GK4, a G-protein-coupled receptor with aphosphatidylinositol phosphate kinase domain inPhytophthora infestans, is involved in sporangiadevelopment and virulence
Chenlei Hua,1† Harold J. G. Meijer,1*†
Jeroen de Keijzer,1‡
Wei Zhao,2 Yuanchao Wang2 andFrancine Govers1,3
1Laboratory of Phytopathology, Wageningen University,Droevendaalsesteeg 1, 6708 PB, Wageningen, theNetherlands.2Department of Plant Pathology, Nanjing AgriculturalUniversity, Nanjing 210095, China.3Centre for BioSystems Genomics, PO Box 98, 6700 ABWageningen, the Netherlands.
Summary
For dispersal and host infection plant pathogenslargely depend on asexual spores. Pathogenesis andsporulation are complex processes that are governedby cellular signalling networks including G-proteinand phospholipid signalling. Oomycetes possess afamily of novel proteins called GPCR-PIPKs (GKs) thatare composed of a seven-transmembrane spanning(7-TM) domain fused to a phosphatidylinositol phos-phate kinase (PIPK) domain. Based on this domainstructure GKs are anticipated to link G-protein andphospholipid signal pathways; however, their func-tions are currently unknown. Expression analyses ofthe 12 GK genes in Phytophthora infestans and theirorthologues in Phytophthora sojae, revealed differen-tial expression during asexual development. PiGK1and PiGK4 were fused to monomeric red fluorescentprotein (mRFP) and ectopically expressed in P. in-festans. In growing hyphae different subcellular distri-bution patterns were observed indicating that thesetwo GKs act independently during development. Wefocused on the functional analyses of PiGK4. Its locali-zation suggested involvement in cell differentiation
and elongation and its 7-TM domain showed a canoni-cal GPCR membrane topology. Silencing of GK4and overexpression of full-length and truncated con-structs in P. infestans revealed that PiGK4 is not onlyinvolved in spore germination and hyphal elongationbut also in sporangia cleavage and infection.
Introduction
Phytophthora species are filamentous organisms thatcause severe diseases in many plant species and areresponsible for huge economic losses in agriculture world-wide. The genus Phytophthora belongs to the oomycetes,a class in the stramenopile lineage that comprises sapro-phytes and pathogens of plants, animals and insects. Withrespect to growth morphology, propagation and spore dis-persal, oomycetes resemble fungi but, evolutionarily thetwo groups are unrelated (Latijnhouwers et al., 2003).Oomycetes that are notorious for their aggressiveness anddevastating effects on plants include the potato late blightpathogen Phytophthora infestans, the soybean stem androot rot pathogen Phytophthora sojae, downy mildewssuch as Bremia lactucae on lettuce and Hyaloperonosporaarabidopsidis on Arabidopsis thaliana, and variousPythium spp. causing seedling damping-off (Agrios, 2005).
For plant pathogens efficient spore production and dis-persal are prerequisites for success. The typical asexualspores produced by Phytophthora spp. are sporangia,oval-shaped structures that germinate directly or indirectly.In the case of P. infestans, sporangia germinate directly athigher temperatures whereas at lower temperatures cyto-plasmic cleavage results in a zoosporangium and theformation of zoospores. Upon release, the biflagellatezoospores swim around until they are triggered to encystand germinate. Germ tubes emerging from sporangia orencysted zoospores swell at the tip to form an appresso-rium, a structure that strongly adheres to the plant surface.A peg protruding from the appressorium penetrates theepidermis and, subsequently the host tissue is colonizedby the pathogen resulting in expanding lesions. After a fewdays sporulating hyphae emerge from the stomata, the
sporangia are dispersed and initiate a new infection cycle(Latijnhouwers et al., 2003; Agrios, 2005).
Although the environmental conditions that favour Phy-tophthora infection are relatively well known, the precisecues that trigger and activate the pathogen are still poorlyunderstood (Judelson and Blanco, 2005). Developmentalprocesses such as sporulation, zoosporogenesis and ger-mination rely on signalling cascades that are often drivenby biologically active molecules which are sensed byreceptors located in cell membranes. One class of recep-tors comprises G-protein-coupled receptors (GPCRs),typical seven-transmembrane spanning (7-TM) proteinswhich, upon activation, change their conformation. In theactivated conformation the C-terminal tail of the GPCRlocated at the cytosolic side of the membrane, interactswith the heterotrimeric G-protein, which then dissociatesinto the a subunit and the bg dimer. Subsequently, Ga andGbg further transmit signals into intracellular signallingcascades, thereby activating or repressing various compo-nents including phospholipid modifying and signallingenzymes (Oude Weernink et al., 2007). There is increasingevidence that in Phytophthora the signalling networksdirected by G-proteins and phospholipids are important forzoospore differentiation, asexual development and infec-tion. For example, Latijnhouwers et al. (2002), observedthat the G-protein activator mastoparan induces phos-pholipase D (PLD) activity and triggers the accumulation ofphosphatidic acid (PA) in zoospores, and that both mas-toparan and PAinduce zoospore encystment. Mutation of aPLD gene in P. sojae resulted in reduced growth (Lamouret al., 2006). Moreover, P. infestans mutants in which theGb subunit gene is silenced no longer sporulate whereasmutants of P. infestans and P. sojae with a silenced Gasubunit gene lose pathogenicity and produce abnormalzoospores that are impaired in swimming behaviourand chemotaxis (Latijnhouwers and Govers, 2003;Latijnhouwers et al., 2004; Hua et al., 2008).
Genome analysis revealed that each Phytophthoraspecies has over 50 genes encoding GPCRs (Tyler et al.,2006) but thus far hardly anything is known about theirfunctions. The only Phytophthora GPCR gene for which afunction has been hypothesized is GPR11 in P. sojae.Wang et al. (2010) showed that silencing GPR11 causeddefects in zoospore release, encystment and germination.Since this phenotype was not associated with down-regulation of genes that are suppressed in Ga-silencedmutants, GPR11 seems to function independent of the Gasubunit (Hua et al., 2008; Wang et al., 2010). The findingthat GPCRs drive signalling pathways independent of theheterotrimeric G-protein has been described previously forplants and humans (Heuss and Gerber, 2000; Ferguson,2001; Chen et al., 2004). Such alternative bypasses mightalso exist in Phytophthora. All species sequenced so farhave only one Ga subunit gene and one Gb subunit gene
(Laxalt and Govers, 2002; Tyler et al., 2006). In P. in-festans and P. sojae these genes can be silencedwithout losing viability (Latijnhouwers and Govers, 2003;Latijnhouwers et al., 2004; Hua et al., 2008) and it is there-fore unlikely that all GPCR-mediated activities requireheterotrimeric G-proteins. Moreover, Phytophthora pos-sesses a novel class of GPCR proteins named GPCR-PIPKs (hereafter abbreviated as GKs), in which anN-terminal 7-TM domain typical for GPCRs is combinedwith a C-terminal phosphatidylinositol phosphate kinase(PIPK) domain (Bakthavatsalam et al., 2006; Meijer andGovers, 2006). PIPKs phosphorylate phosphatidylinositolphosphate (PIP) isomers at their inositol ring and, depend-ing on their catalytic specificity, this phosphorylation resultsin different PIP2 isomers (Clarke et al., 2007). Theseisomers can act directly as second messengers or areprecursors for other second messengers; e.g. PI(4,5)P2 byitself is known to modulate many proteins whereas cleav-age of PI(4,5)P2 by phospholipase C results in the forma-tion of the second messengers inositol trisphosphate (IP3)and diacylglycerol (Meijer and Munnik, 2003).
The combination of a GPCR domain and a PIPK domainin a single protein is rather unique. Remarkably, among theGPCRs identified in P. sojae, 12 have this unique combi-nation (Tyler et al., 2006). In contrast, in all other non-oomycete eukaryotes only one GK gene has been found sofar and that is RpkA, one out of 55 GPCR genes in the slimemould Dictyostelium discoideum (Bakthavatsalam et al.,2006). Unlike the D. discoideum GK RpkA, which wasshown to be involved in cell density sensing, phagocytosisand bacterial defence (Bakthavatsalam et al., 2007; Riyahiet al., 2011), the Phytophthora GKs have not been func-tionally characterized.
The aims of this study were to determine the occurrenceof GK genes in plant pathogenic oomycetes outside thegenus Phytophthora and to gain insight into potential func-tions of GKs in oomycetes. Mining the genomes of thebroad-host-range species Pythium ultimum, (Levesqueet al., 2010) and the downy mildew H. arabidopsidis(Baxter et al., 2010), and comparing their GK gene familieswith those in P. infestans (Haas et al., 2009), P. sojae andP. ramorum, showed that GKs belong to the core set ofoomycete proteins that are well conserved. Furthermore,expression profiling of the 12 GK genes in P. infestans andP. sojae indicated potential involvement of some GKsin pre-infection stages. This was confirmed by functionalanalyses of one GK; gene silencing and overexpressionof P. infestans GK4 revealed a role of PiGK4 in the initiationof plant infection. Moreover, expression of fluorescentlytagged GKs showed that the subcellular localization ofGK4 differs from that of another GK. This spatial distribu-tion and the differential expression patterns during devel-opment suggest distinct functions for the various GKs inthe Phytophthora life cycle.
Previously, the full set of PIPKs was identified in P. sojaeand P. ramorum and divided into four classes named A, B,C and D (Meijer and Govers, 2006). Here we surveyed thegenome databases of P. infestans, H. arabidopsidis andPy. ultimum and identified 16 PIPK genes in P. infestansand Py. ultimum and 15 in H. arabidopsidis with ortho-logues for each of the four classes that were identified inP. sojae and P. ramorum. In all species the genes encod-ing type B and type C PIPKs are single copy geneswhereas for type A PIPKs there are two genes in eachspecies. All remaining PIPK genes encode type D PIPKs,the novel class of GPCR-PIPKs (GKs). In the genomes,PIPK genes are randomly dispersed without significantclustering and located in gene-dense regions that arelargely syntenic (not shown). ID codes, nucleotidesequences and predicted proteins of the PIPK genes inP. infestans, Py. ultimum and H. arabidopsidis are listed insupplemental files (Table S1).
Since the high number of the GKs in oomycetes couldbe due to ancestral gene duplication events we investi-gated their relationship by constructing a phylogram of allknown GKs, including D. discoideum RpkA, the only GKfound outside oomycetes. As shown in Fig. 1 there are 12clusters, one for each of the 12 GKs and a separate
branch for RpkA. Each cluster, except one, comprises fivegenes from five oomycete species suggesting one-to-oneorthologous relationships. The one exception is GK6with four orthologues; it is lacking in H. arabidopsidis.This phylogenetic distribution suggests that the GK genefamily already existed in the last common ancestor ofthese five species whereas the isolated position of RpkAreflects the evolutionary distance between oomycetesand D. discoideum. Taken together, the analyses showthat GK genes are well conserved among oomycetes.Since the same holds for the distribution over the fourPIPK classes and the gene numbers it seems that PIPKsfunction in processes and/or signalling pathways that arepreserved among plant pathogenic oomycetes.
Phytophthora GK genes are differentially expressedduring development
To get a first insight into the overall expression patterns ofGK genes we analysed whole-genome expression dataand EST data that are publicly available for P. infestansand P. sojae. Based on the P. infestans Nimblegen data(Haas et al., 2009) it seemed that all 12 PiGKs areexpressed during in vitro growth of mycelium on variousmedia. There was no indication for a strong up- or down-regulation during in planta growth at 2–5 days post inocu-lation (dpi). Moreover, the majority of the PiGK ESTs werederived from mycelium, sporangia and zoospores ESTlibraries whereas only one was obtained from in plantastages (Randall et al., 2005; Table S2). Similar GKexpression patterns were observed in P. sojae for whichwe extracted data from the P. sojae RNA-seq transcrip-tional database presented by Ye et al. (Ye et al., 2011;Yang et al., 2013, accompanying paper).
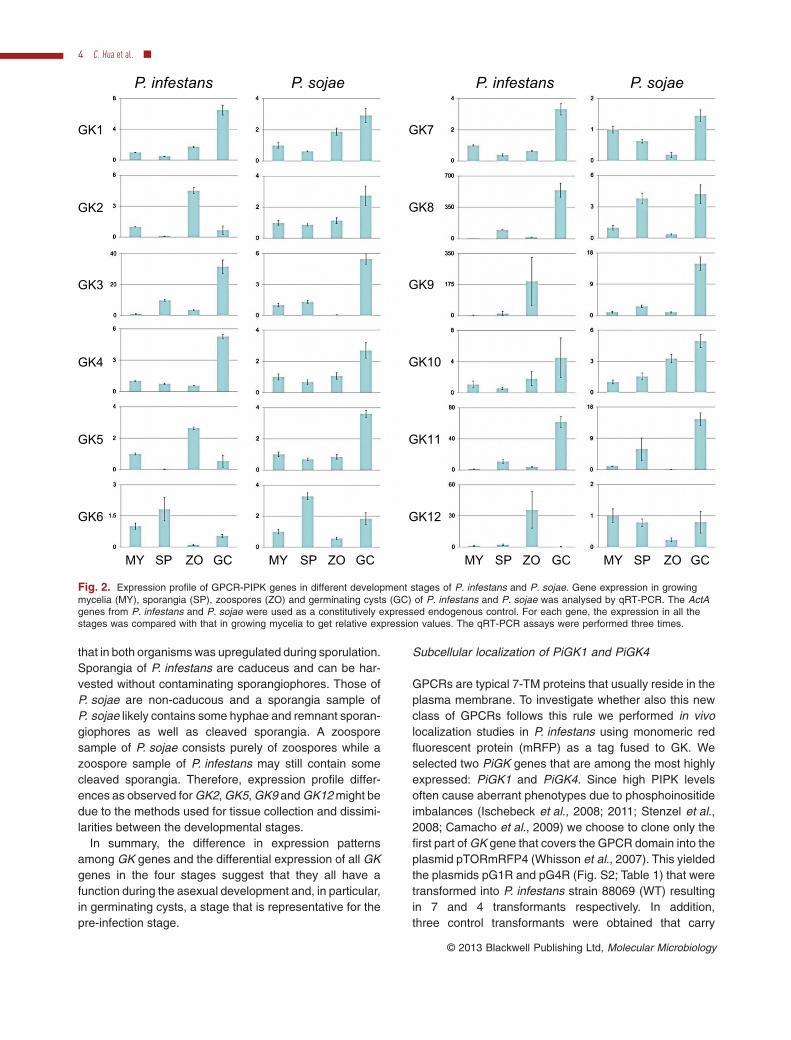
Based on the above in silico expression analyses wedecided to focus on GK gene expression during asexualdevelopment of P. infestans and P. sojae. Four stageswere analysed; growing mycelia, sporangia, zoosporesand germinating cysts. By using quantitative RT-PCR(qRT-PCR) we could show that seven P. infestans GKgenes have the highest expression during cyst germination(PiGK1, 3, 4, 7, 8, 10 and 11), four in zoospores (i.e. PiGK2,5, 9 and 12) and one (PiGK6) in sporangia (Fig. 2). Whenusing the expression level of the actin gene ActA asstandard for comparison, the highest expression amongthe 12 PiGK genes was observed for PiGK1, PiGK4 andPiGK11 (Fig. S1). In P. sojae all PsGK genes, with theexception of PsGK6, showed the highest expressionduring cyst germination. As in P. infestans, PsGK6 had thehighest expression in sporangia. When comparing theoverall profiles for the four stages in the two species, it isevident that the majority of the GK genes share theirhighest expression during cyst germination (Fig. 2). Theonly correlated exception is the PiGK6–PsGK6 gene pair
Fig. 1. Phylogeny of GPCR-PIPKs. Neighbour-Joinedphylogenetic tree of GPCR-PIPKs from Dictyostelium discoideum(Dd), Phytophthora infestans (Pi), Phytophthora sojae (Ps),Phytophthora ramorum (Pr), Hyaloperonospora arabidopsidis (Ha)and Pythium ultimum (Pu). The deduced full-length amino acidsequences of GPCR-PIPK genes were aligned by CLUSTAL_X 1.83and the phylogenetic tree was constructed by using Mega 4.0 byNeighbour-Joining method with 1000 bootstrap replicates.
Phytophthora GK4 in sporangia development and virulence 3
that in both organisms was upregulated during sporulation.Sporangia of P. infestans are caduceus and can be har-vested without contaminating sporangiophores. Those ofP. sojae are non-caducous and a sporangia sample ofP. sojae likely contains some hyphae and remnant sporan-giophores as well as cleaved sporangia. A zoosporesample of P. sojae consists purely of zoospores while azoospore sample of P. infestans may still contain somecleaved sporangia. Therefore, expression profile differ-ences as observed for GK2, GK5, GK9 and GK12 might bedue to the methods used for tissue collection and dissimi-larities between the developmental stages.
In summary, the difference in expression patternsamong GK genes and the differential expression of all GKgenes in the four stages suggest that they all have afunction during the asexual development and, in particular,in germinating cysts, a stage that is representative for thepre-infection stage.
Subcellular localization of PiGK1 and PiGK4
GPCRs are typical 7-TM proteins that usually reside in theplasma membrane. To investigate whether also this newclass of GPCRs follows this rule we performed in vivolocalization studies in P. infestans using monomeric redfluorescent protein (mRFP) as a tag fused to GK. Weselected two PiGK genes that are among the most highlyexpressed: PiGK1 and PiGK4. Since high PIPK levelsoften cause aberrant phenotypes due to phosphoinositideimbalances (Ischebeck et al., 2008; 2011; Stenzel et al.,2008; Camacho et al., 2009) we choose to clone only thefirst part of GK gene that covers the GPCR domain into theplasmid pTORmRFP4 (Whisson et al., 2007). This yieldedthe plasmids pG1R and pG4R (Fig. S2; Table 1) that weretransformed into P. infestans strain 88069 (WT) resultingin 7 and 4 transformants respectively. In addition,three control transformants were obtained that carry
GK1
GK2
GK3
GK4
GK5
GK6
GK7
GK8
GK9
GK10
GK11
GK12
MY SP ZO GC MY SP ZO GC MY SP ZO GC MY SP ZO GC
P. infestans P. sojae P. infestans P. sojae
Fig. 2. Expression profile of GPCR-PIPK genes in different development stages of P. infestans and P. sojae. Gene expression in growingmycelia (MY), sporangia (SP), zoospores (ZO) and germinating cysts (GC) of P. infestans and P. sojae was analysed by qRT-PCR. The ActAgenes from P. infestans and P. sojae were used as a constitutively expressed endogenous control. For each gene, the expression in all thestages was compared with that in growing mycelia to get relative expression values. The qRT-PCR assays were performed three times.
the vector pTORmRFP4. Epifluorescence microscopyrevealed that five pG1R (OXG1R1–5), two pG4R (OXG4R1and OXG4R2), and all three control transformants (C1–3)exhibited red fluorescence. Fluorescence intensity andlocalization of the mRFP signal was comparable betweenindividual transformants within each set of OXG1R, OXG4R
and C transformants. The presence and expression of thetransgenes was confirmed by PCR and RT-PCR respec-tively (data not shown). Overall, the colony morphologyand growth rate of the OX transformants was similar to thatof the recipient strain 88069 and the control transformants.
Sporangia obtained from two transformants per con-struct were germinated overnight after which the germtubes were examined by epifluorescence and confocalmicroscopy. In the control transformants C1 and C3, mRFPfluorescence was prominently found in the cytosol and insome motile vesicles of various sizes. These vesicles weredistributed randomly in the germ tubes but were absent inthe extreme tip region (Fig. 3A). In transformants OXG1R1and OXG1R3 fluorescence caused by the G1::mRFP fusionprotein was concentrated in membranes localized justbehind the tip of growing mycelium and surrounding larger
Table 1. Phytophthora infestans transformants used in this study.
a. pTORmRFP4 is the backbone vector of all other plasmids listed in the table. For details see supplemental files (Fig. S2).b. the recipient strain of all transformants is P. infestans 88069.
mR
FP
Brigh
tM
erg
ed
A B
C T=0s
T=1s
T=3s
T=4s
T=5s
T=6s
T=7s
T=8s
T=9s
T=10s
T=11s
T=2s
*
*
*
*
*
D
mR
FP
Brigh
tM
erg
ed
Fig. 3. Subcellular localization of mRFP andG1::mRFP in the hyphal tip of P. infestans.Sporangia of P. infestans C1 and OXG1R
transformants were germinated in RSM onbiofoil slides overnight, and the growing tipwas monitored by confocal microscopy.A. mRFP is present throughout the cytosoland in some motile vesicles of various sizeswhich are distributed randomly in the germtubes but seem absent from the extreme tipregion.B. G1::mRFP concentrated in membranessurrounding larger vacuolar-like compartmentsand occasionally, in vesicles moving along themembranes.C. G1::mRFP concentrated in the membranesof vacuolar-like compartments, inmicrofilament-like structures betweenvacuolar-like compartments, and in vesiclesconcentrated around vacuolar-likecompartments.D. Time series images illustrating movementsof G1::mRFP localized vesicles as shown inVideo S1. Asterisks indicate a small vesiclemerging with a large vesicle. Arrows indicatean appearing vesicle that moves subsequentlyover a long distance.Bars: 5 mm.
Phytophthora GK4 in sporangia development and virulence 5
vacuolar compartments (Fig. 3B). Microfilament-like struc-tures with fluorescence were often observed betweenthose vacuolar compartments (Fig. 3C). Motile punctatestructures with strong fluorescence were continuouslymoving along the larger cellular compartments and alongthe microfilament-like structures, occasionally concentrat-ing around the vacuolar compartments (Fig. 3D andVideo S1). During the microscopic monitoring we havenever seen the motile structures migrating into the hyphalapex, (defined as the extreme end of the hyphal tip) (Fig. 3and Video S2).
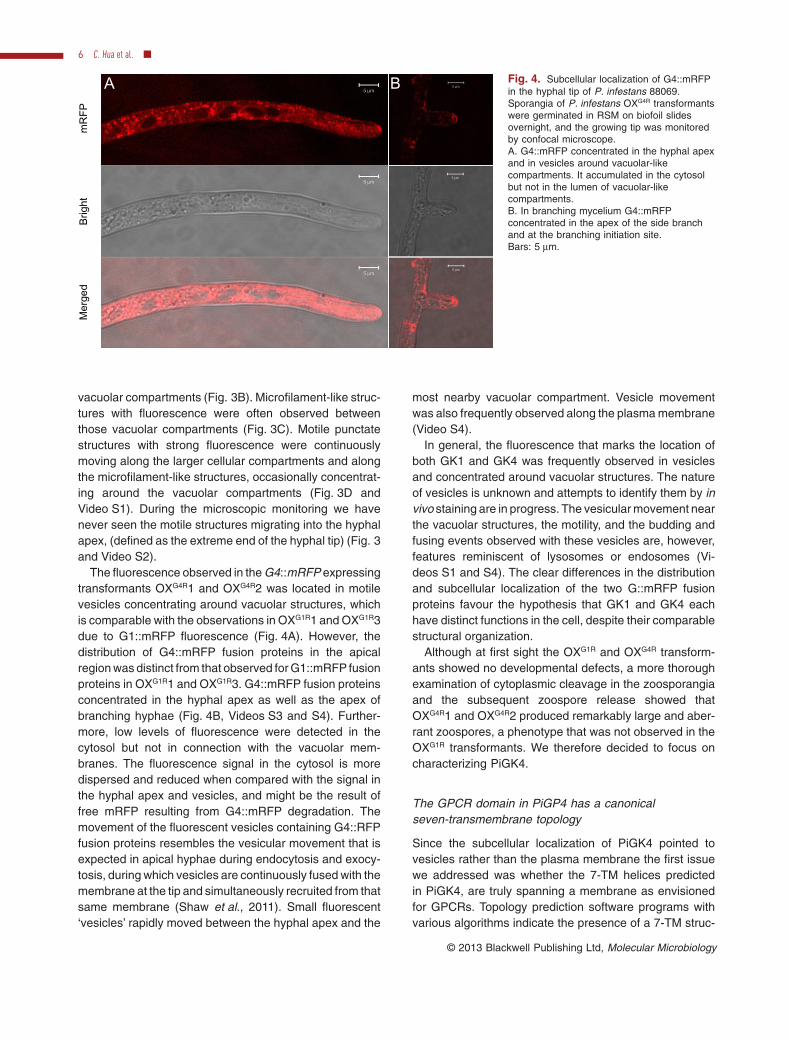
The fluorescence observed in the G4::mRFP expressingtransformants OXG4R1 and OXG4R2 was located in motilevesicles concentrating around vacuolar structures, whichis comparable with the observations in OXG1R1 and OXG1R3due to G1::mRFP fluorescence (Fig. 4A). However, thedistribution of G4::mRFP fusion proteins in the apicalregion was distinct from that observed for G1::mRFP fusionproteins in OXG1R1 and OXG1R3. G4::mRFP fusion proteinsconcentrated in the hyphal apex as well as the apex ofbranching hyphae (Fig. 4B, Videos S3 and S4). Further-more, low levels of fluorescence were detected in thecytosol but not in connection with the vacuolar mem-branes. The fluorescence signal in the cytosol is moredispersed and reduced when compared with the signal inthe hyphal apex and vesicles, and might be the result offree mRFP resulting from G4::mRFP degradation. Themovement of the fluorescent vesicles containing G4::RFPfusion proteins resembles the vesicular movement that isexpected in apical hyphae during endocytosis and exocy-tosis, during which vesicles are continuously fused with themembrane at the tip and simultaneously recruited from thatsame membrane (Shaw et al., 2011). Small fluorescent‘vesicles’ rapidly moved between the hyphal apex and the
most nearby vacuolar compartment. Vesicle movementwas also frequently observed along the plasma membrane(Video S4).
In general, the fluorescence that marks the location ofboth GK1 and GK4 was frequently observed in vesiclesand concentrated around vacuolar structures. The natureof vesicles is unknown and attempts to identify them by invivo staining are in progress. The vesicular movement nearthe vacuolar structures, the motility, and the budding andfusing events observed with these vesicles are, however,features reminiscent of lysosomes or endosomes (Vi-deos S1 and S4). The clear differences in the distributionand subcellular localization of the two G::mRFP fusionproteins favour the hypothesis that GK1 and GK4 eachhave distinct functions in the cell, despite their comparablestructural organization.
Although at first sight the OXG1R and OXG4R transform-ants showed no developmental defects, a more thoroughexamination of cytoplasmic cleavage in the zoosporangiaand the subsequent zoospore release showed thatOXG4R1 and OXG4R2 produced remarkably large and aber-rant zoospores, a phenotype that was not observed in theOXG1R transformants. We therefore decided to focus oncharacterizing PiGK4.
The GPCR domain in PiGP4 has a canonicalseven-transmembrane topology
Since the subcellular localization of PiGK4 pointed tovesicles rather than the plasma membrane the first issuewe addressed was whether the 7-TM helices predictedin PiGK4, are truly spanning a membrane as envisionedfor GPCRs. Topology prediction software programs withvarious algorithms indicate the presence of a 7-TM struc-
A Bm
RF
PB
righ
tM
erg
ed
Fig. 4. Subcellular localization of G4::mRFPin the hyphal tip of P. infestans 88069.Sporangia of P. infestans OXG4R transformantswere germinated in RSM on biofoil slidesovernight, and the growing tip was monitoredby confocal microscope.A. G4::mRFP concentrated in the hyphal apexand in vesicles around vacuolar-likecompartments. It accumulated in the cytosolbut not in the lumen of vacuolar-likecompartments.B. In branching mycelium G4::mRFPconcentrated in the apex of the side branchand at the branching initiation site.Bars: 5 mm.
ture in oomycete GKs but the outcome is not alwaysconsistent (Table S3). For PiGK4 for example, Conpred IIpredicts the canonical structure with the N-terminus local-ized at the extracellular or luminal membrane and theC-terminus intracellular, whereas HMMTOP predicts eightTM domains with both, the N-terminus and the C-terminusof PiGK4 extracellular. Since the membrane topology ofPiGK4 determines the location of the PIPK domain, thespatial organization of its GPCR domain was analysedexperimentally using an established method that is basedon the expression of a TM protein fused to the b-lactamasereporter enzyme in the prokaryote Escherichia coli. Thetopological position of the b-lactamase determines theresistance against carbenicillin (Fig. S3, Calamia andManoil, 1990; Lacatena et al., 1994; Schulein et al., 1996;Bhave et al., 2003). Seven truncated fragments of thePiGK4 gene of increasing length and covering one, two,three, four, five, six and seven TM domains, respectively,were cloned in such a way that fusion proteins wereproduced with a b-lactamase at the C-terminus (Fig. 5).The expression in E. coli was controlled by an IPTG-inducible promoter. As shown in Fig. 5, all transformantsgrew in the absence of carbenicillin indicating that theproduction of the chimeric GK4::b-lactamase enzymes didnot hamper bacterial growth. Nearly all transformants werecapable of growth in the presence of 10 mg ml-1 carbenicil-lin at high cell densities, probably due to the release ofb-lactamase from lysed neighbouring cells. Cells produc-ing fusion proteins with an uneven number of putative TMdomains did, however, not survive at lower densities, whilecells producing fusion proteins with an even number of TM
domains gained a discriminative ability to grow. This sen-sitivity is due to the fact that one TM domain in the fusionconstruct targets the b-lactamase enzyme to the cytoplasmwhile the N-terminus resides extracellular. The same holdsfor three, five and seven TM domains. In the case ofresistance, two, four and six TM domains make one, twoor three loops back and forth, respectively, with theb-lactamase enzyme ending up in the periplasmic space.Although cells carrying the larger constructs with four andsix TM domains did not survive at very low cell densities(1000–1 spotted cells), the difference in growth betweeneven and uneven numbers was obvious when the cellswere plated on medium containing lower concentrations ofcarbenicillin (2.5 and 1.25 mg ml-1) (Fig. 5). These resultsshow that the GPCR domain in PiGK4 has the canonical7-TM topology typical for GPCRs. However, the carbeni-cillin resistance of the full-length PiGK4 fusion construct ispuzzling. One would expect that the C-terminal PIPKdomain resides intracellular but it cannot be excluded thatthe PIPK domain itself associates with the plasma mem-brane or repositions the b-lactamase enzyme to the peri-plasmic space.
Overexpression of PiGK4 leads to aberrant sporangiaand stimulates germ tube elongation
The subcellular localization of G4::mRFP in motile vesiclessurrounding vacuolar structures and their aggregation inapical regions of expanding hyphae implicates a putativerole for PiGK4 during cell differentiation and elongation.To investigate this we generated, on the one hand,
LB+IPTG LB+IPTG/Carb
107 101 107 101
Carb2.5
Carb10
Carb1.25
Transmembrane domain Signal peptide-less β-lactamase domainPIPK domain
Fig. 5. Carbenicillin resistance assay of E. coli C41 transformed with b-lactamase fusion constructs in which the b-lactamase gene ispreceded by the full-length PiGK4 gene, the GPCR domain of PiGK4 and truncated versions of this GPCR domain. Suspensions of bacteriacarrying the PiGK4-b-lactamase fusion constructs shown on the left, were spotted in decreasing concentrations on plates without (shown inthe middle) and with (right) carbenicillin. Growth was recorded after 72 h incubation and this confirmed that equal numbers of bacteria werespotted of each construct (middle). On plates with carbenicillin the alternating pattern demonstrates that constructs with an even number ofTM domains enable the bacteria to hydrolyse the b-lactam antibiotic and hence to grow, whereas bacteria carrying constructs with an unevennumber of TM domains cannot grow. The ability to grow is determined by the location of the b-lactamase moiety which is either in theperiplasmic space or in the cytosol. With increasing numbers of TM domains in the fusion constructs decreasing concentrations of carbenicillinwere needed to discriminate between sensitive and resistant transformants. For details on the fusion constructs and the assay see Fig. S4.
Phytophthora GK4 in sporangia development and virulence 7
P. infestans transformants in which PiGK4 expressionis silenced and, on the other hand, transformants withincreased and constitutive expression of the full-lengthPiGK4 gene. For the latter we constructed plasmidpG4K4R with pTORmRFP4 as cloning vector. Expressionof PiGK4 is driven by the HAM34 promoter and should leadto a fusion protein with mRFP at the C-terminus (Fig. S2).Transformation of P. infestans strain 88069 with pG4K4Ryielded six G418-resistant transformants, three of which,named OXG4K4R1, 2 and 3, showed mRFP fluorescence andincreased PiGK4 expression when compared with therecipient WT strain (Fig. 6). The fluorescence was con-siderably weaker than in OXG4R1 and OXG4R2 but thelocalization was comparable (Fig. S4). For obtainingPiGK4-silenced transformants we did not perform DNAtransformations with specific silencing constructs butexploited the experience gained in previous studies thathomology-dependent gene silencing is a common phe-nomenon in Phytophthora (Latijnhouwers et al., 2004). Sowe analysed the PiGK4 mRNA levels in a variety of trans-formants carrying constructs with either full-length or trun-cated versions of PiGK4 and found four transformants inwhich the PiGK4 mRNA levels are reduced (Fig. 6). SP4R1and SP4R2 resulted from transformation with the plasmidpP4R that contains a truncated version of PiGK4 whereas
SG4K4R1 and SG4K4R2 resulted from transformation with theplasmid pG4K4R that contains a full-length of PiGK4(Fig. S2 and Table 1). As expected based on the level ofsequence homology between the GKs, silencing of PiGK4did not affect the mRNA levels of other PiGK genes (datanot shown).
Thus, stable transformants were obtained that eitheroverexpress PiGK4, or were silenced for PiGK4, allowingthe resultant phenotypes to be studied. When the over-expressing (OXG4K4R1, OXG4K4R2 and OXG4K4R3), silenced(SP4R1, SP4R2, SG4K4R1 and SG4K4R2) and control transform-ants (C1 and C3) were grown on rye agar medium theirgrowth behaviour was similar to that of the WT strain88069. Microscopic observations showed that hyphae ofOXG4K4R transformants appear a bit tortuous with a slightlyswollen tip (Fig. S4). There were no additional defectsand similar numbers of sporangia were produced (datanot shown). There were, however, more prominentdifferences during the development of sporangia andzoospores. When the morphology of sporangia, har-vested after 12 days of growth on rye sucrose agar (RSA)plates, was analysed by microscopy, the sporangia ofWT, C1, C3, and the four SP4R/SG4K4R transformants werehyaline (clear) and lemon-shaped. In contrast, OXG4K4R
transformants produced sporangia with aberrant cell
Re
lative
exp
ressio
n
3
2
1
0
4
5
6
Fig. 6. PiGK4 expression in P. infestans transformants. qRT-PCR analysis of PiGK4 expression using as template RNA from germinatingcysts of the wild-type strain 88069, control strains (C1 to C3), and transformants carrying constructs pP4R (T1 to T11) or pG4K4R (T12 toT19). In T1, T2, T15 and T17, PiGK4 expression is significantly reduced when compared with the wild-type strain. These silencedtransformants were renamed SP4R1, SP4R2, SG4K4R1 and SG4K4R2 respectively (Table 1). In contrast, in T12, T13 and T14 PiGK4 expression issignificantly increased. These overexpressing transformants were renamed OXG4K4R1, OXG4K4R2 and OXG4K4R3 respectively (Table 1). ActA fromP. infestans was used as a constitutively expressed endogenous control. The qRT-PCR experiment was repeated twice using RNA isolatedfrom germinating cysts harvested independently and at different dates.
walls, which were less transparent and appeared to belarger (Fig. 7A). The latter was quantified and indeed thesporangia produced by OXG4K4R1, OXG4K4R2 and OXG4K4R3were significantly larger than those produced by WT andother transformants (~ 45 mm versus 30 mm; Fig. 7B).DAPI staining revealed that also the nuclei in the spor-angia produced by the OXG4K4R transformants are consid-erable more fluorescent than those in wild type (Fig. 7A).In sporangia produced by the SP4R, SG4K4R and OXG4R
transformants, nuclei could not be visualized. It seemsthat in those sporangia the uptake of DAPI is blocked butwhether or not this is an aberrant phenotype that is cor-related with GK4 silencing or overexpression remains tobe investigated.
We then investigated whether the aberrant morphologyof the sporangia affects germ tube development. Upongermination, sporangia normally produce one germ tube.OXG4K4R1 and OXG4K4R2, however, developed two or moregerm tubes at a very high frequency (> 70%). In WT, C1,C3 and SP4R transformants this abnormal behaviour wasexceptional (Fig. 8A). Moreover, the germ tubes emerg-ing from sporangia of the OXG4K4R transformants grewvery fast. After overnight germination, the germ tubes ofOXG4K4R transformants had on average reached twice thelength than those of the other strains (Fig. 8A). To inves-tigate if similar phenotypes are observed during cyst ger-mination, we harvested zoospores, prepared cysts andallowed them to germinate in water for 2 h (Fig. 8B). Thegerm tube lengths of WT, C1 and C3 were comparablewhereas the growth of the germ tubes of the PiGK4-silenced strains SP4R1, SP4R2, SG4K4R1 and SG4K4R2 wereslightly impaired. In contrast, cysts derived from OXG4K4R1and OXG4K4R3 showed significantly increased lengths ofthe germ tubes when compared with WT, C1 and C3but this was less significant for OXG4K4R2 (Fig. 8C). Thedevelopment of multiple germ tubes as observed duringsporangial germination was not witnessed during cystgermination.
In summary, these data show that neither PiGK4 over-expression nor PiGK4 silencing affects the viability of thetransformants. In both cases viable sporangia are pro-duced. Overexpression, however, results in aberrant ger-mination of sporangia and increased lengths of germtubes emerging from sporangia and cysts.
Overexpression of the GPCR domain of PiGK4compromises cytoplasmic cleavage in zoosporangia
The increased size and aberrant cell walls of sporangiaproduced by OXG4K4R transformants (Fig. 7) may alsoinfluence the development of the zoosporangia. Thereforewe monitored the indirect germination in the various trans-formants and this pointed to additional defects related tocytoplasmic cleavage and the release of zoospores.
The sporangial cleavage rate of OXG4K4R transformants,based on the distribution of cleaved and uncleaved spo-rangia, was strongly reduced when compared with allother transformants and the WT strain (Fig. 9A). Pro-longed incubations did not improve the zoospore releaseefficiency. Released zoospores of OXG4K4R transformantswere morphologically comparable to those observed inthe control transformants and the WT strain (Fig. S5). Theone exception was the OXG4R1 and OXG4R2 transformants.These showed similar levels of zoospore release (Fig. 9A)but frequently the zoospores derived were remarkablylarge and aberrantly shaped (Figs 9 and S5), probably asa result of incomplete cleavage. The reduction in cleavagein full-length transformants and the deficiency in correctcytoplasmic cleavage in OXG4R1 and OXG4R2 transform-ants suggest that PiGK4 mediated signalling is essentialfor correct indirect germination of sporangia.
Silencing and overexpression of PiGK4 affectspathogenicity on potato and Nicotianabenthamiana leaves
The pathogenicity of all the PiGK4 transformants wastested on detached leaves of potato and Nicotianabenthamiana. WT and control strains had completelycolonized the potato leaves at 5 dpi (Fig. 10A), and pro-duced large amounts of sporangia (Fig. S6). Transform-ants OXG4K4R1, OXG4K4R2 and OXG4K4R3 infected as well asWT and control strains (Fig. 10). In contrast, OXG4R1 andOXG4R2 exhibited significant reduction of pathogenicity onthe host plant potato (Fig. 10A). Importantly, sporulationon infected leaves was diminished both for transformantsand for OXG4R1 hardly any spores were detected (Fig. S6).Also the four silenced transformants SP4R1, SP4R2, SG4K4R1and SG4K4R2 showed reduced pathogenicity but the level ofreduction was less than observed for OXG4R1 and OXG4R2.This was validated by quantification of the lesion sizes(Fig. 10B). On N. benthamiana leaves all transformants,except the control transformants, showed reduced growthat 7 dpi (Fig. S7).
Discussion
GPCR-PIPKs (GKs) belong to a novel class of PIPKs andwere only discovered when the genomes of the two firstsequenced Phytophthora species were annotated (Meijerand Govers, 2006; Tyler et al., 2006). Since then moreoomycete genomes have been sequenced and in thisstudy we show that in these oomycetes the PIPKs arestrongly conserved. They all have the four distinct classesof PIPKs, A to D, with three genes in class A and Bencoding type III PIPKs, one in class C encoding acommon type I/II PIPK and the remaining 11 or 12 genesin class D, the novel class, encoding a fusion between a
Phytophthora GK4 in sporangia development and virulence 9
Fig. 7. Sporangial shape of PiGK4-silenced and -overexpressing transformants.A. Sporangia from the P. infestans wild-type strain 88069 (WT), control transformant C1, GPCR domain-overexpressing transformants OXG4R1and OXG4R2, PiGK4-silenced transformants SP4R1 and SP4R2, and PiGK4-overexpressing transformants OXG4K4R1 and OXG4K4R2. In the panelson the right the sporangia are stained with DAPI. Sporangia were collected from 12-day-old mycelia growing on RSA plates. Bar = 50 mm.B. The average length of the sporangia from WT, control strains, and OXG4R, SP4R, SG4K4R and OXG4K4R transformants (n � 50). Asterisksindicate a significant difference (P < 0.05) using the Student’s t-test.
Fig. 8. Sporangia and cyst germination in PiGK4-silenced and -overexpressing transformants.A and B. (A) Germinating sporangia and (B) germinating cysts of the P. infestans wild-type strain 88069 (WT), control transformant C1, GPCRdomain-overexpressing transformants OXG4R1 and OXG4R2, PiGK4-silenced transformants SP4R1 and SP4R2, and PiGK4-overexpressingtransformants OXG4K4R1 and OXG4K4R2. The photos in (A) were taken after overnight incubation of sporangia at 18°C in the water.Bar = 500 mm. The photos in (B) were taken after 2 h incubation of cysts at 18°C in the water. Bar = 100 mm.C. Quantification of germ tube length of germinating cysts from WT, control strains, and OXG4R, SP4R, SG4K4R and OXG4K4R transformants(n � 50), which were incubated at 18°C for 2 h in the water. Asterisks indicate a significant difference (P < 0.05) using the Student’s t-test.
Phytophthora GK4 in sporangia development and virulence 11
7-TM domain resembling a GPCR and a novel type PIPKclassified as type IV (Bakthavatsalam et al., 2006). Type I,II and III PIPKs are known to function as kinases thatphosphorylate different PIP isomers leading to a variety ofPIP2 isomers. Whether or not type IV PIPKs have thesame catalytic activity with PIP as substrate remains to beinvestigated. It seems plausible, however, that the com-
bination of a GPCR domain and an enzymatic PIPKdomain has biological relevance. In this way, incomingsignals transmitted by the GPCR domain can be routeddirectly into downstream phosphoinositide signals withoutthe need for heterotrimeric G-proteins as intermediate.There is strong evidence that oomycetes need thesekinds of bypasses to stay alive. Most eukaryotes have
A
B
WT C1
OXG4R1 OXG4R2
SP4R1 SP4R2
OXG4K4R1 OXG4K4R2
Cle
ava
ge
(%
)
90
80
70
60
50
40
30
20
10
0
**
*
Fig. 9. Sporangia cleavage inPiGK4-silenced and -overexpressingtransformants.A. Cleaved and uncleaved sporangia of theP. infestans wild-type strain 88069 (WT),control transformant C1, GPCRdomain-overexpressing transformants OXG4R1and OXG4R2, PiGK4-silenced transformantsSP4R1 and SP4R2, and PiGK4-overexpressingtransformants OXG4K4R1 and OXG4K4R2. Thelarge and aberrant zoospores in the OXtransformants are indicated by black triangles.Sporangia of each strain were collected from12-day-old mycelia growing on RSA plates,flooded with cold water, and kept at 4°C for3 h. Subsequently the photos were taken.Bar = 100 mm.B. The percentage of sporangia showingcleavage. The data were obtained in threeindependent experiments, and in eachexperiment, at least 100 sporangia of eachtransformant were examined. Asterisksindicate a significant difference (P < 0.05)using the Student’s t-test.
Fig. 10. Infection assay of PiGK4-silencedand -overexpressing transformants ondetached potato leaves.A. Leaves of potato cultivar Impala 5 daysafter inoculation with the P. infestans wild-typestrain 88069 (WT), control transformant C1,GPCR domain-overexpressing transformantsOXG4R1 and OXG4R2, PiGK4-silencedtransformants SP4R1, SP4R2, SG4K4R1 andSG4K4R2, and PiGK4-overexpressingtransformants OXG4K4R1 and OXG4K4R2.Leaflets were inoculated with a 20 ml dropletcontaining 2 ¥ 103 zoospores and kept at18°C. Each transformant was inoculated on atleast six leaves comprising a total of at least24 inoculation sites.B. Quantification of lesion size. The diameterof each lesion was measured at 5 dpi andstatistics was done by Student’s t-test. Twoindependent infection assays were performedwith similar results. Asterisks indicate asignificant difference (P < 0.05) using theStudent’s t-test.
Phytophthora GK4 in sporangia development and virulence 13
multiple Ga and Gb subunits that often have distinct func-tions in signalling. Fungi, for example, have two to four Gasubunits, and knockout mutagenesis of each of theseleads to distinct phenotypes (Li et al., 2007). In contrast,Phytophthora spp. have only one canonical Ga and onecanonical Gb subunit gene, gpa1 and gpb1 (Laxalt andGovers, 2002). Silencing gpa1 affects, among others,zoospore behaviour and virulence (Latijnhouwers et al.,2004; Hua et al., 2008) whereas silencing gpb1 leads toa severe reduction in sporulation (Latijnhouwers andGovers, 2003). Despite these phenotypes that severelydisable Phytophthora to function as pathogen, thesilenced strains remain viable, pointing to the existence ofalternative signalling routes independent of Ga and Gb.GKs represent a novel family of receptor signalling mol-ecules that may function independently of Ga or Gb. Thisis supported by the fact that oomycetes each have a largenumber of distinct GKs that, similar to the multiple Gasubunits in other eukaryotes, may have specialized func-tions. In fact, this study and the accompanying paper byYang et al. (2013) demonstrate potentially distinct func-tions for Phytophthora GKs because the phenotypesof silenced transformants as well as their subcellularlocalization differ. GKs are also illustrative examples ofoomycete-specific proteins with novel domain combina-tions. Recently, Seidl et al. (2011) showed that, in com-parison with other taxonomic classes, oomycetes have aremarkably high number of proteins with distinct domaincombinations (or bigrams). Several bigrams highlighted inthat study, because of their abundance, include domainsrelated to phospholipid signalling. To our knowledge, ourstudies on GKs are the first addressing biological func-tions of oomycete proteins with a novel domain architec-ture discovered by bioinformatics analysis.
Apart from the single copy gene RpkA in the slime mouldD. discoideum and a few potential candidates in otherslime moulds (Bakthavatsalam et al., 2007; Riyahi et al.,2011), no other GK genes outside oomycetes are knownand this raised the question how the large GK families inoomycetes evolved. Phylogenetic analysis revealed thatthe 12 orthologous pairs of GKs initially identified inP. sojae and P. ramorum (Meijer and Govers, 2006) haveorthologues in all oomycetes included in this study. Hence,the 12 GK genes apparently resulted from gene-duplication events in their last common progenitor andcould point to an ancient functional diversification that wasmaintained after speciation. The pattern of orthologues ineach species without any evidence for additional intraspe-cific duplication events is in contrast to several other genefamilies in oomycetes. For example, a recent studyshowed that sPLD-like-Bs (Meijer et al., 2011) form sub-populations of paralogues per species indicating direc-tional evolution. The large-scale evolutionary dynamicsthat is accompanied with complex and distinct patterns of
gene duplication and losses and considered to be a typicalfeature of oomycete genomes (Seidl et al., 2012), didhardly affect the GK family. Of the 90 genes (16 PIPKsubfamilies including 12 GKs in five species) only one ismissing: GK6, the gene that is most abundantly expressedin sporangia, seems to be lost in H. arabidopsidis. Sincethis is the only species out of five that does not producezoospores during asexual development (Baxter et al.,2010), we speculate that GK6 has a function in zoosporeformation, sporangia cleavage or zoospore behaviour.
Of the many GPCRs that have been described in litera-ture, very few, if any, have additional known domains asC-terminal fusion partner. The peculiar domain composi-tion of GKs raised the question if the GPCR domain inGKs has the canonical 7-TM topology that is typical forGPCRs. We addressed this by using an established topol-ogy assay for TM proteins in E. coli based on a seriesof truncated PiGK4 constructs fused to b-lactamase(Fig. S3). This assay confirmed that each subsequent pre-dicted TM domain of PiGK4 forced another crossing of themembrane. Moreover, based on the alternating patternsof antibiotic resistance, loops connecting uneven to evensegments were found to be located in the cytosol and thisresembles the membrane topology of a human GPCRthat was assayed in a similar manner (Bhave et al., 2003).Two other features support the topology; the observedconfiguration obeys the ‘positive inside’ rule (Nilsson andvon Heijne, 1990) and the predicted extracellular loops 1and 2 contain cysteine residues, as is the case in over90% of the GPCRs (Petäjä-Repo et al., 2005). Accordingto this topology the N-terminus of PiGK4 is located at theextracellular or luminal leaflet of the membrane, and theC-terminus at the inner (cytoplasmic) leaflet of the mem-brane. Altogether, these data support our view that theGPCR domain in the GKs has a canonical 7-TM lay-outwhich is in accordance with other GPCRs.
In the topology assay we also included a full-lengthPiGK4 construct anticipating that we could confirm thatalso in the presence of PIPK domain the 7-TM topologywas maintained. Unexpectedly, we observed a reposition-ing of the full-length protein as opposed to solely theGPCR domain. This could be due to an imbalancedprotein charge, as was postulated by Nilsson and vonHeijne (1990) or to a small hydrophobic window that ispresent directly preceding the activation loop that coversthe conserved IID motif (Bakthavatsalam et al., 2006) anddirectly flanked by the dibasic KK amino acid pair that wasshown to mediate the association of PIPKs to the plasmamembrane (Mikami et al., 2010). This membrane associa-tion, however, should not involve membrane penetration.It cannot be excluded though that a prokaryote host rec-ognizes this hydrophobic fragment as a TM and, hence,that the antibiotic resistance of the full-length construct isdue to an artefact. This awaits further investigation.
To determine whether also in Phytophthora PiGK4resides in membranes as predicted by the 7-TM topologyof the GPCR domain, we generated P. infestans transform-ants in which the mRFP-tagged GPCR domain of PiGK4 isconstitutively expressed (OXG4R1 and 2). For comparisonwe included transformants expressing the GPCR domainof another member of the GK family, PiGK1 (OXG1R1 and3). Microscopic analysis revealed dissimilar localizationpatterns for G1::mRFP and G4::mRFP. None of the twowas prominently detected in the plasma membrane.Instead, most fluorescence resided in small vesicles, manyof which exhibited rapid motility. The main difference wasobserved at the hyphal apex, with a prominent accumula-tion of PiGK4 in the tip of the mycelium and at the site ofbranching whereas PiGK1 did not accumulate in the tips.Transformants expressing full-length mRFP fusion pro-teins (OXG4K4R1, 2 and 3) showed significantly lowerfluorescence intensity but their distribution matched theobservations made for GPCR::mRFP fusion proteins.Similar to PiGK1 and PiGK4, PsGK5 in P. sojae does notlocalize to the plasma membrane but resides in fast movingvesicles that might be different from the ones that containPiGK1 or PiGK4 (Yang et al., 2013, accompanying paper).In other organisms, GPCRs are primarily localized inplasma membranes although presence in intracellularorganelles including ER, Golgi and endosomes has beenreported (Marchese et al., 2008; Toshima et al., 2009;Hislop and von Zastrow, 2011). Exposure to ligands oragonists, frequently causes rapid redistribution of GPCRsfrom plasma membranes into the cell interior, often intopunctuated structures due to ligand induced endocytosis.Similar to GKs in Phytophthora the GK RpkA in D. discoi-deum was not detected in the plasma membrane butrather in intracellular membranes and compartments(Bakthavatsalam et al., 2007). The lack of plasma mem-brane localization and the resemblance in localizationamong GKs indicate that GKs preferably reside inside thecells. One has to be careful, however, to draw too firmconclusions. In all cases these studies are performed ontransformants in which the gene of interest is expressedconstitutively or at high levels and this may result in arte-facts. We cannot exclude that the localization that isobserved reflects only the distribution and dynamic traffick-ing of GKs during steady-state conditions.
Despite the caveats mentioned above, overexpressingtransformants are excellent tools for functional analysis. InOXG4K4R transformants, overexpressing the full-length GK4gene, several morphological and developmental aberra-tions were observed including a significant increase insporangia size, the development of multiple germ tubesand increased germ tube elongation. Multiple hyphal out-growths were previously observed upon silencing Pigpb1in P. infestans, but in those transformants that lack the Gbsubunit, the sporangia were extraordinarily small and there
was no sustained tube growth (Latijnhouwers and Govers,2003). In plants phenomena reminiscent of the tube elon-gation observed in OXG4K4R transformants, have beenassociated with PIPK activity and/or production and locali-zation of PI(4,5)P2 (Munnik and Nielsen, 2011). In Agapan-thus umbellatus, for example, tube elongation is observedwhen low concentrations of caged PI(4,5)P2 are photo-released in pollen tubes (Monteiro et al., 2005) and in A.thaliana or N. benthamiana overexpression of PIP5Kinfluenced pollen tube expansion or root hair formation(Kusano et al., 2008; Stenzel et al., 2008; Zhao et al.,2010; Ischebeck et al., 2011). Moreover, in the moss Phys-comitrella patens PIPK1 was essential for rhizoid elonga-tion and caulonemal cell development (Saavedra et al.,2011). Experiments to determine whether the type IV PIPKencoded by PiGK4 has PIPK activity are in progress.
The significantly larger sporangia produced by theOXG4K4R transformants had aberrant cell walls that seemedto be thicker and showed defects in zoosporogenesis.Cleavage seemed to be incomplete as there werefewer zoospores released. Nevertheless, the zoosporesbehaved as normal and had a normal morphology (data notshown). Also OXG4R transformants that overexpress onlythe GPCR domain of PiGK4 had defects in zoosporogen-esis. However, in contrast to full-length overexpressorsthe number of released zoospores was not reduced (nodecrease in % cleavage) but the zoospores were large withaberrant morphology. This strong effect on indirect germi-nation, likely due to impairment in cytoplasmic cleavagewas also observed in gpa1-silenced transformants(Latijnhouwers et al., 2004). An additional defect in the Gamutants is reduced motility of zoospores, a phenotype thatwas not observed in any of the PiGK4-silenced or-overexpressing transformants analysed in the currentstudy. Incomplete cleavage or no cleavage at all, andresulting in multinucleated ‘superzoospores’, is also aprominent phenotype in P. infestans transformants overex-pressing PiCdc14, a gene with a putative function inflagella and that encodes a phosphatase found to bindmicrotubules in vitro (Ah-Fong and Judelson, 2011).Despite the fact that the various transformants all showdefects in cleavage during zoosporogenesis, the causesprobably differ. It is likely that in one way or the other thecytoskeleton is involved as suggested by the interaction ofPiCdc14 with microtubules (Ah-Fong and Judelson, 2011),and as might be the case in the PiGK4 full-length overex-pressors in which potentially higher levels of PIP2 mayaffect the actin cytoskeleton (Clarke et al., 2007). Thisdoes not explain why cleavage is affected in the OXG4R
transformants in which the transgene lacks the PIPK. Inthose transformants it is possible that the overload of theGPCR domain results in sequestering of a ligand, therebydisturbing the normal signal transduction routes and result-ing in the aberrant phenotypes.
Phytophthora GK4 in sporangia development and virulence 15
A disturbed balance caused by overload of the GPCRdomain might also explain why the OXG4R transformantsand the silenced transformants (SP4R1, SP4R2, SG4K4R1 andSG4K4R2) showed similar phenotypes in the pathogenicityassay on potato. In the case of silencing, there is less orno PiGK4 activity, resulting in loss of downstream signalsnecessary to support pathogenicity. In the case of theOXG4R transformants, the ligand might be fully absorbedby the truncated PiGK4 thereby preventing the activationof the native GPCR-PIPK and thus mimicking the silenc-ing effect. On N. benthamiana also the full-length overex-pressors showed reduced pathogenicity. This might bedue to the fact that N. benthamiana is less sensitive toP. infestans than potato, so that slightly impaired patho-genicity is better noticed. For other organisms like yeast,plants and mammals, it was demonstrated that phospho-inositide signalling is dependent on a delicate balance(Fukami et al., 2010) emphasizing that strict regulation ofGK activity is likely required to efficiently regulate thepathogens’ behaviour.
The data presented here suggest that GKs in oomyce-tes have properties and functions which are distinctivefrom those of individual GPCRs and PIPKs. Based on thelocalization of the GKs in vesicular compartments and thepredicted topology, it seems that the cargo of the vesiclescontains the ligand for the GPCR and thereby mayinfluence the PIPK activity in the cytoplasm. Phospho-inositides play a crucial role in vesicle sorting (Behnia andMunro, 2005; Balla et al., 2009). Therefore, modulation ofPIPK activity could influence the phosphoinositide‘barcode’ of the vesicle and as such recode it for (addi-tional) intracellular transport and subsequent function.The phenotypic changes observed upon silencing andoverexpression of GK genes in P. infestans in this studyand in P. sojae by Yang et al. (2013, accompanyingpaper) imply that GKs are involved in various aspects oflife cycle management. GPCRs are currently the mostimportant and successful target proteins for drug devel-opment (Schlyer and Horuk, 2006; Xue et al., 2008) high-lighting oomycete GKs as interesting tools to support thesearch for new compounds that target aggressive oomy-cete plant pathogens such as Phytophthora.
Experimental procedures
Gene searches and analysis
All PIPK genes previously identified in P. sojae and P. ramo-rum (Meijer and Govers, 2006) were used to searchfor homologues in the genomes of P. infestans (http://www.broadinstitute.org/annotation/genome/phytophthora_infestans/MultiHome.html), H. arabidopsidis (http://vmd.vbi.vt.edu/) and Py. ultimum (http://pythium.plantbiology.msu.edu/) by various BLAST methods. ESTs, when available, wereused to confirm intron–exon structures. All sequences
obtained are incorporated in Table S1. The PIPKs werealigned by CLUSTAL_X 1.83 (Thompson et al., 1997) and theconserved domains were analysed by comparison with non-redundant protein sequences at the NCBI website (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Phylogenetic trees wereconstructed by Mega 4 (Tamura et al., 2007). Transmembranedomains and membrane topology were predicted usingConpred II (http://bioinfo.si.hirosaki-u.ac.jp/~ConPred2/) andHMMTOP (http://www.enzim.hu/hmmtop/). For in silicoexpression analysis of PIPK genes we used P. infestans Nim-bleGen microarray data (Accession No. GSE14480; Haaset al., 2009) and EST data (Randall et al., 2005) and P. sojaeRNA-seq data derived from http://phy.njau.edu.cn/ptd/ (Yeet al., 2011).
Phytophthora culture conditions
Wild-type P. infestans isolate 88069 and all transformantswere maintained at 18°C on rye sucrose ager (RSA) (Catenand Jinks, 1968), or on clarified rye broth prepared by cen-trifuging rye sucrose medium (RSM) for 10 min at 5000 g(Judelson and Roberts, 2002). Non-sporulating hyphaewere obtained from 5-day-old RSM cultures inoculated withmycelia derived from RSA plates. Sporangia were obtainedby flooding 12-day-old RSA cultures with water, rubbing thehyphae with a glass rod, followed by filtration through twolayers of Miracloth (CALBIOCHEM, Cat. No. 475855).Zoospores were generated by incubation of plates or iso-lated sporangia in cold water for 3 h at 4°C. Zoosporeswere purified by filtration through two layers of Miraclothfollowed by centrifugation at 2000 g for 2 min to collect thezoospores and encysting zoospores. Cysts were generatedby vortexing zoospore suspensions for 30 s. Germinatingcysts were obtained by incubating the cysts at room tem-perature for 2 h in water, followed by a centrifugation step at2000 g for 2 min. Tissues of P. sojae from different devel-opmental stages were obtained as described by Hua et al.(2008). P. sojae tissues of strain P6497 were grown andgerminated in V8 liquid medium. Since P. sojae sporangiaare non-caducous, sporulating hyphae were prepared byrepeatedly washing 3-day-old hyphae with water and incu-bating them for 4–8 h at 25°C until most hyphae developedsporangia. P. sojae zoospores were filtered through twolayers of Miracloth and collected by centrifugation at 2000 gfor 2 min. Tissues to be used for RNA isolation were con-centrated by centrifugation, frozen directly into liquid N2 andstored at -80°C.
Nucleic acid manipulations
DNA and RNA isolation from Phytophthora and expressionanalysis by RT-PCR were performed as described previously(Pieterse et al., 1991; Hua et al., 2008). For first-strand cDNAsynthesis 2 mg of total RNA was used as template. qRT-PCRanalysis was performed following the method of Zhao et al.(2011). The ActA gene from P. infestans was used as a con-stitutively expressed endogenous control, and the expressionof each gene was determined relative to ActA using the DDCtmethod. The primers are listed in Table S4. All qRT-PCRswere performed at least three times.
For constructing plasmids for transformation genomic DNA ofmycelium and cDNA from different tissues of P. infestansstrain 88069 were used as templates for amplification of genesand gene fragments. Amplification of coding sequences wasperformed by Pfu proofreading polymerase (Promega) withspecific primers (Table S4) linked to required restriction sites.Upon amplification, the obtained fragments were gel-purifiedand cloned into pGEM-T-Easy vectors (Promega). All frag-ments were confirmed by sequencing using standard M13primers or specific primers (Table S4). The fragments werere-cloned into the ClaI and BsiWI restriction sites of thepTORmRFP4 vector (Fig. S2). Transformation to P. infestansstrain 88069 was performed as described previously (McLeodet al., 2008). Putative transformants were screened by PCRusing primer RFP-F-seq, matching the HAM34 promoter, andRFP-R-seq, matching the mRFP gene. PCR positive trans-formants were analysed for silencing and overexpressing ofPiGK4 by RT-PCR and this was confirmed and quantified byqRT-PCR.
Fluorescence microscopy
Sporangia of transformants with mRFP reporter constructswere germinated overnight in RS liquid medium on biofoilslides, and slides were examined by microscopy. For lightand epifluorescence microscopy we used a Nikon Eclipse90i epifluorescence microscope (Nikon, Badhoevedorp, theNetherlands). mRFP fluorescence (561 nm excitation; emis-sion 600–630 nm) was visualized by using a TRITC filtercube (EX540/25, DM 565, BA 605/55). The NIS-Elementssoftware package and LSM image examiner software wereused to analyse and merge digital pictures. For more detailedstudies, Confocal Laser Scanning Microscopy (CLSM) wasapplied using a 60¥ oil-immersion objective. Excitation/emission settings for mRFP were 543/LP560 nm.
Germination assay
Sporangia or cysts were incubated in Petri dishes or in a24-well microtitre plates in water at 18°C for the time pointsindicated in the text or figure legends. Germination was moni-tored by light microscopy. In case of multiple germ tubes, asfrequently observed in OXG4R transformants, the most pro-longed was considered for measurement. All experimentswere performed at least three times.
Topology construct cloning and expression in E. coli
The bacterial expression vector pbb3 for b-lactamase fusionsis shown in Fig. S3 and was constructed as described in thelegend of that figure. Fragments of PiGK4 encoding one ormore predicted TM segments were amplified and cloned intothe pbb3 vector via the EcoRI and NotI sites enclosed in theprimers (Table S4; Fig. S3) and verified by sequencing. Sub-sequently these were introduced into E. coli strain C41(Miroux and Walker, 1996) and transformants were grownovernight in LB amended with 50 mg ml-1 kanamycin at 28°C.The cultures were diluted to OD600 = 1 and protein production
was induced by addition of IPTG (0.5 mM). After incubation at22°C (2 h, 180 r.p.m.), a 10-fold dilution series was made inLB supplemented with 0.5 mM IPTG. Droplets of 5 ml werespotted on LB agar plates containing 0.5 mM IPTG and 0,1.25, 2.5 or 10 mg ml-1 carbenicillin and incubated at 22°C. Atleast two independent clones were tested for each construct.
Infection assay
Twenty microlitres of zoospores (2 ¥ 103) droplets from eachstrain were inoculated on detached leaves of 20-day-oldpotato cultivar Impala or 15-day-old N. benthamiana. Inocu-lated leaves were incubated at 18°C. The pictures of infectedleaves were taken at 5 dpi for potato cultivar Impala and 7 dpifor N. benthamiana. At least six detached leaves were inocu-lated for each strain. Each lesion was measured and statis-tics were performed by Student’s t-test. The experimentswere performed at least twice.
Acknowledgements
This work was financially supported by the TechnologyFoundation of the Netherlands Organization for ScientificResearch (STW-NWO; VIDI Grant 10281 to H.J.G.M.), thePriority Academic Program Development of Jiangsu HigherEducation Institutions (PAPD), and the 111 InternationalCooperation grant (B07030) from the Chinese government(PAPD to Y.W. and B07030 to Nanjing Agricultural Univer-sity). We thank Norbert de Ruijter and Klaas Bouwmeester forassistance with microscopy, and Rob Weide for assistancewith Phytophthora transformation. We thank Frank Takken forproviding C41 E. coli strain, May Bente Brurberg for providingpETM10 vector and Steve Whisson for providing the plasmidpTORmRFP4.
References
Agrios, G.N. (2005) Plant Pathology. New York, USA: Aca-demic Press.
Ah-Fong, A.M., and Judelson, H.S. (2011) New role forCdc14 phosphatase: localization to basal bodies in theoomycete Phytophthora and its evolutionary coinheritancewith eukaryotic flagella. PLoS ONE 6: e16725.
Bakthavatsalam, D., Meijer, H.J.G., Noegel, A.A., andGovers, F. (2006) Novel phosphatidylinositol phosphatekinases with a G-protein coupled receptor signature areshared by Dictyostelium and Phytophthora. Trends Micro-biol 14: 378–382.
Bakthavatsalam, D., Brazill, D., Gomer, R.H., Eichinger, L.,Rivero, F., and Noegel, A.A. (2007) A G protein-coupledreceptor with a lipid kinase domain is involved in cell-density sensing. Curr Biol 17: 892–897.
Balla, T., Szentpetery, Z., and Kim, Y.J. (2009) Phospho-inositide signaling: new tools and insights. Physiology(Bethesda) 24: 231–244.
Baxter, L., Tripathy, S., Ishaque, N., Boot, N., Cabral, A.,Kemen, E., et al. (2010) Signatures of adaptation to obli-gate biotrophy in the Hyaloperonospora arabidopsidisgenome. Science 330: 1549–1551.
Behnia, R., and Munro, S. (2005) Organelle identity and thesignposts for membrane traffic. Nature 438: 597–604.
Phytophthora GK4 in sporangia development and virulence 17
Calamia, J., and Manoil, C. (1990) lac permease ofEscherichia coli: topology and sequence elements promot-ing membrane insertion. Proc Natl Acad Sci USA 87:4937–4941.
Camacho, L., Smertenko, A.P., Perez-Gomez, J., Hussey,P.J., and Moore, I. (2009) Arabidopsis Rab-E GTPasesexhibit a novel interaction with a plasma-membranephosphatidylinositol-4-phosphate 5-kinase. J Cell Sci 122:4383–4392.
Caten, C.E., and Jinks, J.L. (1968) Spontaneous variability ofsingle isolates of Phytophthora infestans. I. Cultural varia-tion. Can J Bot 46: 329–348.
Chen, J.G., Pandey, S., Huang, J., Alonso, J.M., Ecker, J.R.,Assmann, S.M., and Jones, A.M. (2004) GCR1 can actindependently of heterotrimeric G-protein in response tobrassinosteroids and gibberellins in Arabidopsis seed ger-mination. Plant Physiol 135: 907–915.
Clarke, J.H., Richardson, J.P., Hinchliffe, K.A., and Irvine,R.F. (2007) Type II PtdInsP kinases: location, regulationand function. Biochem Soc Symp (74): 149–159.
Ferguson, S.S. (2001) Evolving concepts in G protein-coupled receptor endocytosis: the role in receptor desen-sitization and signaling. Pharmacol Rev 53: 1–24.
Fukami, K., Inanobe, S., Kanemaru, K., and Nakamura, Y.(2010) Phospholipase C is a key enzyme regulating intra-cellular calcium and modulating the phosphoinositidebalance. Prog Lipid Res 49: 429–437.
Haas, B.J., Kamoun, S., Zody, M.C., Jiang, R.H., Handsaker,R.E., Cano, L.M., et al. (2009) Genome sequence andanalysis of the Irish potato famine pathogen Phytophthorainfestans. Nature 461: 393–398.
Heuss, C., and Gerber, U. (2000) G-protein-independent sig-naling by G-protein-coupled receptors. Trends Neurosci23: 469–475.
Hislop, J.N., and von Zastrow, M. (2011) Role of ubiquitina-tion in endocytic trafficking of G-protein-coupled receptors.Traffic 12: 137–148.
Hua, C., Wang, Y., Zheng, X., Dou, D., Zhang, Z., Govers, F.,and Wang, Y. (2008) A Phytophthora sojae G-protein alphasubunit is involved in chemotaxis to soybean isoflavones.Eukaryot Cell 7: 2133–2140.
Ischebeck, T., Stenzel, I., and Heilmann, I. (2008) Type Bphosphatidylinositol-4-phosphate 5-kinases mediate Arabi-dopsis and Nicotiana tabacum pollen tube growth by regu-lating apical pectin secretion. Plant Cell 20: 3312–3330.
Ischebeck, T., Stenzel, I., Hempel, F., Jin, X., Mosblech, A.,and Heilmann, I. (2011) Phosphatidylinositol-4,5-bisphosphate influences Nt-Rac5-mediated cell expansionin pollen tubes of Nicotiana tabacum. Plant J 65: 453–468.
Judelson, H.S., and Blanco, F.A. (2005) The spores of Phy-tophthora: weapons of the plant destroyer. Nat Rev Micro-biol 3: 47–58.
Judelson, H.S., and Roberts, S. (2002) Novel protein kinaseinduced during sporangial cleavage in the oomycete Phy-tophthora infestans. Eukaryot Cell 1: 687–695.
Kusano, H., Testerink, C., Vermeer, J.E.M., Tsuge, T.,Shimada, H., Oka, A., et al. (2008) The Arabidopsis phos-
phatidylinositol phosphate 5-kinase PIP5K3 is a key regu-lator of root hair tip growth. Plant Cell 20: 367–380.
Lacatena, R.M., Cellini, A., Scavizzi, F., and Tocchini-Valentini, G.P. (1994) Topological analysis of the humanbeta 2-adrenergic receptor expressed in Escherichia coli.Proc Natl Acad Sci USA 91: 10521–10525.
Lamour, K.H., Finley, L., Hurtado-Gonzales, O., Gobena, D.,Tierney, M., and Meijer, H.J.G. (2006) Targeted gene muta-tion in Phytophthora spp. Mol Plant Microbe Interact 19:1359–1367.
Latijnhouwers, M., and Govers, F. (2003) A Phytophthorainfestans G-protein beta subunit is involved in sporangiumformation. Eukaryot Cell 2: 971–977.
Latijnhouwers, M., Munnik, T., and Govers, F. (2002) Phos-pholipase D in Phytophthora infestans and its role inzoospore encystment. Mol Plant Microbe Interact 15: 939–946.
Latijnhouwers, M., de Wit, P.J., and Govers, F. (2003)Oomycetes and fungi: similar weaponry to attack plants.Trends Microbiol 11: 462–469.
Latijnhouwers, M., Ligterink, W., Vleeshouwers, V.G., vanWest, P., and Govers, F. (2004) A Galpha subunit controlszoospore motility and virulence in the potato late blightpathogen Phytophthora infestans. Mol Microbiol 51: 925–936.
Laxalt, M.A., and Govers, F. (2002) Differential expression ofG protein alpha and beta subunit genes during develop-ment of Phytophthora infestans. Fungal Genet Biol 36:137–146.
Levesque, C.A., Brouwer, H., Cano, L., Hamilton, J.P., Holt,C., Huitema, E., et al. (2010) Genome sequence of thenecrotrophic plant pathogen Pythium ultimum reveals origi-nal pathogenicity mechanisms and effector repertoire.Genome Biol 11: R73.
Li, L., Wright, S.J., Krystofova, S., Park, G., and Borkovich,K.A. (2007) Heterotrimeric G protein signaling in filamen-tous fungi. Annu Rev Microbiol 61: 423–452.
McLeod, A., Fry, B.A., Zuluaga, A.P., Myers, K.L., and Fry,W.E. (2008) Toward improvements of oomycete transfor-mation protocols. J Eukaryot Microbiol 55: 103–109.
Marchese, A., Paing, M.M., Temple, B.R., and Trejo, J. (2008)G protein-coupled receptor sorting to endosomes and lyso-somes. Annu Rev Pharmacol Toxicol 48: 601–629.
Meijer, H.J.G., and Govers, F. (2006) Genomewide analysisof phospholipid signaling genes in Phytophthora spp.: nov-elties and a missing link. Mol Plant Microbe Interact 19:1337–1347.
Meijer, H.J.G., and Munnik, T. (2003) Phospholipid-basedsignaling in plants. Annu Rev Plant Biol 54: 265–306.
Meijer, H.J.G., Hassen, H.H., and Govers, F. (2011) Phytoph-thora infestans has a plethora of phospholipase Denzymes including a subclass that has extracellular activ-ity. PLoS ONE 6: e17767.
Mikami, K., Saavedra, L., Hiwatashi, Y., Uji, T., Hasebe, M.,and Sommarin, M. (2010) A dibasic amino acid pair con-served in the activation loop directs plasma membranelocalization and is necessary for activity of plant type I/IIphosphatidylinositol phosphate kinase. Plant Physiol 153:1004–1015.
Miroux, B., and Walker, J.E. (1996) Over-production of pro-teins in Escherichia coli: mutant hosts that allow synthesis
of some membrane proteins and globular proteins at highlevels. J Mol Biol 260: 289–298.
Monteiro, D., Castanho Coelho, P., Rodrigues, C., Camacho,L., Quader, H., and Malho, R. (2005) Modulation of endo-cytosis in pollen tube growth by phosphoinositides andphospholipids. Protoplasma 226: 31–38.
Munnik, T., and Nielsen, E. (2011) Green light for polyphos-phoinositide signals in plants. Curr Opin Plant Biol 14:489–497.
Nilsson, I., and von Heijne, G. (1990) Fine-tuning the topol-ogy of a polytopic membrane protein: role of positively andnegatively charged amino acids. Cell 62: 1135–1141.
Oude Weernink, P.A., Han, L., Jakobs, K.H., and Schmidt,M. (2007) Dynamic phospholipid signaling by G protein-coupled receptors. Biochim Biophys Acta 1768: 888–900.
Petäjä-Repo, U.E., Bouvier, M., and Devi, L.A. (2005) GPCRfolding and maturation. In Contemporary Clinical Neuro-science: The G Protein-Coupled Receptors Handbook.Devi, L.A. (ed.). Totowa, USA: Humana Press, pp. 71–93.
Randall, T.A., Dwyer, R.A., Huitema, E., Beyer, K., Cvitanich,C., Kelkar, H., et al. (2005) Large-scale gene discovery inthe oomycete Phytophthora infestans reveals likely com-ponents of phytopathogenicity shared with true fungi. MolPlant Microbe Interact 18: 229–243.
Riyahi, T.Y., Frese, F., Steinert, M., Omosigho, N.N., Glockner,G., Eichinger, L., et al. (2011) RpkA, a highly conservedGPCR with a lipid kinase domain, has a role in phagocytosisand anti-bacterial defense. PLoS ONE 6: e27311.
Saavedra, L., Balbi, V., Lerche, J., Mikami, K., Heilmann, I.,and Sommarin, M. (2011) PIPKs are essential for rhizoidelongation and caulonemal cell development in the mossPhyscomitrella patens. Plant J 67: 635–647.
Schlyer, S., and Horuk, R. (2006) I want a new drug:G-protein-coupled receptors in drug development. DrugDiscov Today 11: 481–493.
Schulein, R., Rutz, C., and Rosenthal, W. (1996) Membranetargeting and determination of transmembrane topology ofthe human vasopressin V2 receptor. J Biol Chem 271:28844–28852.
Seidl, M.F., Van den Ackerveken, G., Govers, F., and Snel, B.(2011) A domain-centric analysis of oomycete plant patho-gen genomes reveals unique protein organization. PlantPhysiol 155: 628–644.
Seidl, M.F., Van den Ackerveken, G., Govers, F., and Snel, B.(2012) Reconstruction of oomycete genome evolutionidentifies differences in evolutionary trajectories leading topresent-day large gene families. Genome Biol Evol 4: 199–211.
Shaw, B.D., Chung, D.W., Wang, C.L., Quintanilla, L.A., andUpadhyay, S. (2011) A role for endocytic recycling inhyphal growth. Fungal Biol 115: 541–546.
Stenzel, I., Ischebeck, T., Konig, S., Holubowska, A.,
Sporysz, M., Hause, B., and Heilmann, I. (2008) The typeB phosphatidylinositol-4-phosphate 5-kinase 3 is essentialfor root hair formation in Arabidopsis thaliana. Plant Cell20: 124–141.
Tamura, K., Dudley, J., Nei, M., and Kumar, S. (2007) MEGA4:Molecular Evolutionary Genetics Analysis (MEGA) softwareversion 4.0. Mol Biol Evol 24: 1596–1599.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F.,and Higgins, D.G. (1997) The CLUSTAL_X windows inter-face: flexible strategies for multiple sequence alignmentaided by quality analysis tools. Nucleic Acids Res 25:4876–4882.
Toshima, J.Y., Nakanishi, J., Mizuno, K., Toshima, J., andDrubin, D.G. (2009) Requirements for recruitment of a Gprotein-coupled receptor to clathrin-coated pits in buddingyeast. Mol Biol Cell 20: 5039–5050.
Tyler, B.M., Tripathy, S., Zhang, X., Dehal, P., Jiang, R.H.,Aerts, A., et al. (2006) Phytophthora genome sequencesuncover evolutionary origins and mechanisms of patho-genesis. Science 313: 1261–1266.
Wang, Y., Li, A., Wang, X., Zhang, X., Zhao, W., Dou, D.,Zheng, X., and Wang, Y. (2010) GPR11, a putative seven-transmembrane G protein-coupled receptor, controlszoospore development and virulence of Phytophthorasojae. Eukaryot Cell 9: 242–250.
Whisson, S.C., Boevink, P.C., Moleleki, L., Avrova, A.O.,Morales, J.G., Gilroy, E.M., et al. (2007) A translocationsignal for delivery of oomycete effector proteins into hostplant cells. Nature 450: 115–118.
Xue, C., Hsueh, Y.P., and Heitman, J. (2008) Magnificentseven: roles of G protein-coupled receptors in extracellularsensing in fungi. FEMS Microbiol Rev 32: 1010–1032.
Yang, X., Zhao, W., Hua, C., Zheng, X., Jing, M., Li, D., et al.(2013) Chemotaxis and oospore formation in Phytophthorasojae are controlled by G-protein-coupled receptors with aphosphatidylinositol phosphate kinase domain. Mol Micro-biol doi:10.1111/mmi.12191
Ye, W., Wang, X., Tao, K., Lu, Y., Dai, T., Dong, S., et al.(2011) Digital gene expression profiling of the Phytoph-thora sojae transcriptome. Mol Plant Microbe Interact 12:1530–1539.
Zhao, W., Dong, S., Ye, W., Hua, C., Meijer, H.J.G., Dou, X.,et al. (2011) Genome-wide identification of Phytophthorasojae SNARE genes and functional characterization of theconserved SNARE PsYKT6. Fungal Genet Biol 48: 241–251.
Zhao, Y., Yan, A., Feijo, J.A., Furutani, M., Takenawa, T.,Hwang, I., et al. (2010) Phosphoinositides regulateclathrin-dependent endocytosis at the tip of pollen tubes inArabidopsis and tobacco. Plant Cell 22: 4031–4044.
Supporting information
Additional supporting information may be found in the onlineversion of this article at the publisher’s web-site.
Phytophthora GK4 in sporangia development and virulence 19