cally reasonable because Y-linked genes, at least inmammals and Drosophila, are often essential for malefertility. For example, human Y chromosome micro-deletions within Y-palindromic regions are often associ-ated with spermatogenic failure (Noordam and Repping
2006; Lange et al. 2009). In Drosophila melanogaster,mutations in at least three of seven currently Y-annotatedgenes (kl-2, kl-3, and kl-5, as well as an additional set ofunannotated genes: kl-1, ks-1, and ks-2; data obtainedfrom http://flybase.org/) are known to cause male-sterilephenotypes. Nevertheless, the overall agreement be-tween our multilocus and two-locus results (the latterdoes not assume essentiality; see Figure S2) suggests thata violation of the essentiality assumption is unlikely tostrongly affect our conclusions.
For each paralog pair, there are three possiblegenotypes: both loci functional, one functional andone nonfunctional, and both nonfunctional. Transi-tions between genotypic states can occur by mutation,by gene conversion, or by crossing over, with crossoveryielding an abnormal Y chromosome. For individualscarrying a structurally normal Y, fitness follows thefunction w¼ (1 � sh)k(0)j, where j refers to the numberof gene pairs with both copies nonfunctional, and krefers to the number of pairs where one of the two genecopies is functional (0 # k # n). Individuals with j . 0and individuals carrying abnormal Y chromosomes aresterile. After selection, the reproductive contribution ofan individual with k Y-linked mutations is
xkS ¼xkwk
w;
where xk is the zygotic frequency of k-bearing males, wk¼(1 � sh)k is the fitness of a male with k mutations, andmean male fitness with respect to the Y is �w ¼
Pnk¼0 xkwk .
(The reproductive contribution of sterile individuals iszero.)
To facilitate analytical tractability, we assume that therates of recombination and mutation are both smallenough to ignore multiple mutation and multiple re-combination events per generation. In other words, thereis a zero probability of an individual with k mutationsproducing a fertile son with k� 2 or k 1 2 mutations. Thisassumption is justified as long as 2nu > 1 and nd > 1,which requires that the mutation and recombination rateper locus is small, and the number of loci mutable to anonfunctional allele is much smaller than the reciprocalof the mutation or gene conversion rate: n > min[1/u, 1/d]. Because n represents a small fraction of Y-linkednucleotides (i.e., it represents a very specific functional
Figure 2.—The probability of fixation for Y-linked dupli-cate genes. The solid line depicts the analytical approxima-tion from Equation 2. Circles represent the proportion ofduplicate genotypes (out of 100,000 replicate simulationsfor each data point) that eventually become fixed withinthe population. Results are shown for d ¼ 0, N ¼ 1000, andu ¼ 10�5, per locus, per generation. Values of d . 0 yield ap-proximately the same results (see Figure S1).
class), this assumption is biologically reasonable. Never-theless, a violation of these assumptions is expected tomake our results conservative by downwardly biasing thespeed of Muller’s ratchet (which is enhanced by a highermutation rate) and minimizing the positive effect of geneconversion (higher gene conversion rates increasinglycounteract Muller’s ratchet). Extending across the 2nloci, the probability that a Y chromosome experiencesone mutation is Pr(M ¼ 1) ¼ 2nu ¼ U. The probabilitythat zero mutations occur is Pr(M ¼ 0) ¼ 1 � U. Theprobability of a recombination event between one of the nparalog pairs is Pr(R¼ 1)¼ nd¼ D. The probability of norecombination is Pr(R ¼ 0) ¼ 1 � D.
Given a sequence of events of (i) birth, (ii) selection,(iii) mutation, (iv) recombination, and (v) random mat-ing, the frequency of fertile males in the next generationfollows the recursion
xk9 ¼�xk�1ð1� shÞk�1
w
U ðn � k 1 1Þn
1xkð1� shÞk
w
Uk 1 2nð1� U Þ2n
�
3Dð1� cÞðn � kÞ
n1 1� D
� �
1�xkð1� shÞk
w
U ðn � kÞn
1xk11ð1� shÞk11
w
3U ðk 1 1Þ1 2nð1� U Þ
2n
�
3Dbð1� cÞðk 1 1Þ
n:
The ‘‘least-loaded’’ (k¼ 0) and ‘‘most-loaded’’ (k¼ n)classes of fertile males follow the recursion
x09 ¼ x0
w
ð1� U Þð1� DcÞn 1 UDð1� cÞbn
� �
1x1ð1� shÞ
w
U
2n1 1� U
� �Dð1� cÞb
n
� �
and
xn9 ¼ xn�1ð1� shÞn�1
w
U ð1� DÞn
1xnð1� shÞn
w
ð2� U Þð1� DÞ2
;
respectively. The frequency of sterile males in the nextgeneration (via crossover, mutation, or gene conver-sion) will be
xs9 ¼ 1�Xn
k¼0
xk9:
Deterministic equilibria and mean fitness of the Y: Whenthere is no recombination between duplicates (D ¼ 0),
mean Y chromosome fitness as well as the distribution ofmutations among individuals can be analytically de-termined. If mutations that eliminate duplicate genefunction are deleterious (sh . 0), and the number ofunique Y-linked genes is large (n ? U/sh), the pop-ulation approaches the equilibrium: xk � PoisðU=shÞ,xs � 0, and �w � 1� U . This is analogous to the case ofmutation–selection balance with incomplete domi-nance (sh . 0), with a Y-linked genetic load of L ¼ U �1 � e�U (e.g., Haldane 1937; Kimura and Maruyama
1966; Kondrashov and Crow 1988). When knockingout a duplicate yields no fitness effect (sh ¼ 0), or thenumber of Y-linked genes is small (n > U/sh), thepopulation approaches the equilibrium: xn � 1� U=2,xs � U =2, and �w � 1� U =2. Under this scenario, thegenetic load is reduced by a factor of 2, to L¼U/2� 1�e�U/2 (Haldane 1937).
Gene conversion between duplicates increases thefrequency of the least-mutated class (Figure 3 and FigureS3), whether or not there is a gene conversion biasfavoring functional over nonfunctional loci. The fre-quency of the least-loaded class represents a quantity ofparticular importance for adaptation on clonally trans-mitted chromosomes such as the Y (Charlesworth
and Charlesworth 2000). Without recombination,the unit of selection is the chromosome rather than thelocus. Beneficial mutations that are associated withmutation-free genetic backgrounds are relatively likely
Figure 3.—Gene conversion increases the frequency of Ychromosome haplotypes that carry zero deleterious muta-tions (i.e., the ‘‘least-loaded’’ genotypic class). The cost of amutation eliminating function of a copy of each duplicatepair is represented by sh (this cost increases from left to righton the x-axis). The relative proportion of mutation-free Ychromosomes in recombining vs. nonrecombining popula-tions is presented as a ratio of the two scenarios (gene conver-sion increases the proportion of mutation-free Y’s when thisratio is greater than one). The number of distinct, Y-linkedgenes is represented by n. Results are presented for c ¼ 0,b ¼ 0.5, and u ¼ 5 3 10�4, per locus, per generation, andD ¼ U ¼ 2nu. Additional results are presented in Figure S3.
to become fixed (Peck 1994; Orr and Kim 1998) and donot permit hitchhiking of deleterious mutations duringa selective sweep (Rice 1987). However, as the frequencyof the least-loaded class becomes small, virtually allbeneficial mutations will arise in inferior genetic back-grounds. This will limit the adaptive potential of theY chromosome. Because it increases the fraction ofmutant-free Y chromosomes, gene conversion is ex-pected to enhance the fixation probability for beneficialmutations and can reduce the deleterious consequen-ces of hitchhiking.
By shifting the mutational distribution toward rela-tively mutation-free genotypes, gene conversion alsoincreases mean Y chromosome fitness. This effect doesnot depend on a gene conversion bias, but can becomeexacerbated when conversion events favor functionalover nonfunctional variants (for models yielding similarconclusions about the genetic load, albeit by differentapproaches, see Bengtsson 1986, 1990, and especiallyOhta 1989).
These long-term effects of gene conversion can beaccounted for by a straightforward explanation. Whenthe fitness cost of silencing both copies of a duplicatepair is much greater than the cost of silencing one of thecopies (when duplicates partially or completely maskdeleterious mutations: h , 0.5), selection across Y chro-mosomes mimics truncation selection, which is par-ticularly efficient at removing deleterious alleles (e.g.,Kondrashov 1988; Ohta 1989). Truncation selectionarises because mutations on a relatively mutation-free Ywill generally affect one copy of a pair, with the second,functional copy compensating for loss of the first. As thenumber of mutations on a Y increases, so does theprobability of silencing the second copy of a pair. Con-sequently, the deleterious effect of each mutation in-creases faster than linearly with the number of mutationscarried on a Y.
Without recombination, the accumulation of muta-tions is unidirectional, and the population will tend toevolve toward the edge of the truncation point (nmutations at distinct genes), particularly if masking byduplicates is strong (i.e., having two functional copiesprovides the same fitness as one copy). At the extreme ofsh ¼ 0 (complete masking), the population evolves tocontain n functional genes, each distinct. Gene conver-sion restores variability by permitting bidirectionaltransitions (e.g., k to k � 1 and k 1 1 mutations). Ychromosomes that are closer to the truncation pointhave a higher probability of transitioning (by mutationor recombination) beyond the truncation point wherethey are removed by selection. Consequently, thepopulation distribution shifts toward fewer mutationsper Y. However, if selection in favor of functionalduplicates is strong relative to the number Y-linkedgenes (sh . 0; n large), most individuals will carry fewmutations, the truncation point becomes irrelevant to Ychromosome evolution, selection shifts toward multi-
plicative epistasis, and gene conversion does notstrongly influence mean fitness or the distribution ofmutations among Y chromosomes. This explanationaccounts for the decreased impact of gene conversionon mutation-free Y chromosomes, as the strength ofselection (sh) increases (Figure 3 and Figure S3).
Muller’s ratchet and the accumulation of nonfunctionalgenes: The deterministic results (presented above) rep-resent an upper limit for Y chromosome fitness. In finitepopulations, where Muller’s ratchet operates, meanfitness can further decrease with each successive loss of‘‘mutation-free’’ individuals. Once lost from the popula-tion, mutation-free genotypes are unlikely to be recov-ered by back mutation or positive selection because theymust initially arise within the current least-loaded classand subsequently avoid stochastic loss (Peck 1994; Orr
and Kim 1998; Gordo and Charlesworth 2000).To explore the influence of gene conversion on the
rate and severity of Y chromosome degeneration viaMuller’s ratchet, we conducted a series of stochasticsimulations, varying the selection and recombinationalparameters (u, h, n, d, c, b). We first use the recursionspresented above to bring the frequencies of eachgenotypic class to deterministic equilibrium. Conver-gence to equilibrium is followed by 100,000 generationsof simulation under a mutation–selection–drift modeland constant male population size. For each generation,genotype frequencies were sampled from a pseudoran-dom multinomial distribution (pseudorandom num-bers generated with R; R Development Core Team
2005), with genotypes randomly sampled after selec-tion, mutation, and recombination.
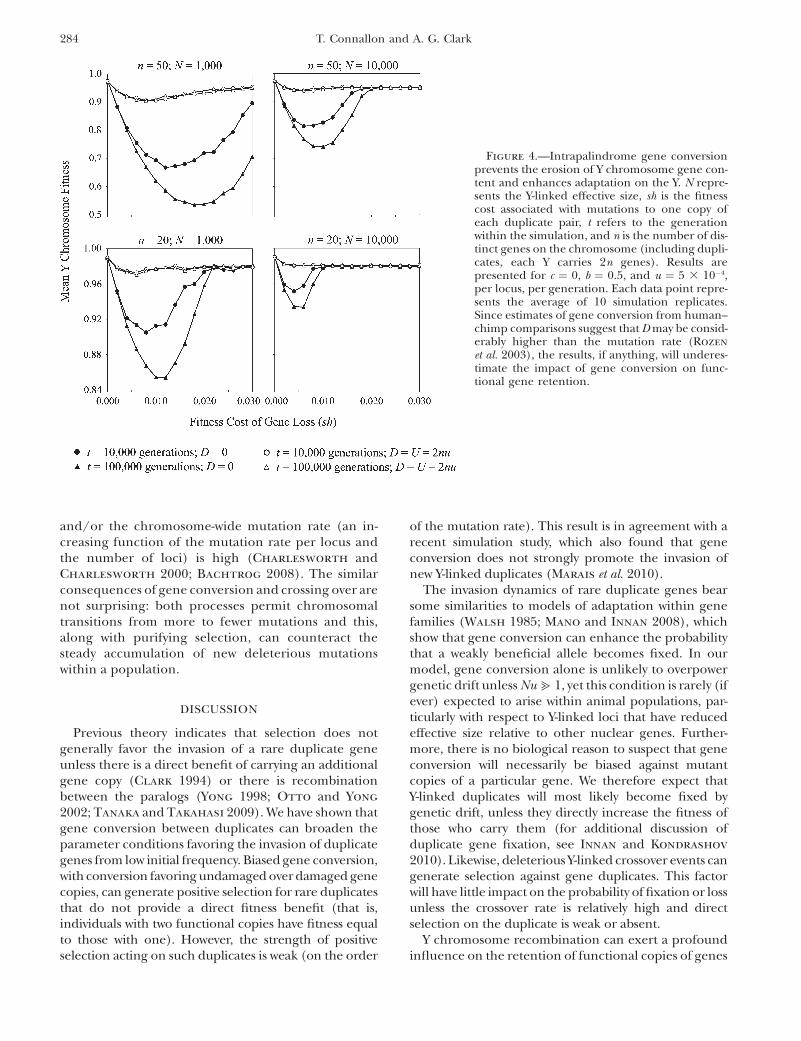
When there is no gene conversion between dupli-cates, Muller’s ratchet can operate rapidly, causingY-linked fitness decay and loss of functional genes.Representative simulation results are shown in Figures4 and 5. In agreement with previous theory (Haigh 1978;Gordo and Charlesworth 2000; Bachtrog 2008), theimpact of the ratchet is strongest when the ancestral Ycarries many functional gene duplicates and whenmutations have small individual fitness effects. Relativelylow rates of gene conversion can rescue Y-linked genesfrom stochastic loss via Muller’s ratchet and therebyincrease mean fitness of the Y (Figures 4 and 5).Increasing the total mutation and gene conversion rateson the Y (U and D, respectively) amplifies the differencesbetween recombining and nonrecombining chromo-somes, whereas a decrease in these compound parame-ters (U, D / 0) eliminates these long-term evolutionarydifferences. This effect occurs both with and withoutbiased gene conversion between duplicates.
Gene conversion appears to constrain accumulationof deleterious mutations in a way that is identical tocrossing over in traditional models of Muller’s ratchet.Under both models, the rate at which the ratchet‘‘clicks’’—the least mutated class of individuals is lost—ishighest when individual mutations are weakly deleterious
and/or the chromosome-wide mutation rate (an in-creasing function of the mutation rate per locus andthe number of loci) is high (Charlesworth andCharlesworth 2000; Bachtrog 2008). The similarconsequences of gene conversion and crossing over arenot surprising: both processes permit chromosomaltransitions from more to fewer mutations and this,along with purifying selection, can counteract thesteady accumulation of new deleterious mutationswithin a population.
DISCUSSION
Previous theory indicates that selection does notgenerally favor the invasion of a rare duplicate geneunless there is a direct benefit of carrying an additionalgene copy (Clark 1994) or there is recombinationbetween the paralogs (Yong 1998; Otto and Yong
2002; Tanaka and Takahasi 2009). We have shown thatgene conversion between duplicates can broaden theparameter conditions favoring the invasion of duplicategenes from low initial frequency. Biased gene conversion,with conversion favoring undamaged over damaged genecopies, can generate positive selection for rare duplicatesthat do not provide a direct fitness benefit (that is,individuals with two functional copies have fitness equalto those with one). However, the strength of positiveselection acting on such duplicates is weak (on the order
of the mutation rate). This result is in agreement with arecent simulation study, which also found that geneconversion does not strongly promote the invasion ofnew Y-linked duplicates (Marais et al. 2010).
The invasion dynamics of rare duplicate genes bearsome similarities to models of adaptation within genefamilies (Walsh 1985; Mano and Innan 2008), whichshow that gene conversion can enhance the probabilitythat a weakly beneficial allele becomes fixed. In ourmodel, gene conversion alone is unlikely to overpowergenetic drift unless Nu ? 1, yet this condition is rarely (ifever) expected to arise within animal populations, par-ticularly with respect to Y-linked loci that have reducedeffective size relative to other nuclear genes. Further-more, there is no biological reason to suspect that geneconversion will necessarily be biased against mutantcopies of a particular gene. We therefore expect thatY-linked duplicates will most likely become fixed bygenetic drift, unless they directly increase the fitness ofthose who carry them (for additional discussion ofduplicate gene fixation, see Innan and Kondrashov
2010). Likewise, deleterious Y-linked crossover events cangenerate selection against gene duplicates. This factorwill have little impact on the probability of fixation or lossunless the crossover rate is relatively high and directselection on the duplicate is weak or absent.
Y chromosome recombination can exert a profoundinfluence on the retention of functional copies of genes
Figure 4.—Intrapalindrome gene conversionprevents the erosion of Y chromosome gene con-tent and enhances adaptation on the Y. N repre-sents the Y-linked effective size, sh is the fitnesscost associated with mutations to one copy ofeach duplicate pair, t refers to the generationwithin the simulation, and n is the number of dis-tinct genes on the chromosome (including dupli-cates, each Y carries 2n genes). Results arepresented for c ¼ 0, b ¼ 0.5, and u ¼ 5 3 10�4,per locus, per generation. Each data point repre-sents the average of 10 simulation replicates.Since estimates of gene conversion from human–chimp comparisons suggest that D may be consid-erably higher than the mutation rate (Rozen
et al. 2003), the results, if anything, will underes-timate the impact of gene conversion on func-tional gene retention.
284 T. Connallon and A. G. Clark
that have already become fixed within the population.Our simulations show that low rates of gene conversionare sufficient to maintain Y-linked genes and counteractdegradation via Muller’s ratchet. These results areconservative, as higher rates enhance the preservationof functional gene copies. Thus, once gene conversionhas evolved, it can potentially provide a degree ofstability on an otherwise evolutionarily unstable Ychromosome. Interestingly, Marais et al. (2010) ob-served that the rate of invasion for gene conversionmodifier alleles does not greatly exceed neutral expect-ations unless they greatly increase the gene conversionrate. This suggests that, while low rates of conversionmay slow the rate of Muller’s ratchet, the evolution ofthe gene conversion rate itself may be much morerestrictive.
The large number of genes within the ‘‘ampliconic’’region of the human Y (Skaletsky et al. 2003) shouldprovide a large target for mutations, creating anopportunity for Muller’s ratchet to act. This role ofgene conversion on the Y is therefore likely to explainpatterns of gene retention on the human Y chromo-some. It is less clear whether similar patterns character-ize other animal species. Current (albeit incomplete)data suggest that gene family amplification and re-tention might be common Y chromosome attributes(Rozen et al. 2003; Verkaar et al. 2004; Murphy et al.2006; Alfoldi 2008; Wilkerson et al. 2008; Krsticevic
et al. 2009), although the prevalence of Y-linked geneconversion outside the human and chimp lineages isless clear (but see Geraldes et al. 2010). Futuresequencing efforts, including evidence for gene conver-sion among Y-linked genes in nonhuman species, willhelp to determine the general relevance of the dupli-cation and gene conversion model presented here.
Within-chromosome crossovers can generate an ab-normal, sterility-inducing Y (Lange et al. 2009) andpotentially represent a deleterious fitness consequenceof Y-linked recombination. This cost also implies that
the number of Y-linked duplicate genes (or in humansthe size of Y-linked palindromes) will have an upperlimit. As the number of Y-linked loci that interact viarecombination increases, so too should the rate ofdeleterious crossovers. This suggests an upper limit toY chromosome gene content, where crossing overbecomes unbearably costly. From this perspective,duplication and recombination represent a costlymechanism of Y chromosome preservation.
In addition to the Y chromosome, our findings haveimplications for asexually reproducing species. Recentreports suggest that the asexual bdelloid rotifers aretetraploid (Mark Welch et al. 2008) and that geneconversion occurs between gene copies (Hur et al. 2008;Mark Welch et al. 2008). Our model supports theverbal claim that gene conversion between homologousgene copies might aid in DNA damage repair andprevent the genomic degradation that is expected toaccompany strict asexual reproduction. Unlike the Ychromosome scenario, crossovers between homolo-gous, tetraploid chromosomes will tend to avoid dele-terious chromosomal aberrations. The relative rate ofnonhomologous crossovers is an empirical questionthat may be difficult to assess, given the likely associationbetween chromosome abnormalities and embryonicdeath, which will lead to a pronounced bias toward‘‘normal’’ chromosomes. On the other hand, crossingover between homologous chromatids is likely togenerate copy number polymorphism, which adds alevel of complexity to the evolutionary dynamics ofautosomal gene duplicates or gene families. This maylead to different evolutionary consequences of crossingover and gene conversion in asexual lineages comparedto the results that we report for the Y chromosome andrepresents an interesting avenue for future theoreticalresearch.
We are grateful to Roman Arguello, Clement Chow, MargaridaCardoso-Moreira, Qixin He, Lacey Knowles, Amanda Larracuente,Rich Meisel, Nadia Singh, and two anonymous reviewers for discussion
Figure 5.—The proportion of loss-of-functionduplicates following 100,000 generations of mu-tation, selection, and genetic drift. Parametersare described in the Figure 4 legend and through-out the text. Results are presented for c ¼ 0, b ¼0.5, u ¼ 5 3 10�4, per locus, per generation, andD on the order of the mutation rate, D¼ U¼ 2nu.Each point represents the average of 10 replicatesimulations.
Gene Conversion and Y Evolution 285
and comments that substantially improved the quality of the manu-script and to Sarah Otto for comments about the eigenvalue-selection-coefficient approximation and for sharing an unpublished manu-script. This work was supported by National Institutes of Health grantGM64590 to A.G.C. and A. B. Carvalho.
LITERATURE CITED
Alfoldi, J. E., 2008 Sequence of mouse Y chromosome. Ph.D. Dis-sertation, MIT, Cambridge, MA.
Bachtrog, D., 2006 A dynamic view of sex chromosome evolution.Curr. Opin. Genet. Dev. 16: 578–585.
Bachtrog, D., 2008 The temporal dynamics of processes underly-ing Y chromosome degeneration. Genetics 179: 1513–1525.
Bengtsson, B. O., 1986 Biased conversion as the primary functionof recombination. Genet. Res. 47: 77–80.
Bengtsson, B. O., 1990 The effect of biased conversion on the mu-tation load. Genet. Res. 55: 183–187.
Carvalho, A. B., L. B. Koerich and A. G. Clark, 2009 Origin andevolution of Y chromosomes: Drosophila tales. Trends Genet. 25:270–277.
Charlesworth, B., 2003 The organization and evolution of the hu-man Y chromosome. Genome Biol. 4: 226.
Charlesworth, B., and D. Charlesworth, 2000 The degenera-tion of Y chromosomes. Philos. Trans. Biol. Sci. 355: 1563–1572.
Clark, A. G., 1994 Invasion and maintenance of a gene duplication.Proc. Natl. Acad. Sci. USA 91: 2950–2954.
Engelstadter, J., 2008 Muller’s ratchet and the degeneration of Ychromosomes: a simulation study. Genetics 180: 957–967.
Geraldes, J., T. Rambo, R. Wing, N. Ferrand and M. W. Nachman,2010 Extensive gene conversion drives the concerted evolutionof paralogous copies of the SRY gene in European rabbits. Mol.Biol. Evol. (in press).
Gordo, I., and B. Charlesworth, 2000 The degeneration of asex-ual haploid populations and the speed of Muller’s ratchet. Genet-ics 154: 1379–1387.
Haigh, J., 1978 Accumulation of deleterious genes in a population—-Muller’s ratchet. Theor. Popul. Biol. 14: 251–267.
Haldane, J. B. S., 1937 The effect of variation on fitness. Am. Nat.71: 337–349.
Heinritz, W., D. Kotzot, S. Heinze, A. Kujat, W. J. Eleemann et al.,2005 Molecular and cytogenetic characterization of a non-mosaic isodicentric Y chromosome in a patient with Klinefeltersyndrome. Am. J. Med. Genet. A 132A: 198–201.
Hughes, J. F., H. Skaletsky, T. Pyntikova, T. A. Graves, S. K. M.van Daalen et al., 2010 Chimpanzee and human Y chromo-somes are remarkably divergent in structure and gene content.Nature 463: 536–539.
Hur, J. H., K. Van Doninck, M. L. Mandigo and M. Meselson,2008 Degenerate tetraploidy was established before bdelloidrotifer families diverged. Mol. Biol. Evol. 26: 375–383.
Innan, H., and F. A. Kondrashov, 2010 The evolution of gene du-plications: classifying and distinguishing between models. Nat.Rev. Genet. 11: 97–108.
Kimura, M., 1957 Some problems of stochastic processes in genet-ics. Ann. Math. Stat. 28: 882–901.
Kimura, M., 1962 On the probability of fixation of mutant genes ina population. Genetics 47: 713–719.
Kimura, M., and T. Maruyama, 1966 Mutational load with epistaticgene interactions in fitness. Genetics 54: 1337–1351.
Koerich, L. B., X. Wang, A. G. Clark and A. B. Carvalho,2008 Low conservation of gene content in the Drosophila Ychromosome. Nature 456: 949–951.
Kondrashov, A. S., 1988 Deleterious mutations and the evolutionof sexual reproduction. Nature 336: 435–440.
Kondrashov, A. S., and J. F. Crow, 1988 King’s formula for the mu-tation load with epistasis. Genetics 120: 853–856.
Krsticevic, F. J., H. L. Santos, S. Januario, C. G. Schrago andA. B. Carvalho, 2009 Functional copies of the Mst77F geneon the Y chromosome of Drosophila melanogaster. Genetics 184:295–307.
Lange, J., H. Skaletsky, S. K. M. van Daalen, S. L. Embry, C. M.Korver et al., 2009 Isodicentric Y chromosomes and sex disor-ders as byproducts of homologous recombination that maintainspalindromes. Cell 138: 855–869.
Mano, S., and H. Innan, 2008 The evolutionary rate of duplicategenes under concerted evolution. Genetics 180: 493–505.
Marais, G., 2003 Biased gene conversion: implications for genomeand sex evolution. Trends Genet. 19: 330–338.
Marais, G., P. R. A. Campos and I. Gordo, 2010 Can intra-Y geneconversion oppose the degeneration of the human Y chromo-some?: A simulation study. Genome Biol. Evol. 2: 347–357.
Mark Welch, D. B., J. L. Mark Welch and M. Meselson,2008 Evidence for degenerate tetraploidy in bdelloid rotifers.Proc. Natl. Acad. Sci. USA 105: 5145–5149.
Murphy, W. J., A. J. P. Wilkerson, T. Raudsepp, R. Agarwala,A. A. Schaffer et al., 2006 Novel gene acquisition on carnivoreY chromosomes. PLoS Genet. 2: e43.
Noordam, M. J., and S. Repping, 2006 The human Y chromosome:a masculine chromosome. Curr. Opin. Genet. Dev. 16: 225–232.
Ohta, T., 1989 The mutational load of a multigene family with uni-form members. Genet. Res. 53: 141–145.
Orr, H. A., and Y. Kim, 1998 An adaptive hypothesis for the evolu-tion of the Y chromosome. Genetics 150: 1693–1698.
Otto, S. P., and D. Bourguet, 1999 Balanced polymorphisms andthe evolution of dominance. Am. Nat. 153: 561–574.
Otto, S. P., and T. Day, 2007 A Biologist’s Guide to Mathematical Mod-eling in Ecology and Evolution. Princeton University Press, Prince-ton, NJ.
Otto, S. P., and D. B. Goldstein, 1992 Recombination and the evo-lution of diploidy. Genetics 131: 745–751.
Otto, S. P., and P. Yong, 2002 The evolution of gene duplicates.Homol. Eff. 46: 451–483.
Peck, J. R., 1994 A ruby in the rubbish: beneficial mutations, delete-rious mutations and the evolution of sex. Genetics 137: 597–606.
R Development Core Team, 2005 R: A Language and Environmentfor Statistical Computing, reference index version 2.2.1. R Founda-tion for Statistical Computing, Vienna.
Repping, S., H. Skaletsky, J. Lange, S. Silber, F. van der Veen et al.,2002 Recombination between palindromes P5 and P1 on thehuman Y chromosome causes massive deletions and spermato-genic failure. Am. J. Hum. Genet. 71: 906–922.
Rice, W. R., 1987 Genetic hitchhiking and the evolution of reducedgenetic activity on the Y sex chromosome. Genetics 116: 161–167.
Rozen, S., H. Skaletsky, J. Lange, S. Silber, F. van der Veen et al.,2003 Abundant gene conversion between arms of palindromesin human and ape Y chromosomes. Nature 423: 873–876.
Skaletsky, H., T. Kuroda-Kawaguchi, P. J. Minx, H. S. Cordum, L.Hillier et al., 2003 The male-specific region of the human Ychromosome is a mosaic of discrete sequence classes. Nature423: 825–837.
Tanaka, K. M., and K. R. Takahasi, 2009 Enhanced fixation andpreservation of a newly arisen duplicate gene by masking delete-rious loss-of-function mutations. Genet. Res. 91: 267–280.
Verkaar, E. L. C., C. Zijlstra, E. M. van ’t Veld, K. Boutaga, D. C.J. Boxtel et al., 2004 Organization and concerted evolutionof the ampliconic Y-chromosomal TSPY genes from cattle.Genomics 84: 468–474.
Walsh, B., 1985 Interaction of selection and biased gene conversionin a multigene family. Proc. Natl. Acad. Sci. USA 82: 153–157.
Wilkerson, A. J. P., F. Raudsepp, T. Graves, D. Albracht, W.Warren et al., 2008 Gene discovery and comparative analysisof X-degenerate genes from the domestic cat Y chromosome.Genomics 92: 329–338.
Yong, P., 1998 Theoretical population genetic model of the inva-sion of an initial duplication. Honours Thesis, Department of Zo-ology, University of British Columbia, Vancouver, BC, Canada.
Yu, Y.-H., Y.-W. Lin, J.-F. Yu, W. Schempp and P. H. Yen,2008 Evolution of the DAZ gene and AZFc region on primateY chromosomes. BMC Evol. Biol. 8: 96.
Gene Duplication, Gene Conversion and the Evolutionof the Y Chromosome
Tim Connallon and Andrew G. Clark
Copyright � 2010 by the Genetics Society of AmericaDOI: 10.1534/genetics.110.116756
T. Connallon and A. G. Clark 2 SI
FILE S1
I. Invasion of gene duplicates on Y chromosomes that carry an arbitrary number of linked genes.
Y-linked duplicate genes evolve within the genetic background of the entire Y chromosome, which is likely to contain
multiple functional genes, particularly during early stages of sex chromosome evolution. To determine the generality of the single
gene duplication scenario in the main text, we developed a second model to examine the evolutionary dynamics of rare, Y-linked
duplicates on ancestral chromosomes carrying an arbitrary number (n) of single-copy genes.
Consider a rare, Y-linked duplicate on Y chromosome carrying n single-copy genes. By duplicating one of the n single-copy
genes, the individual has n – 1 single-copy genes and a single duplicated pair. Though expanding the number of loci greatly
increases the number of possible genotypes to follow within the population, subsequent calculations can be simplified by making
each gene essential. In other words, fitness drops to zero (s = 1) unless each of the n genes has at least one functional copy.
Given this simplification, there are four relevant genotypic classes within the population: (i) individuals with n functional
singletons and no duplicates, each at frequency xn and with fitness wn = 1 – sh; (ii) those with n + 1 functional genes (n – 1
singleton) at frequency xn1 and with fitness wn1 = 1; (iii) those with n + 1 genes (n – 1 singleton), of which n are functional, at
frequency xn0 and with fitness wn0 = 1 – sh; and (4) a class of sterile individuals, at frequency xs and with fitness ws = 1 – s = 0, that
either lack a functional copy of an essential gene, or carry an abnormal Y chromosome.
In an individual carrying n singletons, the Y chromosome deleterious mutation rate per gamete per generation is U = nu, and
the distribution of mutations across gametes is reasonably modeled as a Poisson variable with mean of nu. However, given that the
diploid, genomic deleterious mutation rate is unlikely to be much greater than one, and Y chromosomes typically represent a tiny
fraction of a genome, the number of new mutations should be close to the Bernoulli distribution: U = nu is probability of one
mutation, and 1 – U represents the probability of zero mutations, per generation. For an individual carrying n + 1 total genes, the
overall mutational target will be slightly increased, and the Y chromosome mutation rate becomes Udup = U(n + 1)/n, per
generation. The presence of gene duplicates introduces an opportunity for gene conversion, which as before, are governed by
recombination rate (d), crossover (c), and conversion bias (b) parameters.
Following the events order of (i) birth, (ii) selection, (iii) mutation, (iv) recombination, and (v) fertilization, the Y chromosome
recursions are:
xn1'=xn1[2Ud(1 c)b + (n U Un)(1 dc)]
[xn1 + (xn0 + xn )(1 h)]n+xn0(1 h)(1 U)d(1 c)b
xn1 + (xn0 + xn )(1 h)
xn0 '=2xn1U(1 d)
[xn1 + (xn0 + xn )(1 h)]n+xn0(1 h)(1 U)(1 d)
xn1 + (xn0 + xn )(1 h)
xn '=xn (1 h)(1 U)
xn1 + (xn0 + xn )(1 h)
T. Connallon and A. G. Clark 3 SI
xs '= xn1'+xn0 '+xn '
Stability of the equilibrium xn1 = xn0 = 0, ˆ x n =1 U =1 ˆ x s , and w = (1 U)(1 h) is governed by the eigenvalue:
=2Ud(1 c)b + (n U Un)(1 dc) + (1 h)(1 U)(1 d)n
2(1 h)(1 U)n+
2Ud(1 c)b + (n U Un)(1 dc) + (1 h)(1 U)(1 d)n{ }24(n U Un)(1 dc)(1 d)(1 h)(1 U)n
2(1 h)(1 U)n
When there is no recombination (d = 0), a rare gene duplicate is favored by selection when sh > U/(n – nU). Substituting for
U = nu yields sh > u/(1 – nu). This result differs slightly from the previous model of a duplicate linked to a single essential gene (the
former model predicts that a duplicate invades when sh > u/(1 – u)). Multiple Y-linked genes will therefore decrease opportunities
for positive selection in favor of new duplicates.
When selection is weak (sh 0), recombination can promote selection in favor of the duplicate. For sh = c = 0, the Taylor
series approximation around d = 0 gives a leading eigenvalue of:
=d= 0
+d d= 0
d +O(d2) 1+ d(2b 1)
which is greater than one for b > 0.5, as in the previous model. Numerical simulations of the leading eigenvalue under a broad
range of parameter space show that, as before, the opportunity for positive selection for a new duplicate is greater with
recombination.
T. Connallon and A. G. Clark 4 SI
II. Invasion Probability of Duplicate Genes with Gene Conversion
FIGURE S1.—The probability of fixation for Y-linked duplicate genes. The red line depicts the analytical approximation from
Eq. (2). To facilitate comparison between these results and those of Fig. 2 from the main text, we show the approximation for N =
1000, s = 1, d = 0, and u = 10-5, and present representative simulation results for d > 0 and various combinations of the remaining parameters (c, b). Circles represent the proportion of duplicate genotypes (out of 100,000 replicate simulations for each data point)
that eventually become fixed within the population.
T. Connallon and A. G. Clark 5 SI
III. Maintenance of Functional Gene Duplicates
FIGURE S2.—Gene conversion and the maintenance of functionally redundant paralogs. Results are presented for two
extremes of selection: gene conversion between paralogs of an essential gene (s = 1) and between paralogs of a nonessential gene (s
= 0.001). In each case, gene conversion is unbiased (b = 0.5) and the mutation rate is u = 10-5. Under essentiality and non-
essentiality, fitness is maximized when at least one of the paralog copies is functional (i.e., masking of knockout mutations is
complete: h = 0). Each point represents the fraction of 100 simulation replicates where both copies are maintained as functional
within the population. For each simulation run, the population is initially fixed for two functional Y-linked genes, and then
evolves under mutation, recombination, selection, and genetic drift for 100,000 generations.
T. Connallon and A. G. Clark 6 SI
IV. Frequency of the ‘least loaded class’ under biased gene conversion.
FIGURE S3.—Gene conversion increases the frequency of Y chromosomes haplotypes that carry zero deleterious mutations
(i.e., the “least-loaded” genotypic class). Results use the same parameters as those of Fig. 3 with n = 50, and with the biased gene