Genetic Variation at a Yin-Yang 1 Response Site Regulates theTranscription of Cyclin-Dependent Kinase Inhibitor p18INK4C
Transcript in Lupus-Prone Mice
Hari-Hara S. K. Potula and Laurence Morel
We have previously shown that a novel 274 C-to-T mutation in the promoter of the cyclin-dependent kinase inhibitor p18Ink4c
(p18) gene was associated with a reduced p18 expression in B cells from mice carrying the Sle2c1 lupus susceptibility locus. To
determine the function of the 274 C/T single nucleotide polymorphism, we have characterized the proximal promoter of the
mouse p18 gene. Functional analysis of the 59 flanking region by sequential deletions revealed crucial elements between 2300 and
+1, confirming the in silico prediction that the 274 T allele created a novel Yin-Yang 1 (YY-1) binding site adjacent to an existing
one common to both alleles. Moreover, we found that YY-1, E2F1, and Sp-1 can synergistically enhance the activity of the p18
promoter. Mutational inactivation revealed that YY-1 binding regulates the p18 activity in an allele-dependent fashion. EMSAs
with splenic B cell extracts directly demonstrated that YY-1 binds to the p18 promoter with differences between the C and the T
alleles. We also determined in vivo by chromatin immunoprecipitation that the T allele resulted in increased YY-1 and decreased
Nrf-2 binding to the p18 promoter as compared with the C allele in B cells. Thus, YY-1 is a direct regulator of p18 gene expression
in an allele-dependent fashion that is consistent with the lupus-associated T allele, inducing a lower p18 transcriptional activity by
increasing YY-1 binding. These results establish the p18274 C/T mutation as the leading causal variant for the B1a cell expansion
that characterizes the NZB and NZM2410 lupus-prone strains. The Journal of Immunology, 2012, 188: 4992–5002.
The mammalian mitotic cell cycle is divided into four dis-tinct phases: G1, S, G2, and M (1). Progression through thecell cycle is regulated by the activation and subsequent
inactivation of a family of serine/threonine kinases known ascyclin-dependent kinases (CDKs) (2). In the G1 phase, positive andnegative growth signals regulate CDK activity through numerousmechanisms, including the expression of CDK inhibitors (CDKIs).Association of a CDK with a positively regulating cyclin D or Esubunit leads to its activation, which is prevented by the binding ofthe CDKs by CDKIs (2). Based on binding properties, the CDKIsare divided into two families: the inhibitor of kinase 4 (Ink4:p16Ink4a, p15Ink4b, p18Ink4c [p18], and p19Ink4d) and CDK/kinaseinhibitory proteins (p21Cip1, p27Kip1, and p57). p18 acts primarilyon CDK4 and CDK6, and is involved in cell-cycle arrest in a varietyof cells, including B lymphocytes (3). Increased p18 expression hasbeen associated with specific cell stimulation and differentiationconditions, such as IL-6 stimulation and terminal differentiation ofB cells into plasma cells (4). In contrast, p18 expression is down-regulated by agents that stimulate cell proliferation such as genis-tein in breast cancer cells (5) and HTLV infection in T cells (6).
Relatively little is known about the regulation of p18 expression,most of it from studies in human cell lines. The p18 protein is verystable with a very low turnover (7), implying that p18 is mostlyregulated at the transcriptional level. The human p18 promoter doesnot contain a TATA box, and is activated with the ubiquitous E2F1and Sp-1 transcription factors, without any evidence of downstreamregulatory elements (8).The Cdkn2c gene, which encodes for p18, is located within
Sle2c1, an NZM2410/NZB-derived systemic lupus erythematosus(SLE) susceptibility locus that we have associated with an expan-sion of peritoneal cavity B1a cells (9, 10). Previously, we haveidentified a novel 274 C/T single nucleotide polymorphism (SNP)in the NZB allele of the p18 promoter, which was associated witha significantly reduced p18 expression in the splenic B (spB) cellsand peritoneal cavity B1a cells from Sle2c1-carrying mice. Thepresence of the T allele was also associated with a defective G1cell-cycle arrest in spB cells and an increased proliferation ofperitoneal cavity B1a cells, leading to an expanded B1a cell pop-ulation that has been associated with autoimmunity (10). Overall,these studies identifiedCdkn2c as the lead candidate gene for Sle2c1and suggested the 274 C/T as the causal polymorphism. Geneticlinkage and association studies cannot distinguish a direct effect ofthis polymorphism on gene function from differences mediated byother neighboring polymorphisms that may be in linkage disequi-librium. To better understand the mechanism that modulates p18expression, we performed studies to determine the functional sig-nificance of the274 C/Texchange in the p18 promoter and the basisof its linkage with SLE. An in silico analysis has suggested that the274 T mutation created a new Yin-Yang 1 (YY-1) binding siteadjacent to an existing one (10). YY-1 is a pleiotropic transcriptionfactor that can both upregulate and downregulate gene expressiondepending on the promoter context and the specific cellular envi-ronment (11, 12). It is a ubiquitously expressed 65-kDa protein thatbinds to a consensus 59-CCATNTT-39 sequence (13). The mecha-nism by which YY-1 regulates gene expression is complex and most
Department of Pathology, Immunology, and Laboratory Medicine, University ofFlorida, Gainesville, FL 32610
Received for publication July 8, 2011. Accepted for publication March 6, 2012.
This work was supported by Grant RO1 AI068965 from the National Institutes ofHealth (L.M.).
Address correspondence and reprint requests to Dr. Laurence Morel, Department ofPathology, Immunology and Laboratory Medicine, University of Florida, Gainesville,FL 32610-0275. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: B6, C57BL/6; CDK, cyclin-dependent kinase;CDKI, cyclin-dependent kinase inhibitor; ChIP, chromatin immunoprecipitation;p18, p18Ink4c; SDM, site-directed mutagenesis; siRNA, small interfering RNA;SL2, Schneider line 2; SLE, systemic lupus erythematosus; SNP, single nucleotidepolymorphism; spB, splenic B (cell); YY-1, Yin-Yang 1.
Copyright� 2012 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/12/$16.00
likely involves both coactivator and corepressor complexes regu-lating histone acetylation (11, 12). YY-1–interacting proteins in-clude basal transcription factors such as TBP (14), transcriptionalcoregulators such as p300/CREB, poly(ADP-ribosyl) polymerase,and several transcription factors such as SP-1, c-Myc, and C/EBPb(15). C-to-T promoter polymorphisms creating a novel YY-1 sitehave been identified in several human genes and have been associ-ated with immune-related diseases. The 2571 C/T in the IL-10promoter and 2509 C/T in the TGF-b promoter are associatedwith increased levels of both cytokines and with increased suscep-tibility to asthma (16, 17) and periodontitis (18). The 21993 C/Tpolymorphism in the TBX21 gene encoding for the TBET tran-scription factor has also been associated with an increased expres-sion by the T allele (19), leading to an increased susceptibility toSLE (20), resistance to hepatitis B virus infections (21), and asthma(22). The 21112 C/T mutation in the IL-13 promoter is associatedwith an increased transcription in individuals homozygous for the Tallele, which also have a greater susceptibility to allergic inflam-mation (23). Finally, two other SNPs that increase YY-1 binding onthe FCGRIIB promoter are associated with increased gene expres-sion and susceptibility to SLE (24). Interestingly, mutations thataffect YY-1 binding have not been reported in mice to our knowl-edge. In this study, we show that the274 region on the p18 promoteracts as an activator through the binding of YY-1 and transactivationby E2F1 and Sp-1. The274 C-to-T base exchange is consistent withthe creation of a second YY-1 site that relieves the activator and actsas a repressor of transcription. This is the first study, to our knowl-edge, to characterize the role of YY-1 in regulating the p18 promoterand to demonstrate a functional role for the 274 C-to-T SNP indecreasing p18 expression that is associated with the Sle2c1 lupussusceptibility locus.
Materials and MethodsIn silico p18 promoter analysis
A comprehensive library of hydroxyl radical cleavage profiles (availableonline at: dna.bu.edu/orchid) allows prediction of structural DNA profilesfrom sequence information in silico (25). Transcription factor binding siteson the p18 promoter were predicted using Chip MAPPER (http://mapper.chip.org/) (26) and TFSEARCH (http://www.cbrc.jp/research/db/TFSEARCH.html).
Reagents
All chemicals, Schneider Drosophila medium, glutamine, trypsin, andantibiotics were purchased from Sigma. The luciferase reporter vectorpGL-4 basic and the assay kit were purchased from Promega. RPMI,DMEM-high glucose, and DMEM-F-12 culture media were purchasedfrom Cellgro. FCS was obtained from Atlanta Biotech, and cell cultureplasticware was purchased from Corning.
Mice
The B6.Sle2c1 congenic strain is a C57BL/6 (B6) mouse that carries an ∼6Mb interval derived from the NZB genome on telomeric chromosome 4that contains the Cdkn2c gene (10). B6 mice were obtained from TheJackson Laboratory. All experiments were conducted according to proto-cols approved by the University of Florida Institutional Animal Care andUse Committee.
Cell culture
The NIH3T3 (mouse fibroblasts), Raji (human B lymphocytes), and theDrosophila melanogaster Schneider line 2 (SL2) cell lines were obtainedfrom the American Type Culture Collection. NIH3T3 cells were grown inDMEM supplemented with 10% FCS, 2 mM glutamine, 100 U/ml peni-cillin, and 50 mg/ml streptomycin. Raji cells were grown in RPMI sup-plemented with 10% v/v FCS, 100 U/ml penicillin, and 50 mg/ml strep-tomycin. Cell density was kept between 0.20 and 1.253 106 cells/ml. SL2cells were grown at 24˚C without supplemental CO2 as a loosely attachedmonolayer in Schneider Drosophila medium supplemented with 10% FCSand antibiotics.
Plasmids
pCaags–YY-1 and pCaags empty vector were provided by Prof. YukiYamaguchi (PRESTO, Chiyodaku, Japan), pPac–SP-1 and pPac emptyvector were provided by Prof. Yasumasa Iwasaki (Kochi University, Kochi,Japan), and pSG5L-E2F1 and pSG5L empty vector were provided by Dr.Shiwu Li (University of Florida). All plasmid sequences were confirmedby sequence analysis at the University of Florida Sequencing Core. Side-by-side transfections showed a clear p18 induction associated with theexpression of each transcription factor above that of the empty vectorcontrol (data not shown).
Reporter constructs
The 2279 p18 reporter construct was generated by cloning the promotersegment 279 nucleotide upstream from the putative translation initiation siteof the mouse p18 gene into the luciferase reporter plasmid pGL-4-basic (10).274 C and Tallele containing plasmids were cloned from the B6.Sle2c1 andB6 genomic DNA, respectively. Another252 construct that does not includethe274 SNP was also constructed as a negative control. Promoter DNAwasamplified using touchdown PCR with the primers shown in Table I. Frag-ments were digested with XhoI and Bglll restriction enzymes, and ligatedinto the pGL-4.10 basic vector digested with the same restriction enzymes.All constructs were confirmed by sequencing. Mutations in the p18 promoterwere generated by site-directed mutagenesis (SDM) on both promoterscarrying the C and T alleles with the primers shown in Table I. The locationand sequence of the mutations are depicted in Fig. 1A and 1B. PCR productswere digested with DpnI to cleave the parental strand and transformed intoSuper Competent cells (Invitrogen). The accuracy of the sequence of eachmutated promoter was verified by sequencing before usage.
Transient transfection, small interfering RNA, and luciferaseassays
The activity of the p18 promoter was determined by measuring the ex-pression of the luciferase reporter gene. Transient transfection assays inNIH3T3 cells were performed using Lipofectamine 2000 (Invitrogen) inserum-free medium in 12-well plate at 80290% cell confluence with 1.6mg reporter plasmids plus 160 ng of the Renilla luciferase plasmid pRL-TK (Promega) as an internal control for transfection efficiency. After 6 h ofincubation of the cells with the Lipofectamine:DNA complexes, the me-dium was changed to DMEM supplemented with 10% FCS and incubatedfor an additional 48 h at 37˚C. Primary spB cells were isolated by negativeselection from 8–11-wk-old B6 or B6.Sle2c1 mice using the MiltenyiB Cell Isolation kit. The purity of the preparation was typically ∼95%CD19+ B cells. After 24-h stimulation with LPS (50 mg/ml), cells (33 106)were transfected with 2.5 mg p18/Luc constructs along with 250 ng pRL-TK Renilla by using the Mouse B Cell Nucleofector Kit program Z-001(Amaxa; LONZA). The transfection efficiency of spB cells with the pMax-GFP vector was .90% of the nonadherent cells by fluorescent microscopy(data not shown). For in vitro small interfering RNA (siRNA) transfection,3 3 106 spB cells in 1 ml media were incubated with 5 mM YY-1 siRNA(sc-36864; Santa Cruz Biotech) in the Nucleofector transfection reagent atroom temperature with gentle agitation for 30 min, with or without ex-pression plasmids of YY-1. Transfected cells were incubated for 6–8 hbefore being subjected to the luciferase assays. Similarly, 2 3 106 RajiB cells were transiently transfected with 2.5 mg p18/Luc constructs alongwith 250 ng pRL-TK Renilla by using the Amaxa Nucleofector device(program V-01) with the Nucleofector Kit V. Transfected Raji cells wereincubated for 18 h before being subjected to the luciferase assays.Transfection of SL2 cells was performed using 25 ml Insectogene (Bion-tex) in serum-free medium with 2.5 mg p18 promoter-containing plasmidsand 250 ng pRL-TK. After 4 h of incubation of the cells with the Insec-togene:DNA complexes, the medium was changed to Schneider’s Dro-sophila medium supplemented with 10% FCS, and the cells were main-tained at 24˚C without supplemental CO2 for 48 h.
All transfected cells were collected by centrifugation and lysed in re-porter lysis buffer (Promega). Cell extracts were prepared using the DualLuciferase Assay System (Promega). Luciferase activity in cell extracts wasmeasured using a Lumat LB 9507 luminometer (Berthold Technologies).Firefly luciferase activity was normalized to Renilla luciferase activity.Results were expressed as luciferase fold induction by each construct overthe expression induced by pGL4 basic. These experiments were repeatedwith three independent DNA preparations.
Quantitative RT-PCR and Western blot
cDNAs from transfected spB cells were prepared using the Promega reversetranscription kit. Quantitative RT-PCR was performed with TaqMan probes
Mm00483243_m1 for Cdkn2c and Mm99999915_g1 for Gapdh (ABI)using the comparative cycle threshold method normalized to controlnontransfected spB cells. For Western blots, cells were lysed in lysis buffer(Promega) on ice for 60 min. After centrifugation, the supernatants wereassayed for protein concentration (Bradford assay; Bio-Rad). Sampleswere run on 12% SDS-PAGE gels. Proteins were electroblotted onto 0.45-mm pore size nitrocellulose filters, and the filters were blocked at roomtemperature with 5% nonfat milk in TBS and 1% Tween 20. Filters werethen incubated overnight with rabbit polyclonal anti–YY-1 (sc-1703) oranti-IgG–HRP (Southern Biotech). The blots were washed three times with0.1% Tween 20 in TBS and incubated with donkey anti-rabbit HRP(Sigma). Blots were developed using an ECL detection system (ECL;Amersham).
EMSA
Nuclear extracts were prepared using a kit (Active Motif) according to themanufacturer’s instructions. Protein content was determined using theBradford assay. All the forward strands of oligonucleotide probes (TableI) were biotinylated (ID Technologies) for detection using a light-shiftchemiluminescent EMSA kit (Pierce). After labeling, complementarystrands were mixed together in an equimolar ratio and allowed to annealfor 1 h at 37˚C to form double-stranded probe. EMSAs were performed byincubating 0.5 ng labeled probe with nuclear extract (5–10 mg) with orwithout competing unlabeled oligonucleotides at 10- to 100-fold excess inbinding buffer (10 mm Tris-HCl, pH 7.5, 50 mm Nacl, 1 mM DTT, 1 mMEDTA, 5% glycerol, and 1 mg/ml poly[dI×dC]) at room temperature for 30min. Mixtures were size fractionated on a nondenaturing 6% polyacryl-amide gel followed by transfer to Zeta nitrocellulose membranes (Bio-Rad) and detection by streptavidin-HRP/chemiluminescence for biotin-labeled probes. Supershift reactions were run as described earlier withthe addition of 2 mg rabbit polyclonal anti–YY-1 Ab (TRAPPC4), anti-E2F1 Ab (SC-193), or goat polyclonal anti–SP-1 Ab, all from Santa CruzBiotech. A Sox3 probe was used as an unrelated cold probe as control forbinding specificity. Density values for each band were calculated relativeto the band generated by the T or C allele oligonucleotide without com-petitor. Extracts from spB cells isolated from three B6 and three B6.Sle2c1mice by negative selection (Miltenyi) were analyzed, and each EMSAwasperformed at least three times.
Chromatin immunoprecipitation assays
Chromatin immunoprecipitation (ChIP) assays were performed using theMillipore kit according to the manufacturer’s protocol with minor mod-ifications. In brief, formaldehyde was added to the culture medium toa final concentration of 1%. Cells were incubated for 10 min at 37˚C andwashed twice with ice-cold PBS containing a protease inhibitor mixture(Sigma). After centrifugation, the pellets were resuspended in sonicationbuffer (1% SDS; 10 mM EDTA; 50 mM Tris-HCl, pH 8.1; protease in-hibitor mixture) and sonicated on ice to an average length of 500–1000 bpwith a Sonic Dismembrator (Fisher Scientific), then centrifuged at 12,000rpm. The chromatin solution was diluted 10-fold with dilution buffer(0.01% SDS; 1.1% Triton X-100; 1.2 mM EDTA; 16.7 mM Tris-HCl, pH
8.1; 167 mM NaCl), one-third of which was reserved as a total input ofchromatin. The diluted chromatin solution was precleared with salmonsperm DNA–protein A-agarose for 1 h and incubated with either anti-Nrf2(C-20), anti–YY-1 Ab (sc-1703), nonspecific rabbit IgG, or no Ab for 18 hat 4˚C with rotation. Immunoprecipitation, washing, and elution werecarried out according to the manufacturer’s instructions. Cross-linkedimmunoprecipitates and total chromatin inputs were reversed, and sam-ples were treated with proteinase K (Sigma), then extracted with phenol-chloroform-isoamyl alcohol (25:24:1). DNAwas precipitated with ethanoland resuspended in water. Quantitative PCR was performed with the pri-mers shown in Table I using 5 ml of template, with the Syber Green Real-Time PCR Detection System (Bio-Rad Laboratories) and analyzed usingthe CFX Manager Software v1.0 (Bio-Rad Laboratories).
Statistics
Statistical analyses were performed using the GraphPad Prism 5 Software.Two-tailed unpaired t tests were used, with multiple-test corrections asappropriate. The figures show means and SD, as well as statistical sig-nificance when p # 0.05.
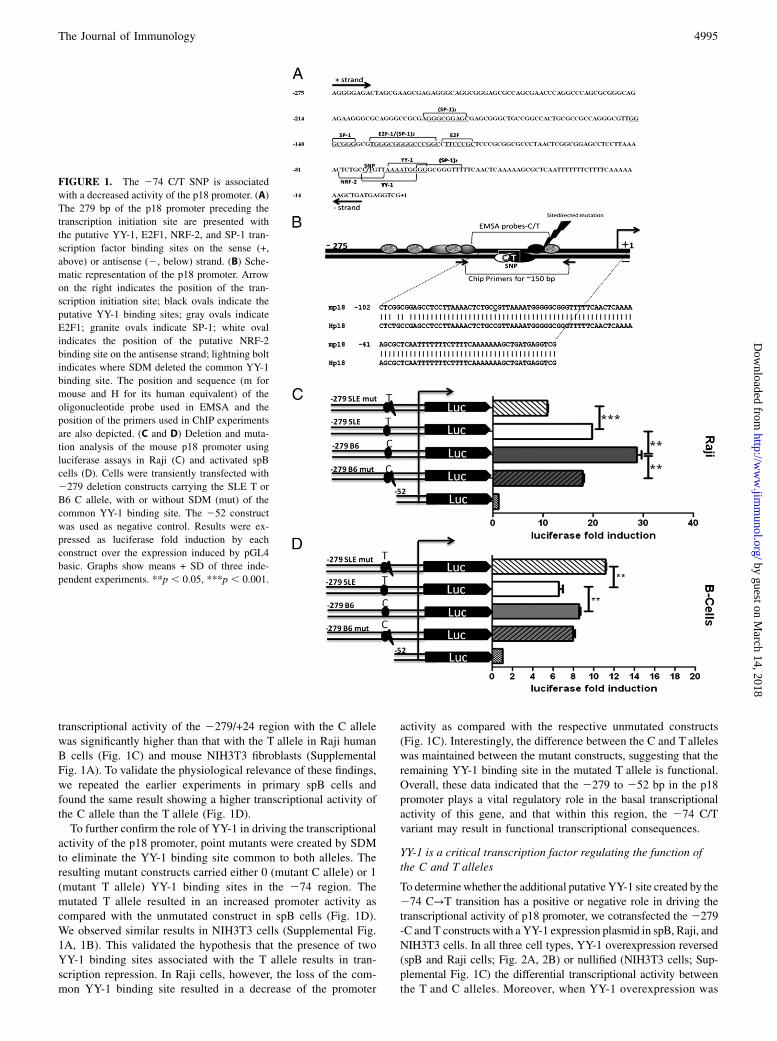
ResultsDecreased transcriptional activity of the p18 promoter with the274 T allele
The sequence of the p18 promoter from 21209 to +24 was ana-lyzed with Mapper and TFSEARCH to find putative transcrip-tional elements. This analysis predicted that the 274 C/T tran-sition results in the loss of binding sites for the NRF2 andHunchback transcription factors, whereas it creates a binding sitefor YY-1 that is adjacent to the already existing YY-1 site (Fig.1A, 1B). The first 150 bp of the promoter have a very high G + Ccontent (Fig. 1A), which is a hallmark of several TATA-less pro-moters (27). Several putative binding sites for E2F1 and SP-1,which transactivate the human p18 promoter (8), were identifiedin the mouse proximal promoter (Fig. 1A). BLAST alignmentrevealed that the first 100 nucleotides of the mouse and human p18promoter share 98% identity (Fig. 1B and data not shown). Thishigh degree of conservation suggests that these elements are likelyto play an important role in the regulation of p18 expression.Using deletion constructs (Table I), we have determined that the
2279 to 252 p18 region contains all elements that are necessaryfor basal transcription in mouse NIH3T3 and human Raji cells,and that the promoter construct carrying the 274 T variant wasassociated with a lower transcriptional activity (10). We extendedthese experiments to determine the functional significance of the274 C → T transition in the p18 promoter. We confirmed that the
Table I. List of oligonucleotides used in this study
aOligonucleotides used for dsDNA EMSA probes. Only the sequences of the biotinylated forward oligonucleotides are shown.bUnderlined bolded nucleotides indicate the 274 C/T SNP.cNucleotides mutated by SDM are indicated by a double underline.dXhoI site is underlined.eBglII site is underlined.
4994 YY-1 REGULATION OF THE Cdkn2c/p18 GENE EXPRESSION
transcriptional activity of the 2279/+24 region with the C allelewas significantly higher than that with the T allele in Raji humanB cells (Fig. 1C) and mouse NIH3T3 fibroblasts (SupplementalFig. 1A). To validate the physiological relevance of these findings,we repeated the earlier experiments in primary spB cells andfound the same result showing a higher transcriptional activity ofthe C allele than the T allele (Fig. 1D).To further confirm the role of YY-1 in driving the transcriptional
activity of the p18 promoter, point mutants were created by SDMto eliminate the YY-1 binding site common to both alleles. Theresulting mutant constructs carried either 0 (mutant C allele) or 1(mutant T allele) YY-1 binding sites in the 274 region. Themutated T allele resulted in an increased promoter activity ascompared with the unmutated construct in spB cells (Fig. 1D).We observed similar results in NIH3T3 cells (Supplemental Fig.1A, 1B). This validated the hypothesis that the presence of twoYY-1 binding sites associated with the T allele results in tran-scription repression. In Raji cells, however, the loss of the com-mon YY-1 binding site resulted in a decrease of the promoter
activity as compared with the respective unmutated constructs(Fig. 1C). Interestingly, the difference between the C and T alleleswas maintained between the mutant constructs, suggesting that theremaining YY-1 binding site in the mutated T allele is functional.Overall, these data indicated that the 2279 to 252 bp in the p18promoter plays a vital regulatory role in the basal transcriptionalactivity of this gene, and that within this region, the 274 C/Tvariant may result in functional transcriptional consequences.
YY-1 is a critical transcription factor regulating the function ofthe C and T alleles
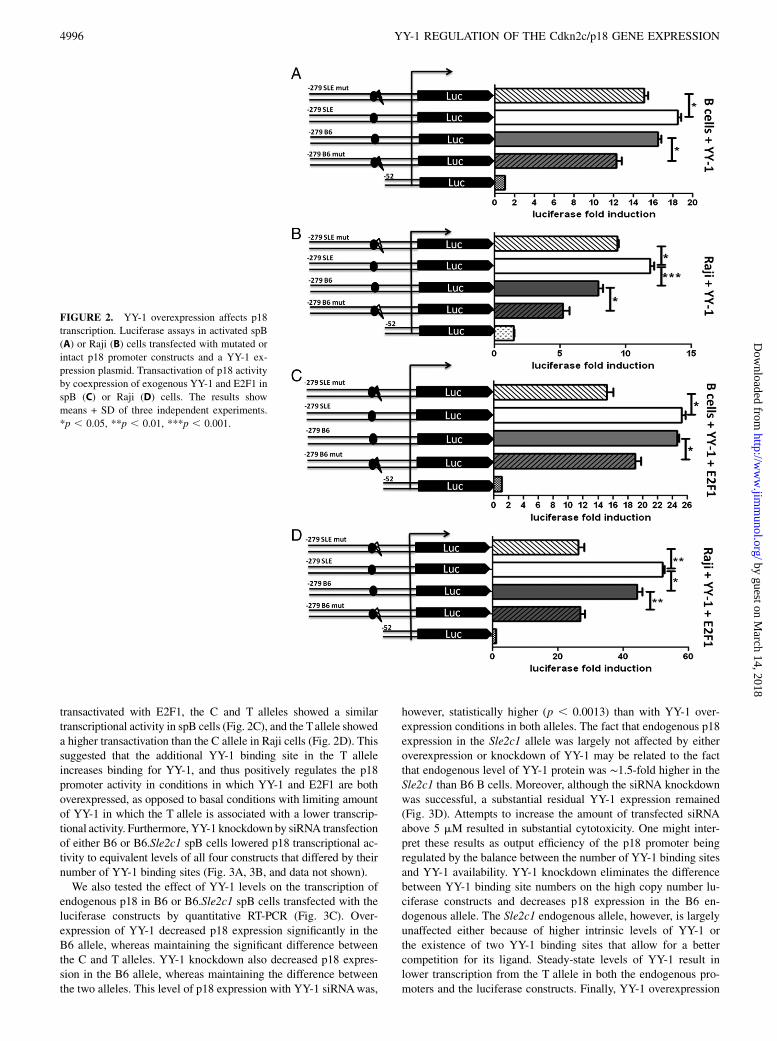
To determinewhether the additional putative YY-1 site created by the274 C→T transition has a positive or negative role in driving thetranscriptional activity of p18 promoter, we cotransfected the 2279-C and T constructs with a YY-1 expression plasmid in spB, Raji, andNIH3T3 cells. In all three cell types, YY-1 overexpression reversed(spB and Raji cells; Fig. 2A, 2B) or nullified (NIH3T3 cells; Sup-plemental Fig. 1C) the differential transcriptional activity betweenthe T and C alleles. Moreover, when YY-1 overexpression was
FIGURE 1. The 274 C/T SNP is associated
with a decreased activity of the p18 promoter. (A)
The 279 bp of the p18 promoter preceding the
transcription initiation site are presented with
the putative YY-1, E2F1, NRF-2, and SP-1 tran-
scription factor binding sites on the sense (+,
above) or antisense (2, below) strand. (B) Sche-
matic representation of the p18 promoter. Arrow
on the right indicates the position of the tran-
scription initiation site; black ovals indicate the
putative YY-1 binding sites; gray ovals indicate
E2F1; granite ovals indicate SP-1; white oval
indicates the position of the putative NRF-2
binding site on the antisense strand; lightning bolt
indicates where SDM deleted the common YY-1
binding site. The position and sequence (m for
mouse and H for its human equivalent) of the
oligonucleotide probe used in EMSA and the
position of the primers used in ChIP experiments
are also depicted. (C and D) Deletion and muta-
tion analysis of the mouse p18 promoter using
luciferase assays in Raji (C) and activated spB
cells (D). Cells were transiently transfected with
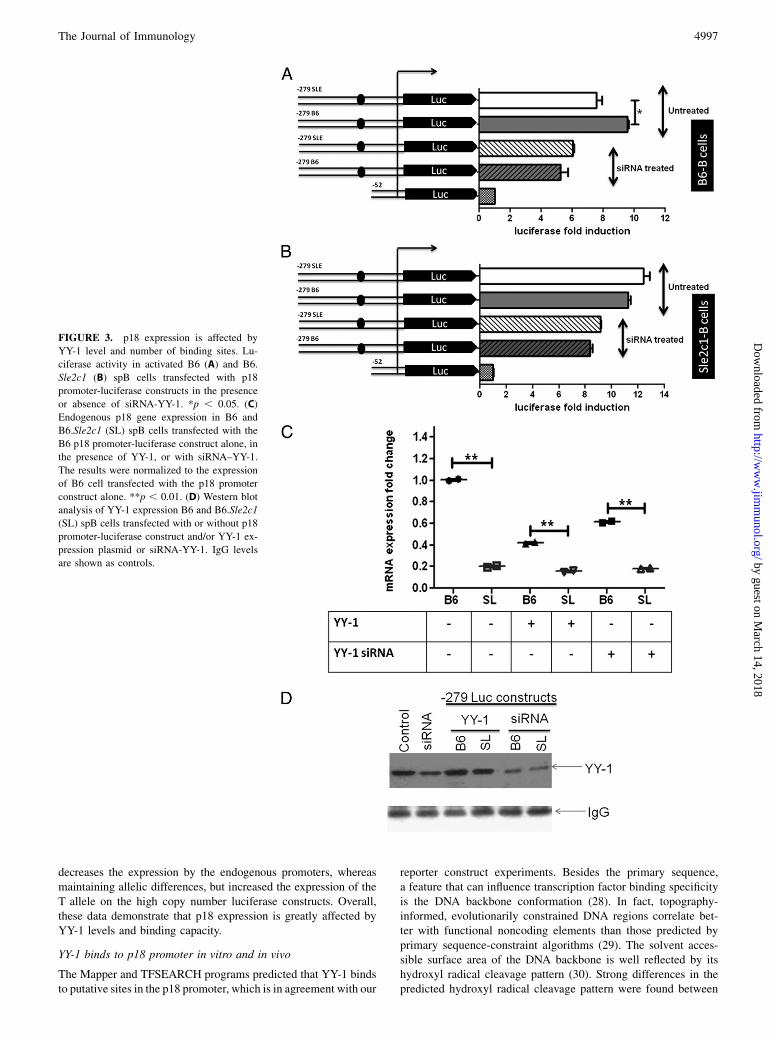
transactivated with E2F1, the C and T alleles showed a similartranscriptional activity in spB cells (Fig. 2C), and the Tallele showeda higher transactivation than the C allele in Raji cells (Fig. 2D). Thissuggested that the additional YY-1 binding site in the T alleleincreases binding for YY-1, and thus positively regulates the p18promoter activity in conditions in which YY-1 and E2F1 are bothoverexpressed, as opposed to basal conditions with limiting amountof YY-1 in which the T allele is associated with a lower transcrip-tional activity. Furthermore, YY-1 knockdown by siRNA transfectionof either B6 or B6.Sle2c1 spB cells lowered p18 transcriptional ac-tivity to equivalent levels of all four constructs that differed by theirnumber of YY-1 binding sites (Fig. 3A, 3B, and data not shown).We also tested the effect of YY-1 levels on the transcription of
endogenous p18 in B6 or B6.Sle2c1 spB cells transfected with theluciferase constructs by quantitative RT-PCR (Fig. 3C). Over-expression of YY-1 decreased p18 expression significantly in theB6 allele, whereas maintaining the significant difference betweenthe C and T alleles. YY-1 knockdown also decreased p18 expres-sion in the B6 allele, whereas maintaining the difference betweenthe two alleles. This level of p18 expression with YY-1 siRNAwas,
however, statistically higher (p , 0.0013) than with YY-1 over-expression conditions in both alleles. The fact that endogenous p18expression in the Sle2c1 allele was largely not affected by eitheroverexpression or knockdown of YY-1 may be related to the factthat endogenous level of YY-1 protein was ∼1.5-fold higher in theSle2c1 than B6 B cells. Moreover, although the siRNA knockdownwas successful, a substantial residual YY-1 expression remained(Fig. 3D). Attempts to increase the amount of transfected siRNAabove 5 mM resulted in substantial cytotoxicity. One might inter-pret these results as output efficiency of the p18 promoter beingregulated by the balance between the number of YY-1 binding sitesand YY-1 availability. YY-1 knockdown eliminates the differencebetween YY-1 binding site numbers on the high copy number lu-ciferase constructs and decreases p18 expression in the B6 en-dogenous allele. The Sle2c1 endogenous allele, however, is largelyunaffected either because of higher intrinsic levels of YY-1 orthe existence of two YY-1 binding sites that allow for a bettercompetition for its ligand. Steady-state levels of YY-1 result inlower transcription from the T allele in both the endogenous pro-moters and the luciferase constructs. Finally, YY-1 overexpression
FIGURE 2. YY-1 overexpression affects p18
transcription. Luciferase assays in activated spB
(A) or Raji (B) cells transfected with mutated or
intact p18 promoter constructs and a YY-1 ex-
pression plasmid. Transactivation of p18 activity
by coexpression of exogenous YY-1 and E2F1 in
spB (C) or Raji (D) cells. The results show
means + SD of three independent experiments.
*p , 0.05, **p , 0.01, ***p , 0.001.
4996 YY-1 REGULATION OF THE Cdkn2c/p18 GENE EXPRESSION
decreases the expression by the endogenous promoters, whereasmaintaining allelic differences, but increased the expression of theT allele on the high copy number luciferase constructs. Overall,these data demonstrate that p18 expression is greatly affected byYY-1 levels and binding capacity.
YY-1 binds to p18 promoter in vitro and in vivo
The Mapper and TFSEARCH programs predicted that YY-1 bindsto putative sites in the p18 promoter, which is in agreement with our
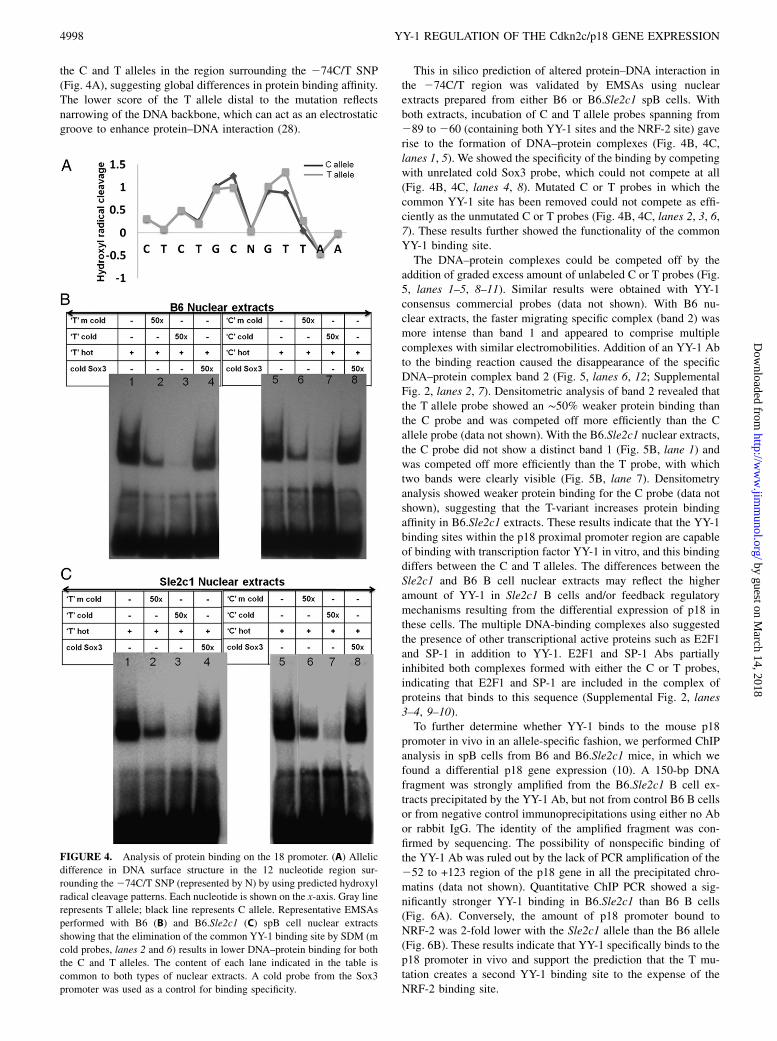
reporter construct experiments. Besides the primary sequence,a feature that can influence transcription factor binding specificityis the DNA backbone conformation (28). In fact, topography-informed, evolutionarily constrained DNA regions correlate bet-ter with functional noncoding elements than those predicted byprimary sequence-constraint algorithms (29). The solvent acces-sible surface area of the DNA backbone is well reflected by itshydroxyl radical cleavage pattern (30). Strong differences in thepredicted hydroxyl radical cleavage pattern were found between
the C and T alleles in the region surrounding the 274C/T SNP(Fig. 4A), suggesting global differences in protein binding affinity.The lower score of the T allele distal to the mutation reflectsnarrowing of the DNA backbone, which can act as an electrostaticgroove to enhance protein–DNA interaction (28).
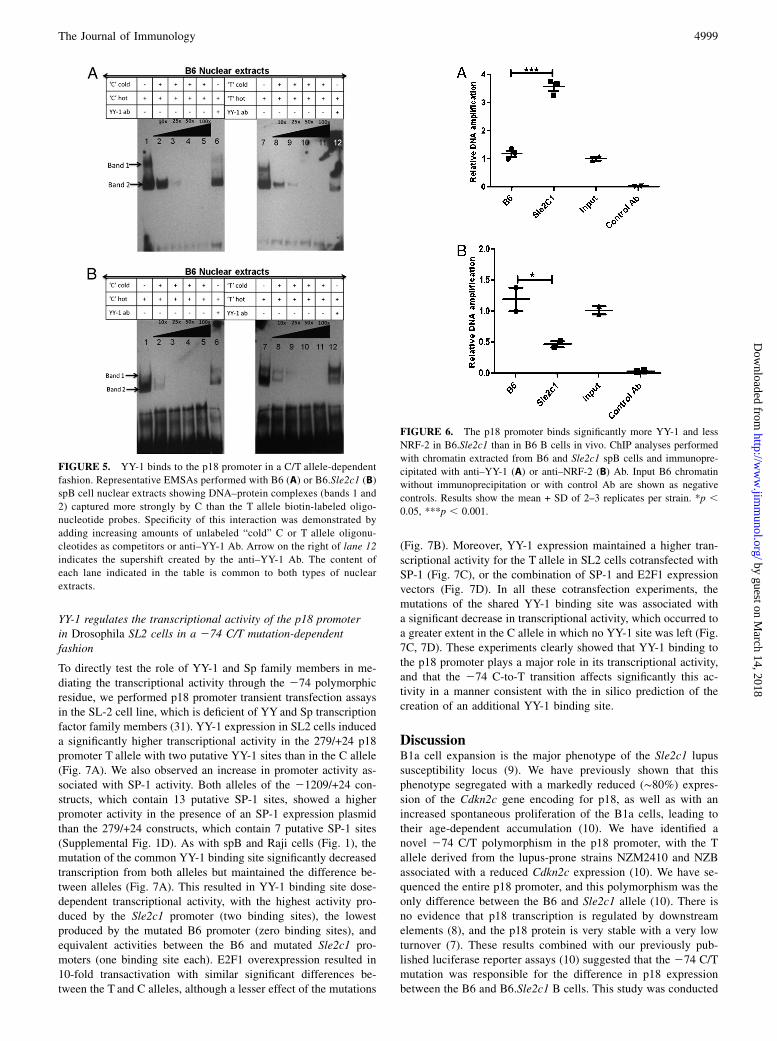
This in silico prediction of altered protein–DNA interaction inthe 274C/T region was validated by EMSAs using nuclearextracts prepared from either B6 or B6.Sle2c1 spB cells. Withboth extracts, incubation of C and T allele probes spanning from289 to 260 (containing both YY-1 sites and the NRF-2 site) gaverise to the formation of DNA–protein complexes (Fig. 4B, 4C,lanes 1, 5). We showed the specificity of the binding by competingwith unrelated cold Sox3 probe, which could not compete at all(Fig. 4B, 4C, lanes 4, 8). Mutated C or T probes in which thecommon YY-1 site has been removed could not compete as effi-ciently as the unmutated C or T probes (Fig. 4B, 4C, lanes 2, 3, 6,7). These results further showed the functionality of the commonYY-1 binding site.The DNA–protein complexes could be competed off by the
addition of graded excess amount of unlabeled C or T probes (Fig.5, lanes 1–5, 8–11). Similar results were obtained with YY-1consensus commercial probes (data not shown). With B6 nu-clear extracts, the faster migrating specific complex (band 2) wasmore intense than band 1 and appeared to comprise multiplecomplexes with similar electromobilities. Addition of an YY-1 Abto the binding reaction caused the disappearance of the specificDNA–protein complex band 2 (Fig. 5, lanes 6, 12; SupplementalFig. 2, lanes 2, 7). Densitometric analysis of band 2 revealed thatthe T allele probe showed an ∼50% weaker protein binding thanthe C probe and was competed off more efficiently than the Callele probe (data not shown). With the B6.Sle2c1 nuclear extracts,the C probe did not show a distinct band 1 (Fig. 5B, lane 1) andwas competed off more efficiently than the T probe, with whichtwo bands were clearly visible (Fig. 5B, lane 7). Densitometryanalysis showed weaker protein binding for the C probe (data notshown), suggesting that the T-variant increases protein bindingaffinity in B6.Sle2c1 extracts. These results indicate that the YY-1binding sites within the p18 proximal promoter region are capableof binding with transcription factor YY-1 in vitro, and this bindingdiffers between the C and T alleles. The differences between theSle2c1 and B6 B cell nuclear extracts may reflect the higheramount of YY-1 in Sle2c1 B cells and/or feedback regulatorymechanisms resulting from the differential expression of p18 inthese cells. The multiple DNA-binding complexes also suggestedthe presence of other transcriptional active proteins such as E2F1and SP-1 in addition to YY-1. E2F1 and SP-1 Abs partiallyinhibited both complexes formed with either the C or T probes,indicating that E2F1 and SP-1 are included in the complex ofproteins that binds to this sequence (Supplemental Fig. 2, lanes3–4, 9–10).To further determine whether YY-1 binds to the mouse p18
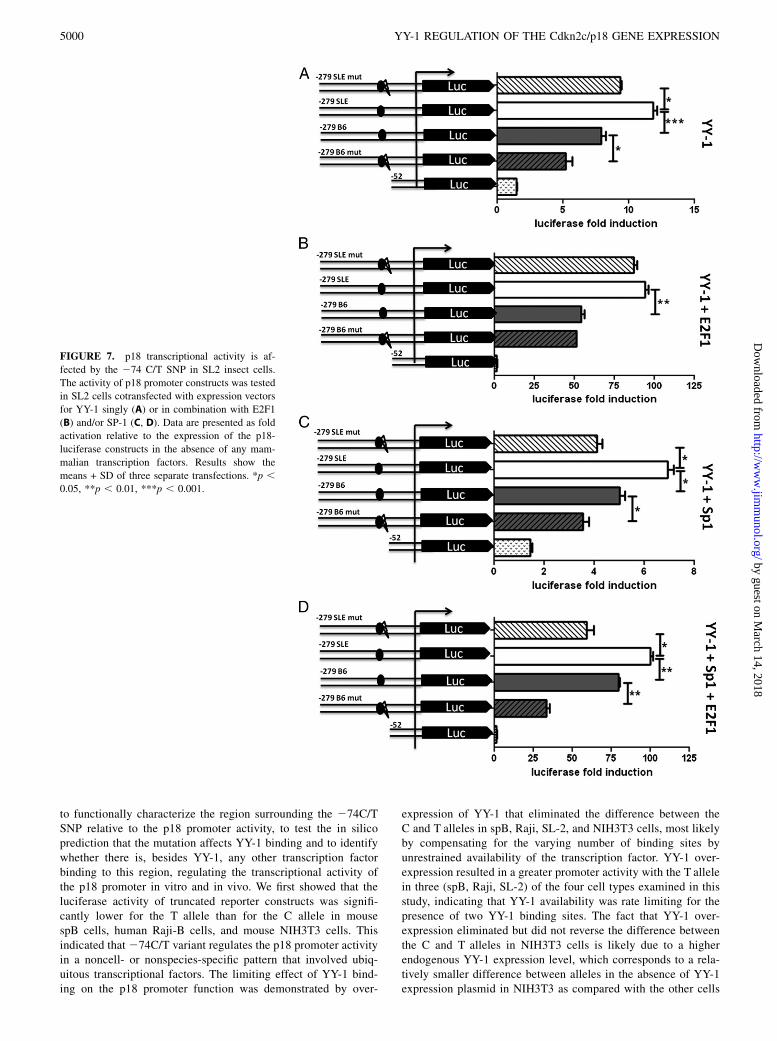
promoter in vivo in an allele-specific fashion, we performed ChIPanalysis in spB cells from B6 and B6.Sle2c1 mice, in which wefound a differential p18 gene expression (10). A 150-bp DNAfragment was strongly amplified from the B6.Sle2c1 B cell ex-tracts precipitated by the YY-1 Ab, but not from control B6 B cellsor from negative control immunoprecipitations using either no Abor rabbit IgG. The identity of the amplified fragment was con-firmed by sequencing. The possibility of nonspecific binding ofthe YY-1 Ab was ruled out by the lack of PCR amplification of the252 to +123 region of the p18 gene in all the precipitated chro-matins (data not shown). Quantitative ChIP PCR showed a sig-nificantly stronger YY-1 binding in B6.Sle2c1 than B6 B cells(Fig. 6A). Conversely, the amount of p18 promoter bound toNRF-2 was 2-fold lower with the Sle2c1 allele than the B6 allele(Fig. 6B). These results indicate that YY-1 specifically binds to thep18 promoter in vivo and support the prediction that the T mu-tation creates a second YY-1 binding site to the expense of theNRF-2 binding site.
FIGURE 4. Analysis of protein binding on the 18 promoter. (A) Allelic
difference in DNA surface structure in the 12 nucleotide region sur-
rounding the274C/T SNP (represented by N) by using predicted hydroxyl
radical cleavage patterns. Each nucleotide is shown on the x-axis. Gray line
represents T allele; black line represents C allele. Representative EMSAs
performed with B6 (B) and B6.Sle2c1 (C) spB cell nuclear extracts
showing that the elimination of the common YY-1 binding site by SDM (m
cold probes, lanes 2 and 6) results in lower DNA–protein binding for both
the C and T alleles. The content of each lane indicated in the table is
common to both types of nuclear extracts. A cold probe from the Sox3
promoter was used as a control for binding specificity.
4998 YY-1 REGULATION OF THE Cdkn2c/p18 GENE EXPRESSION
YY-1 regulates the transcriptional activity of the p18 promoterin Drosophila SL2 cells in a 274 C/T mutation-dependentfashion
To directly test the role of YY-1 and Sp family members in me-diating the transcriptional activity through the 274 polymorphicresidue, we performed p18 promoter transient transfection assaysin the SL-2 cell line, which is deficient of YY and Sp transcriptionfactor family members (31). YY-1 expression in SL2 cells induceda significantly higher transcriptional activity in the 279/+24 p18promoter T allele with two putative YY-1 sites than in the C allele(Fig. 7A). We also observed an increase in promoter activity as-sociated with SP-1 activity. Both alleles of the 21209/+24 con-structs, which contain 13 putative SP-1 sites, showed a higherpromoter activity in the presence of an SP-1 expression plasmidthan the 279/+24 constructs, which contain 7 putative SP-1 sites(Supplemental Fig. 1D). As with spB and Raji cells (Fig. 1), themutation of the common YY-1 binding site significantly decreasedtranscription from both alleles but maintained the difference be-tween alleles (Fig. 7A). This resulted in YY-1 binding site dose-dependent transcriptional activity, with the highest activity pro-duced by the Sle2c1 promoter (two binding sites), the lowestproduced by the mutated B6 promoter (zero binding sites), andequivalent activities between the B6 and mutated Sle2c1 pro-moters (one binding site each). E2F1 overexpression resulted in10-fold transactivation with similar significant differences be-tween the T and C alleles, although a lesser effect of the mutations
(Fig. 7B). Moreover, YY-1 expression maintained a higher tran-scriptional activity for the T allele in SL2 cells cotransfected withSP-1 (Fig. 7C), or the combination of SP-1 and E2F1 expressionvectors (Fig. 7D). In all these cotransfection experiments, themutations of the shared YY-1 binding site was associated witha significant decrease in transcriptional activity, which occurred toa greater extent in the C allele in which no YY-1 site was left (Fig.7C, 7D). These experiments clearly showed that YY-1 binding tothe p18 promoter plays a major role in its transcriptional activity,and that the 274 C-to-T transition affects significantly this ac-tivity in a manner consistent with the in silico prediction of thecreation of an additional YY-1 binding site.
DiscussionB1a cell expansion is the major phenotype of the Sle2c1 lupussusceptibility locus (9). We have previously shown that thisphenotype segregated with a markedly reduced (∼80%) expres-sion of the Cdkn2c gene encoding for p18, as well as with anincreased spontaneous proliferation of the B1a cells, leading totheir age-dependent accumulation (10). We have identified anovel 274 C/T polymorphism in the p18 promoter, with the Tallele derived from the lupus-prone strains NZM2410 and NZBassociated with a reduced Cdkn2c expression (10). We have se-quenced the entire p18 promoter, and this polymorphism was theonly difference between the B6 and Sle2c1 allele (10). There isno evidence that p18 transcription is regulated by downstreamelements (8), and the p18 protein is very stable with a very lowturnover (7). These results combined with our previously pub-lished luciferase reporter assays (10) suggested that the 274 C/Tmutation was responsible for the difference in p18 expressionbetween the B6 and B6.Sle2c1 B cells. This study was conducted
FIGURE 5. YY-1 binds to the p18 promoter in a C/T allele-dependent
fashion. Representative EMSAs performed with B6 (A) or B6.Sle2c1 (B)
spB cell nuclear extracts showing DNA–protein complexes (bands 1 and
2) captured more strongly by C than the T allele biotin-labeled oligo-
nucleotide probes. Specificity of this interaction was demonstrated by
adding increasing amounts of unlabeled “cold” C or T allele oligonu-
cleotides as competitors or anti–YY-1 Ab. Arrow on the right of lane 12
indicates the supershift created by the anti–YY-1 Ab. The content of
each lane indicated in the table is common to both types of nuclear
extracts.
FIGURE 6. The p18 promoter binds significantly more YY-1 and less
NRF-2 in B6.Sle2c1 than in B6 B cells in vivo. ChIP analyses performed
with chromatin extracted from B6 and Sle2c1 spB cells and immunopre-
cipitated with anti–YY-1 (A) or anti–NRF-2 (B) Ab. Input B6 chromatin
without immunoprecipitation or with control Ab are shown as negative
controls. Results show the mean + SD of 2–3 replicates per strain. *p ,0.05, ***p , 0.001.
to functionally characterize the region surrounding the 274C/TSNP relative to the p18 promoter activity, to test the in silicoprediction that the mutation affects YY-1 binding and to identifywhether there is, besides YY-1, any other transcription factorbinding to this region, regulating the transcriptional activity ofthe p18 promoter in vitro and in vivo. We first showed that theluciferase activity of truncated reporter constructs was signifi-cantly lower for the T allele than for the C allele in mousespB cells, human Raji-B cells, and mouse NIH3T3 cells. Thisindicated that 274C/T variant regulates the p18 promoter activityin a noncell- or nonspecies-specific pattern that involved ubiq-uitous transcriptional factors. The limiting effect of YY-1 bind-ing on the p18 promoter function was demonstrated by over-
expression of YY-1 that eliminated the difference between theC and T alleles in spB, Raji, SL-2, and NIH3T3 cells, most likelyby compensating for the varying number of binding sites byunrestrained availability of the transcription factor. YY-1 over-expression resulted in a greater promoter activity with the T allelein three (spB, Raji, SL-2) of the four cell types examined in thisstudy, indicating that YY-1 availability was rate limiting for thepresence of two YY-1 binding sites. The fact that YY-1 over-expression eliminated but did not reverse the difference betweenthe C and T alleles in NIH3T3 cells is likely due to a higherendogenous YY-1 expression level, which corresponds to a rela-tively smaller difference between alleles in the absence of YY-1expression plasmid in NIH3T3 as compared with the other cells
FIGURE 7. p18 transcriptional activity is af-
fected by the 274 C/T SNP in SL2 insect cells.
The activity of p18 promoter constructs was tested
in SL2 cells cotransfected with expression vectors
for YY-1 singly (A) or in combination with E2F1
(B) and/or SP-1 (C, D). Data are presented as fold
activation relative to the expression of the p18-
luciferase constructs in the absence of any mam-
malian transcription factors. Results show the
means + SD of three separate transfections. *p ,0.05, **p , 0.01, ***p , 0.001.
5000 YY-1 REGULATION OF THE Cdkn2c/p18 GENE EXPRESSION
types. Elimination of the YY-1 binding site common to bothalleles by SDM clearly showed a transcriptional activity of thep18 promoter was significantly affected in both alleles. Moreover,siRNA knockdown experiments showed that a low YY-1 ex-pression was also associated with equivalent transcriptional ac-tivity of all four alleles expression 0 to 4 YY-1 binding sites. Ininsect cells in which YY-1 activity can be tested without trans-activation by endogenous cofactors such as SP-1 and E2F1, ex-ogenous YY-1 activated the p18 promoter proportionally to thenumber of predicted YY-1 binding sites. Overall, these resultsdemonstrated that YY-1 by itself regulates the p18 promoter ac-tivity, and that the 274T mutation created a novel YY-1 bindingsite. Unfortunately, these in vitro results cannot be readily vali-dated in vivo because both B cell-specific YY-1 knockout (32)and overexpression of YY-1 in hematopoietic stem cells (33)eliminate mature B cells.YY-1 is a ubiquitous factor that mediates both transcriptional
activation and repression depending on both the sequence andcellular contexts (13). The comparison of sequences surroundingthe YY-1 binding site between promoters that are repressed andpromoters that are activated by YY-1 has determined consensusrepressor and activator sequences (13). Accordingly, the 274 Tpolymorphism creates a YY-1 repressor binding site, which fitswith the results that we have obtained with spB and Raji cells. Thevariable effects of YY-1 on the transcription of a same gene indifferent cell types have been clearly demonstrated for the humanIL-13 gene. The YY-1 binding site created by a C-to-T mutationincreased IL-13 expression in TH2 T cells because of STAT1 andSTAT6 transactivation, had no effect in nonpolarized CD4+ T cellsbecause of the binding of NFAT-2, and repressed IL-13 tran-scription in Jurkat T cells because of the absence of STAT6 andthe binding of OCT-1 (23). These complex gene–environmentinteractions can explain some of our results that showed differenteffects of the 274 C/T SNP in spB and Raji B cells, NIH3T3fibroblasts, and SL2 insect cells. The results obtained in the spBand Raji B cells are probably the most informative for p18 tran-scriptional regulation in B cells, and the repressive effect of the274 T mutation in these cells corresponds indeed to what wehave observed in mouse B cells (10).In vitro and in vivo assays confirmed that YY-1 binds to the
274 region of the p18 promoter. EMSA with B cell nuclearextracts produced multimeric complexes, which most likelyreflect multimer formation of YY-1 with other proteins likeE2F2 and SP-1. Support for the concept that the interactions ofYY-1 with gene promoters involves ternary complexes comesfrom studies on the IL-13 promoter where YY-1 forms a com-plex with STAT1 and a variable number of other factors (23),and on the IFN-g promoter that involves interaction betweenYY-1 and NFAT (34). In this study, a fast migrating complex(band 2) is an integral part of the YY-1/p18 promoter com-plexes, based on competitions with excess unlabeled probes,YY-1 consensus probes, C or T mutant probes, and supershiftassays. Band 2 was strongly bound by the 274 C probe ascompared with the 274 T probe with the B6 extracts, and thereverse was observed with Sle2c1 extracts. The fact that oppo-site results were obtained between the two strains may be a di-rect consequence of their different levels of p18 expression.This may lead to different physiological states through a dys-regulated cell cycle and different milieus of transcription factorsthat transactivate the p18 promoter on the YY-1 binding site inthese cells, in a similar fashion of what has been describedfor the IL-13 promoter between nonpolarized and TH2 CD4+
T cells (23). This hypothesis needs to be formally tested bycomparing the respective amounts of SP-1, NRF-2, and poten-
tially Hunchback available in B cells between the two strains.Additional and yet unidentified binding factors that may alsoplay a role in p18 transcription may be expressed at differentlevels depending on p18 expression.Finally, in vivo binding of YY-1 to the p18 promoter was
detected by ChIP assay, with significantly more binding to the Tallele in B6.Sle2c1 spB cells than to the C allele in spB B6 cells.ChIP assays also verified the prediction that the 274C/T SNPcaused the loss of NRF-2 binding site in T allele. NRF-2 is anantioxidant-activated transcription factor, and interestingly, theNrf2 targeted deletion has been associated with a multiorgan au-toimmune inflammatory syndrome that includes lupus-like mani-festations (35–37). Moreover, a polymorphism that affects Nrf-2expression level has been associated with lupus (38). Nrf-2 defi-ciency affects the expression of a large number of genes besidesthe antioxidant pathway (36), but it has not been reported toregulate p18 expression. It is intriguing that the loss of an Nrf-2binding site is also associated with lupus susceptibility in theSle2c1 locus, suggesting that the cell cycle regulation in B cellsand the size of the B1a cell compartment should be examined inNrf2-deficient mice.YY-1 has been implicated in the onset of the S phase (39) and in
the regulation of p16INK4a expression (40). Our results demon-strate that YY-1 plays a major role in regulating p18INK4c ex-pression, and that alterations in the number of its binding site onthe p18 promoter are associated with substantial consequenceson B cell homeostasis. Furthermore, our results suggest thatthe dominant binding of YY-1 created by the 274T element incombination with SP-1 plays a stronger negative regulatory roleon p18 promoter activity than the binding of NRF-2 in combi-nation with SP-1 that occurs with the C allele. Although the 274region on the p18 promoter is identical between mice and humans,the 274 C/T SNP has not been reported in humans. This does notmean that it does not exist, because the mouse mutation in theNZM2410/NZB genome that we have found was also novel. Itprobably indicates, however, that it may exist at a much lowerfrequency than in the IL-10, IL-13, TGF-B, and TBX21 genes, inwhich it has been found by multiple groups. We believe that ourfindings have provided not only significant insights on the regu-lation of the p18 promoter activity, but also have identified acausative variant responsible for the decreased p18 expression inthe Sle2c1 lupus susceptibility locus. Analysis of the ever in-creasing amount of genomic data should reveal whether the mouseSNP exists in the human p18 promoter and allow to test whether itis associated with the size of the newly identified human B1 cellcompartment (41), which is expanded in lupus patients (42) as inmany mouse models of lupus (43).
AcknowledgmentsWe thank Prof. Yuki Yamaguchi (PRESTO), Prof. Yasumasa Iwasaki (Kochi
University), and Dr. Shiwu Li (University of Florida) for generous gifts of
expression plasmids.
DisclosuresThe authors have no financial conflicts of interest.
References1. Sherr, C. J. 1994. The ins and outs of RB: coupling gene expression to the cell
cycle clock. Trends Cell Biol. 4: 15–18.2. Sherr, C. J., and J. M. Roberts. 1999. CDK inhibitors: positive and negative
regulators of G1-phase progression. Genes Dev. 13: 1501–1512.3. Schrantz, N., G. E. Beney, M. T. Auffredou, M. F. Bourgeade, G. Leca, and
A. Vazquez. 2000. The expression of p18INK4 and p27kip1 cyclin-dependentkinase inhibitors is regulated differently during human B cell differentiation. J.Immunol. 165: 4346–4352.
4. Morse, L., D. Chen, D. Franklin, Y. Xiong, and S. Chen-Kiang. 1997. Inductionof cell cycle arrest and B cell terminal differentiation by CDK inhibitor p18(INK4c) and IL-6. Immunity 6: 47–56.
5. Zhou, J. N., and S. Linder. 1996. Expression of CDK inhibitor genes in im-mortalized and carcinoma derived breast cell lines. Anticancer Res. 16(4A):1931–1935.
6. Suzuki, T., T. Narita, M. Uchida-Toita, and M. Yoshida. 1999. Down-regulationof the INK4 family of cyclin-dependent kinase inhibitors by tax protein ofHTLV-1 through two distinct mechanisms. Virology 259: 384–391.
7. Forget, A., O. Ayrault, W. den Besten, M. L. Kuo, C. J. Sherr, and M. F. Roussel.2008. Differential post-transcriptional regulation of two Ink4 proteins, p18 Ink4cand p19 Ink4d. Cell Cycle 7: 3737–3746.
8. Blais, A., D. Monte, F. Pouliot, and C. Labrie. 2002. Regulation of the humancyclin-dependent kinase inhibitor p18INK4c by the transcription factors E2F1and Sp1. J. Biol. Chem. 277: 31679–31693.
9. Xu, Z., B. Duan, B. P. Croker, E. K. Wakeland, and L. Morel. 2005. Geneticdissection of the murine lupus susceptibility locus Sle2: contributions to in-creased peritoneal B-1a cells and lupus nephritis map to different loci. J.Immunol. 175: 936–943.
10. Xu, Z., H. H. Potula, A. Vallurupalli, D. Perry, H. Baker, B. P. Croker,I. Dozmorov, and L. Morel. 2011. Cyclin-dependent kinase inhibitor Cdkn2cregulates B cell homeostasis and function in the NZM2410-derived murine lupussusceptibility locus Sle2c1. J. Immunol. 186: 6673–6682.
11. Shi, Y., J. S. Lee, and K. M. Galvin. 1997. Everything you have ever wanted toknow about Yin Yang 1....... Biochim. Biophys. Acta 1332: F49–F66.
12. Thomas, M. J., and E. Seto. 1999. Unlocking the mechanisms of transcriptionfactor YY1: are chromatin modifying enzymes the key? Gene 236: 197–208.
13. Shrivastava, A., and K. Calame. 1994. An analysis of genes regulated by themulti-functional transcriptional regulator Yin Yang-1. Nucleic Acids Res. 22:5151–5155.
14. Austen, M., B. Luscher, and J. M. Luscher-Firzlaff. 1997. Characterization of thetranscriptional regulator YY1. The bipartite transactivation domain is indepen-dent of interaction with the TATA box-binding protein, transcription factor IIB,TAFII55, or cAMP-responsive element-binding protein (CPB)-binding protein.J. Biol. Chem. 272: 1709–1717.
15. Seto, E., B. Lewis, and T. Shenk. 1993. Interaction between transcription factorsSp1 and YY1. Nature 365: 462–464.
16. Hobbs, K., J. Negri, M. Klinnert, L. J. Rosenwasser, and L. Borish. 1998.Interleukin-10 and transforming growth factor-beta promoter polymorphisms inallergies and asthma. Am. J. Respir. Crit. Care Med. 158: 1958–1962.
17. Pulleyn, L. J., R. Newton, I. M. Adcock, and P. J. Barnes. 2001. TGFbeta1 alleleassociation with asthma severity. Hum. Genet. 109: 623–627.
18. de Souza, A. P., P. C. Trevilatto, R. M. Scarel-Caminaga, R. B. de Brito, andS. R. Line. 2003. Analysis of the TGF-beta1 promoter polymorphism (C-509T)in patients with chronic periodontitis. J. Clin. Periodontol. 30: 519–523.
19. Li, J. R., J. G. Li, G. H. Deng, W. L. Zhao, Y. J. Dan, Y. M. Wang, and S. Chen.2011. A common promoter variant of TBX21 is associated with allele specificbinding to Yin-Yang 1 and reduced gene expression. Scand. J. Immunol. 73:449–458.
20. You, Y., W. Zhao, S. Chen, W. Tan, Y. Dan, F. Hao, and G. Deng. 2010. As-sociation of TBX21 gene haplotypes in a Chinese population with systemiclupus erythematosus. Scand. J. Rheumatol. 39: 254–258.
21. Chen, S., W. Zhao, W. Tan, B. Xu, Y. Dan, Q. Mao, X. Kuang, Y. Wang, andG. Deng. 2009. Association of TBX21 T-1993C polymorphism with viral per-sistence but not disease progression in hepatitis B virus carriers. Hepatol. Res.39: 716–723.
22. Suttner, K., P. Rosenstiel, M. Depner, M. Schedel, L. A. Pinto, A. Ruether, J.Adamski, N. Klopp, T. Illig, C. Vogelberg, et al. 2009. TBX21 gene variantsincrease childhood asthma risk in combination with HLX1 variants. J. AllergyClin. Immunol. 123: 1062–1068, 1068.e1–8.
23. Cameron, L., R. B. Webster, J. M. Strempel, P. Kiesler, M. Kabesch,H. Ramachandran, L. Yu, D. A. Stern, P. E. Graves, I. C. Lohman, et al. 2006.Th2 cell-selective enhancement of human IL13 transcription by IL13-1112C.T,a polymorphism associated with allergic inflammation. J. Immunol. 177: 8633–8642.
24. Su, K., X. Li, J. C. Edberg, J. Wu, P. Ferguson, and R. P. Kimberly. 2004. Apromoter haplotype of the immunoreceptor tyrosine-based inhibitory motif-bearing FcgammaRIIb alters receptor expression and associates with autoim-munity. II. Differential binding of GATA4 and Yin-Yang1 transcription factorsand correlated receptor expression and function. J. Immunol. 172: 7192–7199.
25. Greenbaum, J. A., B. Pang, and T. D. Tullius. 2007. Construction of a genome-scale structural map at single-nucleotide resolution. Genome Res. 17: 947–953.
26. Marinescu, V. D., I. S. Kohane, and A. Riva. 2005. MAPPER: a search engine forthe computational identification of putative transcription factor binding sites inmultiple genomes. BMC Bioinformatics 6: 79.
27. Azizkhan, J. C., D. E. Jensen, A. J. Pierce, and M. Wade. 1993. Transcriptionfrom TATA-less promoters: dihydrofolate reductase as a model. Crit. Rev.Eukaryot. Gene Expr. 3: 229–254.
28. Joshi, R., J. M. Passner, R. Rohs, R. Jain, A. Sosinsky, M. A. Crickmore,V. Jacob, A. K. Aggarwal, B. Honig, and R. S. Mann. 2007. Functional speci-ficity of a Hox protein mediated by the recognition of minor groove structure.Cell 131: 530–543.
29. Parker, S. C., L. Hansen, H. O. Abaan, T. D. Tullius, and E. H. Margulies. 2009.Local DNA topography correlates with functional noncoding regions of thehuman genome. Science 324: 389–392.
30. Balasubramanian, B., W. K. Pogozelski, and T. D. Tullius. 1998. DNA strandbreaking by the hydroxyl radical is governed by the accessible surface areas ofthe hydrogen atoms of the DNA backbone. Proc. Natl. Acad. Sci. USA 95: 9738–9743.
31. Courey, A. J., and R. Tjian. 1988. Analysis of Sp1 in vivo reveals multipletranscriptional domains, including a novel glutamine-rich activation motif. Cell55: 887–898.
32. Liu, H., M. Schmidt-Supprian, Y. Shi, E. Hobeika, N. Barteneva, H. Jumaa,R. Pelanda, M. Reth, J. Skok, K. Rajewsky, and Y. Shi. 2007. Yin Yang 1 isa critical regulator of B-cell development. Genes Dev. 21: 1179–1189.
33. Pan, X., M. Jones, J. Jiang, K. Zaprazna, D. Yu, W. Pear, I. Maillard, andM. L. Atchison. 2012. Increased expression of PcG protein YY1 negativelyregulates B cell development while allowing accumulation of myeloid cells andLT-HSC cells. PLoS ONE 7: e30656.
34. Soutto, M., F. Zhang, B. Enerson, Y. Tong, M. Boothby, and T. M. Aune. 2002. Aminimal IFN-gamma promoter confers Th1 selective expression. J. Immunol.169: 4205–4212.
35. Ma, Q., L. Battelli, and A. F. Hubbs. 2006. Multiorgan autoimmune inflamma-tion, enhanced lymphoproliferation, and impaired homeostasis of reactive oxy-gen species in mice lacking the antioxidant-activated transcription factor Nrf2.Am. J. Pathol. 168: 1960–1974.
36. Li, J., T. D. Stein, and J. A. Johnson. 2004. Genetic dissection of systemic au-toimmune disease in Nrf2-deficient mice. Physiol. Genomics 18: 261–272.
37. Yoh, K., K. Itoh, A. Enomoto, A. Hirayama, N. Yamaguchi, M. Kobayashi,N. Morito, A. Koyama, M. Yamamoto, and S. Takahashi. 2001. Nrf2-deficientfemale mice develop lupus-like autoimmune nephritis. Kidney Int. 60: 1343–1353.
38. Cordova, E. J., R. Velazquez-Cruz, F. Centeno, V. Baca, and L. Orozco. 2010.The NRF2 gene variant, -653G/A, is associated with nephritis in childhood-onsetsystemic lupus erythematosus. Lupus 19: 1237–1242.
39. Palko, L., H. W. Bass, M. J. Beyrouthy, and M. M. Hurt. 2004. The Yin Yang-1(YY1) protein undergoes a DNA-replication-associated switch in localizationfrom the cytoplasm to the nucleus at the onset of S phase. J. Cell Sci. 117: 465–476.
40. Wang, X., Y. Feng, L. Xu, Y. Chen, Y. Zhang, D. Su, G. Ren, J. Lu, andB. Huang. 2008. YY1 restrained cell senescence through repressing the tran-scription of p16. Biochim. Biophys. Acta 1783: 1876–1883.
41. Griffin, D. O., N. E. Holodick, and T. L. Rothstein. 2011. Human B1 cells inumbilical cord and adult peripheral blood express the novel phenotype CD20+CD27+ CD43+ CD70-. J. Exp. Med. 208: 67–80.
42. Griffin, D. O., and T. L. Rothstein. 2011. A small CD11b(+) human B1 cellsubpopulation stimulates T cells and is expanded in lupus. J. Exp. Med. 208:2591–2598.
43. Duan, B., and L. Morel. 2006. Role of B-1a cells in autoimmunity. Autoimmun.Rev. 5: 403–408.
5002 YY-1 REGULATION OF THE Cdkn2c/p18 GENE EXPRESSION