Genetic differentiation across the Andes in twopan-Neotropical tyrant-flycatcher species
Frank E. Rheindt A,B,D, Janette A. NormanA,B and Les ChristidisA,C
AGenetics Department, University of Melbourne, Melbourne, Vic. 3000, Australia.BSciences Department, Museum Victoria, 11 Nicholson St, Carlton, Vic. 3053, Australia.CAustralian Museum, Research and Collections, 6 College Street, Sydney, NSW 2010, Australia.DCorresponding author. Email: [email protected]
Abstract. Vocal and phylogenetic research has revealed much hidden species-level diversity in tyrant-flycatchers (Tyrannidae), and field ornithological accounts indicate there is a great deal of additional diversity to be uncovered. Using mitochondrial and nuclear DNA sequences, we screened genetic diversity from across the range of two pan-Neotropical species of elaeniine flycatcher – Camptostoma obsoletum (Southern Beardless Tyrannulet) and Phaeomyias murina (Mouse-coloured Tyrannulet) – for which vocal discontinuities have previously been reported. Our results indicate deep genetic divergences across the Andes in both species, and support a previously proposed separation of trans-Andean populations of P. murina into P. tumbezana. Within C. obsoletum, we additionally uncover a deep genetic break between a trans-Andean clade from dry Tumbesia and one from humid Chocó–Panama, and we fail to detect mitochondrial DNA differentiation in western Amazonian C. o. olivaceum, which has been treated as a distinct form based on differences in voice and plumage from surrounding subspecies. Further molecular and vocal sampling is necessary to confirm the division of C. obsoletum into three species.
Introduction
With ~450 currently recognised species in 100 genera, tyrant- (Mouse-coloured Tyrannulet). The latter forms a monospecific flycatchers (Tyrannidae) make up a disproportionately large part genus, while the former is member of a genus of two species that of the diverse Neotropical avifauna (Fitzpatrick 2004). used to be lumped into a single species (Fitzpatrick 2004). One However, the generally subtle geographical differentiation in commonality between the two groups is that field workers have their inconspicuous plumage patterns has obscured insights into observed similar vocal discontinuities in these two species, with their species-level taxonomy. stark differences in call notes between cis-Andean (east of the
In recent years, vocalisations have been shown to be a much Andes) and trans-Andean (west of the Andes) populations better indicator of species boundaries than plumage patterns in (Ridgely and Tudor 1994; Ridgely and Greenfield 2001). Both many inconspicuously coloured suboscine passerines such as species inhabit the same type of dry to deciduous scrub habitat tyrant-flycatchers (Alström and Ranft 2003; Rheindt et al. and the edge of humid forest, and they share a similar incon-2008a). Advances in our knowledge of the vocalisations of spicuous brownish to olive coloration with light underparts that tyrant-flycatchers have seen the description of new species tend to have a yellow tinge in more humid parts of their distri-(Coopmans and Krabbe 2000; Álvarez Alonso and Whitney bution. Their current subspecific taxonomy is mainly based on 2001; Johnson and Jones 2001; Zimmer et al. 2001) and the ele- plumage comparisons of large series of specimens from most vation of vocally distinct subspecies to species level (Reynard parts of their ranges (e.g. Zimmer 1941). The similarities in et al. 1993; Zimmer and Whittaker 2000; Leger and Mountjoy habitat preference as well as in vocal and plumage discontinu-2003). Similarly, molecular studies revealed several examples ities observed in these two species make them an interesting of paraphyletic and polyphyletic arrangements necessitating the object for a screening of the distribution of genetic diversity. elevation of some races to species level (García-Moreno et al. Currently, seven subspecies are widely accepted for 1998; Chesser 2000; Joseph et al. 2003a, 2003b; Joseph and Phaeomyias murina, although Ridgely and Greenfield (2001) Wilke 2004; Rheindt et al. 2008a). There are further indications separated three trans-Andean races (tumbezana, inflava and of vocal discontinuities in a high proportion of currently recog- maranonica) from western Ecuador and Peru (but not nised species of tyrant-flycatcher, which are suggestive of addi- eremonoma from Panama) into P. tumbezana (Tumbes tional cryptic species diversity (Ridgely and Tudor 1994; Tyrannulet) on account of differing calls and plumage. Within Ridgely and Greenfield 2001; Fitzpatrick 2004). the three cis-Andean races (wagae, incompta and murina),
In this study, we examine cryptic genetic diversity in two vocalisations are reported to be homogeneous and no further widely distributed (Fig. 1) members of the Neotropical Elaeniini species divisions have been proposed. However, Zimmer (1941) (Lanyon 1988; Rheindt et al. 2008b), Camptostoma obsoletum documented an interesting pattern of plumage variation in (Southern Beardless Tyrannulet) and Phaeomyias murina which the more yellowish Amazonian race wagae, which ranges
from Bolivia through the Guianas, forms a geographical divi-sion between the drab races murina from south of Amazonia and incompta from Venezuela and northern Colombia.
The genus Camptostoma ranges from Argentina in the south to Arizona in the north, in ~14 currently recognized taxa (Fitzpatrick 2004). Central American populations from Costa Rica through to Arizona are usually thought to constitute a separate monotypic species, C. imberbe (Northern Beardless Tyrannulet), mainly on account of differences in call notes (Fitzpatrick 2004). However, Ridgely and Tudor (1994) pointed out that vocal differences between some races within C. obsole-tum sensu stricto (Southern Beardless Tyrannulet) are more pro-nounced than those between C. imberbe and the neighbouring taxon C. obsoletum flaviventre. Similarly, Fitzpatrick (2004) divided the 13 remaining races of the taxonomically restricted southern C. obsoletum into five geographically and morpho-logically discrete groups, some of which may merit species status: the flaviventre group from Central America, the northern South American pusillum group, the sclateri group from Tumbesia and the Marañón Valley, olivaceum from Western Amazonia, and the nominate group from most of Brazil south to
Fig. 1. Neotropical distribution and sampling localities of Phaeomyias murina (range: striped, sampling localities: circles) and Camptostoma obso-letum (range: gray, sampling localities: stars).
Argentina. Among these assemblages, the trans-Andean sclateri group has been singled out as differing vocally from cis-Andean races (Ridgely and Greenfield 2001), while Fitzpatrick (2004) emphasised the great morphological and vocal divergence of western Amazonian olivaceum towards surrounding subspecies.
We utilised mitochondrial and nuclear DNA sequences of several samples covering most parts of the ranges of both species to screen for patterns of genetic diversity. Within Camptostoma obsoletum, our sampling covered all five taxon groups as proposed by Fitzpatrick (2004), while our coverage of Phaeomyias murina included both trans-Andean and cis-Andean populations. Even though our geographical coverage of both species is wide, our sample size per locality is generally low. Therefore, this work is intended to be a first genetic screen-ing to highlight potential areas of cryptic diversity in these groups. A complete elucidation of phylogenetic patterns in the two species will have to await future studies.
Methods
Sampling regime, laboratory techniques and alignment
We sequenced the mitochondrial coding gene region, NADH dehydrogenase subunit 2 (including flanking bits of tRNA-Met; this partition henceforth called ND2), for five individuals each of Phaeomyias murina and Camptostoma obsoletum. An addi-tional one and two samples of each species, respectively, were also available from Rheindt et al. (2008b). We sequenced the nuclear intron, β-fibrinogen intron 5 (Fib5), for three indi-viduals each of P. murina and C. obsoletum. Two additional sequences for each species were available from Rheindt et al. (2008b). The genetic screening regime covered the largest parts of the range for both species (Table 1, Fig. 1).
For Camptostoma, we used Ornithion inerme as an outgroup, since Ornithion probably constitutes the sister genus (GenBank accession numbers: EF501915, EF501841; Rheindt et al. 2008b). For Phaeomyias, we utilised Capsiempis flaveola as an outgroup (GenBank accession numbers: DQ294563, DQ294475; Tello and Bates 2007); even though Capsiempis is possibly not the sister genus to Phaeomyias, it is closely related (Rheindt et al. 2008b).
Extraction and sequence generation followed Rheindt et al. (2008b). We aligned and edited sequences using the program SEQUENCHER v. 4.1.4 (Gene Codes Corp., Ann Arbor, MI). Heterozygous sites in Fib5 sequences were interpreted as ambi-guities. All sequences were double-checked by eye. ND2 sequences were translated and checked for stop codons, anoma-lous substitution patterns and deviant base composition.
Phylogenetic analysis
Phylogenetic analysis was carried out using maximum parsi-mony (MP) and maximum likelihood (ML) as implemented in the program PAUP* (Swofford 2002), with indels excised before analysis. Default settings were activated for heuristic searches. MP analysis was unweighted since no saturation was detected by plotting pairwise divergences against the number of transitions/transversions (not shown). For ML analysis, we selected a suitable evolutionary model (Table 2) for each parti-tion and for the concatenated dataset by applying the hierarchi-cal likelihood ratio test as implemented in the program
Genetic differentiation in Neotropical flycatchers Emu 263
MODELTEST 3.06 (Posada and Crandall 1998) to the align-ment containing all sampled individuals of both ingroup species. Partition congruence was evaluated using the partition homogeneity test in PAUP*. We ran 100 bootstrap replicates for ML analyses and 1000 replicates for all other analyses and tests. PAUP* was also used to estimate partition parameters.
Results
Genetic characterisation
The alignment of the ND2 partition did not exhibit any anoma-lies and comprised 1088 base pairs (bp), of which 367 bp (33.7%) were variable and 260 bp (23.9%) were parsimony-informative. In Fib5, we found three indels, none of which was parsimony-informative: a 5 bp gap in Capsiempis flaveola with respect to the ingroup Phaeomyias, an 8 bp gap in the sample of Camptostoma obsoletum flaviventre from Panama and a 1 bp insertion in the sample of C. obsoletum griseum from Lambayeque (Peru). Despite the indels, alignment was straight-forward, with sequences totalling 586 bp (not counting indels). Of these, 74 bp (12.6%) were variable and 25 bp (4.3%) were parsimony-informative. In concert with other studies of tyran-nids using the same two gene regions for species and genus-
level phylogenetics (Tello and Bates 2007; Rheindt et al. 2008a, 2008b), the phylogenetic utility of Fib5 was greatly reduced in comparison to ND2.
χ2-tests of homogeneity of base frequencies across taxa did not show any evidence for base compositional bias in either par-tition (not shown). The partition homogeneity test failed to reveal statistically significant incongruence between the two data partitions (P = 0.068). Pairwise uncorrected divergences between samples for ND2 and Fib5 are listed in Table 3.
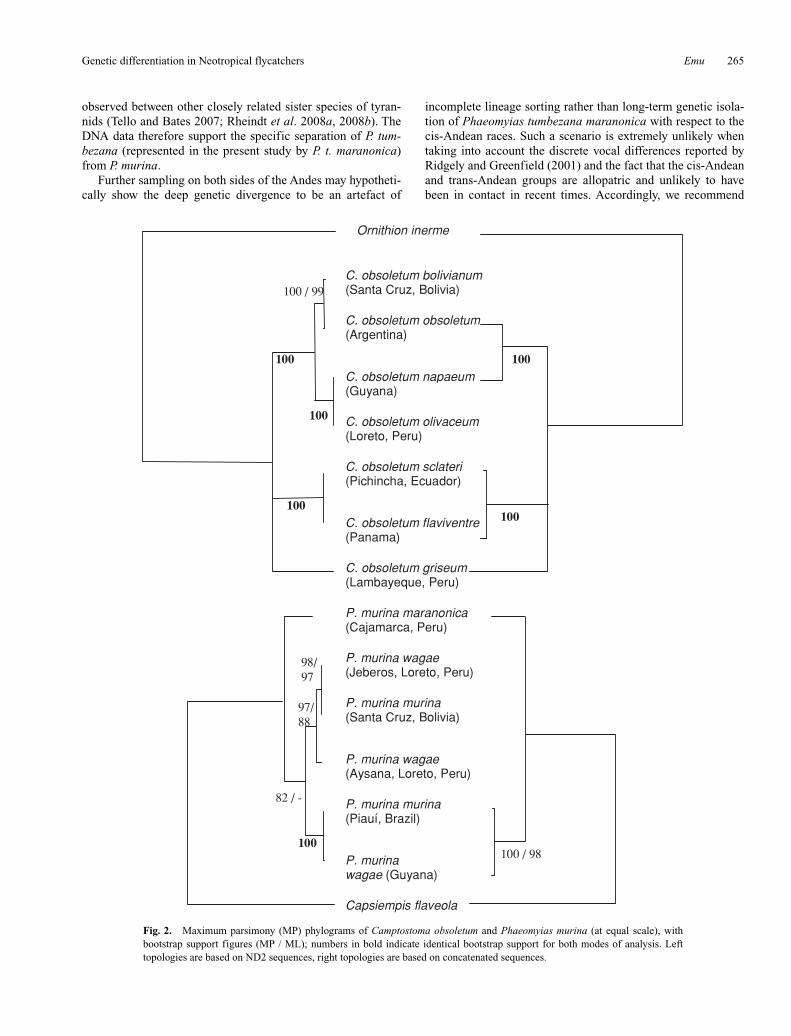
Phylogenetics of Phaeomyias murina MP analysis of the ND2 partition placed the trans-Andean Phaeomyias murina maranonica as sister to a monophyletic cis-Andean clade with moderate bootstrap support (Fig. 2, left). Within the cis-Andean clade, the west-Amazonian and east-Amazonian samples each grouped together across subspecific boundaries with high support, with both P. murina murina and P. murina wagae being represented in both groups (Fig. 2, left). The results of the ML analysis were congruent, but there was less support for the monophyly of the cis-Andean clade (Fig. 2, left).
In accordance with its lower phylogenetic utility, the Fib5 par-tition produced an unresolved topology (not shown). The only node that received somewhat moderate Fib5 bootstrap support of
Table 1. Samples used in this study, including tissue identification numbers and housing institutions, GenBank Accession numbers and collection locality
Abbreviations: LSU, Natural History Museum of Louisiana State University (Baton Rouge, LA, USA); LGEMA, Laboratório de Genética Evolutiva e Molecular de Aves (Universidade de São Paulo, Brazil); USNM, National Museum of Natural History, Washington, DC; N, north; S, south; E, east; W, west
(and standard intermediates)
Institution and GenBank Accession Taxon name Collection locality Reference tissue identification numbers number ND2 Fib5
LSU B7328 EU330880 – Phaeomyias murina Peru: Loreto, Isla Pasto, Rio Amazonas this study wagae opposite Aysana, 80 km NE Iquitos
LSU B6693 EU330877 – Phaeomyias murina Bolivia: Sta Cruz, 1 km N 2km E YPFB this study murina Refinery
LGEMA P2180 EU330871 EU330885 Phaeomyias murina Brazil: Piauí, Serra das Confusões this study murina
LGEMA P2179 – EU330883 Phaeomyias murina Brazil: Piauí, Serra das Confusões this study murina
264 Emu F. E. Rheindt et al.
Table 2. Model parameters used for each partition and for the concatenated dataset Abbreviations: TrN is the Tamura–Nei model (Tamura and Nei 1993); HKY is the Hasegawa–Kishino–Yano model
(Hasegawa et al. 1985); +G stands for rate variation among sites
ND2 Fib5 Combined
Evolutionary model TrN + G HKY + G HKY + G Base frequencies (A, C, G, T) 0.31, 0.31, 0.09, 0.29 0.30, 0.17, 0.23, 0.30 0.31, 0.26, 0.14, 0.29 Transition/transversion ratio n.a. 1.5348 4.1249 Gamma shape parameter 0.2704 0.1952 0.2610 Proportion of invariable sites 0 0 0
88 and 83 in MP and ML analyses, respectively, is along a branch uniting the trans-Andean sample of Phaeomyias murina mara-nonica with the Guyanan sample of P. murina wagae. Since no statistical incongruence was detected between data partitions, we concatenated alignments for those sequences in which both par-titions were available to increase phylogenetic signal. The result-ing tree yielded high support for a monophyletic cis-Andean group in both MP and ML analyses (Fig. 2, right).
Phylogenetics of Camptostoma obsoletum In both ML and MP analyses, the ND2 partition produced a polytomous and deep division into three highly supported clades (Fig. 2, left): a cis-Andean clade, a trans-Andean clade including samples from humid regions (Chocó, Panama) and a trans-Andean clade including a sample from dry regions (Tumbesia). Within the cis-Andean clade, there was a further subdivision into two highly-supported groups: an Amazonian clade (including the samples from Loreto, in Peru, and Guyana) and a clade centred in the Chaco zone south of Amazonia (including the samples from Argentina and southern Bolivia).
Fib5 analysis using both ML and MP yielded a mostly unre-solved topology, with strong bootstrap support limited to the branch uniting the two humid trans-Andean samples from Panama and northern Ecuador (MP 91; ML 86). Analysis of the concatenated dataset was entirely congruent with the ND2-based phylogeny (Fig. 2, right).
Discussion
Phylogenetic patterns in Phaeomyias murina
Ridgely and Greenfield (2001) separated Phaeomyias tum-bezana (incl. races maranonica and inflava) from western Peru and Ecuador as a different species based on its vocal and plumage distinction from cis-Andean taxa. The only trans-Andean race we sampled is P. t. maranonica, for which our analyses indicate a sister relationship with a monophyletic cis-Andean clade (Fig. 2), albeit with moderate branch support. Mitochondrial (ND2) sequence divergence between P. t. mara-nonica and cis-Andean samples ranges between 4.5% and 5.0% (Table 3). This level of divergence is comparable to that
Table 3. ND2 and Fib5 sequence divergences between samples
(P. m. maranonica ↔ other Phaeomyias) Western Amazonian ↔ eastern Amazonian 2.3–2.4 –
(Loreto and Bolivian samples ↔ Guyanan and Brazilian samples) Within western-Amazonian group 0.0–0.8 –
(within Loreto, and Loreto ↔ Bolivia) Within eastern-Amazonian group 0.1 0.0–0.9
(within Brazil, and Guyana ↔ Brazil) Comparisons involving Camptostoma obsoletum
Camptostoma ↔ outgroup (Ornithion) 10.0–11.6 3.2–5.9 Cis-Andean ↔ trans-Andean 6.9–8.4 1.9–4.9 (C. o. griseum–sclateri–flaviventre ↔ all other Camptostoma sampled)Humid group ↔ arid group (west of Andes) 7.7 4.0–7.5
(Panama and Pichincha, Ecuador ↔ Lambayeque, Peru) Within-humid group (west of Andes) 0.0 1.5
(Loreto/Guyana ↔ Bolivia–Argentina) Within-Amazonian clade (east of Andes) 0.0 –
(Loreto ↔ Guyana) Within-Chaco clade (east of Andes) 0.1 –
(Argentina ↔ Bolivia)
Genetic differentiation in Neotropical flycatchers Emu 265
observed between other closely related sister species of tyran- incomplete lineage sorting rather than long-term genetic isola-nids (Tello and Bates 2007; Rheindt et al. 2008a, 2008b). The tion of Phaeomyias tumbezana maranonica with respect to the DNA data therefore support the specific separation of P. tum- cis-Andean races. Such a scenario is extremely unlikely when bezana (represented in the present study by P. t. maranonica) taking into account the discrete vocal differences reported by from P. murina. Ridgely and Greenfield (2001) and the fact that the cis-Andean
Further sampling on both sides of the Andes may hypotheti- and trans-Andean groups are allopatric and unlikely to have cally show the deep genetic divergence to be an artefact of been in contact in recent times. Accordingly, we recommend
100 / 99
100
100
100
98/ 97
82 / -
97/ 88
100
Ornithion inerme
C. obsoletum bolivianum (Santa Cruz, Bolivia)
C. obsoletum obsoletum (Argentina)
C. obsoletum napaeum (Guyana)
C. obsoletum olivaceum (Loreto, Peru)
C. obsoletum sclateri (Pichincha, Ecuador)
C. obsoletum flaviventre (Panama)
C. obsoletum griseum (Lambayeque, Peru)
P. murina maranonica (Cajamarca, Peru)
P. murina wagae (Jeberos, Loreto, Peru)
P. murina murina (Santa Cruz, Bolivia)
P. murina wagae (Aysana, Loreto, Peru)
P. murina murina (Piauí, Brazil)
P. murina wagae (Guyana)
Capsiempis flaveola
100
100
100 / 98
Fig. 2. Maximum parsimony (MP) phylograms of Camptostoma obsoletum and Phaeomyias murina (at equal scale), with bootstrap support figures (MP / ML); numbers in bold indicate identical bootstrap support for both modes of analysis. Left topologies are based on ND2 sequences, right topologies are based on concatenated sequences.
266 Emu F. E. Rheindt et al.
biological species status for P. tumbezana in agreement with Ridgely and Greenfield (2001). We note, however, that the fourth trans-Andean subspecies eremonoma from Panama has not been considered by Ridgely and Greenfield (2001) or in the present study. Taking into account the close avifaunal ties between Panama and the Chocó Region in Ecuador and Colombia (Brumfield and Capparella 1996), eremonoma may well be part of P. tumbezana. However, P. tumbezana is centred south of the Chocó in the dry Tumbesian Region of Ecuador and Peru. Moreover, eremonoma more closely resembles the cis-Andean clade in plumage (Fitzpatrick 2004). The resolution of its position must therefore await further vocal and molecular analysis.
Within the cis-Andean clade, our sampling included speci-mens of the bright Amazonian race Phaeomyias murina wagae and the drabber nominate form, whose distribution is centred south of Amazonia. The ND2 partition showed the cis-Andean clade to be divided into two highly supported sub-clades (Fig. 2) of 2.3–2.4% divergence (Table 3), though interestingly this divi-sion was not along taxonomic or plumage boundaries, but between the west and the east of the cis-Andean lowlands. Small sample size precludes a taxonomic revision, since haplotypes of the two sub-clades may be represented in P. m. wagae and nom-inate P. m. murina in different frequencies, as a result of incom-plete lineage sorting.
Phylogenetic patterns in Camptostoma obsoletum Although there have not been any formal proposals to split Camptostoma obsoletum into two or more species, vocal and plumage discontinuities within the range of the species have led to suggestions that some of the five morphological taxon groups within C. obsoletum may merit species rank (Fitzpatrick 2004). Our analyses included representatives of all five of these taxon groups and demonstrated a division into three deep and highly supported genetic lineages (Fig. 2), each separated by 6.9–8.4% ND2 divergence (Table 3): one cis-Andean clade, one humid trans-Andean clade (Chocó and Panama) and one arid trans-Andean clade (Tumbesia). These levels are on the high end of those recorded for congeneric species level divergences in tyrannids (Tello and Bates 2007; Rheindt et al. 2008a, 2008b).
The deep genetic break between the cis-Andean and trans-Andean taxon groups is analogous to the situation in Phaeomyias in that it is corroborated by vocal differences that have been documented between trans-Andean Camptostoma obsoletum sclateri and cis-Andean C. o. olivaceum (Ridgely and Greenfield 2001). More surprising is the break between the two trans-Andean groups, which display a deep genetic divergence not only in ND2 but also in the nuclear Fib5 partition (Table 3). This congruence between nuclear and mitochondrial divergence between the two clades is coupled with the absence of mito-chondrial divergence within the humid trans-Andean clade (Table 3) across a distance of 900 km (Fig. 1). Both lines of evi-dence combine to indicate that the genetic break between the Chocó–Panama and the Tumbesian clade is not a result of incomplete lineage sorting but reflects long-term reproductive isolation. Using a greatly enlarged dataset, including samples from the presumed contact zone, future researchers may be able to address the question of on-going gene flow versus isolation between the humid and dry trans-Andean clades by utilising
such recently developed methodologies as IM analysis (Nielsen and Wakeley 2001; Hey and Nielsen 2004; Hey 2005).
The division between the wet Panama–Chocó Camptostoma obsoletum clade and the dry Tumbesian Camptostoma obsole-tum clade in our sampling regime is in conflict with Fitzpatrick’s (2004) circumscription of the Central American flaviventre group (consisting of flaviventre, orphnum and majus) and the west Peruvian – Ecuadorian sclateri group (consisting of sclateri, maranonicum and griseum). Our analyses reveal zero mitochondrial divergence between a sclateri sample from Pichincha (Ecuador) and a Panamanian flaviventre sample (Table 3, Fig. 2). In contrast, our griseum sample from western Peru is highly differentiated from Ecuadorian sclateri (Table 3) and emerges in a separate clade (Fig. 2).
It may be concluded that Fitzpatrick’s (2004) assignment of trans-Andean races into taxon groups could have been erro-neous, and that the boundary should have been drawn between western Ecuadorian Camptostoma obsoletum sclateri and western Peruvian Camptostoma obsoletum griseum. However, in his analysis of a large series of specimens, Zimmer (1941) reported on a broad zone of intermediate plumage between these two races in north-western Peru. Additionally, the vocali-sations of these two races appear identical in the field (F. E. Rheindt, pers. obs.). Therefore, we suggest that Fitzpatrick’s (2004) assignment of griseum and sclateri into the same taxon group is likely correct, but that our sample from extreme northern Ecuador may belong to caucae from the adja-cent Colombian Chocó. Fitzpatrick (2004) assigned trans-Andean C. obsoletum caucae to the cis-Andean C. obsoletum pusillum group, while restricting the C. obsoletum flaviventre group to Central America. However, our results indicate that our northern Ecuadorian sample may mark the southernmost exten-sion of the flaviventre group, which should henceforth include caucae. To study the contact zone of the two trans-Andean taxon groups and assess their reproductive isolation, it is necessary to conduct increased molecular sampling and field studies in northern Ecuador near where dry Tumbesian woodland shifts into humid Chocó forest.
Within the cis-Andean clade, genetic structure was shallow. Nevertheless, individuals grouped into two highly supported sub-clades (Fig. 2) that were differentiated by 1.9–2.0% ND2 divergence (Table 3): (1) an Amazonian clade comprising the Camptostoma obsoletum pusillum group (one napaeum sample from Guyana) and the C. obsoletum olivaceum group (one olivaceum sample from Peru), and (2) a clade centred in the Chaco zone south of Amazonia, comprising the nominate group (one bolivianum sample from Bolivia and one obsoletum sample from Argentina). Although western Amazonian C. obso-letum olivaceum is distinctive in plumage pattern (Zimmer 1941; Fitzpatrick 2004), there was no mitochondrial sequence divergence between it and C. obsoletum napaeum. Fitzpatrick (2004) suggested that olivaceum is vocally distinct from sur-rounding races and possibly represents a separate species. Additional data on vocalisations are needed to confirm the pur-ported acoustic differentiation of olivaceum, in addition to nuclear DNA data to assess whether the lack of mitochondrial divergence may be a result of introgression.
In summary, our results indicate that Camptostoma obsole-tum likely comprises at least three species. Additional species-
Genetic differentiation in Neotropical flycatchers Emu 267
level lineages may have been overlooked by our sampling regime or, in the case of olivaceum, by the absence of data from more than one marker. On the other hand, the deep genetic divi-sions between the three C. obsoletum clades (Fig. 2) are unlikely to be an artefact of incomplete lineage sorting. Nevertheless, before formally proposing the elevation of trans-Andean C. flaviventre and C. sclateri to species level, we would like to wait for additional molecular sampling or the documentation of consistent morphological and vocal differences between them, or both datasets, preferably accompanied by studies of their contact zone in northern Ecuador. Future molecular work should also aim to include northern C. imberbe into the frame-work of a Camptostoma phylogeny, as it may well be nested within C. obsoletum (Ridgely and Tudor 1994).
Comparison of the distribution of genetic breaks in Camptostoma and Phaeomyias
As expected from accounts of vocal discontinuities (Ridgely and Tudor 1994; Ridgely and Greenfield 2001), both Phaeomyias murina and Camptostoma obsoletum share a deep genetic divergence between cis-Andean and trans-Andean sub-species (Table 3). These divergences are comparable to or exceed genetic divergences between species-level taxa in other tyrannid flycatchers (Tello and Bates 2007; Rheindt et al. 2008a, 2008b) and, coupled with vocal data, suggest biological species status of the taxon groups on either side of the Andes. In the case of Camptostoma obsoletum, however, we found an additional and equally deep genetic break (in both mitochon-drial and nuclear DNA) between subspecies from humid and dry regions west of the Andes, but we lack samples from the area where these two taxon groups meet. Therefore, we refrain from making taxonomic proposals for Camptostoma on the basis of this study and encourage phylogeneticists to address these issues in future studies using more samples. The distribution and depth of trans-Andean genetic breaks in Camptostoma and Phaeomyias may not be as dissimilar as indicated by the present data: Phaeomyias drops out in the wet Chocó lowlands north of the dry Tumbesian zone, but the presently unsampled sub-species eremonoma occurs in Panama. Unlike Camptostoma, trans-Andean races of Phaeomyias from humid and dry regions do not come into contact. However, future studies may well show that Phaeomyias displays a similar threefold division into a Tumbesian, Chocó and cis-Andean clade, each of which may deserve species status under the biological species concept.
Acknowledgements We would like to thank the following people and institutions for the provi-sion of tissue samples for this study: Donna Dittmann, Van Remsen and Robb Brumfield (Louisiana State University Museum of Natural Sciences, Baton Rouge, LA) as well as Cristina Y. Miyaki and G. Sebastián Cabanne (Laboratório de Genética Evolutiva e Molecular de Aves, Universidade de São Paulo, Brazil). Two anonymous referees are acknowledged for helping improve the manuscript. This work was undertaken at the Population and Evolutionary Genetics Unit, Museum Victoria, which was established with the generous support of the Ian Potter Foundation and Amersham Biosciences (now GE Healthcare). The first author was supported by the fol-lowing grants: Joseph Grinnell Student Research Award 2004 awarded by the Cooper Ornithological Society; Sigma Xi Grant-in-Aid of Research 2004; Systematics Research Fund 2006 awarded by the Linnean Society;
Museum Victoria 1854 Student Scholarship; and University of Melbourne IPRS/MIRS Scholarships.
References Alström, P., and Ranft, R. (2003). The use of sounds in bird systematics, and
the importance of bird sound archives. Bulletin of the British Ornithologists’ Club 123A, 114–135.
Álvarez Alonso, J., and Whitney, B. M. (2001). A new Zimmerius tyrannulet (Aves:Tyrannidae) from white-sand forests of northern Amazonian Peru. Wilson Bulletin 113, 1–9. doi:10.1676/0043-5643(2001)113 [0001:ANZTAT]2.0.CO;2
Brumfield, R. T., and Capparella, A. P. (1996). Historical diversification of birds in northwestern South America: a molecular perspective on the role of vicariant events. Evolution 50, 1607–1624. doi:10.2307/2410897
Chesser, R. T. (2000). Evolution in the high Andes: the phylogenetics of Muscisaxicola ground-tyrants. Molecular Phylogenetics and Evolution 15, 369–380. doi:10.1006/mpev.1999.0774
Coopmans, P., and Krabbe, N. (2000). A new species of flycatcher (Tyrannidae: Myiopagis) from eastern Ecuador and eastern Peru. Wilson Bulletin 112, 305–312. doi:10.1676/0043-5643(2000)112[0305: ANSOFT]2.0.CO;2
Fitzpatrick, J. W. (2004). Family Tyrannidae (Tyrant-flycatchers) and Genera Camptostoma and Phaeomyias. In ‘Handbook of the Birds of the World. Vol. 9: Cotingas to Pipits and Wagtails.’ (Eds J. del Hoyo, A. Elliott and D.A. Christie.) pp. 170–257 and 270–287. (Lynx Edicions: Barcelona.)
García-Moreno, J., Arctander, P., and Fjeldså, J. (1998). Pre-Pleistocene dif-ferentiation among chat-tyrants. Condor 100, 629–640. doi:10.2307/ 1369744
Hasegawa, M., Kishino, H., and Yano, T. (1985). Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. Journal of Molecular Evolution 22, 160–174. doi:10.1007/BF02101694
Hey, J. (2005). On the number of New World founders: a population genetic portrait of the peopling of the Americas. PLoS Biology 3, e193. doi:10.1371/journal.pbio.0030193
Hey, J., and Nielsen, R. (2004). Multilocus methods for estimating popu-lation sizes, migration rates and divergence time, with applications to the divergence of Drosophila pseudoobscura and D. persimilis. Genetics 167, 747–760. doi:10.1534/genetics.103.024182
Johnson, N. K., and Jones, R. E. (2001). A new species of tody-tyrant (Tyrannidae:Poecilotriccus) from northern Peru. Auk 118, 334–341. doi:10.1642/0004-8038(2001)118[0334:ANSOTT]2.0.CO;2
Joseph, L., and Wilke, T. (2004). When DNA throws a spanner in the taxo-nomic works: testing for monophyly in the Dusky-capped Flycatcher, Myiarchus tuberculifer, and its South American subspecies, M. t. atri-ceps. Emu 104, 197–204. doi:10.1071/MU03047
Joseph, L., Wilke, T., Bermingham, E., Alpers, D., and Ricklefs, R. (2003a). Towards a phylogenetic framework for the evolution of shakes, rattles and rolls in Myiarchus tyrant-flycatchers (Aves:Passeriformes: Tyrannidae). Molecular Phylogenetics and Evolution 31, 139–152. doi:10.1016/S1055-7903(03)00259-8
Joseph, L., Wilke, T., and Alpers, D. (2003b). Independent evolution of migration on the South American landscape in a long-distance temper-ate-tropical migratory bird, Swainson’s flycatcher (Myiarchus swain-soni). Journal of Biogeography 30, 925–937. doi:10.1046/j.1365-2699. 2003.00841.x
Lanyon, W. E. (1988). A phylogeny of the thirty-two genera in the Elaenia assemblage of tyrant-flycatchers. American Museum Novitates 2914, 1–57.
Leger, D. W., and Mountjoy, D. J. (2003). Geographic variation in song of the Bright-rumped Attila (Tyrannidae:Attila spadiceus): implications for species status. Auk 120, 69–74. doi:10.1642/0004-8038(2003)120 [0069:GVISOT]2.0.CO;2
Nielsen, R., and Wakeley, J. (2001). Distinguishing migration from isola-tion: a Markov chain Monte Carlo approach. Genetics 158, 885–896.
268 Emu
Posada, D., and Crandall, K. A. (1998). MODELTEST: testing the model of DNA substitution. Bioinformatics 14, 817–818. doi:10.1093/ bioinformatics/14.9.817
Reynard, G. B., Garrido, O. H., and Sutton, R. L. (1993). Taxonomic revi-sion of the Greater Antillean Pewee. Wilson Bulletin 105, 217–226.
Rheindt, F. E., Norman, J. A., and Christidis, L. (2008a). DNA evidence shows vocalizations to be a better indicator of taxonomic limits than plumage patterns in Zimmerius tyrant-flycatchers. Molecular Phylogenetics and Evolution 48, 150–156. doi:10.1016/j.ympev. 2008.04.016
Rheindt, F. E., Norman, J. A., and Christidis, L. (2008b). Phylogenetic rela-tionships of tyrant-flycatchers (Aves:Tyrannidae), with an emphasis on the elaeniine assemblage. Molecular Phylogenetics and Evolution 46, 88–101. doi:10.1016/j.ympev.2007.09.011
Ridgely, R. S., and Greenfield, P. J. (2001). ‘The Birds of Ecuador. Vols 1 and 2.’ (Comstock Publishing Associates: Ithaca, NY.)
Ridgely, R. S., and Tudor, G. (1994). ‘The Birds of South America. Vol. II. The Suboscine Passerines.’ (University of Texas Press: Austin, TX.)
Swofford, D. L. (2002). ‘PAUP*: Phylogenetic Analysis Using Parsimony (*and other methods). Version 4.0.’ (Sinauer Associates: Sunderland, MA.)
F. E. Rheindt et al.
Tamura, K., and Nei, M. (1993). Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzee. Molecular Biology and Evolution 10, 512–526.
Tello, J. G., and Bates, J. M. (2007). Molecular phylogenetics of the tody-tyrant and flatbill assemblage of tyrant-flycatchers (Tyrannidae). Auk 124, 134–154. doi:10.1642/0004-8038(2007)124[134:MPOTTA]2.0. CO;2
Zimmer, J. T. (1941). Studies of Peruvian birds. No. XXXVII. The genera Sublegatus, Phaeomyias, Camptostoma, Xanthomyias, Phyllomias and Tyrannus. American Museum Novitates 1109, 1–25.
Zimmer, K. J., and Whittaker, A. (2000). Species limits in Pale-tipped Tyrannulets (Inezia: Tyrannidae). Wilson Bulletin 112, 51–66. doi:10.1676/0043-5643(2000)112[0051:SLIPTT]2.0.CO;2
Zimmer, K. J., Whittaker, A., and Oren, D. C. (2001). A cryptic new species of flycatcher (Tyrannidae:Suiriri) from the cerrado region of central South America. Auk 118, 56–78. doi:10.1642/0004-8038(2001)118 [0056:ACNSOF]2.0.CO;2
Manuscript received 30 April 2008, accepted 04 July 2008