Page 1
Author's Accepted Manuscript
Glutamate level in anterior cingulate predictsanxiety in healthy humans: A magneticresonance spectroscopy study
Shilpi Modi, Poonam Rana, Prabhjot Kaur,Nisha Rani, Subash Khushu
PII: S0925-4927(14)00056-0DOI: http://dx.doi.org/10.1016/j.pscychresns.2014.03.001Reference: PSYN10178
To appear in: Psychiatry Research: Neuroimaging
Received date: 7 August 2013Revised date: 28 January 2014Accepted date: 7 March 2014
Cite this article as: Shilpi Modi, Poonam Rana, Prabhjot Kaur, Nisha Rani,Subash Khushu, Glutamate level in anterior cingulate predicts anxiety inhealthy humans: A magnetic resonance spectroscopy study, PsychiatryResearch: Neuroimaging, http://dx.doi.org/10.1016/j.pscychresns.2014.03.001
This is a PDF file of an unedited manuscript that has been accepted forpublication. As a service to our customers we are providing this early version ofthe manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting galley proof before it is published in its final citable form.Please note that during the production process errors may be discovered whichcould affect the content, and all legal disclaimers that apply to the journalpertain.
www.elsevier.com/locate/psychresns
Page 2
Glutamate level in anterior cingulate predicts anxiety in healthy humans: A magnetic
resonance spectroscopy study
Shilpi Modi, Poonam Rana, Prabhjot Kaur, Nisha Rani, and Subash Khushu*
NMR Research Centre, Institute of Nuclear Medicine and Allied Sciences (INMAS), Lucknow Road, Timarpur, Delhi, India
*Corresponding author.
Dr. Subash Khushu,
Scientist ‘G’ and Head,
NMR Research Center, INMAS,
Lucknow Road, Timarpur,
Delhi -110054, India.
Tel.: +91-11-23905313; Fax: +91-11-23919509
E-mail address: [email protected]
Page 3
Glutamate level in anterior cingulate cortex predicts anxiety in healthy humans: A
magnetic resonance spectroscopy study
Abstract
Anxiety, a personality dimension in healthy humans, has been found to be associated with
many functional consequences such as increased distractibility and attentional bias in favour
of threat-related information, along with morphological and microstructural changes in the
brain. The associated metabolic/ neurochemical alterations are sparsely studied. In the present
magnetic resonance spectroscopy (MRS) study, we investigated the possible relationship
between regional brain chemistry within anterior cingulate cortex (4-cm3 voxel) and
hippocampus (2.5-cm3 voxel) and anxiety (measured by State-Trait Anxiety Inventory) in our
subject group. In the anterior cingulate cortex, multivariate analysis of covariance showed an
increase in myo-inositol and combined glutamate and glutamine levels in the high anxiety
subject group as compared with the low anxiety group. In the partial correlation analysis
between neurochemicals and anxiety, glutamate and combined glutamate and glutamine also
showed a predictive value for anxiety. On analysing the trait anxiety sub-score separately, we
found glutamate, inositol and combined glutamate and glutamine levels to be increased in the
high trait anxiety group as compared with the low trait anxiety group. All three resonances
also had a predictive value for trait anxiety. In the hippocampus, none of the neurochemicals
showed significant difference between high and low anxiety groups. The study provides a
first account of alterations in anterior cingulate cortex neurochemistry in relation to anxiety in
healthy subjects. The study thus contributes to the limited literature available on altered
metabolism and neural mechanisms underlying sub-clinical anxiety.
Keywords: Trait anxiety; State anxiety; Neuroimaging; Hippocampus
Page 4
1. Introduction
Anxiety is an aversive emotional and motivational state that can be characterized by harm
avoidance behaviour such as worrying, irritability, and difficulty in relaxing, or by a
predisposition to interpret ambiguous situations as threatening (Grachev and Apkarian, 2000;
Eysenck et al., 2007). These effects are seen not only in clinical anxiety but also within the
normal population as a personality dimension that is generally assessed by measures of
anxiety such as Spielberger’s State-Trait Anxiety Inventory (STAI) (Spielberger, 1983;
Eysenck et al., 2007). The STAI consists of two scales – Y2, which measures the
participant’s disposition to anxiety (trait), and Y1, which measures how anxious the
participant feels at the moment (state). The model proposed by Sandi and Richter-Levin
(Sandi and Richter-Levin, 2009) supports the etiological hypotheses that the high anxiety trait
is a vulnerability factor to develop both depression and anxiety disorders. Therefore,
identifying hallmarks of anxiety to identify individuals that are at risk for the development of
clinical anxiety disorders and depression becomes important, so that timely preventive
interventions may be given to them. Both state and trait anxiety have been found to be
associated with many functional consequences such as, increased distractibility, attentional
bias in favour of threat-related information and a hyper-responsive amygdala even for
unattended threat-related stimuli (Bishop et al., 2004; Eysenck et al., 2007; Bishop, 2009).
Similarly, morphological and microstructural changes have been reported to be associated
with anxiety-related personality traits (Pujol et al., 2002; Westlye et al., 2011; Baur et al.,
2012; Lochner et al., 2012; Montag et al., 2012; Modi et al., 2013). Pujol et al. (2002)
showed that a large right anterior cingulate cortex is related to a temperamental disposition to
fear and anticipatory worry. Similarly, in another volumetric study, a positive correlation was
obtained between trait anxiety and the volume of the left amygdala and right hippocampus
(Baur et al., 2012). Lochner et al. (2012) found white matter abnormalities in the anterior
Page 5
limb of the internal capsule (ALIC) and cingulum that were related to obsessive-compulsive
disorder (OCD). Similarly, increased harm avoidance behaviour has been shown to be
associated with decreased white matter (WM) microstructure in major WM tracts such as
corticolimbic pathways, the anterior thalamic radiations, inferior fronto-occipital cortex and
right superior longitudinal fasciculus (Westlye et al., 2011). Individual differences in negative
emotionality have also been found to be positively correlated with the fractional anisotropy
(FA) values of several WM tracts in the temporal lobe of the left hemisphere (Montag et al.,
2012). A recent diffusion tensor tractography study showed a positive correlation between
trait anxiety scores and the mean FA value in the fornix and left uncinate fasciculus (Modi et
al., 2013).
The functional, morphological and microstuctural changes associated with anxiety might also
be related to alterations in regional chemistry of the brain. In vivo proton magnetic resonance
spectroscopy (1H-MRS) has been used primarily to measure concentrations of
neurochemicals in brain tissue such as N-acetyl-l-aspartate (NAA), which is a marker of
neuronal viability (Miller, 1991); glutamate (Glu) and glutamine (Gln), which are excitatory
neurotransmitters (Whiteside et al., 2006); choline (Cho), which is an indirect marker of
myelination and cell membrane metabolism (Miller, 1991); myo-inositol (mI), which is
involved in phospholipid metabolism (Whiteside et al., 2006); and creatine (Cr), which is a
marker of cellular energetics and commonly used as a reference level (Miller, 1991;
Whiteside et al., 2006). In vivo MRS studies on the neurobiology of anxiety-related disorders
are relatively few and confined to DSM-IV anxiety disorders like panic disorder (PD) (Dager
et al., 1999), OCD (Whiteside et al., 2006), posttraumatic stress disorder (PTSD)
(Mahmutyaz�c�o�lu et al., 2005; Kimbrell et al., 2005), and social anxiety disorder (SAD)
(Phan et al., 2005). MRS studies in non-clinical populations in which trait or state anxiety is
assessed as a personality dimension are very few. The first ever MRS study examining the
Page 6
relationship between regional brain chemistry within the orbital frontal cortices and
physiologic anxiety in normal subjects demonstrated an increase in chemical concentrations
of most of the metabolites studied in the group with higher levels of anxiety as compared
with the lower anxiety group. Out of all the metabolites studied, NAA was identified as the
strongest chemical marker for anxiety (Grachev and Apkarian, 2000). The authors explained
these findings on the basis of a chemical-behavioral network in the brain of healthy subjects
as a possible mechanism for the development of anxiety. They proposed that the genetic
predisposition for anxiety may be linked to the number of neurons and their receptors in the
brain. An increase in the number of axons and synaptic connections across anxiety-related
regions (i.e., sprouting) might result in an increase in the concentration of NAA (Grachev and
Apkarian, 2000). In another study on unmedicated OCD patients, Fan et al. (2010) also
reported a correlation between the NAA/Cr ratio in the medial prefrontal cortex and trait
anxiety scores on the STAI among the control group.
Besides the orbital frontal cortex, the anterior cingulate cortex (ACC) and the hippocampus
have been previously implicated in the pathophysiology of several psychiatric disorders, like
bipolar disorder, depression, and schizophrenia, as well as anxiety disorders such as PTSD,
OCD, and SAD (Heckers et al., 2001; Sala et al., 2004; Mahmutyaz�c�o�lu et al., 2005; Phan
et al., 2005; Whiteside et al., 2006; Hong et al., 2007; Hermens et al., 2012). The anterior
cingulate cortex plays a role in the modulation of conditional fear responses and is critically
involved in performance monitoring and cognitive control. It also has a key role in emotional
and social behaviour (Mahmutyaz�c�o�lu et al., 2005; Etkin et al., 2011; Hermens et al.,
2012). The hippocampus is also one of the most important structures of the limbic system,
being involved in memory functions, processing of emotional information, and stress
response via the hypothalamo-pituitary-adrenal axis (Mahmutyaz�c�o�lu et al., 2005). In the
Page 7
study by Grachev and Apkarian (2000), the authors did not find any relation between
neurochemistry in the cingulate cortex and anxiety. They attributed this negative finding to
the fact that only the middle portion of cingulate area was studied. Instead, the anterior
portion is considered to be an affective area and implied in a variety of emotional tests such
as emotional conflict regulation (Egner et al., 2008), reappraisal (Etkin et al., 2011), affect
labeling of emotional faces (Liebermen et al., 2007), and self-distraction from a fear-
conditioned stimulus (Delgado et al., 2008). Therefore, in the present study, we examined the
relationship between regional brain neurochemicals in the anterior cingulate cortex and the
anxiety levels of healthy subjects using in vivo 1H-MRS. In addition, because of the
relationship between the hippocampus and anxiety, the hippocampus was chosen as another
region of interest (ROI) for MRS. Spectroscopic findings in the anterior cingulate cortex and
the hippocampus in relation to anxiety levels of healthy subjects have the potential to
improve our understanding of the neurobiology of sub-clinical anxiety. Given the limited
literature available on the relationship between anxiety in healthy populations and regional
brain chemistry, we did not restrict our analyses to a particular neurochemical; rather, we
analysed all the possible metabolites.
2. Methods
2.1. Subjects
Twenty-four right-handed, healthy, educated participants drawn from the institute (males =
11, females = 13, mean age = 23.16 years, SD = 2.20 years) were screened for current or past
medical and psychiatric illness using the Medical History Questionnaire of the Hindi version
of the Diagnostic Interview for Genetic Studies (version 2) (Deshpande et al., 1998). None of
the subjects recruited for the study had any clinical evidence of stroke, head injury,
cardiovascular diseases, history of alcohol or drug dependence, hypertension, neurological or
psychiatric disorder or sensori-cognitive impairment; nor did they have any cortical
Page 8
infarctions on the T2-weighted magnetic resonance images. In the case of females, care was
taken that while undergoing spectroscopy session they were not in their premenstrual or
menstrual periods. The procedure followed in the current study was in accordance with the
guidelines of the ethical committee of the Institute. Further, all subjects gave their consent to
participate in the study, and the procedure was thoroughly explained to them. After the
spectroscopy session, participants’ state-anxiety (inside the scanner) and trait-anxiety levels
were assessed using the STAI self-report questionnaires for adults (Spielberger, 1983).
Following Grachev and Apkarian (2000), the scores of state and trait anxiety were added
together to obtain the total anxiety scores of the participants. To evaluate the current
depression levels, participants were also asked to complete the Beck Depression Inventory
(BDI) (Beck et al., 1996).
2.2. Localized invivo single voxel 1H-MRS
The study was carried out using a 3-T whole body MR system (Magnetom Skyra, Siemens,
Germany) with a 32-channel head coil. The subject’s head was immobilized using
expandable ear cushions. Anatomical imaging was performed in all three orthogonal planes
for positioning the MRS voxels. T2-weighted multislice images (repetition time (TR) = 5600
ms, echo time (TE) = 100 ms, number of excitations = 1, 312×512 matrix, field of view =
180×220 mm, 25 slices, slice thickness = 4.0 mm, distance factor = 1.2 mm) covering the
entire brain were obtained.
For the anterior cingulate cortex, a 20 × 20 × 10 mm3 voxel was positioned in the midline
(covering samples from both the right and left sides of the brain) on the axial slice in which
the caudate nucleus was well formed visually. The voxel posteriorly abuts the genu of the
corpus callosum. For the hippocampus, axial oblique sections were obtained by planning
Page 9
slices parallel to the body of the hippocampus in a parasagittal section; coronal oblique
sections were obtained by positioning slices perpendicular to the body of the hippocampus. A
voxel of 25 × 10 × 10 mm3 was then positioned on these axial oblique, coronal oblique, and
sagittal sections for better delineation and coverage of the hippocampus. Representative voxel
placement are shown in Fig. 1a and Fig. 1b.
MRS was obtained using a PRESS (point-resolved spectroscopy) sequence with the
following acquisition parameters: TR/TE = 2000 ms/33 ms; 2048 spectral points; 1200-Hz
spectral bandwidth and 196 averages. Automated global shimming was used to minimize the
B0 inhomogeneties, and localized shimming was done to further minimize B0 field variations
over the voxel of interest. Unsuppressed water (with 10 averages) spectra were also acquired
immediately before the water-suppressed metabolic acquisition and was used for spectral
quantifications.
The spectra were processed using LCModel software (Provencher, 1993). Only the
metabolites/neurochemicals with Cramer-Rao Lower Bounds less than 20% were analysed.
Relative concentrations of Glu, mI, NAA, combined peak of glycerophosphocholine and
choline (tCh = total choline), combined peak of NAA and N-acetylaspartyl-glutamate
(NAA+NAAG), and combined peak of glutamate and glutamine (Glu+Gln) relative to total
creatine (tCr, creatine + phosphocreatine) were measured. The tCr values were used as
reference because the total amount of Cr is a measure of general brain metabolism and
appears to be stable in subjects who are naïve to psychotropic medications (Keshavan et al.,
1991). Representative spectra from the anterior cingulate cortex and the hippocampus are
shown in Fig. 2a and Fig. 2b, respectively.
Page 10
2.3. Statistical analysis
Controlling for the effects of age, sex, sub-clinical depression (BDI scores) and spectral
quality, anxiety group (high anxiety (total STAI score = 70–95) vs. lower anxiety (total STAI
score = 40–69)) and metabolite ratios were analyzed in both the ROIs with multivariate
analysis of covariance (MANCOVA) using the general linear model (SPSS (version 15.0,
SPSS Inc., Chicago, IL, USA) statistical software). To control for the effect of age, sex, sub-
clinical depression and spectral quality on the observed spectral differences between the two
groups, age, sex, BDI scores, full width at half maximum (FWHM) and signal-to-noise ratio
(SNR) of the spectra were taken as covariates of no interest. Adjustment for multiple
comparisons was done by applying Bonferroni correction. To determine the predictive value
of anxiety on neurochemical concentration, a partial correlation analysis (one-tailed)
including age, sex, sub-clinical depression and spectral quality (FWHM and SNR) as
covariates of no interest was computed between total anxiety scores and the neurochemical
ratios in both the ROIs, with the assumption that there was no correlation between
neurochemical ratios and anxiety (Ho=0). Alternatively, if a correlation of greater than 0.001
is observed at �=0.05 and 90% power of the test, the null hypothesis was rejected. P values of
� 0.05 were considered to be significant. A separate analysis for the trait anxiety sub-score of
the STAI was also carried out.
3. Results
3.1. Behavioral data
Descriptive statistics for the self-report measures are as follows: state anxiety score (STAI-
Y1) = 35.21 ± 9.16 (range 21-48), trait-anxiety score (STAI-Y2) = 37.21 ± 10.07 (range 20-
52), STAI total anxiety = 72.42 ± 16.42, BDI = 6.17 ± 4.51 (range 0-13). Subjects with total
STAI scores between 40 and 69 were considered as the low anxiety group (in ACC: males =
Page 11
6, females = 5, mean age = 23.91 years, SD = 2.34 years; in hippocampus: males = 6, females
= 4, mean age = 23.50 years, SD = 2.01 years), and those with total STAI scores above 70
were considered as the high anxiety group (in ACC: males = 5, females = 8, mean age =
22.54 years, SD = 1.94 years; in hippocampus: males = 3, females = 8, mean age = 22.27
years, SD = 1.95 years) (Grachev and Apkarian, 2000). There was no significant difference
between the BDI scores of the two groups (high anxiety group: BDI score = 6.62 + 5.04; low
anxiety group: BDI score = 5.64 + 3.96; p = 0.607). For the analysis taking trait anxiety sub-
score separately, subjects with trait anxiety scores between 20 and 35 were considered as the
low trait anxiety group (in ACC: males = 7, females = 4; in hippocampus: males = 6, females
= 3), and those with trait anxiety scores above 35 were considered as the high trait anxiety
group (median split on the basis of trait anxiety scores) (in ACC: males = 4, females = 9; in
hippocampus: males = 3, females = 9).
3.2. Between-group differences in neurochemical variables
There was no significant group difference in tCr concentration (institutional units) in any of
the ROIs as obtained using the LCModel (ACC, p = 0.89 (high anxiety group, tCr = 5.36 +
0.38, n = 13; low anxiety group, tCr = 5.33 + 0.44, n = 10); hippocampus, p = 0.36 (high
anxiety group, tCr = 5.82 + 0.50, n = 9; low anxiety group, tCr = 6.02 + 0.44, n = 10)). In the
ACC, the levels of mI/tCr (p = 0.036) and (Glu+Gln)/tCr (p = 0.026) were significantly
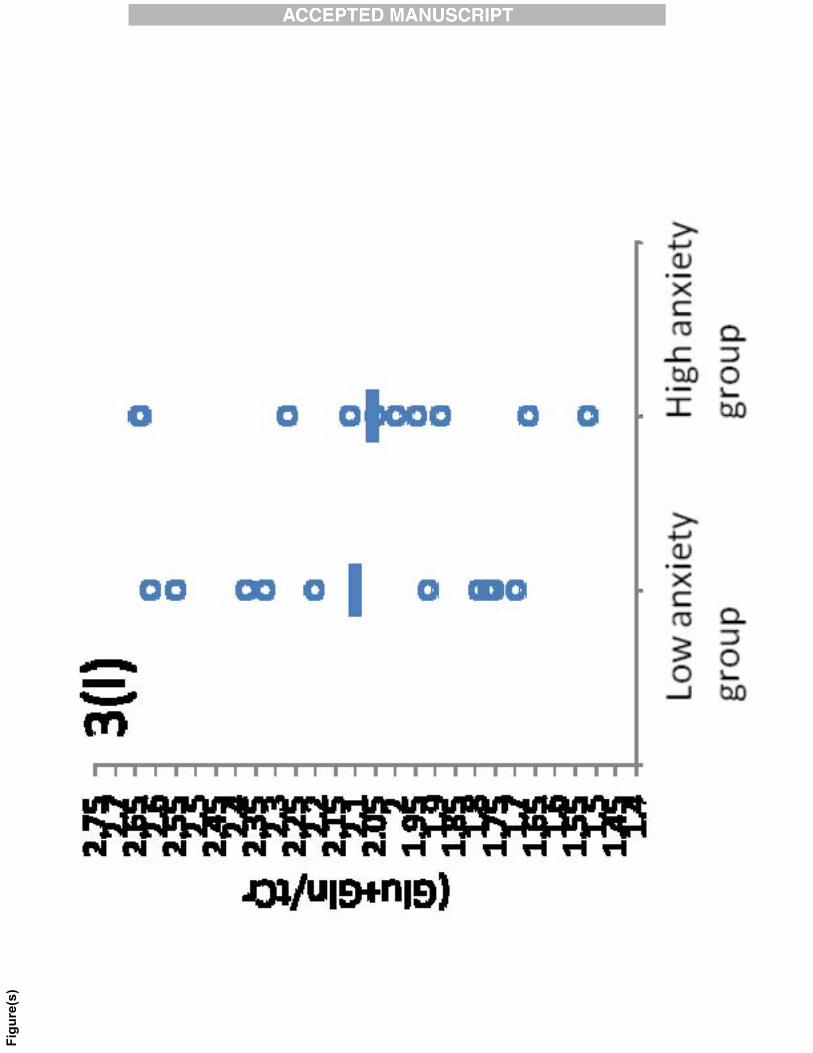
higher in high anxiety group than in the low anxiety group (Table 1, Fig. 3a, 3b, 3c, 3d, 3e,
3f, 3g, 3h, 3i, 3j, 3k, 3l). The level of Glu/tCr also showed an increase in the high anxiety
group as compared with the low anxiety group, but with marginal significance (p = 0.051). In
the hippocampus, none of the metabolites/neurochemicals showed significant differences
between the two groups (Table 1).
Page 12
When the subjects were divided into two groups based on their trait anxiety sub-score, the
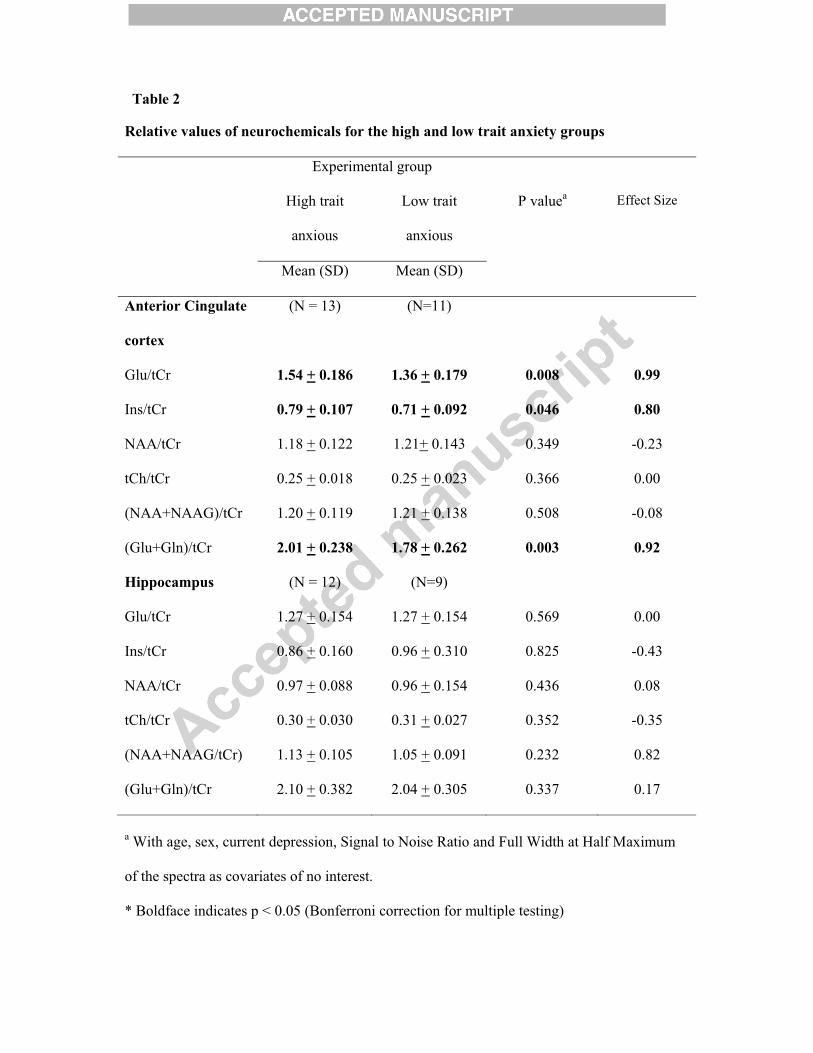
levels of Glu/tCr (p = 0.008), mI/tCr (p = 0.046) and (Glu+Gln)/tCr (p = 0.003) were found
to be significantly elevated in the high trait anxiety group as compared with the low trait
anxiety group in the anterior cingulate cortex (Table 2). In the hippocampus, none of the
metabolites/neurochemicals showed a significant difference between the two groups (based
on trait anxiety scores) (Table 2).
3.3. Correlation between neurochemical variables and anxiety
In the anterior cingulate cortex, partial correlation analysis revealed that the total anxiety
scores of the subjects were positively correlated with the Glu/tCr ratio and the Glu+Gln/tCr
ratio (r =0.462, p = 0.023 for Glu and r = 0.573, p = 0.005 for Glu+Gln, Fig. 4). mI/tCr and
tCh/tCr also showed a positive correlation with total anxiety scores but with marginal
significance (r =0.377, p = 0.056 for mI and r = 0.358, p = 0.066 for tCh). In the
hippocampus, a marginally significant positive correlation was obtained between total
anxiety and the Glu/tCr ratio (r = 0.415, P = 0.055).
Similarly, when the trait anxiety sub-score was analysed separately, the Glu/tCr (r =0.402, p
= 0.044) and Glu+Gln/tCr (r =0.461, p = 0.024) ratios showed a predictive value for trait
anxiety in the anterior cingulate cortex (Fig. 5). mI/tCr also showed a positive correlation
with trait anxiety scores but with marginal significance (r =0.380, p = 0.054) On the other
hand, in the hippocampus, no correlation was obtained between trait anxiety scores and any
of the metabolites/neurochemicals analysed.
4. Discussion
Ours is one of the few reports that demonstrate the relationship between regional
neurochemical changes in the brain and anxiety in healthy individuals. Considering the
Page 13
importance of the anterior cingulate cortex (ACC) and the hippocampus in regulating
emotional processing and their implication in anxiety, they were chosen as ROIs. In the ACC,
we obtained an increase in mI, Glu and the combination Glu+Gln in high anxious subjects as
compared with low anxious ones. Preliminary findings further suggest that total anxiety
predicts the concentration of Glu and the combination Glu+Gln in the ACC. The Glu levels
in the hippocampus were also found to be positively correlated with total anxiety, although at
a marginally significant level. It has been consistently suggested in the literature that changes
in the unresolved glutamate and glutamine (Glu+Gln) resonance are predominantly due to
changes in Glu (Phan et al., 2004; Cortese and Phan, 2005). Glutamate is the major excitatory
neurotransmitter in the mammalian brain. Limbic and associated paralimbic structures (that
include the hippocampus and the ACC, respectively) have been identified as being richly
innervated by glutamatergic pyramidal cells (Cortese and Phan, 2005). In a recent study that
carried out metabolomic analyses on a trait anxiety mouse model, the major excitatory
neurotransmitter glutamate, which binds to the N-methyl-D-aspartate (NMDA) receptor, was
found at higher levels in the plasma of a High Anxiety Behaviour mouse as compared with a
Low Anxiety Behaviour mouse (Zhang et al., 2011). Our findings are also consistent with
previous MRS studies providing evidence that anxiety disorders are associated with
alterations in the glutamate system. A study on generalized SAD (Phan et al., 2005) showed
an increase in Glu/Cr in the ACC of patients as compared with controls along with a
correlation of Glu/Cr with the intensity of social anxiety symptoms. The authors explained
the findings on the basis of proposed models for glutamate’s role in anxiety according to
which an increased glutamatergic transmission and/or excessive glutamate release within the
limbic system is associated with fear-related learning and reactivity (Walker and Davis,
2002). Further, the hyperresponsive limbic system in response to social threat/scrutiny and
anxiety-provoking situations in patients with social anxiety, as well as its attenuation on
Page 14
successful treatment, suggests the functional significance of glutamate in SAD (Phan et al.,
2005). Similarly, Grachev and Apkarian (2000) reported an increase in Glu in the orbito-
frontal cortex in healthy individuals with high state-trait anxiety. A recent study on pediatric
generalized anxiety disorder (GAD) patients showed that Glu/Cr is related to the severity of
anxiety symptoms and suggested the possibility that dysregulation of Glu within the ACC
may be linked to the pathophysiology of pediatric GAD (Strawn et al., 2013). Dynamic
changes in Glu concentrations have also been observed in the ACC during experimentally
induced panic (Zwanzger et al., 2013), suggesting a disturbance of the inhibitory-excitatory
equilibrium. Human genetic studies and clinical drug trials also present evidence for the
involvement of glutamate in anxiety disorders (Cortese and Phan, 2005). In a study by Arnold
et al. (2004), a significant association between a glutamate system gene, GRIN2B (glutamate
NMDA receptor subtype 2B), and OCD diagnosis and lifetime symptom severity was
obtained, thereby supporting glutamate system changes in the pathophysiology of anxiety.
Clinical drug trials using compounds such as phenytoin and topiramate that have direct
actions on glutamate neurotransmission have been reported for the treatment of anxiety
disorders (Cortese and Phan, 2005). Earlier, neuroimaging-based morphological and functional
studies have also shown that glutamate-rich brain regions such as the hippocampus,
amygdala, and ACC are either structurally altered or functionally hyperactive in patients
diagnosed with anxiety disorders (Cortese and Phan, 2005). Our findings of increased Glu/tCr
in the ACC and hippocampus document a direct relationship between the glutamate system
and the neurobiology of anxiety in healthy individuals, thereby suggesting a role of glutamate
in mediating the physiological and behavioral consequences associated with anxiety even at a
sub-clinical level.
Other metabolites that showed a relation with anxiety were mI and tCh. mI is an intermediate
in the cerebral inositol poly-phosphate (IPP) cascade and plays an important role in
Page 15
intracellular signaling pathways. It acts as a second messenger that liberates Ca2+ from the
endoplasmic reticulum and is involved in the recognition of chemical signals (Grachev and
Apkarian, 2000). In a recent study on monozygotic twin pairs, it was found that GAD genetic
risk predicted increases in myo-inositol in the amygdala (Hettema et al., 2012). The
proteomic and metabolomic study by Zhang et al. (2011) has also shown the inositol pathway
to be critically involved in the anxiety phenotype with several proteins and metabolites that
are part of phosphatidylinositol signaling having altered expression levels. However,
according to the authors, there is not enough evidence available to know whether these
pathways are causative or the result of distinct anxiety endophenotypes. Grachev and
Apkarian (2000) also reported an increase in mI in the orbitofrontal cortex of high anxiety
individuals. Our findings further suggest that inositol pathways are implicated in the anxiety
phenotype even at a sub-clinical level. An increased tCh in the ACC might suggest an
increase in overall cell density and/or the rate of membrane turnover as a function of anxiety.
Similar findings in Glu, Glu+Gln and mI were obtained when the trait anxiety sub-score of
the STAI was used as the parameter to classify the subjects into two groups – high trait
anxious and low trait anxious. The results further provide neuroimaging-based evidence that
both Glu and mI levels are altered as a function of sub-clinical anxiety.
In contrast to the findings in healthy subjects (Grachev and Apkarian, 2000; Fan et al., 2010)
and subjects with various anxiety disorders like SAD and OCD (Phan et al., 2005; Whiteside
et al., 2006), i.e., that increased NAA concentration was associated with anxiety symptoms,
we did not find any significant difference between NAA/tCr ratios between the two anxiety
groups. NAA is a pre-product of the excitatory neurotransmitter aspartate and is considered to
be a marker of neuronal density and integrity. Therefore, earlier MRS researchers had
hypothesized that a perpetual level of high anxiety leads to an increase in excitatory
neurotransmitter release and subsequent neuronal reorganization. It may further lead to an
Page 16
increase in the number of axons and synaptic connections, which is suggested by an increase
in the levels of NAA (Grachev and Apkarian, 2000; Phan et al., 2005). One of the reasons for
this discrepancy in findings might be the region of the brain studied. The earlier findings
were mainly obtained in the orbito-frontal cortex or the medial prefrontal cortex, as against
the ACC or hippocampus in our study. Moreover, as suggested by the authors of earlier
studies (Grachev and Apkarian, 2000; Phan et al., 2005), the findings in NAA should be
considered preliminary and should be reexamined using morphometric techniques. Moreover,
a recent morphometric study in major depressive disorder and anxiety (SAD and PD),
showed lower grey matter volumes in the rostral anterior cingulate gyrus extending into the
dorsal anterior cingulate gyrus in both depression and anxiety patients (van Tol et al., 2010).
Therefore, the putative association between NAA and sub-clinical anxiety in healthy
populations needs further investigation.
In summary, this is the first study suggesting alterations in ACC neurochemistry in relation
with anxiety in healthy subjects, with glutamate, combined glutamate and glutamine and
myo-inositol showing the maximum changes. The study thus contributes to the limited
literature available on altered metabolism and neural mechanisms underlying sub-clinical
anxiety. The limitations of our study include a small sample size and the absence of measures
of absolute concentrations of the neurochemicals. Moreover, even though the healthy subjects
were recruited by screening them for current or past medical and psychiatric illness using the
medical history questionnaire of the Hindi version of the Diagnostic Interview for Genetic
Studies (version 2) (Deshpande et al., 1998), the absence of full psychiatric evaluation, for
example, the structured Clinical Interview for DSM-IV Axis I Disorders (SCID-I) (First et
al., 2002), is another limitation of the study. Along with earlier etiological models that
suggest high anxiety trait to be a precursor to anxiety disorders and depression, the observed
Page 17
neurochemical changes in our study, which are similar to those obtained in various anxiety
disorders, further support the need for preventive interventions in high anxiety individuals.
Acknowledgement This work was supported by DRDO R&D Project No. INM 311 (4.1).
References
Arnold, P.D., Rosenberg, D.R., Mundo, E., Tharmalingam, S., Kennedy, J.L., Richter, M.A.,
2004. Association of a glutamate (NMDA) subunit receptor gene (GRIN2B) with obsessive-
compulsive disorder: a preliminary study. Psychopharmacology (Berl) 174, 530-538.
Baur, V., Hänggi, J., Jäncke, L., 2012. Volumetric associations between uncinate fasciculus,
amygdala, and trait anxiety. BMC Neuroscience 13, 4.
Beck, A.T., Steer, R.A., Brown, G.K., 1996. Beck Depression Inventory: Manual, 2nd ed. The
Psychological Corporation, San Antonio, TX.
Bishop, S.J., Duncan, J., Brett, M., Lawlence, A.D., 2004. Prefrontal cortical function and
anxiety: controlling attention to threat related stimuli. Nature Neuroscience 7(2), 184-188.
Bishop, S.J., 2009. Trait anxiety and impoverished prefrontal control of attention. Nature
Neuroscience 12 (1), 92-98.
Cortese, B.M., Phan, K.L., 2005. The role of glutamate in anxiety and related disorders. CNS
Spectrums 10 (10), 820-830.
Page 18
Dager, S.R., Friedman, S.D., Heid, A., Layton, M.E., Richards, T., Artru, A. Strauss, W.,
Hayes, C., Posse, S., 1999. Two-dimensional proton echo-planar spectroscopic imaging of
brain metabolic changes during lactate-induced panic. Archives of General Psychiatry 56, 70–
77.
Delgado, M.R., Nearing, K.I., Ledoux, J.E., Phelps, E.A., 2008. Neural circuitry underlying the
regulation of conditioned fear and its relation to extinction. Neuron 59, 829–838.
Deshpande, S.N., Mathur, M.N.L., Das, S.K., Bhatia, T., Sharma, S.D., Nimgaonkar, V.L.,
1998. A Hindi version of the diagnostic interview for genetic studies. Schizophrenia Bulletin
24 (3), 489–493.
Egner, T., Etkin, A., Gale, S., Hirsch, J., 2008. Dissociable neural systems resolve conflict
from emotional versus nonemotional distracters. Cerebral Cortex 18, 1475–1484.
Etkin, A., Egner, T., Kalisch, R., 2011. Emotional processing in anterior cingulate and medial
prefrontal cortex. Trends in Cognitive Sciences 15 (2), 85-93.
Eysenck, M.W., Derakshan, N., Santos, R., Calvo, M.G., 2007. Anxiety and cognitive
performance: attentional control theory. Emotion 7 (2), 336–353.
Fan, Q., Tan, L., You, C., Wang, J., Ross, C.A., Wang, X., Zhang, T., Li, J., Chen, K., Xiao,
Z., 2010. Increased N-Acetylaspartate/creatine ratio in the medial prefrontal cortex among
unmedicated obsessive-compulsive disorder patients. Psychiatry and Clinical Neurosciences 64
(5), 483-490.
Page 19
First, M. B., Spitzer, R. L., Miriam, G., Williams, J. B.W., 2002. Structured Clinical Interview
for DSM-IV-TR Axis I Disorders, Research Version, Patient edition (SCID-I/P). Biometrics
Research, New York State Psychiatric Institute, New York, NY.
Grachev, I.D., Apkarian, A.V., 2000. Anxiety in healthy humans is associated with orbital
frontal chemistry. Molecular Psychiatry 5, 482–488.
Heckers, S., 2001. Neuroimaging studies of the hippocampus in schizophrenia. Hippocampus
11 (5), 520-528.
Hermens, D.F., Lagopoulos, J., Naismith, S.L., Tobias-Webb, J., Hickie, I.B., 2012. Distinct
neurometabolic profiles are evident in the anterior cingulate of young people with major
psychiatric disorders. Translational Psychiatry 2, e110.
Hettema, J.M., Kettenmann, B., Ahluwalia, V., McCarthy, C., Kates, W.R., Schmitt, J.E.,
Silberg, J.L., Neale, M.C., Kendler, K,S., Fatouros, P., 2012. A pilot multimodal twin imaging
study of generalized anxiety disorder. Depression and Anxiety 29 (3), 202–209.
Hong, S.B., Shin, Y.W., Kim, S.H., Yoo, S.Y., Lee, J.M., Kim, I.Y. Kim, S.I., Kwon, J.S.,
2007. Hippocampal shape deformity analysis in obsessive-compulsive disorder. European
Archives of Psychiatry and Clinical Neurosciences 257 (4), 185-190.
Keshavan, M.S., Kapur, S., Pettegrew, J.W., 1991. Magnetic resonance spectroscopy in
psychiatry: potential, pitfalls, and promise. American Journal of Psychiatry 148 (8), 976-85.
Page 20
Kimbrell, T., Leulf, C., Cardwell, D., Komoroski, R.A., Freeman, T.W., 2005. Relationship of
in vivo medial temporal lobe magnetic resonance spectroscopy to documented combat
exposure in veterans with chronic posttraumatic stress disorder. Psychiatry Research:
Neuroimaging 140 (1), 91-94.
Lieberman, M.D. Eisenberger, N.I., Crockett, M.J., Tom, S.M., Pfeifer, J.H., Way, B.M., 2007.
Putting feelings into words: affect labeling disrupts amygdala activity in response to affective
stimuli. Psychological Science 18, 421–428.
Lochner, C., Fouché, J.-P., Plessis, S., Spottiswoode, B., Seedat, S., Fineberg, N.,
Chamberlain, S.R., Stein, D.J., 2012. Evidence for fractional anisotropy and mean diffusivity
white matter abnormalities in the internal capsule and cingulum in patients with obsessive–
compulsive disorder. Journal of Psychiatry and Neuroscience 37 (3), 193–199.
Mahmutyaz�c�o�lu, K., Konuk, N., Özdemir, H., Atasoy, N., Atik, L., Gündo�du, S., 2005.
Evaluation of the hippocampus and the anterior cingulate gyrus by proton MR spectroscopy in
patients with post-traumatic stress disorder. Diagnostic and Interventional Radiology 11, 125-
129.
Miller, B.L., 1991. A review of chemical issues in 1H NMR spectroscopy: N-acetyl-L-
aspartate, creatine and choline. NMR in Biomedicine 4, 47-52.
Modi, S., Trivedi, R., Singh, K., Kumar, P., Rathore, R.K.S., Tripathi, R.P., Khushu, S., 2013.
Individual differences in trait anxiety are associated with white matter tract integrity in fornix
and uncinate fasciculus: preliminary evidence from a DTI based tractography study.
Behavioural Brain Research 238, 188–192.
Page 21
Montag, C., Reuter, M., Weber, B., Marketta, S., Schoene-bake, J-C., 2012. Individual
differences in trait anxiety are associated with white matter tract integrity in the left temporal
lobe in healthy males but not females. Neuroscience 217, 77–83.
Phan, K.L., Fitzgerald, D.A., Cortese, B.M., Seraji-Bozorgzad, N., Tancer, M.E., Moore, G.J.,
2004. Anterior cingulate neurochemistry in social anxiety disorder: 1H-MRS at 4Tesla.
Neuroreport 16 (2), 183-186.
Provencher, S.W., 1993. Estimation of metabolite concentrations from localized in vivo NMR
spectra. Magnetic Resonanace in Medicine 30, 672–679.
Pujol, J., Lo´pez, A., Deus, J., Cardoner, N., Vallejo, J., Capdevila, A., Paus, T., 2002.
Anatomical variability of the anterior cingulate gyrus and basic dimensions of human
personality. NeuroImage 15, 847–855.
Sala, M., Perez, J., Soloff, P., Ucelli di Nemi, S., Caverzasi, E., Soares, J.C., Brambilla, P.,
2004. Stress and hippocampal abnormalities in psychiatric disorders. European
Neuropsychopharmacology 14 (5), 393-405.
Sandi, C., Richter-Levin, G., 2009. From high anxiety trait to depression: a neurocognitive
hypothesis. Trends in Neuroscience 32 (6), 312-320.
Spielberger, C.D., 1983. Manual for the State-Trait Anxiety Inventory. Consulting
Psychologists Press, Palo Alto, CA.
Page 22
Strawn, J.R., Chu, W.J., Whitsel, R.M., Weber, W.A., Norris, M.M., Adler, C.M., Eliassen,
J.C., Phan, K.L., Strakowski, S.M., DelBello, M.P., 2013. A pilot study of anterior cingulate
cortex neurochemistry in adolescents with generalized anxiety disorder. Neuropsychobiology
67 (4), 224-229.
van Tol, M.J., van der Wee, N.J., van den Heuvel, O.A., Nielen, M.M., Demenescu, L.R.,
Aleman, A., Renken, R., van Buchem, M.A., Zitman, F.G., Veltman, D.J., 2010. Regional
brain volume in depression and anxiety disorders. Archives of General Psychiatry 67 (10),
1002-1011.
Walker, D.L., Davis, M., 2002. The role of amygdala glutamate receptors in fear learning, fear-
potentiated startle, and extinction. Pharmacology, Biochemistry and Behavior 71, 379–392.
Westlye, L.T., Bjørnebekk, A., Grydeland, H., Fjell, A.M., Walhovd, K.B., 2011. Linking an
anxiety-related personality trait to brain white matter microstructure diffusion tensor imaging
and harm avoidance. Archives of General Psychiatry 68 (4), 369–377.
Whiteside, S.P., Port, J.D., Deacon, B.J., Abramowitz, J.S., 2006. A magnetic resonance
spectroscopy investigation of obsessive–compulsive disorder and anxiety. Psychiatry
Research: Neuroimaging 146, 137–147.
Zhang, Y., Filiou, M.D., Reckow, S., Gormanns, P., Maccarrone, G., Kessler, M.S., Frank, E.,
Hambsch, B., Holsboer, F., Landgraf, R., Turck, C.W., 2011. Proteomic and metabolomic
profiling of a trait anxiety mouse model implicate affected pathways. Molecular & Cellular
Proteomics 10, 12.
Page 23
Zwanzger, P., Zavorotnyy, M., Gencheva, E., Diemer, J., Kugel, H., Heindel, W., Ruland, T.,
Ohrmann, P., Arolt, V., Domschke, K., Pfleiderer, B., 2013. Acute shift in glutamate
concentrations following experimentally induced panic with cholecystokinin tetrapeptide--a
3T-MRS study in healthy subjects. Neuropsychopharmacology 38 (9), 1648-1654.
Figure captions:
Fig. 1: Location of (a) 4-cm3 voxel in anterior cingulate cortex and (b) 2.5-cm3 voxel in right
hippocampus of a normal subject.
Fig. 2: Typical spectrum from (a) anterior cingulate cortex and (b) right hippocampus as
analysed by the LCModel. The concentrations of the metabolites and their associated
Cramer-Rao bounds are listed in the box on the right.
Fig. 3: Scatter plots for (a) Glu/tCr (b) Ins/tCr (c) NAA/tCr (d) tCh/tCr (e)
(NAA+NAAG)/tCr (f) (Glu+Gln)/tCr in the anterior cingulate cortex and (g) Glu/tCr (h)
Ins/tCr (i) NAA/tCr (j) tCh/tCr (k) (NAA+NAAG)/tCr (l) (Glu+Gln)/tCr in the hippocampus
in low and high anxiety groups. The horizontal lines in the scatter plot represent the mean
values.
Fig. 4: Partial correlation plots (controlling for age, sex, BDI, FWHM and SNR of the
spectra) between levels of (a) Glu/tCr and total anxiety scores and (b) (Glu+Gln)/tCr and total
anxiety scores in anterior cingulate cortex of the healthy subjects.
Fig. 5: Partial correlation plots (controlling for age, sex, BDI, FWHM and SNR of the
spectra) between levels of (a) Glu/tCr and trait anxiety scores and (b) (Glu+Gln)/tCr and trait
anxiety scores in anterior cingulate cortex of the healthy subjects.
Page 24
Table 1
Relative values of neurochemicals for the high and low anxiety groups
Experimental group
High anxious Low anxious P valuea Effect Size
Mean (SD) Mean (SD)
Anterior Cingulate
Cortex
(N = 13) (N=11)
Glu/tCr 1.52 (0.166) 1.36 (0.210) 0.051# 0.85
Ins/tCr 0.79 (0.109) 0.71 (0.087) 0.036* 0.82
NAA/tCr 1.22 (0.131) 1.16 (0.127) 0.435 0.46
tCh/tCr 0.25 (0.016) 0.24 (0.025) 0.107 0.49
(NAA+NAAG)/tCr 1.23 (0.123) 1.17 (0.128) 0.418 0.48
(Glu+Gln)/tCr 2.03 (0.227) 1.76 (0.252) 0.026* 1.13
Hippocampus (N = 11) (N=10)
Glu/tCr 1.268 (0.146) 1.274 (0.186) 0.605 -0.04
Ins/tCr 0.820 (0.171) 0.992 (0.271) 0.356 -0.77
NAA/tCr 0.998 (0.104) 0.955 (0.132) 0.561 0.36
tCh/tCr 0.311 (0.032) 0.300 (0.024) 0.626 0.39
(NAA+NAAG/tCr) 1.110 (0.102) 1.085 (0.114) 0.511 0.23
(Glu+Gln)/tCr 2.057 (0.352) 2.101 (0.353) 0.868 -0.12
a With age, sex, current depression, Signal to Noise Ratio and Full Width at Half Maximum
of the spectra as covariates of no interest.
* Boldface indicates p < 0.05 (Bonferroni correction for multiple testing); # marginally
significant differences at p < 0.10 are indicated in italics.
Page 25
Table 2
Relative values of neurochemicals for the high and low trait anxiety groups
Experimental group
High trait
anxious
Low trait
anxious
P valuea Effect Size
Mean (SD) Mean (SD)
Anterior Cingulate
cortex
(N = 13) (N=11)
Glu/tCr 1.54 + 0.186 1.36 + 0.179 0.008 0.99
Ins/tCr 0.79 + 0.107 0.71 + 0.092 0.046 0.80
NAA/tCr 1.18 + 0.122 1.21+ 0.143 0.349 -0.23
tCh/tCr 0.25 + 0.018 0.25 + 0.023 0.366 0.00
(NAA+NAAG)/tCr 1.20 + 0.119 1.21 + 0.138 0.508 -0.08
(Glu+Gln)/tCr 2.01 + 0.238 1.78 + 0.262 0.003 0.92
Hippocampus (N = 12) (N=9)
Glu/tCr 1.27 + 0.154 1.27 + 0.154 0.569 0.00
Ins/tCr 0.86 + 0.160 0.96 + 0.310 0.825 -0.43
NAA/tCr 0.97 + 0.088 0.96 + 0.154 0.436 0.08
tCh/tCr 0.30 + 0.030 0.31 + 0.027 0.352 -0.35
(NAA+NAAG/tCr) 1.13 + 0.105 1.05 + 0.091 0.232 0.82
(Glu+Gln)/tCr 2.10 + 0.382 2.04 + 0.305 0.337 0.17
a With age, sex, current depression, Signal to Noise Ratio and Full Width at Half Maximum
of the spectra as covariates of no interest.
* Boldface indicates p < 0.05 (Bonferroni correction for multiple testing)