J. Mol. Biol. (1971) 56, 491-506 Genetic Information in Oocyte RNA ERIC H. DAVIDSON AND BARBARA R. HOIJGH The Rockefeller University, New York, N.Y. 10021, U.S.A. (Received4 September 1970, and in revised form 20 November 1970) RNA of the mature Xenopus oocyte was hybridized with the isolated non- repetitive DNA fraction of the Xenopm genome. The hybrids had the thermal stability and other characteristics expected of non-repetitive DNA-RNA hybrids, and DNA recovered from them reassociated at the rate of non-repetitive DNA. By saturating the non-repetitive DNA with oocyte RNA a direct minimum estimate was obtained of the amount of non-repetitive sequence in the oocyte RNA, i.e. its genomic information content. The oocyte RNA contains about 20 x lo6 nucleotidepairs of diverse sequence, which is 4.5 times the total genomic information present in the Eecherichia coli chromosome. 1. Introduction Transcription of the genetic program for development begins in oogenesis. Stored messenger RNA inherited from oogenesis has been demonstrated in a variety of early embryos (Gross, 1967; Kedes & Gross, 1969), though the diversity of the proteins coded for by maternal messenger RNA can not yet be appreciated. In Xenopus protein synthesis proceedsactively during the first few cleavagesbefore any new high- molecular weight RNA synthesis can be detected. RNA-DNA hybridization studies indicate that heterogeneous RNA’s synthesized in oogenesis remain present in the Xenopus embryo after fertilization (Davidson, Crippa, Kramer & Mirsky, 1966). The population of these RNA’s transcribed from repetitive DNA sequences during oogenesis appears to persist until well into blastulation (Crippa, Davidson & Mirsky, 1967 ; Crippa & Gross, 1969), and homology cannot be detected between the RNA’s synthesized by the embryo at this stage and those inherited from oogenesis (Davidson, Crippa & Mirsky, 1968). No previous studies, however, can provide an estimate of the informational complexity of whatever developmental program is carried in the oocyte RNA. It is the purpose of the studiesreported here to estimate the complexity of the RNA stored in the unfertilized Xenopus egg. The term complexity, as used in this paper, denotes specifically the amount of diverse nucleotide sequence, i.e. sequences lacking homology to one another at a given experimental criterion. The quantitative estima- tion of complexity is of course dependent on the criterion used to establish sequence homology (the criterion is set by the cation concentration and the incubation tempera- ture). With our criterion, about 45% of the Xenopus genome displays internal sequence homology, the remainder behaving asa non-repetitive sequence eomponent (Davidson & Hough, 1969). To measure the complexity of the oocyte RNA we hybridized it with the isolated non-repetitive DNA fraction. In this way the predominant forma- tion of imperfect hybrids with repetitive sequence transcripts is avoided. The mass of RNA hybridized at saturation of the non-repetitive DNA provides a measureof the 491 22
Transcript
J. Mol. Biol. (1971) 56, 491-506
Genetic Information in Oocyte RNA
ERIC H. DAVIDSON AND BARBARA R. HOIJGH
The Rockefeller University, New York, N.Y. 10021, U.S.A.
(Received 4 September 1970, and in revised form 20 November 1970)
RNA of the mature Xenopus oocyte was hybridized with the isolated non- repetitive DNA fraction of the Xenopm genome. The hybrids had the thermal stability and other characteristics expected of non-repetitive DNA-RNA hybrids, and DNA recovered from them reassociated at the rate of non-repetitive DNA. By saturating the non-repetitive DNA with oocyte RNA a direct minimum estimate was obtained of the amount of non-repetitive sequence in the oocyte RNA, i.e. its genomic information content. The oocyte RNA contains about 20 x lo6 nucleotide pairs of diverse sequence, which is 4.5 times the total genomic information present in the Eecherichia coli chromosome.
1. Introduction Transcription of the genetic program for development begins in oogenesis. Stored messenger RNA inherited from oogenesis has been demonstrated in a variety of early embryos (Gross, 1967; Kedes & Gross, 1969), though the diversity of the proteins coded for by maternal messenger RNA can not yet be appreciated. In Xenopus protein synthesis proceeds actively during the first few cleavages before any new high- molecular weight RNA synthesis can be detected. RNA-DNA hybridization studies indicate that heterogeneous RNA’s synthesized in oogenesis remain present in the Xenopus embryo after fertilization (Davidson, Crippa, Kramer & Mirsky, 1966). The population of these RNA’s transcribed from repetitive DNA sequences during oogenesis appears to persist until well into blastulation (Crippa, Davidson & Mirsky, 1967 ; Crippa & Gross, 1969), and homology cannot be detected between the RNA’s synthesized by the embryo at this stage and those inherited from oogenesis (Davidson, Crippa & Mirsky, 1968). No previous studies, however, can provide an estimate of the informational complexity of whatever developmental program is carried in the oocyte RNA.
It is the purpose of the studies reported here to estimate the complexity of the RNA stored in the unfertilized Xenopus egg. The term complexity, as used in this paper, denotes specifically the amount of diverse nucleotide sequence, i.e. sequences lacking homology to one another at a given experimental criterion. The quantitative estima- tion of complexity is of course dependent on the criterion used to establish sequence homology (the criterion is set by the cation concentration and the incubation tempera- ture). With our criterion, about 45% of the Xenopus genome displays internal sequence homology, the remainder behaving as a non-repetitive sequence eomponent (Davidson & Hough, 1969). To measure the complexity of the oocyte RNA we hybridized it with the isolated non-repetitive DNA fraction. In this way the predominant forma- tion of imperfect hybrids with repetitive sequence transcripts is avoided. The mass of RNA hybridized at saturation of the non-repetitive DNA provides a measure of the
491 22
492 E. H. DAVIDSON AND B. R. HOUGH
frtmtion of the total non-repetitive sequence represented in the oocyte RNA, and hence of the complexity of this RNA. In an initial series of experiments reported previously (Davidson & Hough, 1969), we found that the newly synthesized RNA of the lampbrush stage Xenopus oocyte hybridizes with non-repetitive DNA. Our present study deals with the RNA of mature oocytes, rather than that of lampbrush stage oocytes, and thus with the RNA stockpile destined to be inherited directly by the embryo. Some characteristics of the non-repetitive DNA-RNA hybrids are presented, as well as the saturation experiments, which indicate that at least 1.2% of the non-repetitive sequence in the genome is represented in the oocyte RNA.
2. Materials and Methods (a) Reassociation of DNA
DNA was prepared from Xenopua red blood cells by standard methods, as previously described (Davidson & Hough, 1969). All DNA used in this study wae sheared to about 450 nucleotides by forcing it through a needle valve under 50,000 lb. ins pressure in the apparatus built for that purpose by Dr Roy Britten, and kindly made available to us. To obtain the date on which the Cot curve is based, the various sheared DNA solutions were allowed to reassociate for different periods of time at 60°C and the fractions re- associated were assayed by hydroxyapatite chromatography, as described by Britten & Kohne (1966, 1968). Cot is expressed in terms of moles nucleotide per 1. x time of anneal- ing, in seconds, for solutions annealed at 60°C in 0.12 M-phosphate buffer (0.18 M-N&+), composed of equimolar quantities of Na2HP04 and NaH2P0,. Various molarities of this buffer (POc buffer) are used in all the experiments reported in this paper. Reassocietion rate is accelerated at higher salt concentrations, and the parameter used in this paper is the “equivalent Cot,” the Cot multiplied by the appropriate factor for rate increase due to salt concentrations above 0.18 M-N& + . The salt correction factors for rate relative to rate in 0.12 M-PO~ buffer are calculated by Britten & Smith (1969) from data of Wetmur & Davidson (1968).
(b) Isolation of non-repetitive DNA
To isolate the non-repetitive sequence fraction the sheared DNA was incubated to a Cot of 2600, which is not significantly different from the half-reassociation point for the non-repetitive fraction, and the non-reassociated portion of the annealing mixture was harvested from an hydroxyapetite column. The DNA was then dialyzed free of salt and concentrated by lyophilization. Samples of the DNA fraction thus isolated were allowed to reanneal and the duplexes formed were melted. The thermal stability of these duplexes, relative to that of unsheared DNA, is the same as that of duplexes formed from equally sheared &cherichia co.% DNA (Davidson & Hough, 1969). Thus the non-repetitive DNA duplexes melt with the same hyperchromicity as native DNA (39%), in contrast to the less stable duplexes formed from a reassociated repetitive DNA fraction isolated at a Cot of 60, the hyperchromicity of which is only 27%. The melting behavior of the DNA fraction isolated from the column at a C,t of 2500 is consistent with its definition as a non-repetitive sequence fraction.
(c) Isolation of 14C-labeEed non-repetitive DNA
14C-labeled DNA was prepared from cultured Xenopwr kidney cells kindly provided by Dr Joseph Gall. The cells were exposed to [21-14C]thymidine (25 ,uc/ml. medium; 52.8 me/m-mole), and after 1 week, during which the cells grew to confluence, they were harvested and the 14C-labeled DNA extracted. This DNA was fraction&ed as above at C,t = 2500 and the non-repetitive fraction obtained.
(d) Preparation of oocyte RNA
Mature oocyte RNA was extracted from ripe ovarian oocytes. The oocytes were manuslly removed from their follicles with watchmakers’ forceps, after immersion in Barth’s solution X (Barth t Barth, 1959) supplemented with an additional IO.3 g N&l/l. 3000 to 6000
GENETIC INFORMATION IN OOCYTE RNA 493
oocytes were used for each RNA preparation. In some experiments follicular membrane
fragments adhering to the oocytes after removal of the oocyte from the follicle were washed off in Ca- and Mg-free Steinberg’s medium containing 0.001 M-EDTA, as suggested
by Masui (1967). RNA extracted from such EDTA-cleaned oocytes behaved in hybridiza- tion studies in exactly the same way as RNA extracted from oocytes not washed with
EDTA; thus, surviving follicular membrane fragments contribute, if any, a negligible amount of DNA or RNA complementary to non-repetitive Xenqw DNA sequences.
The RNA was extensively purified as described previously (Crippa et al., 1967), except that the DNase treatment was omitted and a third phenol extraction was sometimes performed after pronase and prior to methylation. The RNA was twice precipitated in
ethanol from 0.3 M-potassium acetate. The RNA was methylated in w&o with dimethyl sulfate (110 me/m-mole), according to Smith, Armstrong & McCarthy (1967). Final
specific activities of 8000 to 19,000 cts/min/pg were obtained. Before use the RNA was purified of adherent [3H]dimethylsulfate by passage through Sephadex G50 and two
further ethanol precipitations.
(e) RNA-DNA hybridization
To prepare the annealing mixtures for hybridization, the RNA was precipitated from ethanol and dissolved in 100 to 200 ~1. of denatured non-repetitive DNA made up in 0.41 M-
PO, buffer. To remove heavy metal cations we passed all solutions used over a Chelex (Bio-Red) column prior to annealing.
In order to prevent evaporation during the extended annealing periods, we utilized specially constructed Teflon vials containing conical wells. The wells could be hermetically sealed by the insertion of an overlying threaded plug. To ensure tight sealing during the
incubation the plug is made of Kel-F, which is slightly harder than Teflon, and lateral expansion of the threaded female cylinder is prevented by an external stainless steel band.
The vials were immersed in a 60°C water bath for the periods of time necessary to attain the desired C’,t.
Following the annealing period the samples were diluted to 0.24 M-PO, buffer and treated with 20 fig RNase A/ml. for 2 hr at room temperature. They were then passed
over a Sephadex G200 column, which removes the RNase and digested RNA fragments, while excluding the DNA and any hybridized RNA. The exclusion peak was separated
into single-stranded and duplex fractions by hydroxyapatite and the thermal stability of the RNA-DNA hybrids was routinely assayed by melting them from the hydroxyapatite
column. The samples were introduced to a 1.3-ml. hydroxyapatite column operating at 60°C and washed with 36 ml. of 0.12 M-PO4 buffer. After ribonuclease treatment and Sephadex, all non-hybridized RNA is usually washed out of the column in this volume of buffer at
60°C. Any remaining fragments of RNA are eluted in a 6-ml. wash at 65”C, as shown by experiments in which no DNA was present. To melt RNA-DNA hybrids the temperature
was raised to 65”C, then to 7O”C, and in 4 deg.C steps to 98°C. At each temperature 6 ml. of effluent were collected. The effluents were assayed for 0.D.260 in 4-cm cuvettes and
then individually lyophilized and counted in a scintillation spectrometer.
3. Results
(a) The non-repetitive sequence fraction of Xenopus DNA
The kinetics of reassociation of sheared Xenopus DNA are portrayed in Figure 1
(solid line) as a hydroxyapatite Cot curve. The ordinate of Figure 1 represents the fraction of the DNA remaining bound to the hydroxyapatite column, i.e. behaving as a structure containing double-stranded regions. The controlling parameter of the reassociation reaction is the concentration of the annealing solution times the time of incubation, termed Cot by Britten & Kohne (1966,1968), who originated this method of studying DNA reassociation kinetics. The most slowly reassociating fraction, representing about 56% of the genome at the 60°C criterion employed, is the non- repetitive sequence fraction. The sequences reassociating more rapidly than the
. .
. . e~
-e-a
+ .
20-
‘s
\ E 1
- 8 f K 0
40-
.Lf
: __
___ -
-- -
----
--
----
----
----
----
-_
_-
-z
2 a al
- E i?
-
CT 80
-
yo-3
I
I I
10-Z
10
-l 10
0 IO
’ IO
" 10
4 10
5
Eouw
olen
t co
t
0
FIG
. 1.
Hy
drox
yapa
tite
C,t
curv
e of
sh
eare
d Xe
nopu
s DN
A,
base
d on
da
ta
obta
ined
fro
m
the
reas
socia
tion
of
13 D
NA
conc
entra
tions
, ea
ch
at
one
of
6 sa
lt co
ncen
tratio
ns
rang
ing
from
0.
12
to
0.32
M
-PO,
bu
ffer.
For
deta
ils
of
thes
e so
lutio
ns
see
our
prev
ious
repo
rt (D
avids
on
& Ho
ugh,
19
69).
Ord
inate
an
d ab
scis
sa
are
expl
aine
d in
th
e te
xt.
The
curv
e is
fit
ted
to
the
poin
ts
by
a co
mpu
ter,
oper
atin
g ac
cord
ing
to
a le
ast-s
quar
es
met
hod
(Brit
ten
& Ko
hne,
19
68).
The
dotte
d lin
e re
pres
ents
th
e co
mpu
ter’s
pr
edict
ion
of
the
reas
socia
tion
reac
tion
of
the
non-
repe
titive
co
mpo
nent
al
one.
Th
e no
n-re
petiti
ve
fract
ion
acco
unts
fo
r 55
’$!o
of
the
tota
l DN
A,
at
the
60°C
cr
iterio
n ap
plie
d in
th
ese
expe
rimen
ts.
GENETIC INFORMATION IN OOCYTE RNA 495
non-repetitive fractions (i.e. at C,t values lower than 300 or so) are repetitive, since their higher reassociation rate depends on their presence in higher concentration in the annealing mixtures. Identification of the most slowly reassociating fraction as the non-repetitive fraction is based on its reassociation rate, and on its rate of reassociation relative to that of a known non-repetitive DNA (E. COG) ; details of this and other experimental criteria for the Xenopus non-repetitive DNA fraction are given in our earlier publication (Davidson t Hough, 1969).
Figure 1 also displays an ideal second order reaction component (dotted line) which predicts the contribution of the non-repetitive sequence fraction alone. This theoretical curve was derived by a computer directed to decompose the over-all Cot curve into second-order reaction components, according to a program designed by Roy Britten.
(b) Reassociation of the isolated non-repetitive DNA fraction
The reassociation rate of the isolated non-repetitive DNA fraction is shown in Figure 2. Only 1+3% of the DNA binds to the column at low Cot’s, while the bulk of the DNA reassociates at C,t’s greater than 100. The range of C,t’s over which re- association takes place is slightly greater than that expected of a single kinetic
IO- a
20.
60-
.
70- 60- \
t
90 ~-
100’ 10-z
I 0’
I 10-O
I I I I IO’ 102 103 104
Equivalent Cot
FIQ. 2. Reassooiation of the isolated non-repetitive DNA oomponent expressed es an equivalent C,t curve with the same axes as that in Fig. 1.
Hydroxyapatite determinations were made on non-repetitive DNA solutions of 11 &ml. in 0.12 aa-PO1 buffer for equivalent C&s from 0.04 to 2.2; l-1 mg/ml. in 0.40 w-PO4 buffer for equivalent 0,$‘s from 100 to 11,000 and 3.3 mg/ml. in 0.12 aa-PO1 buffer for the equivalent C,t of 37,600.
496 E. H. DAVIDSON AND B. R. HOUGH
component. That is, more than a single rate of reassociation is included in the data plotted. For Xenopus, with about 55% non-repetitive DNA in a 3 pg genome, the ideal half-C,t for the isolated non-repetitive fraction would be about 1300. The early portion of the reassociation reaction seems to occur in accordance with this prediction, while the later points appear to be retarded. Thus, DNA reassociation should be underway by a Cot of 100 to 200, as is the case in Figure 2, but ideally the reaction should have been essentially complete by C,t = 1300. The relatively high viscosity of the concentrated solutions used to obtain the curve in Figure 2 is certainly involved in this retardation (Wetmur & Davidson, 1968). Any DNA degradation might also have affected the over-all kinetics, and in addition the rate could have been affected by complex events particular to long-term incubations of such isolated non-repetitive sequence fractions. Figure 2 indicates that other than the I+G”~ of the DNA reassociating early, no significant sequence repetition could be present in the non-repetitive sequence fraction.
(c) Reassociation of 14C-lubeled non-repetitive DNA
The non-repetitive DNA fraction isolated from 14C-labeled DNA as described in Materials and Methods, when mixed with excess unlabeled non-repetitive DNA, reassociated at an identical rate with the unlabeled DNA. Though extracted from
TABLE 1
Reassociation of “C-ilabeled non-repetitive DNA incubated with whole DNA carrier.
Reassociation Reassociation of whole DNA of labeled
(by O.D.) DNA (by cts/min)
(%) (%)
Reassociation of non-repetitive
DNA alone (from Fig. 2)
(%I
1.26 21.1 1.6 1.4 12.6 33.3 2.7 1.7
125 34.9 8.2 9.5 1000 60.2 32.0 33.5
Whole sheared DNA was present at 0.211 mg/ml. in 0.12 M-PO* buffer in the Cot 1.26 sample and at 2.11 mg/ml. for the other samples. Each sample contained about 4300 cts/min of ‘*C-labeled non-repetitive DNA, at a specific aotivity of 6700 cts/min/rg in the counting mode used. Values in the last ~cohnnn are derived from the experiment of Fig. 2 which, as described in the text, was carried out with unlabeled non-repetitive DNA.
kidney tissue culture cells rather than red blood cells, this labeled DNA fraction could thus be regarded as indistinguishable in composition from the unlabeled non-repetitive DNA fraction. Its rate of reassociation in the presence of excess whole DNA is shown in Table 1. Within the precision of the determinations, no difference between the rate of reassociation of the labeled non-repetitive DNA in the presence of whole DNA, and the rate of reassociation of the non-repetitive DNA alone can be observed. Ideally, reassociation of the labeled DNA in the presence of whole DNA should occur at a slower rate than does reassociation of the isolated non-repetitive fraction, since the non-repetitive sequences are diluted with other sequences in whole DNA. How- ever, as we have already noted, the rate of reassociation of the isolated non-repetitive
GENETIC INFORMATION IN OOCYTE RNA 497
fraction deviates from expectation in being too slow, thus obscuring this effect. Table 1 demonstrates in any case that no appreciable fraction of the non-repetitive DNA preparation represents sequences which are present repetitively in the genome. At C,t values of 125 and 125, this experiment provides a particularly sensitive test for the presence of low frequency repetitive sequences. Were such sequences present in detectable quantity in the labeled preparation, they would have reassociated with the homologous repetitious families in the whole DNA, and more rapid reassociation of the labeled segments would have been observed.
(d) Precocious binding of a small fraction of non-repetitive DNA
As does the unlabeled non-repetitive DNA preparation, when incubated alone 1.8% of the labeled non-repetitive DNA binds to the column at low C,t’s. A sample of this preparation was incubated to a Cot of 10 and the 1.8o/o bound was harvested. Any repetitive sequences contaminating the non-repetitive fraction should be represented in the DNA binding to the hydroxyapatite column after incubation to Cot 10, since portions of all known repetitive sequence families will be present in the duplex population at C,t 10. The 14C-labeled, bound fraction was then mixed with whole unlabeled DNA and incubated to Cot 1OOO.46o/o of the counts reassociated. Had the labeled fraction consisted of a repetitive sequence contaminant in the non-repetitive fraction, lOOo/o of the counts would have reassociated in the presence of whole DNA. About 150/O more label reassociated by C,t 1000 than expected for pure non-repetitive DNA, on the other hand, suggesting that 15% of 1*8o/o of the original non-repetitive fraction, or O+27o/o, might belong to repetitive sequence families. The remainder of the l*8o/o binding to the column after incubation to Cot 10 thus appears to bind as an artifact. This interpretation was checked by reannealing to C,t 10 a fraction from which the 1.8% of the DNA binding early had already been stripped: 1.4% of the DNA again bound. This behavior also suggested that the hydroxyapatite we used binds a small amount of single-stranded DNA. When the column was washed before use with a few micrograms of denatured DNA fragments early binding was
reduced by 50%, thus demonstrating that a small amount of artifactual binding of single-stranded DNA occurs. These data, together with those of Table 1, suggest that these non-repetitive DNA fractions are as free from repetitive sequence contamination as kinetic methods permit. In the limit, a non-repetitive sequence fraction will contain a single copy of each repetitive sequence, and in Xenopus we calculate that at our criterion this could contribute about 0*30/O of the t,otal non-repetitive sequence length.
(e) Characteristics of the RNA-non-repetitive DNA hybrids
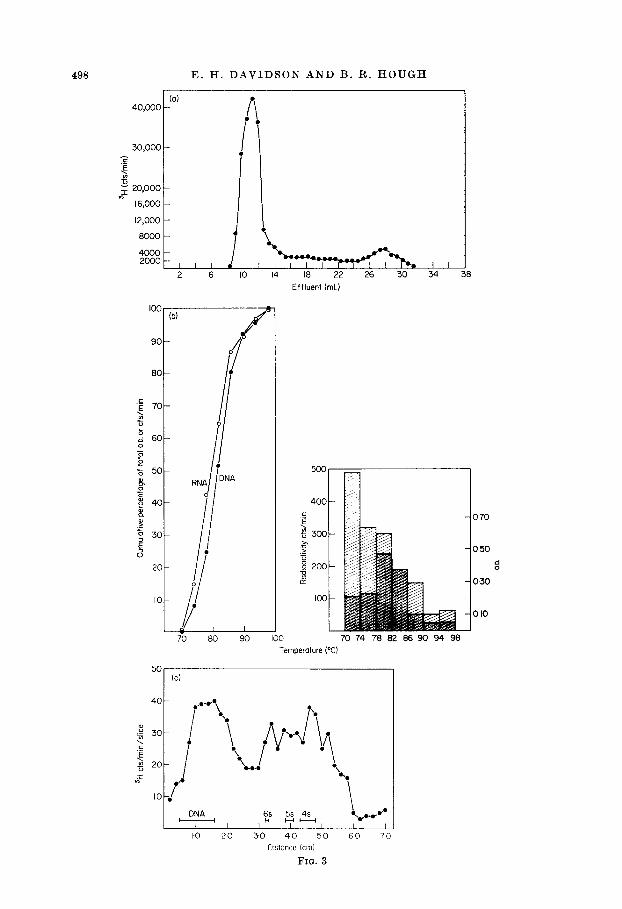
In order to attain the C&s at which non-repetitive DNA sequences will hybridize with complementary RNA it is necessary to use extended annealing times which involve the risk of RNA degradation. Therefore, the integrity of the RNA was routinely checked after annealing by comparing its pattern of elution from Sephadex G200 with that of the starting material. Figure 3(a) shows such an elution pattern, obtained in this case after an incubation of five days at 40 mg RNA/ml. This pattern is similar to those we reported earlier (Davidson & Hough, 1969), for RNA extracted from a different source, lampbrush-stage oocytes, and incubated at concentrations about one-tenth as great. Little degradation of the RNA can be observed, a result which in our experience depends on the absence of both nuclease activity and heavy metal cat,ions.
498 E. H. DAVIDSON AND B. R. HOUGH
30,oco -
16,000 -
12,000 -
I 2 6 IO 14 18 22 26 30 34
Effluent (ml)
go-
80-
20-
IO-
~ I/ 70 80 90 100 70 74 78 82 86 90 94 98
050
8 0 2 200 8
0-x)
100
010
100 1 (b)
8
Temperature (“C)
, DNA , 6,”
5s 4s ‘.p.v u- HH
/ I I I-O 20 30 40 50 GO 7 0
GENETIC INFORMATION IN OOCYTE RNA 499
The melting behavior of RNA-DNA hybrids was examined as a measure of the excellence of base pairing. Figure 3(b) portrays an hydroxyapatite melt of an annealing sample after ribonuclease treatment and exclusion from Sephadex. It is evident that the mean thermal stability of the RNA-non-repetitive DNA hybrids is only 1 to 2 deg. C below that of the non-repetitive DNA-DNA duplexes. A similar result was obtained in our previous study of lampbrush stage oocyte RNA-non-repetitive DNA hybrids (Davidson t Hough, 1969). It is worth noting that the present experi- mental series includes RNA:DNA ratios over an order of magnitude higher than in these earlier experiments, without resulting in any decrease in hybrid thermal stability.
Hybrid formation with the related DNA sequences of repetitive sequence families results in significant nucleotide mispairing, and hence a lowered thermal stability (see for example Shearer & McCarthy, 1970). On the other hand, the high thermal stability of the hybrids portrayed in Figure 3(b) indicates that these hybrids contain little base pair mismatch. The oocyte RNA-DNA hybrids thus display the thermal stability characteristics expected of RNA-non-repetitive DNA hybrids. Hybrid duplexes of comparable thermal stability, relative to DNA-DNA duplexes, have previously been reported only for bacterial systems, which also lack repetitive DNA sequences. For example Kohne (1968) reported an 8 deg. C difference in T, between E. coli ribosomal RNA-ribosomal DNA hybrids and ribosomal DNA-ribosomal DNA duplexes. Similar T, values were reported by Moore & McCarthy (1967), and Bolton & McCarthy (1964) in studies of E. coli messenger RNA-DNA hybrids.
A further criterion of the excellence of base pairing in these hybrids, and thus of their interpretability as bona jide non-repetitive DNA-RNA hybrids, is the size of the hybridized RNA molecules. In Figure 3(c) is displayed the polyacrylamide gel electrophoresis profile of RNA’s recovered from the hybrid duplexes by melting.
Fra. 3. Criteria for hybridization of oocyte RNA with non-repetitive DNA. (a) Sephadex G200 profile of [3H]methyl-labeled mature oocyte RNA after 6 days incubation
at 60°C. 6.0 mg of RNA (40 mg/ml. in 0.40 M-PO~ buffer) were sealed in a Teflon vial together with 100 pg of non-repetitive DNA. Almost all of the RNA elutes at the exclusion peak, showing that it survives the annealing periods.
(b) Hydroxyapatite melt of hybrids between [3H]methyl-labeled oocyte RNA and the isolated non-repetitive DNA fraction. The input was 2.1 mg RNA, at 16 mg/ml., plus 140 ti DNA, and incubation was for 10 days, to an equivalent C,t of 13,600. After annealing, the hybrids were treated with ribonuclease, excluded from a Sephadex G200 column and melted, as described in Materials and Methods. The data are plotted both integrally (left) and incrementally (right) as a bar graph.
(c) Polyacrylamide gel electrophoresis of [sHImethyl-labeled RNA melted from a hybrid with non-repetitive DNA. 166 mg of RNA were annealed at a concentration of 63.8 mg/ml. with 400 H non-repetitive DNA in 0.46 M-PO* buffer, to an equivalent C,t of 8600. The sample was treated with ribonuclease, and the oligoribonucleotides (and ribonuclease) removed in a Sephadex G200 column as usual. The sample was then placed on an hydroxyapatite column, washed with 48 ml. of 0.12 M-PO, buffer at 60°C, 12 ml. at 66”C, and 12 ml. at 70°C. The column temperature was then abruptly raised to 98°C. The 98°C effluent was dialysed overnight against running distilled water, to remove salts and dissociate any weak interactions among the nucleic acids which might have occurred after the sample left the column, and lyophilized. 68 ~1. of a 100.4. water solution of the lyophilizate, containing 3400 counts, were placed in an 8% gel, according to Goldstein & Harewood (1969), and run for 80 min at 6 milltube. Marker gels were stained with acridine orange and experimental gels were sliced and hydrolyzed according to Weinberg, Loening, Willems & Penman (1967) and oounted. The 4 s, 6 s and 6 s RNA’s used as markers had been purified from E. coli and were generously provided by Dr Goldstein.
500 E. H. DAVIDSON AND B. R. HOUGH
Present in the gel are some low molecular weight marker RNA’s and the 400 to 500 nucleotide non-repetitive DNA fragments. Since hybrids formed with repetitive DNA usually contain significant nucleotide mismatch, ribonuclease action at the mispaired sites makes it possible to recover only small RNA fragments in such an experiment. In contrast, the more perfectly matched hybrids expected with non-repetitive DNA should yield some RNA’s of molecular size equivalent to that of the DNA to which they were hybridized. A relevant example is at hand in the work of Jeanteur $ Attardi (1969), who have recovered high molecular weight RNA molecules from well- matched hybrids formed with ribosomal RNA’s, in contrast to low molecular weight RNA recovered from imperfectly base-paired hybrids. Figure 3(c) shows that a significant fraction of the oocybe RNA molecules melted from the hybrids are of about t,he same size as the DNA fragments. The ribonuclease has apparently clipped off the non-hybridized ends of RNA molecules, since the gel contains little or no RNA migrating more slowly than the DNA fragments. Much of the hybridized RNA is shorter than the DNA fragments, as is expected, since the shearing t’o which the DNA has been subjected almost certainly breaks the DNA molecules at random with respect to the units of transcription in vivo. The shortest RNA’s present are about half the length of the 4 s RNA marker.
(f) Recovery of non-repetitive DNA from RNA-DNA hybrids
As a demonstration that the oocyte RNA is actually hybridized with non-repetitive DNA sequences, we annealed a small amount of non-repetitive 14C!-labeled DNA with an excess of oocyte RNA, and isolated the duplex fraction on an hydroxyapatite column. Half of this fraction was used for an estimate of RNA-DNA hybrid content. From the other half we recovered the 14C-labeled DNA from the hybrids by heat denaturation. The recovered DNA was then mixed with an excess of unlabeled whole DNA, and its reassociation rate measured as an index of its non-repetitiveness. The experiment (Table 2, first annealing) wa,s carried out as follows: 20,000 pg of oocyte RNA and 8 pg of non-repetitive 14C-labeled DNA were dissolved in 0.81 ml. of 045 M-
PO, buffer and annealed to an equivalent Cot of 80, with respect to the DNA. The RNA concentration in this experiment, 2500 times the DNA concentration, was high enough to ensure reaction of RNA with non-repetitive DNA (this follows from the data obtained in the saturation experiments presented below). Thus the rate of the hybridization reaction of those DNA sequences represented in the RNA population will be controlled by the RNA Cot value rather than the DNA Cot. After ribonuclease and Sephadex treatment as usual, the duplexes (about 8.7% of the DNA) were separated on the column. The duplex fraction now contained both RNA-DNA hybrids and DNA-DNA duplexes, those expected to form by DNA Cot 80 (see Table 1). As shown below, approximately one-sixth of the total 14C-labeled DNA in duplex form was bound to RNA. To a portion of the duplex fraction containing 3700 cts/min of 14C-labeled DNA (0.5 pg) we added 3 mg of whole unlabeled DNA (Table 2, second annealing). This mixture was denatured and allowed to reassociate to Cot 50. Had the 14C-labeled DNA engaged in RNA-DNA hybridization consisted of repetitive sequences these sequences would have completely or almost completely annealed with the homologous sequences in the unlabeled DNA. Consequently, by Cot 50, the reassociation of labeled with unlabeled DNA would have been increased by 17% above that predicted if all the 14C-labeled DNA recovered were non-repetitive. If on the other hand the DNA which had been hybridized with RNA were non-repetitive,
TABL
E 2
Res
ults
of
ex
perim
ents
w
ith
“C-li
zbel
ed
DN
A re
cove
red
from
hy
brid
s
Non-
repe
titive
[‘W
]DNA
An
neal
ed
with
DN
A %
of
[‘V
]DNA
eq
uiva
lent
in
du
plex
Cd
fract
ion
Frac
tion
of
tota
l [‘*
C]DN
A hy
bridi
zed
with
RNA
DNA
(rele
ased
by
RN
ase)
Firs
t an
neal
ing
53,0
00
cts/
min
(8
CL&
20
mg
oocy
te
RNA
80
8.7
74%
0.
9%
Seco
nd
anne
alin
g 37
00
cts/
min
of
3
mg
whol
e 50
8
[‘W]D
NA
(O+g
) DN
A m
elte
d fro
m
abov
e du
plex
fract
ion
2 0.5
0 -
-2
B B : 0.4
0 -
P f 0 0.3
0 -
0”
” 2 2 2 02
0 -
g
040
t
v 13
,600
lncub
otion
sa
mple
s
/ A
Input
Equiv
. RN
A:DN
A co
t RN
A DN
A hg
/ml.)
(p
g )
\ Ti
me
M%
)
I 13
,600
268
I34
6
2.5
13,60
0 6.7
13
4 6
5 13
,600
I34
134
6
15
6800
15
0 M
O
I5
13,60
0 15
0 I40
15
6800
15
.0 14
0
30
6800
30
-O
I40
30
13,60
0 30
0 I40
60
5000
25
.5 70
40
8600
53
-8 loo
40
8600
53
-8 30
0
50
50
Il.400
5700
400
40.0
100
100
70
5700
56
.2 10
0
60
2500
52
6 96
60
5600
52
-6 96
5 0 5 5 0 0 4 4 0 5 5 2 4-5
Input
RNA:
DNA
FIG.
4.
Sa
turati
on
of
non-
repe
titive
DNA
with
m
atur
e oo
cyte
RNA.
Ch
arac
terist
ics
of
each
inc
ubati
on
sam
ple
arc
given
in
the
Table
to
the
rig
ht
of
the
satur
ation
cu
rve;
meth
ods
are
as
desc
ribed
in
text.
Th
e nu
mbe
rs ad
jacen
t to
ea
ch
point
re
pres
ent
the
DNA
C&s
to
which
the
sa
mple
s we
re
incub
ated.
At
RNA:
DNA
inputs
15
an
d 50
an
alt
erati
on
in an
neali
ng
proce
dure
was
emplo
yed,
as
an
addit
ional
contr
ol,
dupli
cate
sam
ples
were
inc
ubate
d fo
r 5
days
, the
n ex
pose
d to
IO
O’C
and
reinc
ubate
d fo
r a
seco
nd
5-day
pe
riod.
This
contr
ol sh
ows
that
the
se
cond
5
days
of
the
lo-
day
incub
ation
pe
riods
us
ed
in oth
er
sam
ples
at
these
inp
uts
are
as
effec
tive
as
the
first
5 da
ys,
since
no
ch
ange
in
the
amou
nt of
hy
bridiz
ation
wa
s ob
serve
d. Du
ring
the
anne
aling
pe
riod
the
RNA
neith
er
hydro
lyzes
(F
ig.
3(a)
)
nor
suffe
rs
any
other
m
ore
subtl
e fo
rms
of
degr
adati
on
detec
table
by
the
criter
ion
of
hybr
idiza
tion.
GENETIC INFORMATION IN OOCYTE RNA 603
only about 8% should have reassociated by Cot 60. As Table 2 shows, the experiment yielded exactly 8.0% reassociation of the labeled DNA.
The content of DNA-RNA hybrids in the annealing mixture was estimated in an experiment in which half of the original duplex fraction was digested exhaustively with ribonuclease (40 pg/ml. for 60 hours at 37°C). Similar ribonuclease conditions were used by Gelderman, Rake & Britten (1968,197O) to digest mouse RNA-non- repetitive DNA hybrids. The 14C-labeled DNA which had been engaged in hybrid formation with RNA was rendered single stranded by this treatment, so that on passage through a second hydroxyapatite column it no longer bound to the column (incubation with ribonuclease under these conditions had almost no effect on DNA- DNA duplexes in a control experiment). The fraction of the total non-repetitive 14C-labeled DNA entering the experiment which was released from the duplex form by ribonuclease was about O-9%, and this value provides an independent estimate of the fraction of non-repetitive DNA sequences represented in the oocyte RNA.
(g) Saturation of the non-repetitive DNA with oocyte RNA
By increasing the ratio of RNA to DNA in the annealing mixtures the concentration of rarer RNA species is increased so that they react significantly with the non- repetitive DNA. The results of a series of experiments with increasing RNA:DNA ratio is presented in Figure 4. At saturation no further sequences of the non-repetitive DNA react with increasing RNA concentration, indicating that all DNA sequences represented in the RNA population are hybridized. The percentage of the total mass of DNA present in duplexes which is RNA-DNA hybrid is plotted on the ordinate of Figure 4, that is, the fraction of the non-repetitive DNA reassociated at a given Cot which is hybridized with RNA. Each point in Figure 4 represents a sample which was treated with ribonuclease, excluded from Sephadex and bound to an hydroxyapatite column, as described above. DNA-DNA duplexes and RNA-DNA hybrids were then melted together from the column, as in Figure 3(b). All the hybrid preparations represented in Figure 4 displayed the appropriate thermal stability. Unacceptable results in these experiments occurred only on those unusual occasions when, according to Sephadex G200 assay, extensive RNA breakdown had taken place during the annealing period.
In the samples at the lower RNA:DNA ratios in Figure 4 the represented DNA sequences are probably in excess and the reaction rate is controlled by the DNA Cot value. As saturation is approached the concentration of hybridizable RNA mole- cules will begin to affect the rate of the reaction while beyond saturation hybridizable RNA will be present in excess and the reaction rate should depend primarily on RNA concentration. As the equivalent C&s adjacent to each point in Figure 4 demonstrate, the percentage of DNA hybridized with RNA in these experiments appears to be a function of the RNA: DNA ratios in the annealing mixtures rather than of the DNA Cot. While the reaction is controlled by the DNA concentration this is expected to be the case. RNA: DNA ratios in Figure 4 increase beyond the saturation point by less than a factor of two, however, and the rate of RNA-DNA hybrid formation com- pared to DNA-DNA reassociation will increase by only a factor of two or less. A twofold relative rate increase is not observed in these experiments, since both RNA-DNA and DNA-DNA reactions are approaching completion at the Cot’s used. Furthermore, at very high RNA concentrations the annealing solutions tend to gel below 60°C and are difficult to handle ; the result is increased scatter in the points
504 E. H. DAVIDSON AND B. R. HOUGH
representing the higher RNA:DNA ratios, where any relative increase in the rate of RNA:DNA hybridization might be expected to be observed.
Figure 4 indicates that about 0.6% of the non-repetitive sequences in the genome are represented in the RNA of the mature oooyte. This value is close to the 0.9% representation obtained at an RNA: DNA ratio of 2500 in the RNA-driven reaction described above. We are inclined to regard the value of 0.6% obtained from Figure 4 as a more reliable, minimum estimate in as much as it is based on a much larger amount of data. Nonetheless the correspondence of these two estimates is reassuring. On the basis that only one strand is transcribed, 0.6% representation means that about 1.2% of the total genomio information is present in the RNA stored in the mature Xenopus oooyte.
4. Discussion The oooyte RNA-DNA duplexes studied here display the predicted characteristics
of hybrids with non-repetitive DNA. These hybrids have high thermal stability, only 1 to 2 deg. C below that of the non-repetitive DNA-DNA duplexes, and after ribo- nuolease treatment they yield RNA of molecular size close to that of the DNA. Both of these properties depend on precise base pairing in the hybrids, as is indicated by prior studies with precisely paired hybrids between bacterial RNA and DNA (Moore & McCarthy, 1967) and ribosomal RNA and DNA (Jeanteur & Attardi, 1969). These characteristics would not be observed if the oooyte RNA-DNA hybrids were actually formed with a repetitive sequence contaminant in the non-repetitive DNA fraction. RNA-DNA hybrids involving the related but non-identical DNA sequences of repetitive sequence families include a variable amount of nuoleotide mismatch, as do most DNA-DNA duplexes formed from repetitive sequence oom- ponents (Britten & Kohne, 1966,196s; Shearer & McCarthy, 1970). These data indicate, furthermore, that the amount of repetitive sequence contamination in the non-repetitive DNA fraction is very small. Though these items of evidence strongly support the position that the RNA is hybridized with non-repetitive DNA, the independent evidence of the experiment in Table 2 is even more persuasive on this point. Here the DNA actually engaged in duplexes with RNA was trapped on hydroxy- apatite and in the presence of a 6000-fold excess of whole DNA was shown to re- associate exactly at the rate of non-repetitive DNA. Additional proof of the nature of the hybridized RNA would be afforded by recovering the hybridized RNA and studying its rate of hybridization. This is not practicable in this experimental system since the i?a vitro labeled RNA is not of high enough specific activity.
The estimate that 0.6% of the non-repetitive sequence is represented in oooyte RNA is necessarily a minimum one. As noted above the RNA-driven reaction at RNA:DNA ratio 2500 yielded a slightly higher value. Furthermore, in the experi- ments of Figure 4 some fraction of the hybrids could have been lost in the lengthy processing steps. Some exceedingly rare species of RNA might also exist which represent other DNA sequences than those whose reaction was observed, though there is little suggestion in the shape of the saturation curve of a gradually increasing spectrum of RNA frequencies. Accepting a representation of 1.2% of the genomio information in the oooyte RNA as a minimum estimate, the minimum complexity of this RNA can be expressed in terms of the nuoleotide pairs of diverse sequence present. The Xenopw genome contains about 3 x 10’ nuoleotide pairs (Dawid, 1965), of which 55% is non-repetitive at the 60°C criterion we have applied. Thus, 1.2% of the non-
GENETIC INFORMATION IN OOCYTE RNA 505
repetitive DNA represents a complexity in the RNA equivalent to about 20 x 10s nucleotide pairs.
From the very large amount of RNA required to attain saturation in Figure 4 it is clear that those species of RNA molecules responsible for the major portion of the hybridization with non-repetitive DNA are present in relatively low copy number. To saturate the repetitive sequence fraction of Xenopus DNA with exactly similar oocyte RNA preparations, RNA: DNA ratios of only 1 or 2 are needed (Davidson et al., 1968; Crippa et al., 1967). This suggests that some non-repetitive transcripts may be present at as little as 1150th the concentration of the less common repetitive sequence transcripts. It would seem most probable that different non-repetitive sequences are represented in the transcript population by different numbers of copies, though the linear rise of the curve in Figure 4 is consistent with a single frequency of representation for most of the non-repetitive sequences hybridizing. If for simplicity we assume that there is a single frequency class we can estimate how much of the oocyte RNA is transcribed from non-repetitive DNA. According to earlier estimates the population of stored heterogeneous RNA molecules, which in the oocyte consists mainly of repetitive sequence transcripts, represents about 10W2 of the RNA of the mature oocyte. If this is so, the non-repetitive sequence transcripts would account for only low4 to 10m3 of the oocyte RNA. Since each oocyte contains about 4 pg of RNA (Brown & Littna, 1964) the total number of nucleotides of RNA hybridizable with non-repetitive DNA sequences would be at least 7 x 1011 nucleotides (4 x 10m4 pg x 6 x 10z3 nucleotides/mole/3*3 x 10s pg/mole). Thus the average number of RNA copies stored in the oooyte for each active non-repetitive DNA sequence would be given by 7 x loll/20 x 106, or of the order of at least lo4 copies.
The informational complexity of the oocyte RNA is surprisingly high. Complexity of at least 20 x lo6 nucleotide pairs is equivalent to at least 4.5 times the total genomic information in the E. coli chromosome (45 x IO6 nucleotide pairs) (Cairns, 1963). The complexity of the RNA in the oocyte, though it is but a single cell type, would be sufficient to account for 20,000 diverse structural genes of 1000 nucleotide pairs each, or over 40,000 diverse genes the size of that coding for the p-chain of hemoglobin. There is only one other published study of RNA-non-repetitive DNA hybridization using higher cell material known to us, viz that of Gelderman et al. (1968,197O). These workers reported that at least 12% of the non-repetitive DNA sequences are represented in the RNA of whole fetal mouse. However, data are not as yet published regarding the informational complexity of RNA in any single somatic cell type, and we therefore do not know whether the utilization of genomic information in the oocyte is unusually large. In any case the data reported here indicate the magnitude of the task which must be accomplished by the gene regulation machinery for at least one cell type, the growing oocyte. Recognition and activation of perhaps lo* to lo5 different genomic sequences must ultimately be accomplished by the regulation machin- ery in order to specify the unique pattern of gene activity characteristic of the oocyte.
This research was supported by the American Cancer Society grant no. E334 and by the National Institute of Child Health and Development grant no. HD-02412. We are extremely grateful to Dr Jack Goldstein of the New York Blood Center for assisting us with the polyacrylamide gel determinations. We are deeply indebted to Dr Roy Britten, of the Carnegie Institution of Washington and to Dr Bruce R. Voeller of this University for many penetrating and helpful discussions and for rritical advice on this manuscript. We take great pleasure in acknowledging the skill of Mr William Viets and also of Mr Harris Huberman in manually collecting oocytes, and the intelligent and professional assistance of Miss Maggie Chamberlin.
606 E. H. DAVIDSON AND B. R. HOUGH
REFERENCES
Barth, L. G. & Barth, L. J. (1959). J. Eq. Morphol. 7, 210. Bolton, E. T. & McCarthy, B. J. (1964). J. Mol. Biol. 8, 201.
Britten, R. 5. & Kohne, D. E. (1966). Yea&. Carnegie In&n, no. 65, p. 78. Britten, R. J. & Kohne, D. E. (1968). Science, 161, 529.
Britten, R. J. & Smith, J. (1969). Yea&. Carnegie In&n, no. 68, p. 378. Brown, D. D. & Littna, E. (1964). J. Mol. Biol. 8, 669. Cairns, J. (1963). CoId SF. Had. Symp. Quant. Biol. 28, 43.
Crippa, M., Davidson, E. R. & Mirsky, A. E. (1967). Proc. Nut. Acad. Sci., Wash. 57, 885.
Crippa, M. & Gross, P. E. (1969). Proc. Nat. Acad. Sci., Wash. 62, 121.
Davidson, E. H., Crippa, M., Kramer, F. R. & Mirsky, A. E. (1966). Proc. Nat. Acud. Sci., Wash. 56, 856.
Davidson, E. H., Crippa, M. Bi Mirsky, A. E. (1968). Proc. Nut. Acad. Sci., WC&. 60, 152.
Davidson, E. H. & Hough, B. R. (1969). Proc. Nat. Acad. Sci., Wash. 63, 342.
Dawid, I. B. (1965). J. Mol. BioE. 12, 581.
Gelderman, A. H., Rake, V. A. & Britten, R. J. (1968). Yew-b. Cwnegie In&n, no. 67, p. 320.
Gelderman, A. H., Rake, V. A. dt Britton, R. J. (1971). Proc. Nut. Acad. Sci., Wmh.
in the press. Goldstein, 5. & Harewood, K. (1969). J. Mol. BioZ. 39, 383.
Gross, P. R. (1967). Current Topics in Develop. BioZ. 2, 1. Jeanteur, P. & Attardi, G. (1969). J. Mol. BioZ. 45, 305. Kedes, L. & Gross, P. R. (1969). J. Mol. Biol. 42, 559.
Kohne, D. E. (1968). Biophys. J. 8, 1104.
Masui, Y. (1967). J. Exp. Zool. 166, 365. Moore, R. L. & McCarthy, B. J. (1967). J. Bact. 94, 1066.
Shearer, R. W. & McCarthy, B. J. (1970). &o&em. Genet. 4, 395.
Smith, L. D., Armstrong, J. 1,. & McCarthy, B. J. (1967). Biochim. Zriophys. Actcc, 142, 323.
Weinberg, R. A., Loening, V. E., Willems, M. & Penman, 8. (1967). Proc. Nat. Acad. Sk., Wmh. 58, 1088.
Wetmnr, J. G. & Davidson, N. (1968). J. Mol. BioZ. 31, 349.