Pacific University CommonKnowledge Faculty Scholarship (PHRM) School of Pharmacy 2013 Genetic Manipulation of Leishmania Parasites Facilitates the Exploration of the Polyamine Biosynthetic Pathway as a Potential erapeutic Target Sigrid C. Roberts Pacific University Follow this and additional works at: hp://commons.pacificu.edu/phrmfac Part of the Pharmacy and Pharmaceutical Sciences Commons is Book Chapter is brought to you for free and open access by the School of Pharmacy at CommonKnowledge. It has been accepted for inclusion in Faculty Scholarship (PHRM) by an authorized administrator of CommonKnowledge. For more information, please contact CommonKnowledge@pacificu.edu. Recommended Citation Roberts, S. C. (2013). Genetic manipulation of Leishmania parasites facilitates the exploration of the polyamine biosynthetic pathway as a potential therapeutic target. In K.V. Urbano (Ed.), Advances in Genetics Research, Volume 10 (29-54). Hauppauge, NY: Nova Science Publishers.
Transcript
Pacific UniversityCommonKnowledge
Faculty Scholarship (PHRM) School of Pharmacy
2013
Genetic Manipulation of Leishmania ParasitesFacilitates the Exploration of the PolyamineBiosynthetic Pathway as a Potential TherapeuticTargetSigrid C. RobertsPacific University
Follow this and additional works at: http://commons.pacificu.edu/phrmfac
Part of the Pharmacy and Pharmaceutical Sciences Commons
This Book Chapter is brought to you for free and open access by the School of Pharmacy at CommonKnowledge. It has been accepted for inclusion inFaculty Scholarship (PHRM) by an authorized administrator of CommonKnowledge. For more information, please [email protected].
Recommended CitationRoberts, S. C. (2013). Genetic manipulation of Leishmania parasites facilitates the exploration of the polyamine biosynthetic pathwayas a potential therapeutic target. In K.V. Urbano (Ed.), Advances in Genetics Research, Volume 10 (29-54). Hauppauge, NY: NovaScience Publishers.
Genetic Manipulation of Leishmania Parasites Facilitates the Explorationof the Polyamine Biosynthetic Pathway as a Potential Therapeutic Target
AbstractParasites of the genus Leishmania cause a variety of devastating and often fatal diseases, ranging fromcutaneous ulcerative lesions to fatal visceralizing infections that affect an estimated 12 million peopleworldwide. Unfortunately, vaccines are not available and the cturent arsenal of drugs used to treatleishmaniasis is far from ideal. Thus the need for new therapeutic targets and a better understanding of host-parasite interactions is urgent. One biochemical pathway that has been successfully exploited for the treatmentof a related parasitic disease, African trypanosomiasis, is the polyamine biosynthetic pathway. In order toelucidate the polyamine biosynthetic pathway and to explore its potential as a therapeutic target inLeishmania, we have generated and characterized gene deletion mutants and polyamine enzyme overproducerstrains. These studies revealed that the polyamine pathway in Leishmania is significantly different from that ofthe mammalian host and polyamines were found to be essential for parasite proliferation. Infectivity studies inmacrophages and mice with gene deletion mutants revealed that at least two polyamine biosynthetic enzymes,ornithine decarboxylase and spermidine synthase, are necessary for Leishmania donovani to establish asuccessful infection. However, arginase gene deletion mutants of Leishmania mexicana are still capable ofeliciting an infection, albeit at lower levels than wild type parasites. Ongoing studies address whether thedisparities in infectivity are due to the loss of specific enzymes within the pathway or to differences betweenthe two Leishmania species. Furthermore, the gene deletion mutants are useful tools to investigate the relativecontribution of host and parasite polyamine biosynthetic enzymes in parasite infectivity. This chapter willsummarize how genetic manipulations in Leishmania have advanced our understanding of the polyaminepathway and its role in host-parasite interactions.
The license for this PDF is unlimited except that no part of this digital document may be reproduced, stored in a retrieval system or transmitted commercially in any fonn or by any means. The publisher has taken reasonable care in the preparation of this digital doc\llllent, but makes no expressed or implied warranty of any kind and asswnes no responsibility for any errors or omissions. No liability is asswned for incidental or consequential damages in connection with or arising out of information contained herein. This digital document is sold with the clear understanding that the publisher is not engaged in rendering legal, medical or any other professional services.
Chapter II
Genetic Manipulation of Leishmania Parasites Facilitates the Exploration
of the Polyamine Biosynthetic Pathway As a Potential Therapeutic Target
Sigrid C Roberts* Pacific University School ofPhrumacy, Hillsboro, Oregon, US
Abstract
Parasites of the genus Leishmania cause a variety of devastating and often fatal diseases, ranging from cutaneous ulcerative lesions to fatal visceralizing infections that affect an estimated 12 million people worldwide. Unforttmately, vaccines are not available and the cturent arsenal of drugs used to treat leishmaniasis is far from ideal. Thus the need for new therapeutic targets and a better tmderstanding of host-parasite interactions is urgent. One biochemical pathway that has been successfully exploited for the treatment of a related parasitic disease, African trypanosomiasis, is the polyamine biosynthetic pathway. In order to elucidate the polyamine biosynthetic pathway and to explore its potential as a therapeutic target in Leishmania, we have generated and characterized gene deletion mutants and polyamine enzyme overproducer strains. These studies revealed that the polyamine pathway in Leishmania is significantly different from that of the mammalian host and polyamines were fotmd to be essential for parasite proliferation. Infectivity studies in macrophages and mice with gene deletion mutants revealed that at least two polyamine biosynthetic enzymes, omithine decarboxylase and spemlidine synthase, are necessary for Leishmania donovani to establish a successful infection. However, arginase gene deletion mutants of Leishmania mexicana are still capable of eliciting an infection, albeit at lower levels than wild type parasites. Ongoing studies address whether the disparities in infectivity are due to the loss of specific enzymes within the pathway or to differences between the two Leishmania species. Furthe1more, the gene deletion mutants are useful tools to investigate the relative
• Com:sponding author: Sigrid C. Roberts. Pacific University School of Phannacy, 222 SE gth Avenue, Hillsboro, OR 97123, E-mail: [email protected].
30 Sigrid C. Robe1ts
contribution of host and parasite polyamine biosynthetic enzymes in parasite infectivity. This chapter will smmnarize how genetic manipulations in Leishm.ania have advanced om tmderstanding of the polyamine pathway and its role in host-parasite interactions.
Leishmania Parasites and Their Relevance for Human Health
Parasites of the genus Leishmania cause a variety of devastating and often fatal diseases in humans and domestic animals worldwide. Leishmania belong to the order of trypanosomatids, which include Trypanosoma brucei, the causative agent of African sleeping sickness, and Trypanosoma cruzi, which produces Chagas disease. The spectmm of leishmaniasis ranges from cutaneous ulcerative lesions to fatal visceralizing infections and affects an estimated 12 million people worldwide [1 , 2]. Visceral leishmaniasis targets primarily the liver and spleen and is caused by L. donovani, L. infantum (Africa and Asia), and L. chagasi (Central and South America). This type of leishmaniasis is invariably fatal if not treated. Cutaneous leishmaniasis, which produces skin ulceration at the bite site, is caused by about 20 different Leishmania species, including L. mexicana, L. major, and L. amazonen.sis. A third f01m of the disease, mucocutaneous leishmaniasis, which targets and destroys the mucous membranes of nose and mouth, is caused by L. braziliensis in South Ame1ica. The specific host tissue tropism of the different Leishmania species is poorly understood and exceptions occur. For example, L. chagasi may lead to cutaneous leishmaniasis in Central America, while do1mant L. donovani parasites can trigger cutaneous disease years after the visceral f01m has been successfully treated as obse1ved in post kalaazar de1mal leishmaniasis (PKDL) in India and Africa [3, 4].
Leishmania has a digenetic life cycle in which the extracellular promastigote f01m resides in the sand fly vector, whereas the intr·acellular amastigote f01m inhabits the phagolysosome of macrophages in the infected host. The flagellated promastigote resides in the gut of the sand fly but will eventually migrate to the proboscis of the insect. Here, metacyclic or infective promastigotes will be regurgitated into the bite site when the sand fly takes a blood meal. Leishmania promastigotes are then phagocytosed by host macrophages. Usually, phagosomes fuse with lysosomes to f01m phagolysosomes, which are a highly effective in killing foreign pathogens. However, Leishmania parasites inhibit phagolysosomal maturation and withstand the hostile environment. Promastigotes conve1t into the smaller, non-flagellated amastigotes, which proliferate inside this host cell compartment. When a sandfly takes a blood meal from an infected host, it ingests parasitized macrophages and the released amastigotes tr·ansfOim back into promastigotes, completing the life cycle.
Due to the absence of effective vaccines, therapeutic treatment has offered the only avenue of defense against leishmaniasis and other parasitic diseases [5-7] . Dmg treatment of the different fo1ms of leishmaniasis, however, is far from ideal with toxic side effects and dmg resistance limiting the effectiveness of a small number of dmgs. The toxic antimonials are still considered the first line of treatment and the only new dmg, miltefosine, which is also the only anti-leishmania! medicine that can be administered orally, is teratogenic [8]. Thus, the need for new therapeutic targets and a better tmderstanding of host-parasite interactions is urgent.
Genetic Manipulation of Leishmania Parasites ... 31
The Polyamine Pathway
One pathway that has ah·eady been targeted for parasitic diseases is that of polyamine biosynthesis. Difluoromethylornithine (DFMO or eflomithine), which targets the first and rate-limiting step of tlris pathway, is a cllirically approved treatment for Afi.ican sleeping sickness caused by T. brucei [9-12], a parasite closely related to Leishmania. Polyamines are essential and ubiquitous cations that are especially important for rapidly proliferating cells such as parasites and cancer cells. The three biologically relevant polyanrines, putrescine, spemridine, and spemrine have an aliphatic carbon backbone and contain multiple amino groups that are positively charged under physiological conditions (Figure 1). These compotmds play c1itical roles in key cellular processes such as growth, differentiation, and macromolecular biosynthesis. However, despite numerous studies on the ftmctions of polyamines, their roles for cell stmcture and ftmction are still not fully tmderstood.
The polyamine pathway has been characte1ized in detail in the mammalian host (Figure 2). The first and rate-linriting enzyme is omithine decarboxylase (ODC), which catalyzes the conversion of the anrino acid omithine to putrescine. Subsequently, putrescine is metabolized to spemridine by the action of spernridine synthase (SPD). This enzyme catalyzes the addition of an aminopropyl group, which is donated fi:om decarboxylated S-adenosyhnethionine. The enzyme S-adenosyhnetlrionine decarboxylase (ADOMETDC) is responsible for the decarboxylation of S-adenosylmethionine, which comnrits the compotmd for polyamine biosynthesis. A second aminopropylgroup is added to spemridine to produce spemrine in a reaction catalyzed by spemrine synthase (SPM). In mammals, spemrine can be backconveited to spemridine and putrescine by the conceited actions of spemridine/spemrine N(l)-acetyltransferase (SSAT) and polyamine oxidase (POA). Spennidine is an essential polyamine that also plays a vital role in a unique modification reaction that is necessruy to activate eukruyotic translation initiation factor (eiFSA), a protein involved in translation and RNA metabolism [13, 14]. The enzyme deoxyhypusine synthase (DHS) catalyzes the addition of an aminopropyl group derived from spernridine to a lysine residue of eiFSA to f01m deoxyhypsuine and the enzyme deoxyhypusine hydroxylase (DOHH) completes the reaction by f01ming hypusine.
. ~ ~ .,:-.~H • N II t , """" """" . -'
Putrescine
~ ~ HJ• NHJ•r ~ 'NIIl+~
Spermidine
Nil + '1111 .. KI!3+~Nlll~ 1~ J
Spermine
Figme 1. Structures of the common polyamines.
32 Sigrid C. Robe1ts
Arclnlne
11 Spermine
Hypu$1n.ated eiFSA
The enzyme abbreviations utilized are ARG: arginase; ODC: omithine decarboxylase; SPD: spe1midine synthase; SPM: spennine synthase; ADOMETDC: S-adenosyhnethionine decarboxylase; ADC: arginine decarboxylase; AGM: agmatinase; DHS: deoxyhypusine synthase; DOHH: deoxyhypusine hydroxylase. DFMO is difluoromethylomithine, a suicide inhibitor of ODC. The question mark denotes the controversial hypothesis that mammalian cells contain an ADC activity.
Figme 2. The polyamine biosynthetic pathway in the mammalian host.
Evidence from yeast and mammalian cells suggest that the hypusination reaction is indispensable for activation of eiF5A and cell viability [15, 16].
Spe1midine is also used in the synthesis of tlypanothione, a reaction that occms only in tlypanosomatid parasites. T1ypanothione is composed of two molecules of glutathione linked by spe1midine and plays a cmcial role in fighting oxidative stress in these pathogens [17-21]. Although tlypanothione metabolism is an essential and tmique pathway for tlypanosomatids, this therapeutically relevant pathway has been the subject of several recent review articles [22-25] and will therefore not be discussed ftuther in this chapter.
An altemative pathway for pun·escine production exists in plants and some microorganisms where ru·ginine decarboxylase (ADC) conve1ts the amino acid arginine to agmatine, which is ftuther metabolized to putrescine by agmatinase (AGM) or agmatine deiminase and N-cru·bamoylputrescine hydrolase. While the presence of AGM in manunalian cells has been desc1ibed and is generally recognized, the existence of an ADC is disputed [26-28].
Because polyamines ru·e a validated tru·get in the related African tlypanosomes, we used genetic methods to elucidate this pathway in Leishmania with the goal to evaluate its potential as a therapeutic tru·get.
Genetic techniques Available for the Manipulation of Leishmania parasites
Genetic manipulations can be perfo1med in Leishmania and facilitate the characterization of metabolic processes and the identification of proteins that ru·e impo1tant for pru·asite
Genetic Manipulation of Leishmania Parasites ... 33
proliferation and infectivity. The most significant tool is the ability to generate gene deletion mutants via targeted gene replacement. Leishmania are diploid organisms that readily tmdergo homologous recombination and knockout strategies are hence straightf01ward. Gene deletion sn1dies are an example of reverse genetic analysis where the outcome or phenotype of a targeted mutation is examined. The more classical method, fo1ward genetic analysis, attempts to discover the genetic basis of an obse1ved phenotype.
In order to generate gene deletion mutants, constructs containing a dmg resistance cassette flanked by upstr·eam and downstream sequences of the target gene need to be created (Figure 3). Linea1ized DNA constructs are then purified and intr·oduced into the parasite by electr·oporation. Transfected parasites are subsequently plated on semi-solid agar plates in the presence of the selective dmg and colonies are picked to achieve the genetic homogeneity of a clonal cell line. Because Leishmania are diploid organisms, a second round of targeted gene replacement is necessary to create knockout par·asites. Loss of heterozygosity can also be induced, however, a negative selection scheme is necessa1y to stimulate the loss of the second gene copy. Southem blotting and/or polymerase chain reactions (PCR) are utilized to char·acterize the resulting genotype and to ve1ify that the anticipated homologous recombination event occmTed in both heterozygous and homozygous par·asites.
The deletion of an essential gene is only possible if the mutants can be rescued by nutr·ient supplementation or other salvage str·ategies. For example, the removal of polyamine biosynthetic pathway genes from Leishmania is possible because the auxotrophic mutants smvive in the presence of exogenous polyamines. Knockouts of essential genes callllot be perfo1med if rescue str·ategies ar·e unavailable.
A Electroporation C Selection
s Q ~
Homologous recombination and gene replacement
~
~
D Expansion
Confirmation of genotype by Southern blot or PCR
0 ....,_, ....
AReplacement constructs are introduced into parasites via electroporation. BThe target gene is replaced with the dmg resistance cassette when homologous recombination occms. <1>arasites are plated on semi-solid agar plates containing the selective dmg and colonies are picked. °Colonies are expanded in liquid media. TI1e genotype is confmned by Southem blotting or PCR analysis. The process is subsequently repeated to replace the second gene with a different dmg resistance marker.
Figme 3. Targeted gene replacement.
34 Sigrid C. Robe1ts
However, the ability to generate chromosomal gene deletions in the presence, but not absence, of episomes that express the targeted gene is one way to demonstrate that a gene is indispensable [29, 30].
An important control experiment is to generate complemented or add-back strains, where the deleted gene is re-introduced into the knockout strain either on an episome or as a chromosomal integration [31-38]. Compmison of the knockout cell line and the complemented strain ensure that the obse1ved phenotypic changes are indeed caused by the gene deletion event and not by other sporadic mutations or adaptations that might have OCCUlTed.
The opposite of gene deletion, the introduction of genes into Leishmania pm·asites, is also possible. Constlucts containing endogenous genes can be inn·oduced to generate ove1producer sn·ains and the phenotypic characte1ization of the resultant mutant can provide valuable info1mation about the target gene. Fmthe1more, genes from different species can be n·ansfected to create n·ansgenic pm·asites. Endogenous or exogenous genes can be inn·oduced as pa1t of an episome or stably integrated into the chromosome. Advantages of these expression systelllS range from the introduction of repo1ter genes to facilitate imaging of fusion proteins or whole organisms in in vivo infections [39-41], expression and pmification of recombinant proteins [42-44], and ftmctional studies that help to identify genes involved in dmg sensitivity, protism, vimlence or the host immune response [ 45-48].
It is impo1tant to note that the cultivation of Leishmania pm·asites is straightfmwm·d. Exn·acellular parasites can be maintained as the promastigote (insect) fmm indefinitely and in large quantities in liquid culture medium. Amastigotes can also be cultured as an exn·acellular or axenic fmm by utilizing acidic media and higher temperatures, which mimic the host phagolysosomal environment. Although axenic amastigotes m·e only a model system and not t111e inn·acellular amastigotes, the advantage is that pm·asite metabolism can be nun·itionally and biochemically characterized without the contamination of host components. Fmthe1more, infectivity studies in vitro in macrophages as well as in vivo in mice and halllSters are now routinely perfo1med.
The Polyamine Pathway in Promastigotes
We initiated our analysis of the polyamine pathway in Leishmania by generating gene deletion mutants in L. donovani promastigotes. Three knockout lines, LdAodc, LdAspd, and LdAadometdc, were created [32, 33, 35]. In addition, complemented conn·ol cell lines were generated that contained copies of the deleted genes on episomes; LdAodc [ODC}, LdAspd [SPD}, LdAadometdc [ADOMETDC]. The knockout mutants were not able to produce pun·escine (LdAodc) or spe1midine (LdAodc, LdAspd, LdAadometdc) confuming that no altemative pathways for polyamine production m·e present in Leishmania parasites. As expected, the genetically manipulated promastigotes require polyamine supplementation for smvival. The LdAodc parasites are able to grow in spe1midine; however, much higher concenn·ations of spe1midine compared to pun·escine are required [32].
Thus, spe1midine is sufficient and essential for promastigote proliferation; but pun·escine is necessa1y for optimal proliferation. This obse1vation suggests that pun·escine is not merely a precursor for spe1midine synthesis but plays an additional and impo1tant role in Leishmania.
Genetic Manipulation of Leishmania Parasites ... 35
Polyrunine pool analysis of pru·asites revealed that spennine is not being produced by the parasite (Figure 4) and a spennine synthase gene is not present in the annotated genome databases of Leishmania species. This is an unexpected discove1y, as spennine is postulated to be a crucial metabolite in mammalian cells [49].
Transp01ters for arginine (AAP3) and for putrescine/spemlidine (POTl) have been identified on the molecular level. Ornithine transp01t has been characterized. Individual transpotters for putrescine and spennidine have been proposed. Parasite ARG is located in the glycosome, necessitating import of arginine into the glycosome and expoti of omithine from the glycosome to the cytosol. ODC, SPD, and ADOMETDC (not shown) are located in the cytosol.
Figure 4. Polyrunine biosynthesis and transpoti in Leishmania promastigotes.
Analysis of the knockout parasites also revealed that the back·conversion pathway present in mammalian cells is lacking in Leishmania (Figure 4). Spemtidine cannot be conve1ted to putrescine. Sintilarly, spemrine, although taken up by pru·asites, is not ftuther metabolized or utilized [32]. Fmthe1more, the half·live of the leishmania! ODC and ADOMETDC proteins ru·e much longer than that of their human counterpruts [35, 50]. This is a relevant obse1vation since the selectivity of the suicide inhibitor DFMO for T brucei is based on the fact that ODC is a stable enzyme in the pru·asite but is rapidly tmned over in the human host; DFMO binds to human and pru·asite ODC with sinrilar affmities [51] but the human protein is re·synthesized continuously.
More recently, investigations into the hypusine pathway have been initiated in protozoan pathogens [29, 52-56]. Leishmania pru·asites contain two putative DHS genes (DHSL20, DHS34) and one putative DOHH gene [29, 52]. Biochenrical characterization of the proteins encoded by the two putative DHS genes revealed that one protein (DHSL20) was not ftmctional as a DHS but the other protein, DHS34, expressed the expected catalytic activity [29]. Unf01tunately, gene deletion mutants of DHS or DOHH cannot be generated if the enzymes ru·e essential, because rescue by supplementing modified eiF5A is not possible. As expected, a DHS null mutant could not be produced. However, chromosomal knockouts were generated in cell lines that cruTied an episome containing the DHS coding sequence [29]. This
36 Sigrid C. Robe1ts
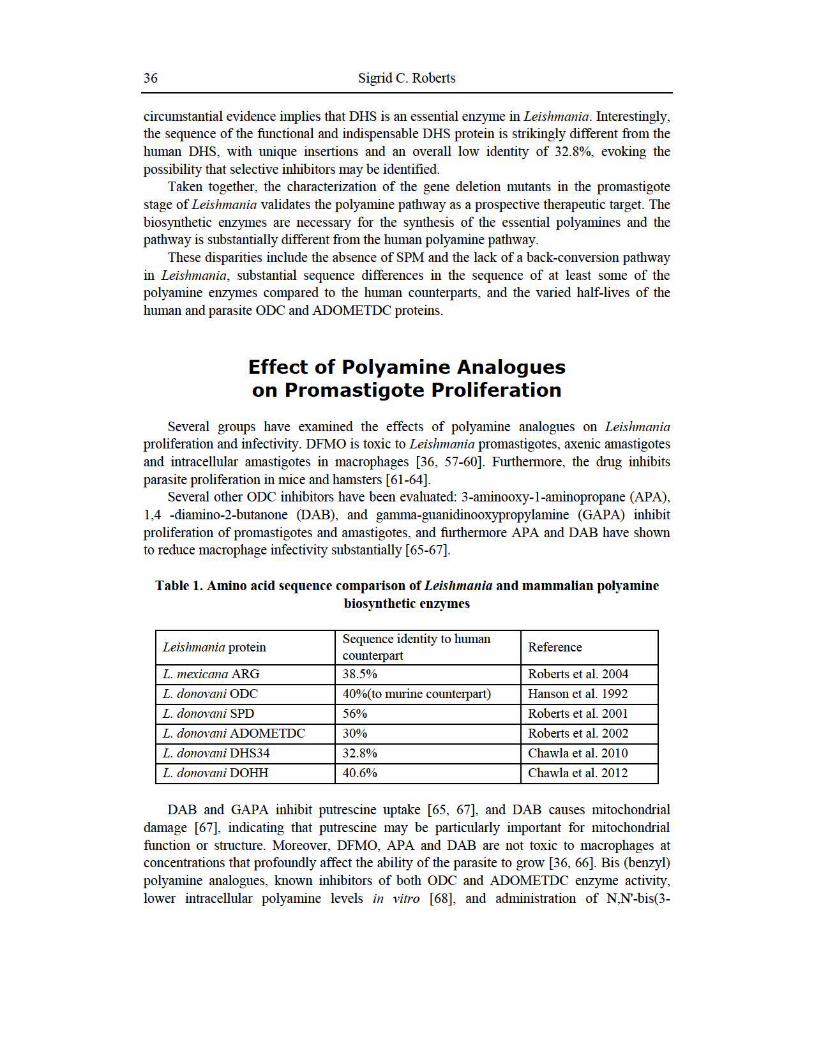
circumstantial evidence implies that DHS is an essential enzyme in Leishmania. Interestingly, the sequence of the ftmctional and indispensable DHS protein is strikingly different from the human DHS, with unique inse1t ions and an overall low identity of 32.8%, evoking the possibility that selective inhibitors may be identified.
Taken together, the characterization of the gene deletion mutants in the promastigote stage of Leishmania validates the polyamine pathway as a prospective therapeutic target. The biosynthetic enzymes are necessary for the synthesis of the essential polyamines and the pathway is substantially different fi"om the human polyamine pathway.
These dispa1ities include the absence of SPM and the lack of a back-conversion pathway in Leishmania, substantial sequence differences in the sequence of at least some of the polyamine enzymes compar·ed to the human cmmterpa1ts, and the varied half-lives of the human and par·asite ODC and ADOMETDC proteins.
Effect of Polyamine Analogues on Promastigote Proliferation
Several groups have examined the effects of polyamine analogues on Leishmania proliferation and infectivity. DFMO is toxic to Leishmania promastigotes, axenic amastigotes and intracellular· amastigotes in macrophages [36, 57-60]. Fmthe1more, the dmg inhibits par·asite proliferation in mice and hamsters [61-64].
Several other ODC inhibitors have been evaluated: 3-aminooxy-1-aminopropane (APA), 1,4 -diamino-2-butanone (DAB), and gamma-guanidinooxypropylamine (GAPA) inhibit proliferation ofpromastigotes and amastigotes, and ftnthe1more APA and DAB have shown to reduce macrophage infectivity substantially [ 65-67].
Table 1. Amino acid sequence comparison of Leisltmallia and mammalian polyamine biosynthetic enzymes
Leishmania protein Sequence identity to human cotmterpatt
Reference
L. mexicana ARG 38.5% Roberts et al. 2004
L. donovani ODC 40%(to murine counterpart) Hanson et al. 1992
L. donovani SPD 56% Roberts et al. 2001
L. donovani ADOMETDC 30% Roberts et al. 2002
L. donovani DHS34 32.8% Chawla et al. 201 0
L. donovani DOHH 40.6% Chawla et al. 2012
DAB and GAPA inhibit putrescine uptake [65, 67], and DAB causes mitochondrial damage [ 67], indicating that putrescine may be pa1ticular·ly impo1tant for mitochondlial ftmction or stmctme. Moreover, DFMO, APA and DAB ar·e not toxic to macrophages at concentrations that profmmdly affect the ability of the par·asite to grow [36, 66]. Bis (benzyl) polyamine analogues, known inhibitors of both ODC and ADOMETDC enzyme activity, lower intracellular· polyamine levels in vitro [68], and administration of N,N'-bis(3-
Genetic Manipulation of Leishmania Parasites .. . 37
[(phenylmethyl) amino] propyl)-1,7-diaminoheptane (MDL27695) suppresses L. donovani liver burden in mice by 50% [63] or 99% [69]. Recently, the stmcture of the leishmania! ODC bmmd to AP A has been modeled according to the solved crystal stmcnu·e of the hmnan ODC - APA complex [70]. The stmcnu·e, kinetic propetties, and half-life of the leishmania! ODC are substantially different from the hmnan cotmterprut [50, 70] and the stmcnu·al information may prove useful for the design or computer-based screening of novel ODC inhibitors.
Overproducer Strains As Tools to Investigate the Mode of Action of Polyamine Analogues
To test whether polyamine analogues indeed target enzymes of the polyamine pathway, a cell based screening assay has been developed that takes advantage of polyamine enzyme overproducing strains. The genetically manipulated pathogens are used as tools to investigate the mode of action of cytotoxic analogs in Leishmania based on the prediction that enzyme overproducer strains will be more resistant to a dmg that specifically targets the overproduced protein. Wild type pru·asites were transfected with episomal constmcts containing the ODC, SPD, or ADOMETDC gene. The episomal constmcts also contained dmg resistance markers that allowed selection of transfected parasites. Fmthetmore, increased dmg concentrations facilitated the overexpression of the polyrunine biosynthetic proteins, presumably due to the high copy number of the episomes. Wild type and oveiproducer strains were then incubated in serial inhibitor dilutions and the EC [50] values were compared. These expetiments confumed that DFMO, 5-([(Z)-4-amino-2-butenyl] methylamino)-5-deoxyadenosine (AbeAdo or MDL73811), and n-butylamine tru·get ODC, ADOMETDC, and SPD, respectively, as had previously been proposed [71-73]. Conversely, pentrunidine, berenil, and methylglyoxal bis (guanylhydrazone) (mitoguazone or MGBG) did not kill pru·asites via polyrunine enzyme inhibition. These compounds have previously been speculated to be toxic solely due to inhibition of the polyamine biosynthetic enzymes but other cellulru· tru·gets have also been proposed [74-80]. The overproducer strains were utilized for a pilot screen of 25 compmmds and we demonstrated that this system offers a rapid cell-based screen for assessing whether synthetic polyrunine analogs exett their toxicity predominantly by targeting the polyamine biosynthetic enzymes in Leishmania. Ultimately, the dmg resistance induced by amplification of tru·get genes and resulting overproduction of the encoded protein offers a general strategy for evaluating the mode of action of therapeutic agents.
Relevance of the Polyamine Biosynthetic Pathway for Leishmania Infectivity
Although stt1dies in promastigotes were promising, the ultimate validation of the polyrunine pathway as a medical target requires the evaluation of the infective runastigote stage of the pru·asite. A long debated but tmsolved question remained whether amastigotes were capable of salvaging polyamines from the phagolysosomal comprutment in macrophages. On the one hand, sh01t tetm infectivity studies in mice and hrunsters revealed
38 Sigrid C. Robe1ts
substantial growth inhibition of intracellular parasites with administration ofDFMO, although parasites were not eradicated [61-64]. On the other hand, the phagolysosome has been posnllated to be lich in nuuients, including polyamines [81-84], and polyamine uptake has been demonstrated in both the promastigote and amastigotes fmm [85-88]. Thus, the prevailing belief has been that Leishmania parasites reside in a polyamine rich environment and that the polyamine biosynthetic pathway would therefore not be a promising therapeutic target. Gene deletion mutants present an ideal tool to solve this conu·oversy since these organisms are not able to synthesize endogenous polyamines and thus depend on salvage of polyarnines for proliferation.
When peritoneal macrophages were infected with wild type and LdAodc parasites, a substantial reduction in pathogen per macrophage were obse1ved 72 hours after infection [36]. These results were minored in murine infectivity studies that revealed a profound reduction in parasite number in both liver and spleen after 4 weeks of infection. The average number of parasites per gram liver was ~ 108 in mice infected with wild type L. donovani but only ~102 parasites in mice infected with LdAodc cell lines [36]. Adminisu·ation ofpuu·escine to the dlinking water of the infected mice partially restored the loss of infectivity of LdAodc par·asites, demonsu·ating that the reduced inu·acellular· smvival was indeed due to the deletion of ODC and lack of endogenous puu·escine synthesis [ 64]. Similar results were obtained when LdAspd par·asites were used in mmine infectivity studies [38]. However, a slightly higher number of LdAspd par·asites were recovered from the livers of infected mice ( ~ 103 organisms per gram liver) compared to LdAodc (~102 organisms per gram liver). The cause for this dispa1i ty in infectivity phenotype between the LdAodc and LdAspd par·asites is unclear· but could be due to differences in either the am01mt of available puu·escine versus spe1midine in the phagolysosomal compa1tment or differential uptake abilities of inu·acellular· amastigotes. These infectivity studies demonsu·ated that ODC and SPD ar·e essential for a robust infection and that insufficient amounts of polyamines ar·e present in the phagolysosomal compa1tment. Because L. donovani par·asites cannot salvage sufficient amounts of puu·escine or spe1midine, the polyamine biosynthetic enzymes ODC, SPD, and ADOMETDC are potential targets for therapeutic u·eatment ofvisceral leishmaniasis.
The Role of Parasite Arginase for Promastigote Proliferation
In Leishmania, arginase (ARG) conve1ts the essential amino acid ar·ginine to omithine, which is then directly funneled into polyamine biosynthesis [89]. ARG gene deletion mutants have been generated in L. mexicana, L. major, and L. amazonensis [89-91]; all three are agents of cutaneous leishmaniasis. Stlldies in the promastigote stage of the par·asite dete1mined that the sole essential role of ARG is the synthesis of omithine for polyamine biosynthesis and that the gene deletion mutants required omithine or puu·escine supplementation for smvival [89-91]. These sn1dies in the promastigote stage of the parasite establish the par·asite ARG as an essential enzyme for polyamine biosynthesis.
Intriguingly, the leishmania! ARG sequence contains a SKL motif, which directs the enzyme to the glycosomal compartment, while the remaining polyamine biosynthetic enzymes ar·e localized to the cytosol [89]. The glycosome, an essential cellular· organelle
Genetic Manipulation of Leishmania Parasites .. . 39
unique to Leishmania and similar parasites, is evolutionaty related to the peroxisomes of mammalian cells. hl Leishmania, glycosome biogenesis is essential and the organelle houses several cmcial metabolic and biosynthetic pathways; however, it is unknown why these pathways are restricted to the glycosome [92, 93]. To investigate the importance of the glycosomal localization for ARG ftmction, a SKL deletion mutant was expressed in the L. mexicana flarg backgrotmd [89]. Phenotypic characterization confnmed that the mutant protein was expressed at similar levels as the wild type protein and that it was tnislocalized to the cytosol. Proliferation studies revealed that the glycosomal localization of ARG was not necessruy for its ftmction in L. mexicana promastigotes [89]. Similar expetiments in L. amazonensis , however, found that a mislocalized ru·g!!..skl protein was expressed at only vety low levels in the cytosol, indicating that proper localization is significant in L. amazonensis promastigotes [90]. However, the promastigotes were able to smvive without omithine or putrescine supplementation, presumably due to residual ru·ginase activity of the ru·g!!..skl protein [90].
Another intriguing obsetvation is that although omithine is the direct product of the ARG reaction, much higher concentrations of exogenous omithine (>1mM) than putrescine (5- 10 JlM) ru·e necessruy to supp01t optimal proliferation of L. mexicana flarg promastigotes [89]. Similar discrepancies between omithine and putrescine requirements were obsetved in L. major flarg promastigotes [91]. Uptake assays established that transp01t capabilities are similru· for omithine and putrescine in L. mexicana and thus do not accotmt for the disparity in nutrient requirements [89]. It is feasible, that ODC is the rate limiting enzyme for polyamine biosynthesis and the finding that L. me.:-cicana parasites contain much higher levels of intracellulru· omithine (~130 nmol/107 parasites) than putrescine (~2.5 nmol/107 parasites) supp01t s this conjectme [89]. The obsetvation that parasites contain elevated pools of omithine also offers a hypothesis for the comprutmentalization of ARG to the glycosome. Since the pru·asites generate a high atnotmt of omithine, cytosolic ru·ginine would be rapidly depleted and not available for protein synthesis if ARG were localized to the cytosol.The expetiments desctibed above were perf01med in ru·ginine tich cultme media, which may explain why pru·asites proliferated well despite a mislocalized ru·g!!..skl enzyme.
The Leishmania genome also contains a putative AGM gene (L. mexicana CBZ27141.1); however, extensive genetic and biochemical expetiments have demonstrated that this gene does not encode a ftmctional AGM or ARG, and targeted gene deletion studies revealed that the gene product is not an essential protein (Riley, Robetts, and Ullman, personal obsetvations). hlterestingly, an 01t holog of this putative AGM sequence is also present in the genomes of the other two tiypanosomatids, T. brucei and T. cntzi. The ADC/AGM altemative pathway for pun·escine production clearly does not exist in Leishmania as LJodc null mutants ru·e not viable without pun·escine supplementation [32].
The Role of Host and Parasite Arginase for Infectivity
Mammalian host cells contain two types of ARG: type I, which is cytosolic and expressed at high levels in the liver, and type II, which is located in the mitochondria [94]. hl macrophages arginine is a key amino acid for two competing pathways: arginine can be
40 Sigrid C. Robe1ts
conve1ted to omithine by the action of ARG or altematively to the potent anti-leishmania! agent nitric oxide by the inducible nitric oxide synthase (iNOS) [95, 96]. Murine infectivity models have documented an increased expression of ARG I in susceptible Balb/c mice associated with a TH-2 cell response and increased parasite proliferation [95, 97-101]. Conversely, the expression of iNOS has been conelated with a TH-1 cell response and decreased parasite loads in the Leishmania resistant CBA mouse model [97, 102, 103]. Numerous experiments have now conelate.d increased ARG I activity with augmented parasite loads [96, 97, 100]. In addition, phannaceutical inhibition of host ARG has been fmmd to reduce parasite numbers in macrophages and 1nice [97, 98, 100]. Thus, the mammalian ARG has been fl1mly established as a key factor for Leishmania infections [97-101, 104-108]. It is not completely understood how increased levels of host ARG I cause or contribute to disease exacerbation. One effect of higher ARG activity may be a depletion of arginine levels, which could reduce the production of nitric oxide. A local reduction of arginine has also been shown to impair the development ofT cells leading to suppression of the immune response and increased parasitemia [ 109-111]. Fmthe1more, it has been speculated that increased ARG activity enhances the levels of host polyamines, which may be salvaged by intracellular parasites and thus increase their proliferation [96, 100, 101].
To dete1mine the role of parasite ARG for infectivity, in vitro and in vivo infectivity studies have been perfmmed with all three b.arg strains (L. me;-cicana, L. major, and L. amazonen.sis b.arg) [37, 90, 91 , 112]. In vitro infectivity studies established that both L. mexicana wild type and b.arg parasites were able to scavenge omithine and putrescine from the phagolysosome of macrophages incubated in supplemented media. However, investigations in BALB/c mice demonstrated that infectivity with L. mexicana b.arg was markedly reduced compared to wild type parasites. [37] Similarly, the L. major b.arg and L. amazonen.sis b.arg deficient strains exhibited a reduced infectivity phenotype in mice [90, 91, 112]. The diminished infectivity of the L. me;-cicana b.arg parasites appeared to conelate with an increased potential to produce nitric oxide by macrophages infected with L. mexicana b.arg parasites compared to macrophages infected with wild type parasites [37]. Fmt he1more, no difference in infectivity was obse1ved between L. mexicana wild type and b.arg parasites in iNOS-deflcient macrophages. One possible explanation for the obse1vation that L. mexicana b.arg parasites are less infective is that the ARG deficient parasites cannot use arginine for polyamine production, and consequently have a reduced uptake and need of host arginine. This, in tum, would leave the host cell with more arginine available for the synthesis of nitlic oxide. Such a conjecture is also plausible because studies have demonstrated that the level of the L. donovani LdAAP3 arginine transpo1ter and thus, arginine uptake, is regulated by intracellular arginine levels [113]. In addition, it is likely that arginine levels in macrophages are limited since they are govemed solely by availability and uptake from the plasma. Indeed, macrophages that lack the arginine transpo1ter mCAT2B exhibit reduced levels of both polyarnines and nitric oxide [96, 114]. Thus, intracellular amastigotes and the host compete for a limited arginine pool and the balance between parasite and host metabolism of arginine may be cmcial in dete1mining the outcome of leishmania! infections.
Intt·iguingly, a conelation between reduced infectivity and increase nitlic oxide production was not fmmd in macrophages infected with L. major b.arg, suggesting that differences in the various cutaneous Leishmania species and their interactions with the mammalian host exist. Overall, a rational explanation for the reduced infectivity of the different cutaneous b.arg Leishmania stt·ains is that although omithine or putt·escine can be
Genetic Manipulation of Leishmania Parasites .. . 41
scavenged from the phagolysosome, endogenous biosynthesis produces more of these essential nutrients.
Taken together, these studies suggest impo11ant roles for both host and parasite arginase for optimal parasite infections. Thus, although inhibition of parasite arginase alone would not be a sufficient therapeutic strategy, the dual inhibition of host and parasite arginase may provide a novel treatment strategy. Inhibition of both enzymes would prevent polyamine biosynthesis and may furthe1more stimulate host defense mechanisms like nitric oxide production. The amino acid sequence of the Leishmania and human ARG enzyme is ~38% identical and the kinetic and biochemical characteristics of the recombinant enzymes are similar [115]. A preliminruy screen identified inhibitors that target both recombinant enzymes [115], supp01t ing the concept that dual inhibition of human and pru·asite enzyme may be possible.
Polyamine Uptake by leishmania Parasites
Both promastigotes and amastigotes ru·e able to synthesize polyamines de novo [32, 36] and have the ability to transp01t polyamines [85-88]. Studies in L. donovani and L. mexicana promastigotes found multiple polyamine transp01t systems, which suggests that different transp01ters exist for putrescine and spe1midine uptake [85, 88]. The gene of the first eukruyotic polyamine transp01ter was cloned fi:om L. major (LmPOTl) [87]. In order to ftmctionally chru·acteiize the putative transp01ter, the gene was expressed in Xenopus laevis oocytes and ftuthe1more transfected into the related pru·asite T. brucei, which has poor endogenous putrescine transp01t.
The LmPOTl transp01t er has high affinity for both putrescine and spermidine. However, while LmPOTl is expressed in L. major promastigotes, the transp01ter could not be detected in amastigotes. A compru·ison between L. me.·dcana promastigotes and amastigotes also suggest that dispru·ate transport systems exist in the two different developmental stages of the pru·asite [85].
Studies moreover revealed that at least putrescine uptake is regulated. Inhibition of ODC by DFMO resulted in increased putrescine uptake in promastigotes and conversely, elevated concentration of putrescine in the media resulted in decreased putrescine uptake [86, 88] . It is well established that intracellulru·levels of polyamines ru·e intiicately regulated in mammalian cells and it apperu·s that the same may be tiue for Leishmania parasites. The obse1vation that polyamine enzyme ove1producer strains only modestly increase intracellular polyamine levels despite profoundly elevated enzyme levels also argues that polyamine pools ru·e conu·olled in the parasite [ 46].
While polyamine uptake is not c1itical for Leishmania, the imp01t of the amino acid ru·ginine is essential. Arginine is vital for both polyamine and protein synthesis. An arginine u·anspOit er, AAP3 , has been identified and characte1ized in L. donovani and L. amazonenesis [113, 116]. In promastigotes, intracellulru· arginine pools regulate the ammmt of Leishmania AAP3 ru·ginine u·anspOiter and thus ru·ginine uptake [113, 116]. It is likely that arginine uptake is also regulated in amastigotes and that both pru·asite and host cell compete for intracellulru· ru·ginine [37].
42 Sigrid C. Robe1ts
Interaction of Host and Parasite Polyamine Pathways in Leishmania infections
Infectivity studies with gene deletion mutants can be used to address whether host polyamine metabolism influences parasite polyamine homoeostasis. Ld!lode parasites exhibit dramatically reduced infectivity phenotypes in macrophages and mice [36]. Since amastigotes are able to transpo1t putrescine, the logical explanation is that insufficient ammmts of host putrescine are available.
Such a conjecture is not unreasonable because host polyamine metabolism is intricately regulated and putrescine levels are maintained at constant levels. Thus, it appears that L. donovani parasites do not benefit from host putrescine pools but depend on endogenous putrescine biosynthesis.The Ld!lspd parasites also show a severely reduced infectivity phenotype but to a lesser degree than Ld!lode [38].
It should be noted, that spe1midine is the essential and sufficient polyamine in promastigotes [32]. Thus, the obse1vation that intracellular Ld!lode amastigotes are more incapacitated than Ld!lspd amastigotes is somewhat smprising. Possible explanations include that less putrescine than spe1midine is present in the phagolysosome or that L. donovani amastigotes have unequal uptake capabilities for the two polyamines. In contrast, Leishmania deficient in ARG exhibit only a moderate loss in infectivity compared to the severely reduced infectivity of Ld!lode and Ld!lspd parasites. All three ARG deficient strains that were generated, L. mexieana, L. major, and L.amazonensis !larg, were still able to establish infections [37, 90, 91, 112].
The differences in infectivity phenotypes can be rationalized by the assumption that omithine is present in higher quantities than putrescine or spe1midine in the phagolysosome or that omithine uptake in amastigotes is much more robust than uptake of the polyamines. One should recall thought that !larg promastigotes require much higher concentration of exogenous omithine than putrescine to allow optimal growth [89, 91], an obse1vation that contradicts the hypothesis that amastigotes would be more successful in scavenging omithine than putrescine.
Fmthe1more, it needs to be emphasized that the !lode and !lspd parasites have been generated in L. donovani, a visceralizing strain, whereas the !larg deletions were introduced into Leishmania species that cause a cutaneous disease. It is feasible that the omithine and polyamine milieu of macrophages of the skin is different from that of macrophages residing in liver and spleen.
The generation and direct comparison of !larg, !lode, !lspd mutants within the same species is necessary to solve this conundmm. Nevertheless, a model of host and pru·asite polyamine metabolism and salvage has been constlucted that can be used as a basis for fmther investigations (Figure 5).
This model speculates that high amounts of omithine are available for pru·asite uptake but only limited ammmts of pun·escine and spe1midine ru·e present in the phagolysosome. Although the augmented host ARG activity in Leishmania infections has always been speculated to produce increased amounts of polyamines for pru·asite salvage, omithine may actually be more impo1tant than polyamines for parasite scavenging and enhanced proliferation.
Genetic Manipulation of Leishmania Parasites ...
Comparison of Host-Parasite Interactions Among Trypanosomatid Parasites
43
The polyamine pathway has been investigated in several other protozoan parasites, including the tlypanosomatids T. bmcei and T. cniZi. Similar genetic manipulations have been petfonned to elucidate the metabolic pathway in T. brucei. In addition to classical knockout studies, the technique of RNA interference (RNAi) has been utilized [ 117-120]. RNAi has the advantage of allowing inducible knockdown of target gene expression and thus facilitates the study of essential genes. However, a drawback of RNAi is that sometimes the knockdown is achieved at only prutiallevels and residual enzyme activity may be present. It is of interest to note that most Leishmania species have lost the protein repettoire necessruy for the RNA silencing machinety.
Investigations in T. brucei and T. cruzi have discovered a unique regulat01y protein -prozyme - that is required for ADOMETDC activation [118, 121-123]. This regulat01y mechanism is distinct from that of mammalian cells. Prozyme has not been studied in Leishmania, but the gene encoding this protein is present in the genome database of several Leishmania species and it is thus likely that the Leishmania ADOMETDC also requires heterodimetization with prozyme for full activation.
arginine
8 1 ornithine IJQ
9 1 putrescine
8 1 spermidine ..
macrophage cytosol
ornithine
~1 ·- putrescine
9 1
Leishmania amastfgote
The polyamine biosynthetic enzymes of Leishmania amastigotes and the host polyamine biosynthetic enzymes are displayed. Arginine uptake by amastigotes is essential and indicated with a solid black an·ow. Salvage of omithine is postulated to be efficient and indicated with a solid grey atrow. Uptake of putrescine and spennidine is likely to be minimal and thus indicated with hashed atTOWS.
Figw-e 5. A model of host and parasite polyatnine metabolism atld pat·asite scavenging.
44 Sigrid C. Robe1ts
T brucei parasites contain a polyamine biosynthetic pathway similar to Leishmania consisting of ARG, ODC, SPD and ADOMETDC [1 17-120, 124-126]. However, in contrast to Leishmania, T. brucei parasites are extracellular pathogens that are present in the host's blood stream. Polyamine levels in blood are low and thus T brucei is dependent on endogenous polyamine biosynthesis [118]. The lack of salvageable polyamines offers an explanation for the efficacy of the ODC inhibitor DFMO in the treatment of Aftican sleeping sickness.
The polyamine pathway in T cntzi is more austere than that of the other trypanosomatids; they are not able to synthesize putrescine fi.·om amino acid precmsors [127-130]. Only two other eukruyotes ru·e known to be incapable of de novo polyamine biosynthesis: Toxoplasma gondii and Cryptosporidium parvum [ 131, 132].
T cntzi parasites lack both ARG and ODC and although an ADC activity was initially repo1ted [133, 134], it is now generally accepted that the altemative ADC/AGM pathway does not exist in T cntzi. It has been speculated that the trypanosomatids have lost their ODC gene followed by ho1izontal gene tr·ansfer of the ODC sequence fi.·om a ve1t ebrate host into T brucei [135]. Suppo1t for this hypothesis comes fi.·om phylogenetic analyses, which show the T brucei ODC clustered closer to the ODC genes fi.·om ve1t ebrates than protozoans. Because of the lack of de novo putr·escine synthesis, T cntzi pru·asites ru·e absolutely dependent on putrescine scavenge fi.·om the host. Like Leishmania, T cntzi is an intracellulru· pru·asite taken up by phagocytosis. However, T cruzi pru·asites escape the phagosome to reside in the host cell cytosol. Thus, the pathogen has access to presumably more abtmdant polyamine pools compared to Leishmania pru·asites, which live in phagolysosomes.
In summruy , the interactions between host and pru·asite that are necessruy to maintain polyrunine homeostasis in the pathogen are strikingly different among the three trypanosomatid parasites (Figme 6).
Leishmania parasites reside inside the phagolysosome and although they apperu· to rely on endogenous polyamine biosynthesis (L. donovani) they ru·e capable of at least limited omithine salvage (L. me;-cicana, L. major, L. amazonensis). T cruzi pru·asites inhabit the cytosol and absolutely depend on putr·escine salvage because they ru·e not able to synthesize polyrunines de novo. Finally, T brucei parasites ru·e extr·acellulru· pathogens that live in the bloodstr·eam - a polyamine poor milieu -and rely on endogenous polyamine biosynthesis.
Conclusion
Genetic techniques, foremost the ability to generate gene deletion mutants, have facilitated the elucidation of the polyamine biosynthetic pathway in Leishmania pru·asites. Fmthe1more, the gene deletion mutants ru·e invaluable tools to investigate the relevance of polyrunine biosynthesis for parasite infectivity and the interplay between host and pru·asite polyrunine pathways.
In addition, the ability to generate overproducer strains can be utilized for the development of cell-based screening assays that test the mode of action of inhibito1y compounds. In this manner, the process of genetic manipulation has been applied to validate the polyamine pathway in Leishmania pru·asites as a potential therapeutic target.
Genetic Manipulation of Leishmania Parasites ... 45
A
T. cruzi amastigotes reside free in the host
cell cytosol
B c Leishmania amastigotes
proliferate in phagolysosomcs inside
the host cell
T. brucei are extracellular paras ites tbat live in the
bloodstream
AT. cruzi amastigotes evade the phagosome to reside free in the cytosol and thus have access to the host polyamines putrescine ( + ) and spermidine (• ). Because they are incapable of de novo putrescine synthesis they depend on polyamine salvage. BLeishmania anutstigotes proliferate in phagolysosomal compartments. They are able to synthesize polyamines and although they can
salvage ornithine (* ), polyamine uptake appears to be limited. cT. brucei parasites are extracellular pathogens and rely on endogenous polyamine synthesis because polyamine levels in blood are minute.
Figme 6. Comparison of host-parasite interactions among trypanosomatid parasites.
Substantial differences between the polyamine pathways of host and parasite have been revealed. Leishmania pathogens lack a back-conversion pathway, do not express spermine synthase, and the polyamine biosynthetic enzymes are more stable than their human counte1pruts.
An altemative pathway of putrescine production via ADC and AGM is not present in Leishmania, and the putative AGM that is present in the Leishmania gen01nic databases does not function as such. Hence, the Leishmania polyaniine pathway consists of only four enzymes: ARG, ODC, SPD and ADOMETDC. The hypusination pathway that utilizes spemridine for the modification and activation of eiFSA is present in Leishmania and both enzymes, DHS and DOHH, have recently been described. Sequence comparisons between the host and pru·asite enzymes reveal that similru·ities ru·e moderate, and especially low for DHS and ADOMETDC, which may offer tru·gets for selective inhibitor development. Infectivity studies with L. donovani gene deletion mutants revealed that the Ld!:J.odc parasites show profmmdly reduced infectivity phenotypes compru·ed to wild type pru·asites and validated ODC as a therapeutic target. Similru·ly, Ld!:J.spd pru·asites exhibited severely decreased infectivity, although not as severely impaired as obse1ved in the LdtJ.odc cell line. These
46 Sigrid C. Robe1ts
obse1vations challenged the conventional belief that polyamines are abundant in the phagolysosomal compartment.
Conversely, !:,arg strains that were generated in L. mexicana, L. major and L.
amazonenesis show only moderate reduction in infectivity. These obse1vations suggest that omithine levels are much higher than putrescine ar1d spe1midine pools in the phagolysosome. Studies suggesting that an increased host ARG activity is cmcial for the exacerbation of leishmaniasis and that host polyamine synthesis is tightly regulated suppo1t tltis conjecn1re. However, it should be noted that L. donovani is a visceralizing strain whereas L. mexicana, L. major ar1d L. amazonenesiscause cutaneous leishmaniasis. Thus, the discrepancies in infectivity phenotypes may also be due to differences in the Leishmania species and/or host cell type (macrophages of skin versus liver and spleen).
In conclusion, studies with genetically manipulated Leishmania strains revealed that the enzymes of the polyamine biosynthetic pathway, especially ODC and SPD, are potential therapeutic targets. FU1the1more, the dual inhibition of host and par·asite polyantine synthesis, part icular·ly targeting the host ARG, may have a synergistic effect and is w01thwhile to explore.
Acknowledgment
The author 's resear·ch that is desc1ibed in this article was suppo1ted in part by Grant AI041622 fi:om the National Institute of Allergy and Infectious Disease.
References
[1] Kedzierski, L. Leishmaniasis. Hum. Vaccin. 2011 ; 7:1204-14. [2] Kobets, T. , Grekov, 1., Lipoldova, M. Leishmaniasis: prevention, par·asite detection and
treatment. Cu.rr. Med. Chern. 2012; 19:1443-74. [3] Ganguly, S., Das, N. K., Bar·bhuiya, J. N., Chatte1jee, M. Post-kala-azar de1mal
leishmaniasis--an ove1view. Int. J. Dermatol. 2010; 49:921-31. [ 4] Singh, S., Sharma, U., Mishra, J. Post-kala-azar de1mal leishmaniasis: recent
developments. Int. J. Dermatol. 2011; 50:1099-108. [5] Kaye, P., Scott, P. Leishmaniasis: complexity at the host-pathogen interface. Nat. Rev.
Microbial. 2011 ; 9:604-15. [6] Kedzierski, L. , Sakthianandeswaren, A., CUit is, J. M., Andrews, P. C., Junk, P. C.,
Kedzierska, K. Leishmaniasis: CUITent treatment and prospects for new dmgs and vaccines. Curr. Med. Chern. 2009; 16:599-614.
[7] Muller, R. Advances in Parasitology. San Diego, CA: Academic Press, 2007. [8] Sinde1mann, H., Engel, J. Development of miltefosine as an oral treatment for
leishmaniasis. Transactions of the Royal Society of Tropical Medicine and Hygiene 2006; 100 Suppl. 1:S17-20.
[9] Bacchi, C. J MP. Parasitic protozoa and polyamines. In: MCann, P. P. P. A. , Sjoerdsma, A., ed. Inhibition of Polyamine Metabolism: Biological Significance and Basis for New Therapies. Orlando, FL: Academic Press, 1987:3 17-44.
Genetic Manipulation of Leishmania Parasites .. . 47
[10] Buni, C., Bnm, R. Eflornithine for the treatment of human African trypanosorniasis. Parasitol. Res. 2003; 90 Supp. 1:S49-52.
[11] Docampo, R. , Moreno, S. N. Cunent chemotherapy of human African trypanosomiasis. Parasitol. Res. 2003; 90 Supp. 1:S10-3.
[12] Schechter, P. J. B. J., Sjoerdsma, A. Clinical aspects of inhibition of omithine decarboxylase with emphasis on therapeutic tiials of eflomithine (DFMO) in cancer and protozoan diseases. h1: McCann, P. P. P. A., Sjoerdsma, A., ed. Inhibition of Polyamine Metabolism: Biological Significance and and Basis for New Therapies, Orlando, FL: Academic Press, 1987:345-64.
[13] Park, M. H. The post-translational synthesis of a polyantine-derived anlino acid, hypusine, in the eukaryotic translation initiation factor SA (eiFSA). J. Biochem. 2006; 139:161-9.
[14] Park, M. H., Nishimma, K. , Zanelli, C. F., Valentini, S. R. Functional significance of eiFSA and its hypusine modification in eukaryotes. Amino Acids 2010; 38:491-500.
[15] Nishimma, K., Lee, S. B., Park, J. H., Park, M. H. Essential role of eiFSA-1 and deoxyhypusine synthase in mouse embryonic development. Amino acids 2012; 42: 703-10.
[16] Park, M. H., Joe, Y. A., Kang, K. R. Deoxyhypusine synthase activity is essential for cell viability in the yeast Saccharomyces cerevisiae. The Journal of biological chemistry 1998; 273: 1677-83.
[17] Aliyanayagam, M. R. , Oza, S. L., Guther, M. L., Fairlamb, A. H. Phenotypic analysis of tlypanothione synthetase knockdown in the African trypanosome. Biochem. J. 2005; 391 :425-32.
[18] Dumas, C. , Ouellette, M., Tovar, J., Cunningham, M. L. , Fairlamb, A. H., Tamar, S., et al. Dismption of the tlypanothione reductase gene of Leishmania decreases its ability to smvive oxidative str·ess in macrophages. EMBO J. 1997; 16:2590-8.
[19] Faii·lamb, A. H., Blackbmn, P., Uhich, P. , Chait, B. T., Cerami, A. Tiypanothione: a novel bis(glutathionyl)spennidine cofactor for glutathione reductase in tlypanosomatids. Science 1985; 227: 1485-7.
[20] Krieger, S., Schwarz, W., AI·iyanayagam, M. R., Faii·lamb, A. H., KI·auth-Siegel, R. L., Clayton, C. Trypanosomes lacking tlypanothione reductase are aviiulent and show increased sensitivity to oxidative str·ess. Mol. Microbiol. 2000; 35:542-52.
[21] Tovar, J., Wilkinson, S., Mottram, J. C., Faii·lamb, A. H. Evidence that tlypanothione reductase is an essential enzyn1e in Leishmania by targeted replacement of the tlyA gene locus. Mol. Microbiol. 1998; 29:653-60.
[22] KI·auth-Siegel, L. R., Comini, M. A., Schlecker, T. The trypanothione system. Subcell. Biochem. 2007; 44:231-51.
[23] KI·auth-Siegel, R. L., Comini, M. A. Redox contr·ol in tlypanosomatids, parasitic protozoa with tlypanothione-based thiol metabolism. Biochim. Biophys. Acta 2008; 1780:1236-48.
[24] KI·auth-Siegel, R. L. , hlhoff, 0 . Parasite-specific tlypanothione reductase as a dmg target molecule. Parasitol. Res. 2003; 90 Suppl. 2:877-85.
[25] Muller, S. , Liebau, E., Walter, R. D. , KI·auth-Siegel, R. L. Thiol-based redox metabolism of protozoan parasites. Trends Parasitol. 2003; 19:320-8.
[26] Coleman, C. S., Hu, G., Pegg, A. E. Putrescine biosynthesis in mammalian tissues. Biochem. J. 2004; 379:849-55.
48 Sigrid C. Robe1ts
[27] Monis, S. M., Jr. Recent advances in arginine metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2004; 7:45-51.
[28] Sastre, M., Galea, E. , Feinstein, D., Reis, D. J., Regunathan, S. Metabolism of agmatine in macrophages: modulation by lipopolysaccharide and inhibito1y cytokines. Biochem. J. 1998; 330 ( Pt 3):1405-9.
[29] Chawla, B., Jhingran, A., Singh, S., Tyagi, N., Park, M. H., Srinivasan, N., et al. Identification and characte1ization of a novel deoxyhypusine synthase in Leishmania donovani. J. Biol. Chem. 2010; 285:453-63.
[30] Mukhe1jee, A. , Roy, G., Guimond, C., Ouellette, M. The gamma-glutamylcysteine synthetase gene of Leishmania is essential and involved in response to oxidants. Molecular microbiology 2009; 74:914-27.
[31] Denise, H., Poot, J. , Jimenez, M., Ambit, A., Henmann, D. C., Ve1meulen, A. N., et al. Studies on the CPA cysteine peptidase in the Leishmania infantum genome strain JPCM5. BMC Mol. Biol. 2006; 7:42.
[32] Jiang, Y. , Robe1ts, S. C., Jardim, A. , Ca1ter, N. S., Shih, S., Ariyanayagam, M., et al. 01nithine decarboxylase gene deletion mutants of Leishmania donovani. J. Bioi. Chem. 1999; 274:3781-8.
[33] Robe1ts, S. C., Jiang, Y., Jardim, A., Carter, N. S., Heby, 0 ., Ullman, B. Genetic analysis of spe1midine synthase fi:om Leishmania donovani. Mol. Biochem. Parasitol. 2001; 115:217-26.
[34] Robe1ts, S. C., Kline, C., Liu, W. , Ullman, B. Generating knock-in parasites: integration of an 01nithine decru·boxylase transgene into its chromosomal locus in Leishmania donovani. &<perimental parasitology 2011 ; 128: 166-9.
[35] Robe1ts, S. C., Scott, J., Gasteier, J. E., Jiang, Y. , Brooks, B., Jru·dim, A., et al. Sadenosylmethionine decarboxylase from Leishmania donovani. Molecular, genetic, and biochelnical chru·acte1ization of null mutants and overproducers. J. Bioi. Chem. 2002; 277:5902-9.
[36] Boitz, J. M., Yates, P. A., Kline, C., Gaur, U. , Wilson, M. E., Ullman, B. , et al. Leishmania donovani 01nithine decru·boxylase is indispensable for pru·asite smvival in the mammalian host. Infect. Immun. 2009; 77:756-63.
[37] Gaur, U., Roberts, S. C., Dalvi, R. P., Conaliza, I., Ullman, B., Wilson, M. E. An effect of pru·asite-encoded ru·ginase on the outcome of murine cutaneous leishmaniasis. J. Immunol. 2007; 179:8446-53.
[38] Gilroy, C. , Olenyik, T., Robe1ts, S. C., Ullman, B. Spe1midine synthase is required for vimlence ofLeishmania donovani. Infection and immunity 2011; 79:2764-9.
[39] Buckner, F. S., Wilson, A. J. Cololimetric assay for screening compounds against Leishmania amastigotes grown in macrophages. The American journal of tropical medicine and hygiene 2005; 72:600-5.
[40] Ha, D. S., Schwru·z, J. K. , Turco, S. J. , Beverley, S. M. Use of the green fluorescent protein as a mru·ker in transfected Leishmania. Molecular and biochemical parasitology 1996; 77:57-64.
[41] Roy, G., Dumas, C., Sereno, D., Wu, Y. , Singh, A. K., Tremblay, M. J., et al. Episomal and stable expression of the luciferase rep01t er gene for quantifying Leishmania spp. infections in macrophages and in animal models. Molecular and biochemical parasitology 2000; 110: 195-206.
Genetic Manipulation of Leishmania Parasites .. . 49
[42] Basile, G., Peticca, M. Recombinant protein expression in Leishmania tarentolae. Mol. Biotechnol. 2009; 43:273-8.
[43] Kushnir, S. , Cirstea, I. C., Basiliya, L., Lupilova, N., Breitling, R., Alexandrov, K. Alt ificial linear episome-based protein expression system for protozoon Leishmania tarentolae. Molecular and biochemical parasitology 2011; 176:69-79.
[44] Mureev, S., Kushnir, S., Kolesnikov, A. A., Breitling, R., Alexandrov, K. Constmction and analysis of Leishmania tarentolae transgenic strains free of selection markers. Molecular and biochemical parasitology 2007; 155:71-83.
[45] Beattie, L. , Evans, K. J., Kaye, P. M., Stnith, D. F. Transgenic Leishmania and t11e immtme response to infection. Parasite immunology 2008; 30:255-66.
[46] Robetts, S. C. , Jiang, Y., Gasteier, J., Ftydman, B., Matton, L. J., Heby, 0 ., et al. Leishmania donovani polyamine biosynthetic enzyme ovetproducers as tools to investigate the mode of action of cytotoxic polyamine analogs. Antimicrob. Agents Chemother. 2007; 51:438-45.
[47] Zhang, W. W. , Chan, K. F., Song, Z. , Matlashewski, G. Expression of a Leishmaniadonovani nucleotide sugar transpmt er in Leishmaniamajor enhances smvival in visceral organs. R-rperimental parasitology 2011; 129:337-45.
[48] Zhang, W. W., Mendez, S., Ghosh, A., Myler, P., Ivens, A., Clos, J. , et al. Comparison of the A2 gene locus in Leishmania donovani and Leishmania major and its control over cutaneous infection. The Journal of biological chemistry 2003; 278:35508-15.
[49] Pegg, A. E. , Michael, A. J. Spemrine synthase. Cell. Mol. Life Sci. 2010; 67: 113-21. [50] Hanson, S., Adelman, J., Ullman, B. Amplification and molecular cloning of the
omithine decarboxylase gene of Leishmania donovani. J. Bioi. Chern. 1992; 267: 2350-9.
[51] Ghoda, L. , Phillips, M. A., Bass, K. E. , Wang, C. C., Coffino, P. T!ypanosome omithine decarboxylase is stable because it lacks sequences found in the carboxyl temrinus of the mouse enzyme which target the latter for intracellular degradation. J. Bioi. Chern. 1990; 265: 11823-6.
[52] Chawla, B., Kumar, R. R., Tyagi, N., Subramanian, G., Siinivasan, N., Park, M. H., et al. A unique modification of the eukruyotic initiation factor SA shows the presence of the complete hypusine pathway in Leishmania donovani. PLoS One 2012; 7:e33138.
[53] Kaiser, A., Gottwald, A., Wiersch, C., Lindenthal, B., Maier, W., Seitz, H. M. Effect of diugs inhibiting spemtidine biosynthesis and metabolism on the in vitro development of Plasmodium falcipamm. Parasitol. Res. 2001; 87:963-72.
[54] Njugtma, J. T. , Nassru·, M., Hoerauf, A., Kaiser, A. E. Cloning, expression and functional activity of deoxyhypusine synthase from Plasmodium vivax. BMC Microbiol. 2006; 6:91.
[55] Schwentke, A., Krepstakies, M., Muller, A. K., Hammerschmidt, C. , Motaal, B., Bemhru·d, T., et al. In vitro and in vivo silencing of plasmodial dbs and elF-SA genes in a putative, non-canonical RNAi-related pathway. BMC microbiology 20 12; 12: 107.
[56] Specht, S., Sru·ite, S. R. , Hauber, 1. , Hauber, J. , Gm·big, U. F., Meier, C. , et al. The gttanylliydi·azone CNI-1493: an inhibitor with dual activity against malruia-inhibition of host cell pro-inflanilllatmy cytokine release and pru·asitic deoxyhypusine synthase. Parasitology research 2008; 102:1177-84.
50 Sigrid C. Robe1ts
[57] Kaur, K., Emmett, K., McCallil, P. P., Sjoerdsma, A., Ulhnan, B. Effects ofDL-alphadifluoromethylomithine on Leishmania donovani promastigotes. J. Protozool. 1986; 33:518-21.
[58] Mukhopadhyay, R., Kapoor, P., Madhubala, R. Characte1ization of alphadifluoromethylomithine resistant Leishmania donovani and its susceptibility to other inhibitors of the polyamine biosynthetic pathway. Pharmacol. Res. 1996; 34:43-6.
[59] Reguera, R. M., Fouce, R. B., Cublia, J. C. , Bujidos, M. L., Ordonez, D. Fluorinated analogues of L-omithine are powerftd inhibitors of omithine decarboxylase and cell growth of Leishmania infantum promastigotes. Life Sci. 1995; 56:223-30.
[60] Sanchez, C. P. , Mucci, J. , Gonzalez, N. S., Ochoa, A., Zakin, M. M., Algranati, I. D. Alpha-difluoromethylomithine-resistant cell lines obtained after one-step selection of Leishmania mexicana promastigote cultures. Biochem. J. 1997; 324 ( Pt 3):847-53.
[61] Gradoni, L. , Io1io, M. A., Gralniccia, M., Orsini, S. In vivo effect of eflomithine (DFMO) and some related compounds on Leishmania infantum prelilninruy communication. Farmaco. 1989; 44: 1157-66.
[62] J. S. Keithly A. F. Inhibition of Leishmania species by alpha-difluoromethylomithine. In: Hrut , D., ed. Leishmaniasis: The current status and new strategies for control. New York, US: Plenum Press, 1987:749-56.
[63] Mukhopadhyay, R., Madhubala, R. Effect of a bis (benzyl) polyamine analogue, and DL-alpha-difluoromethylomithine on pru·asite suppression and cellulru· polyamine levels in golden hamster during Leishmania donovani infection. Pharmacol. Res. 1993; 28:359-65.
[64] Olenyik, T., Gih·oy, C., Ulhnan, B. Oral putrescine restores vilulence of omithine decru·boxylase-deficient Leishmania donovani in Inice. Molecular and biochemical parasitology 2011; 176:109-11.
[65] Singh, S., Jhingran, A., Sha1ma, A., Simonian, A. R., Soininen, P., Vepsalainen, J. , et al. Novel agmatine analogue, gamma-guanidinooxypropylamine (GAPA) efficiently inhibits proliferation of Leishmania donovani by depletion of intracellular polyamine levels. Biochem. Biophys. Res. Commun. 2008; 375:168-72.
[66] Singh, S., Mukhe1jee, A., Khomutov, A. R., Persson, L., Heby, 0., Chatte1jee, M., et al. Antileishmanial effect of 3-aminooxy-1-aminopropane is due to polyamine depletion. Antimicrob. Agents Chemother. 2007; 51 :528-34.
[67] Vallllier-Santos, M. A. , Menezes, D., Oliveira, M. F., de Mello, F. G. The putrescine analogue 1,4-diamino-2-butanone affects polyamine synthesis, transport, ultrastmcture and intracellular smv ival in Leishmania amazonensis. Microbiology 2008; 154:3104-11.
[68] Mukhopadhyay, R., Madhubala, R. Effects of bis(benzyl)polyamine analogs on Leishmania donovani promastigotes. R-rp. Parasitol. 1995; 81:39-46.
[69] Baumann, R. J., McCallil, P. P., Bitonti, A. J. Suppression of Leishmania donovani by oral administration of a bis (benzyl) polyrunine analog. Antimicrob. Agents Chemother. 1991; 35:1403-7.
[70] Dufe, V. T. , Ingner, D., Heby, 0 ., Khomutov, A. R., Persson, L., Al-Kru·adaghi, S. A stmctmal insight into the inhibition of human and Leishmania donovani omithine decru·boxylases by 1-alnino-oxy-3-aminopropane. Biochem. J. 2007; 405:261-8.
Genetic Manipulation of Leishmania Parasites .. . 51
[71] Danzin, C., Marchal, P., Casara, P. hTeversible inhibition of rat S-adenosyhnethionine decarboxylase by 5'-([ (Z)-4-arnino-2-butenyl]methylarnino )-5'-deoxyadenosine. Biochemical pharmacology 1990; 40:1499-503.
[72] Goda, H., Watanabe, T. , Takeda, N., Kobayashi, M., Wada, M., Hosoda, H., et al. Mammalian spennidine synthase--identification of cysteine residues and investigation of the putrescine binding site. Biol. Pharm. Bull. 2004; 27:1327-32.
[73] Grishin, N. V., Oste1man, A. L,. Brooks, H. B., Phillips, M. A., Goldsmith, E. J. X-ray stmcture of omithine decarboxylase fi-om Trypanosoma bmcei: the native stmcture and the stmcture in complex with alpha-difluoromethylomithine. Biochemistry 1999; 38:15174-84.
[74] Basselin, M., Badet-Denisot, M. A., Lawrence, F., Robe1t-Gero, M. Effects of pentamidine on polyamine level and biosynthesis in wild-type, pentamidine-treated, and pentamidine-resistant Leishmania. Experimental parasitology 1997; 85:274-82.
[75] Basselin, M., Badet-Denisot, M.A., Robe1t-Gero, M. Modification of kinetoplast DNA rninicircle composition in pentamidine-resistant Leishmania. Acta tropica 1998; 70:43-61.
[76] Basselin, M., Denise, H., Coombs, G. H., Banett, M. P. Resistance to pentamidine in Leishmania mexicana involves exclusion of the dmg from the mitochond1ion. Antimicrobial agents and chemotherapy 2002; 46:3731-8.
[77] Bitonti, A. J., Dumont, J. A. , McCann, P. P. Characte1ization of T1ypanosoma bmcei bmcei S-adenosyl-L-methionine decarboxylase and its inhibition by Berenil, pentamidine and methylglyoxal bis(guanylhydrazone). The Biochemical journal 1986; 237:685-9.
[78] Leon, W. Selective inhibition of k-DNA synthesis of T1ypanosoma cmzi and Leishmania tarentolae by berenil and ethidium bromide [proceedings]. An. Acad. Bras. Cienc. 1978; 50:597-8.
[79] Mukhe1jee, A., Padmanabhan, P. K., Sahani, M. H., Banett, M. P., Madhubala, R.
Roles for mitochondria in pentamidine susceptibility and resistance in Leishmania donovani. Molecular and biochemical parasitology 2006; 145: 1-10.
[80] Singh, R., Siddiqui, K. A., Valenzuela, M. S., Majumder, H. K. Kinetoplast DNA rninicircle binding proteins in a Leishmania Spp: interference of protein DNA interaction by berenil. Indian J. Biochem. Biophys . 1995; 32:437-41.
[81] Cohen, S. A guide to polyamines. Oxford, UK: Oxford University Press, 1998. [82] McConville, M. J., de Souza, D., Saunders, E., Likic, V. A., Naderer, T. Living in a
phagolysosome; metabolism of Leishmania amastigotes. Trends in parasitology 2007; 23:368-75.
[83] Naderer, T. , McConville, M. J. The Leishmania-macrophage interaction: a metabolic perspective. Cell Microbiol. 2008; 10:301-8.
[84] Satmders, E. C., D.P. Des, Naderer, T. , Semee, M. F., Ralton, J. E., Doyle, M. A. , et al. Central carbon metabolism of Leishmania parasites. Parasitology 2010; 137:1303-13.
[85] Basselin, M., Coombs, G. H. , Banett, M. P. Putrescine and spe1midine transpo1t in Leishmania. Mol. Biochem. Parasitol. 2000; 109:37-46.
[86] Gonzalez, N. S., Algranati, I. D. Regulation of putrescine uptake in Leishmania mexicana promastigotes. Cell. Mol. Biol. (Noisy-le-grand) 1994; 40:907-14.
[87] Hasne, M. P., Ulhnan, B. Identification and characterization of a polyamine pe1mease from the protozoan parasite Leishmania major. J. Bioi. Chern. 2005; 280: 15188-94.
52 Sigrid C. Robe1ts
[88] Kandpal, M., Tekwani, B. L. Polyamine transport systems of Leishmania donovani promastigotes. Life Sci. 1997; 60:1793-801.
[89] Roberts, S. C. , Ta11eer, M. J., Polinsky, M. R., Gibson, K. M., Heby, 0., Ullman, B. Arginase plays a pivotal role in polyamine precursor metabolism in Leishmania. Charactelization of gene deletion mutants. J. Biol. Chem. 2004; 279:23668-78.
[90] DaSilva, M. F., Zampie1i , R. A., Muxel, S.M., Beverley, S.M., Floeter-Winter, L. M. Leishmania amazonensis arginase compartmentalization in the glycosome is impo1t ant for parasite infectivity. PLoS One 2012; 7:e34022.
[91] Reguera, R. M. , Balana-Fouce, R., Showalter, M. , Hickerson, S., Beverley, S. M. Leishmania major lacking ar·ginase (ARG) are auxotrophic for polyamines but retain infectivity to susceptible BALB/c mice. Mol. Biochem. Parasitol. 2009; 165:48-56.
[92] Michels, P. A., Bringaud, F., He1man, M., Hannae1t, V. Metabolic ftmctions of glycosomes in tlypanosomatids. Biochimica et biophysica acta 2006; 1763:1463-77.
[93] Par·sons, M. Glycosomes: parasites and the divergence of peroxisomal pmpose. Molecular microbiology 2004; 53:717-24.
[94] Monis, S. M., Jr. Recent advances in arginine metabolism: roles and regulation of the ar·ginases. Br. J. Phannacol. 2009; 157:922-30.
[95] Mauel, J., Ransijn, A. , Buchmuller-Rouiller, Y. Killing of Leishmania parasites in activated mmine macrophages is based on an L-ar·ginine-dependent process that produces nin·ogen derivatives. Journal of leukocyte biology 1991; 49:73-82.
[96] Wanasen, N., Soong, L. L-ar·ginine metabolism and its impact on host immtmity against Leishmania infection. Immunol. Res. 2008; 41: 15-25.
[97] Iniesta, V., Carcelen, J., Molano, 1. , Peixoto, P. M., Redondo, E., PatTa, P., et al. Arginase I induction dming Leishmania major infection mediates the development of disease. Infect. Immun. 2005; 73:6085-90.
[98] Iniesta, V., Gomez-Nieto, L. C. , Conaliza, I. The inhibition of arginase by N(omega)hydroxy-1-ar·ginine conn·ols the growth of Leishmania inside macrophages. J. R-rp. Med. 2001; 193:777-84.
[99] Iniesta, V., Gomez-Nieto, L. C., Molano, I., Mohedano, A. , Car·celen, J., Miron, C., et al. Arginase I induction in macrophages, n·iggered by Th2-type cytokines, suppo1ts the growth of inn·acellular Leishmania par·asites. Parasite immunology 2002; 24:113-8.
[100] Kropf, P., Fuentes, J. M. , Fahnrich, E., Alpa, L. , Herath, S., Weber, V., et al. AI·ginase and polyamine synthesis are key factors in the regulation of experimental leishmaniasis in vivo. FASEB J. 2005; 19:1000-2.
[101] Kropf, P., Herath, S., Weber, V. , Modolell, M., Muller, I. Factors influencing Leishmania major infection in IL-4-deficient BALB/c mice. Parasite Immunol. 2003; 25:439-47.
[102] AI·endse, B., Van Snick, J., Brombacher, F. IL-9 is a susceptibility factor in Leishmania major infection by promoting den·imental Th2/type 2 responses. Journal of immunology 2005; 174:2205-11.
[104] Gallardo-Soler, A. , Gomez-Nieto, C., Campo, M. L., Mar·athe, C., Tontonoz, P., Casnillo, A., et al. AI·ginase I induction by modified lipoproteins in macrophages: a peroxisome proliferator-activated receptor-gamma/delta-mediated effect that links lipid metabolism and immunity. Mol. Endocrinol. 2008; 22:1394-402.
Genetic Manipulation of Leishmania Parasites .. . 53
[105] Goto, H., Gomes, C. M., Corbett, C. E., Monteiro, H. P., Gidhmd, M. Insulin-like growth factor I is a growth-promoting factor for Leishmania promastigotes and amastigotes. Proc. Natl. Acad. Sci. US 1998; 95:13211-6.
[106] Muller, I. , Hailu, A., Choi, B. S., Abebe, T., Fuentes, J. M., Munder, M., et al. Agerelated alteration of arginase activity impacts on sevetity of leishmaniasis. PLoS neglected tropical diseases 2008; 2:e235.
[107] Rogers, M., Kropf, P., Choi, B. S., Dillon, R., Podinovskaia, M., Bates, P., et al. Proteophosophoglycans regurgitated by Leishmania-infected sand flies target the Larginine metabolism of host macrophages to promote parasite survival. PLoS Pathog. 2009; 5:e1000555.
[108] Shweash, M., Adtienne McGachy, H., Schroeder, J., Neamatallah, T., Btyant, C. E., Millington, 0 ., et al. Leishmania mexicana promastigotes inhibit macrophage IL-12 production via TLR-4 dependent COX-2, iNOS and arginase-! expression. Mol. Immunol. 2011; 48:1800-8.
[109] Choi, B.S., Matt inez-Falero, I. C., Corset, C. , Munder, M., Modolell, M., Muller, I., et al. Differential impact of L-arginine deptivation on the activation and effector functions ofT cells and macrophages. J Leukoc. Bioi. 2009; 85:268-77.
[110] Modolell, M., Choi, B. S., Ryan, R. 0 ., Hancock, M., Titus, R. G., Abebe, T., et al. Local suppression of T cell responses by arginase-induced L-arginine depletion in nonhealing leishmaniasis. PLoS Neg/. Trop. Dis. 2009; 3:e480.
[Ill] Munder, M., Choi, B. S., Rogers, M., Kropf, P. L-arginine deprivation impairs Leishmania major-specific T-cell responses. Eur. J Immunol. 2009; 39:2161-72.
[112] Muleme, H. M., Reguera, R. M., Berard, A. , Azinwi, R. , Jia, P., Okwor, I. B., et al. Infection with arginase-deficient Leishmania major reveals a parasite numberdependent and cytokine-independent regulation of host cellular arginase activity and disease pathogenesis. J Immunol . 2009; 183:8068-76.
[113] Darlyuk, I. , Goldman, A., Robetts, S. C., Ullman, B., Rentsch, D., Zilberstein, D. Arginine homeostasis and transp01t in the human pathogen Leishmania donovani. J Bioi. Chem. 2009; 284: 19800-7.
[114] Wanasen, N., MacLeod, C. L. , Ellies, L. G., Soong, L. L-arginine and cationic amino acid transporter 2B regulate growth and smvival of Leishmania amazonensis amastigotes in macrophages. Infection and immunity 2007; 75:2802-10.
[115] Riley, E., Robetts, S. C. , Ullman, B. Inhibition profile of Leishmania mexicana arginase reveals differences with human arginase I. International journal for parasitology 2011; 41 :545-52.
[116] Castilho-Martins, E. A. , Laranjeira da Silva, M. F., dos Santos, M. G., Muxel, S. M., Floeter-Winter, L. M. Axenic Leishmania amazonensis promastigotes sense both the extemal and intemal arginine pool distinctly regulating the two transp01ter-coding genes. PLoS One 2011; 6:e27818.
[117] Taylor, M. C., Kaur, H., Blessington, B., Kelly, J. M., Wilkinson, S. R. Validation of spemlidine synthase as a dt11g target in African tlypanosomes. Biochem. J. 2008; 409:563-9.
[118] Willett , E., Phillips, M. A. Regulation and function of polyanlines in Afiican tlypanosomes. Trends in parasitology 2012; 28:66-72.
[119] Xiao, Y., McCloskey, D. E., Phillips, M. A. RNA interference-mediated silencing of omithine decarboxylase and spemlidine synthase genes in Ttypanosoma bmcei
54 Sigrid C. Robe1ts
provides insight into regulation of polyamine biosynthesis. Eukaryot Ce/12009; 8:747-55.
[120] Li, F., Hua, S. B., Wang, C. C., Gottesdiener, K. M. Trypanosoma brucei brucei: characterization of an ODC null bloodstream f01m mutant and the action of alphadifluoromethylomithine. Exp. Parasitol. 1998; 88:255-7.
[121] Wille1t, E. K., Fitzpatrick, R., Phillips, M. A. Allosteric regulation of an essential tlypanosome polyamine biosynthetic enzyme by a catalytically dead homolog. Proc. Natl. Acad. Sci. US2007; 104:8275-80.
[122] Wille1t, E. K., Phillips, M. A. Regulated expression of an essential allosteric activator of polyamine biosynthesis in Afiican tlypanosomes. PLoS Pathog. 2008; 4:e1000183.
[123] Wille1t, E. K., Phillips, M. A. Cross-species activation of tlypanosome Sadenosylmethionine decarboxylase by the regulato1y subunit prozyme. Mol. Biochem. Parasitol. 2009; 168:1-6.
[124] Bacchi, C. J. , Yarlett, N. Polyamine metabolism as chemotherapeutic target in protozoan parasites. Mini. Rev. Med. Chem. 2002; 2:553-63.
[125] Heby, 0 ., Persson, L. , Rentala, M. Targeting the polyamine biosynthetic enzymes: a promising approach to therapy of Afiican sleeping sickness, Chagas' disease, and leishmaniasis. Amino Acids 2007; 33:359-66.
[126] Heby, 0 ., Robe1ts, S. C., Ullman, B. Polyamine biosynthetic enzymes as drug targets in parasitic protozoa. Biochem. Soc. Trans 2003; 31 :415-9.
[127] Aliyanayagam, M. R., Fairlamb, A. H. Diamine auxott·ophy may be a universal feature of Trypanosoma cruzi epimastigotes. Mol. Biochem. Parasitol. 1997; 84:111-21.
[128] Canillo, C., Cejas, S., Gonzalez, N. S., Algranati, I. D. T1ypanosoma cruzi epimastigotes lack omithine decarboxylase but can express a foreign gene encoding this enzyme. FEES Lett. 1999; 454:192-6.
[129] Canillo, C., Cejas, S., Huber, A., Gonzalez, N. S., Algranati, I. D. Lack of arginine decarboxylase in Trypanosoma cruzi epimastigotes. J. Eu.karyot Microbiol. 2003; 50:312-6.
[130] Sena, M. P., Senn, A. M., Algranati, I. D. Post-tt·anslational processing, metabolic stability and catalytic efficiency of oat arginine decarboxylase expressed in T1ypanosoma cruzi epimastigotes. R-rp. Parasitol. 2009; 122:169-76.
[131] Keithly, J. S., Zhu, G., Upton, S. J., Woods, K. M. , Martinez, M. P., Yarlett, N. Polyamine biosynthesis in C1yptosp01idium pa1vum and its implications for chemotherapy. Mol. Biochem. Parasitol. 1997; 88:35-42.
[132] Seabra, S. H., DaMatta, R. A., de Mello, F. G., de Souza, W. Endogenous polyamine levels in macrophages is sufficient to suppo1t growth of Toxoplasma gondii. J. Parasitol. 2004; 90:455-60.
[133] Hemandez, S., Schwru·cz de Tarlovsky, S. AI·ginine decarboxylase in Trypanosoma cruzi, chru·actedstics and kinetic prope1ties. Cell. Mol. Bioi. (Noisy-le-grand) 1999; 45:383-91.
[134] Schwru·cz de Tarlovsky, M. N. , Hemandez, S. M. , Bedoya, A. M. , Lammel, E. M., Isola, E. L. Polyamines in T1ypanosoma cruzi. Biochem. Mol. Biol. Int. 1993; 30: 547-58.
[135] Steglich, C., Schaeffer, S. W. The omithine decru·boxylase gene of T1ypanosoma brucei: Evidence for h01izontal gene tt·ansfer fi·om a ve1t ebrate source. Infect. Genet. Evol. 2006; 6:205-19.