j ourna l homepage: www.e lsev ie r .com/ locate /phb
Glutamate/GABA balance in ACC modulates the nociceptive responses ofvocalization: An expression of affective-motivational component of painin guinea pigs
João Zugaib 1, Márcio Ramos Coutinho 1, Mateus Dalbem Ferreira, Leda Menescal-de-Oliveira ⁎Department of Physiology, Faculty of Medicine of Ribeirão Preto, University of São Paulo, 14.049-900 Ribeirão Preto, SP, Brazil
H I G H L I G H T S
• The anterior cingulate cortex (ACC) modulates nociceptive response in guinea pigs.• Glutamate agonist (DLH) injection in the ACC produced pronociception.• GABAA antagonist bicuculline injected in the ACC caused robust pronociception.• Muscimol microinjection in the ACC after MK-801 produced antinociception.
⁎ Corresponding author at: Department of Physiology,Preto University of São Paulo, 14049-900 Ribeirão Preto3333; fax: +55 16 3602 0221.
E-mail address: [email protected] (L. Menescal-d1 These authors contributed equally to this work
Evidence corroborates the role of the anterior cingulate cortex (ACC) in themodulation of cognitive and emotion-al functions. Its involvement in themotivational-affective component of pain has beenwidely investigated usingdifferent methods to elucidate the specific role of different neurotransmitter systems. We used the peripheralnoxious stimulus-induced vocalization algesimetric test to verify glutamatergic and GABAergic neurotransmis-sion in the guinea pig ACC. Microinjection of homocysteic acid (DLH; 30 nmol) in the left guinea pig ACC in-creased the amplitude of vocalizations (pronociception) compared to controls injected with saline. Moreover,microinjection ofMK-801 (3.6 nmol), an NMDA receptor antagonist, did not alter the amplitude of vocalizations,but its microinjection prior to DLH prevented the increase in vocalizations induced by this drug. Regarding theGABAergic system, blockade of GABAA receptors with bicuculline (1 nmol) increased the amplitude of vocaliza-tions, while three different doses of the GABAA agonist muscimol (0.5, 1 and 2 nmol) did not influence nocicep-tive vocalization responses. Finally, a combination of MK-801 (3.6 nmol) and muscimol (1 nmol) reduced theamplitude of vocalizations (antinociception), suggesting that a combination of glutamate and GABA in the ACCmodulates the expression of affective-motivational pain response.We suggest that activation of NMDA receptorsor blockade of GABAergic neurotransmission promotes pronociception and that the antinociceptive effect ofmuscimol depends on the blockade of NMDA receptors.
The anterior cingulate cortex (ACC) is known to modulate severalmotivational responses, such as anxiety, conditioned fear, post traumaticdisorder and tonic immobility [1–4]. Interestingly, several studies usingvarious approaches have shown that the ACC is involved in affectiveandmotivational painmediation [5–7]. Several neurotransmitters appearto participate in this process [8,9].
School of Medicine of Ribeirão, SP, Brazil. Tel.: +55 16 3602
e-Oliveira).
ghts reserved.
Within this context, Johansen and Fields [6] demonstrated thatglutamatergic activation in the rat ACC is necessary and sufficient forformalin-induced conditioned place aversion, an affective and learnedpain response. Behavioral and immunohistochemistry studies stronglysupport that the NMDA glutamatergic receptors are crucially involvedin this mechanism [7]. Others neurotransmitters are also involved inaffective pain. For instance, Bozkurt et al. [10] mapped the distributionof the various receptors in primate ACC and revealed high concentrationsof GABAA inhibitory receptors. LaGraize and Fuchs [11] demonstrated theattenuation of escape/avoidance behavior induced by noxious stimulusin L5-ligated rats after muscimol (GABAA agonist) administration inthe ACC. The results from whole-cell patch clamp studies associatedwith behavioral analyses have shown that the ACC GABAergic systemis involved in affective pain and interacts with the ACC glutamatergicsystem [8].
In our laboratory, the neural substrates involved in non-anesthetized guinea pigs (Cavia porcellus) nociception have frequentlybeen investigated by means of vocalization nociceptive test, in whichvocalization is induced by the application of a peripheral noxious stim-ulus [12–16]. We have been using this approach since 1977 [12] andprovided substantial contributions to the understanding of some neuralcircuits that modulate the affective and emotional aspects of pain. Infact, showing the same phenomenon in different animal models whenit presents itself preserved in different species, strengthens inferencesabout its adaptive significance. This idea is supported by our studysince vocalizations are elicited in various high aversive emotional states[17–19] and has been considered an expression of affective pain.Moreover, the vocalization control circuits are composed of the samesupra-spinal structures that are involved in pain mediation [20–23].
To our knowledge, there are no studies analyzing the role of theguinea pig ACC in pain affective-motivational response. Likewise,there are no investigations concerning the glutamatergic andGABAergicmechanisms involved in this response. However, these approaches maybe important to understanding the neural mechanisms of unpleasantpain. Thus, the current work proposed to evaluate the effects of gluta-matergic and GABAergic agonists and antagonists in the guinea pigACC on the vocalization induced by peripheral noxious stimulus appli-cation. In addition, the present study aimed to analyze the interactionbetween these two systems in this motivational nociceptive response.
2. Material and methods
2.1. Animals
In the present study, we used male guinea pigs (Cavia porcellus)weighing 400–500 g, provided by the animal facilities of the Facultyof Medicine of Ribeirão Preto (FMRP). The animals were housed inplexiglass cages (56 cm × 37 cm × 39 cm) with a controlled tem-perature (24 ± 1 °C), 12/12 h cycles of light/dark and food andwater ad libitum. The conditions of animal maintenance and experi-mental procedures complied with the standards of the guide for thecare and use of laboratory animals, according to the Brazilian Collegeof Animal Experimentation (COBEA). The experiments were submittedto and approved by the Ethics Committee on Animal Experimentation(CETEA) of the FMRP, University of São Paulo (protocol number145/2010). The guidelines of the Committee for Research and EthicalIssues of IASP [24] were followed throughout the experiments.
2.2. Surgical procedures
The animals were deeply anesthetized with a combination of40 mg/kg ketamine hydrochloride and 5 mg/kg xylazine (intramus-cular injection) and placed in a stereotaxic apparatus (Davis-KopfInstruments, USA). A guide cannula (10 mm long and 0.6 mm outerdiameter) was then implanted 1 mm dorsal to the left ACC usingthe following coordinates from the atlas of Rössner [25] for guineapigs: 1.2 mm rostral to bregma, 0.4 mm lateral to the midline and4.7 mm above the intra-aural line. The guide cannula was fixed withautopolymerizing resin and anchored with a screw that was threadedthrough the skullcap. Then, the animals received a subcutaneous injec-tion of Banamine (Schering-Plough, flunixin meglumine, 2.5 mg/kg,10 mg/ml, 0.2 ml), which has analgesic and anti-inflammatoryproperties.
2.3. Vocalization algesimetric test
This test consists of applying an acute peripheral noxious stimulus(electric shock) that causes the emission of a vocalization response,interpreted as an expression of pain. A pair of non-insulated electrodes(1.0 cm long) was implanted in the subcutaneous region of the rightthigh. The animals were placed in an acrylic box lined with nylon
foam. After 20 min of acclimation, the experimental protocol was initi-ated. The electrodes were connected an electric stimulator that releasedthree intermittent stimulus with pulses (AC current with square waves,100-Hz frequency, and 0.5-ms duration) of varying intensity (0.6 to4.0 mA) sufficient to induce audible vocalization, which is the most fre-quent response elicited by guinea pigs during nociceptive stimulation.The intensity of stimulus was the minimum needed to produce thevocalization in each guinea pig. Once the threshold value (baseline)was established for each animal, the intensity was maintained at aconstant level throughout the experiment. Vocalizations were emittedduringnoxious stimulation, and only a small number of animals showedpost-stimulus vocalization. Electrical stimulation (3 second duration)induced vocalizations that did not persist in the short inter-stimulus in-tervals. After completing the baseline, noxious peripheral stimulationwas applied at 2, 5, 15, 30, 45 and 60 min after the treatment withvarious drugs.
Vocalization was recorded by an Aiwa DM-64 microphone con-nected to the preamplifier of a polygraph. The peak amplitude ofthe polygraphic recordings was proportional to the intensity of theanimal's vocalization. These peaks were measured in millimeters,and the average response for each period was used for quantitativeevaluation.
2.4. Drugs
The drugs used in this study were DLH (DL-homocysteic acid; excit-atory amino acid agonists), MK-801 (NMDA glutamate receptor antag-onist), bicuculline methiodide (GABAA antagonist), and muscimol(GABAA receptor agonist). All drugs were purchased from SigmaSt. Louis — USA, and doses were based on previous work from ourlaboratory [4,15].
2.5. Experimental procedures
After 5–7 days of recovery from surgery, the different groups weresubmitted to a vocalization test. Glutamatergic and GABAergic neuro-transmission, as well as their interaction, was tested with differenttreatments. Initially, we investigated the effect of microinjection ofunilateral homocysteic acid (DLH) and MK-801 (NMDA antagonist) inthe guinea pig ACCs on the amplitude of vocalization during a noxiousperipheral stimulation. In Group 1 (n = 7), animals received amicroin-jection of saline (0.9% NaCl) as the vehicle control. In groups 2 (n = 8)and 3 (n = 6), animals received amicroinjection of DLH (30 nmol) andMK-801 (3.6 nmol), respectively. Group 4 (n = 8)was pretreated withMK-801 10 min prior to the microinjection of DLH. In group 5 (n = 8),we investigated the effect of an isolated microinjection of 1 nmol ofbicuculline (GABAA antagonist) on the amplitude of vocalizations.Then, groups 6 (n = 8), 7 (n = 8), and 8 (n = 6) were treated with amuscimolmicroinjection in three different doses (0.5, 1 and 2 nmol, re-spectively). Furthermore, we tested the microinjection of 1 nmol ofmuscimol 10 min prior to microinjection of 1 nmol of bicuculline(Group 9, n = 8). Finally, we investigated the interaction of glutamate/GABA in group 10 (n = 8) with a microinjection of 3.6 nmol of MK-80110 min prior to a microinjection of 1 nmol of muscimol. It is impor-tant to note that each group of animals was used for one treatment only.
Microinjections were performed with a Hamilton microsyringe(10 μl) connected to PE-10 polyethylene tubing and attached to aMizzy needle segment (0.3 mm outer diameter; 1.0 mm longer thanthe guide cannula). In all experimental groups, 0.2 μl wasmicroinjectedover a period of 1 min, and theMizzy needle was left in place for an ad-ditional 40 s to avoid reflux.
2.6. Histological analysis
After the behavioral tests were completed, the injection sites wereidentified by microinjection of 0.2 μl of 2% pontamine sky blue dye.
10 J. Zugaib et al. / Physiology & Behavior 126 (2014) 8–14
Each animal was deeply anesthetized with sodium pentobarbital andperfused intracardiallywith saline, followed by 10% formalin. The brainswere removed and fixed in 10% formalin. Routine histological proce-dures were used for tissue sectioning, and the stained tissues wereobserved under a light microscope to determine the location of thestimulation sites according to the atlas of Rössner [25].
2.7. Statistical analysis
The amplitude of the peaks in the graphic record of vocalization wasmeasured in millimeters (mm), and the average of each response wasinterpreted as a quantitative measure of vocalization. The results inmmwere converted to vocalization index (VI) scores using the follow-ing formula: VI = mean vocalization − value of baseline / value ofbaseline. Data are reported as the mean ± SEM, and statistical analysisof data was performed by a two-way repeated measures analysis ofvariance (ANOVA) followed by the Duncan post-hoc test. In addition,the data were considered [25] significantly different at p b 0.05.
3. Results
3.1. Nociceptive response of vocalization: Glutamatergic neurotransmissionin the ACC
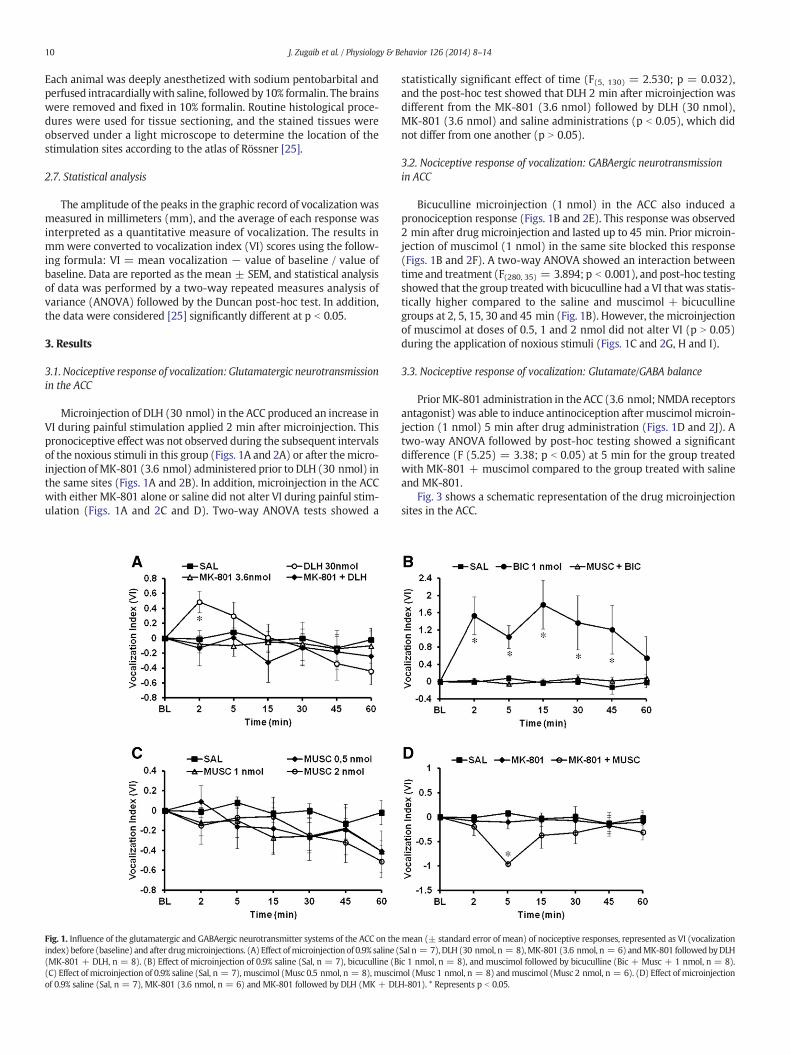
Microinjection of DLH (30 nmol) in the ACC produced an increase inVI during painful stimulation applied 2 min after microinjection. Thispronociceptive effect was not observed during the subsequent intervalsof the noxious stimuli in this group (Figs. 1A and 2A) or after themicro-injection ofMK-801 (3.6 nmol) administered prior to DLH (30 nmol) inthe same sites (Figs. 1A and 2B). In addition, microinjection in the ACCwith either MK-801 alone or saline did not alter VI during painful stim-ulation (Figs. 1A and 2C and D). Two-way ANOVA tests showed a
Fig. 1. Influence of the glutamatergic and GABAergic neurotransmitter systems of the ACC on theindex) before (baseline) and after drugmicroinjections. (A) Effect ofmicroinjection of 0.9% saline ((MK-801 + DLH, n = 8). (B) Effect of microinjection of 0.9% saline (Sal, n = 7), bicuculline (B(C) Effect of microinjection of 0.9% saline (Sal, n = 7), muscimol (Musc 0.5 nmol, n = 8), muscimof 0.9% saline (Sal, n = 7), MK-801 (3.6 nmol, n = 6) and MK-801 followed by DLH (MK + DL
statistically significant effect of time (F(5, 130) = 2.530; p = 0.032),and the post-hoc test showed that DLH 2 min after microinjection wasdifferent from the MK-801 (3.6 nmol) followed by DLH (30 nmol),MK-801 (3.6 nmol) and saline administrations (p b 0.05), which didnot differ from one another (p N 0.05).
3.2. Nociceptive response of vocalization: GABAergic neurotransmissionin ACC
Bicuculline microinjection (1 nmol) in the ACC also induced apronociception response (Figs. 1B and 2E). This response was observed2 min after drug microinjection and lasted up to 45 min. Prior microin-jection of muscimol (1 nmol) in the same site blocked this response(Figs. 1B and 2F). A two-way ANOVA showed an interaction betweentime and treatment (F(280, 35) = 3.894; p b 0.001), and post-hoc testingshowed that the group treated with bicuculline had a VI that was statis-tically higher compared to the saline and muscimol + bicucullinegroups at 2, 5, 15, 30 and 45 min (Fig. 1B). However, the microinjectionof muscimol at doses of 0.5, 1 and 2 nmol did not alter VI (p N 0.05)during the application of noxious stimuli (Figs. 1C and 2G, H and I).
3.3. Nociceptive response of vocalization: Glutamate/GABA balance
PriorMK-801 administration in the ACC (3.6 nmol; NMDA receptorsantagonist) was able to induce antinociception after muscimol microin-jection (1 nmol) 5 min after drug administration (Figs. 1D and 2J). Atwo-way ANOVA followed by post-hoc testing showed a significantdifference (F (5.25) = 3.38; p b 0.05) at 5 min for the group treatedwith MK-801 + muscimol compared to the group treated with salineand MK-801.
Fig. 3 shows a schematic representation of the drug microinjectionsites in the ACC.
mean (± standard error of mean) of nociceptive responses, represented as VI (vocalizationSal n = 7), DLH (30 nmol, n = 8),MK-801 (3.6 nmol, n = 6) andMK-801 followedbyDLHic 1 nmol, n = 8), and muscimol followed by bicuculline (Bic + Musc + 1 nmol, n = 8).ol (Musc 1 nmol, n = 8) andmuscimol (Musc 2 nmol, n = 6). (D) Effect of microinjection
H-801). * Represents p b 0.05.
Fig. 2. Polygraphic record of the vocalization amplitude (mm) of a representative animal from each group after microinjection in the guinea pig ACC. (A) DLH (30 nmol); (B) DLH(30 nmol) followed by MK-801 (3.6 nmol); (C) MK-801 (3.6 nmol); (D) 0.9% saline (0.2 μl); (E) bicuculline (1 nmol); (F) muscimol (1 nmol) followed by bicuculline (1 nmol);(G), muscimol (0.5 nmol); (H) muscimol (1 nmol); (I) muscimol (2 nmol); and (J) MK-801 (3.6 nmol) followed by muscimol (1 nmol). The horizontal bars indicate the period ofapplication of noxious stimulation (3 s) at different time intervals (minutes). The baseline (BL) is the record of vocalizations prior to microinjection of drugs.
Fig. 3. Schematic representation of the ACC sections of guinea pigs. Filled circles (•) indicate the sites of microinjections: (A) Saline (0.9% NaCl); (B) DLH (30 nmol); (C) MK-801(3.6 nmol); (D) MK-801 (3.6 nmol) followed by DLH (30 nmol); (E) bicuculline (1 nmol); (F) muscimol (1 nmol) followed by bicuculline (1 nmol); (G) muscimol (0.5 nmol);(H) muscimol (1 nmol); (I) muscimol (2 nmol); and (J) MK-801 (3.6 nmol) followed by muscimol (1 nmol). AP = anteroposterior, CC = corpus callosum, V = lateral ventricle. Theschemes were modified from the atlas of Rössner (1965) for guinea pigs.
12 J. Zugaib et al. / Physiology & Behavior 126 (2014) 8–14
4. Discussion
The present data suggest that during the application of noxious stim-uli, peripheralNMDA receptors of theACC are critical for pronociceptioninduced by glutamatergic stimulation. However, these receptors are notthe only mediators of nociceptive vocalization response because theirblockade alone does not reduce the response amplitude. RegardingGABAergic neurotransmission, microinjection of the GABAA receptorantagonist bicuculline increased VI for a longer period than treatmentwith DLH. However, microinjection of muscimol in three differentconcentrations did not alter the magnitude of VI but did induceantinociception when microinjected after MK-801. These resultssuggest that glutamatergic and GABAergic neurotransmission balanceis important in ACC nociceptive vocalization response and that thisvocalization response depends on the blockage of NMDA receptors.
The VI increase after ACC glutamatergic activation observed in thepresent study provides additional support for the assumption that theACC is a key brain structure to the processing of pronociceptive re-sponses following distinct noxious stimuli [26–29]. The blockage ofthis pronociception by MK-801 contributes to the idea that excitatoryamino acids and their NMDA receptors in the ACC play an importantrole in this processing [5,7,30–32]. Within this context, we must notethat these results were obtained by analyzing the glutamatergic systemof the caudal portion of the guinea pig ACC, specifically where thecorpus callosum is closed. This fact is important because the ACC isvery extensive and distinct responses may be elicited in different partsof this structure [33–35].
In the currentwork, an increase in VI after bicucullinemicroinjectionin the ACC was also demonstrated. This pronociceptive effect wasprevented by previous muscimol ACC administration. These datacorroborate the evidence that GABAA receptors in the ACCmay also con-siderably affect the processing of nociceptive responses [8,10,11]. In thepresent study, we also obtained interesting results from a novel proce-dure, in which we used a GABAA agonist to prevent pronociceptioncaused by a GABAA antagonist. We used this approach to investigatehow exogenous muscimol affects this behavioral response. In our opin-ion, one possible reason for this effect is the fact that muscimol may
Fig. 4. Schematic diagram of the nociceptive circuitry in the ACC. (A) At baseline, the ACC neurorons in the ACC by intense noxious stimulus, DLH stimulation or GABAergic tone inhibitionwithpronociception; (C) the inhibitory effect of themuscimol (GABAA receptor agonist) is not sufficand the result is nociception; (D) the block of NMDA receptors (glutamatergic receptors) byMKACC = anterior cingulate cortex; DLH = DL-homocysteic acid, excitatory amino acid agonists
prevail over bicuculline in the competition for GABAA receptor binding.This mechanism may have provoked a reduction in bicuculline bindingto GABAA receptors or some inhibitory cell effects [36,37]. Additionally,the influence of GABAc receptors must not be disregarded becausemuscimol is capable of binding to these receptors, while bicuculline isnot [38]. It is also important to note that muscimol activity could havebeen potentiated by the GABA endogenous pool in the interruption ofthe bicuculline effect. This synergism, however, was not enough toblock ACC excitatory discharge, as shown by the nociceptive responseselicited by the noxious stimuli.
Curiously, muscimol microinjection alone in the ACC, in three differ-ent concentrations, did not alter the vocalization nociceptive responses.However, muscimolmicroinjection in the ACC afterMK-801 injection inthe same site significantly decreased the VI. These results corroborateevidence from in vitro electrophysiological studies that have shownthat ACC neurons of mice and rats submitted to persistent pain exhibitan increase in the probability of glutamate release and a depression ininhibitory post synaptic current [39,40]. Interestingly, ACC slices per-fused by bicuculline exhibited an evident increase in action potentials,indicating the existence of GABAergic tonus [8,41]. Considering that ex-citatory amino acids in the ACC are essential during pain mediation[6,7], we have hypothesized that the muscimol microinjected singly inthe ACC was not sufficient to inhibit the pronociceptive effects fromthe glutamate released immediately after noxious stimuli. Nevertheless,the blocking of glutamatergic neurotransmission in the ACC byMK-801led to the antinociceptive effect from muscimol microinjection. If thishypothesis is correct, we may suppose that the GABA system exerts arole in the regulation of ACC neuronal hyperactivity. In physiologicalconditions, this system would appear to reduce the amount of excitatoryresponses to painful stimuli by a tonic action [8,41]. However, the GABAsystem is not capable of blocking the ACC neuronal hyperactivity [42]caused by intense noxious stimuli. Fig. 4 shows a schematic diagram ofthe presumed circuitry in the ACC.
The blockade of GABAergic neurotransmission by bicuculline pro-duced more pronociception than stimulation with DLH. We interpretthis finding based on the previous work demonstrating increased levelsof glutamate in the ACC during noxious stimulation [39,43]. Glutamate
ns are tonically inhibited by GABAergic interneurons; (B) activation of glutamatergic neu-bicuculline produces a hyperactivity of the neurons involved in nociception,which inducesient to overcome the glutamatergic effect on the efferent neurons involved in nociception,-801 was able to show the inhibitory effect of muscimol, and the result is antinociception.; MK-801 = NMDA glutamate receptor antagonist.
release primarily activates ionotropic non-NMDA (AMPA/kainate)receptors, which induce a depolarization in the neuron membrane,activating NMDA receptors by dislodging Mg2+ ions, which physio-logically block the inside of the receptor pore [44,45]. Due to theirhigh permeability to Ca2+, NMDA receptors exhibit excitotoxicityand alternatively may become desensitized and/or internalized as aprotective mechanism against neuronal death [46,47]. Based onthese data, we hypothesize that animals that received DLH microin-jection in the ACC may have inactivated the receptors and preventedthe persistent effect of glutamate in their effector neurons. In con-trast, the animals that received bicuculline microinjection exhibiteda persistent response due to the absence of GABAergic inhibition andthe facilitation of glutamate discharge.
The literature has highlighted different algesimetricmodels to inves-tigate the motivational-affective component of pain [5,18,48]. Ourmodel of vocalization should be understood as a behavioral reaction toemotional content, produced in a state of acute pain, in which theanimal cannot escape from the source of stimulation. In contrast, studieshave often demonstrated the role of the ACC in themodulation of antic-ipatory nociceptive responses aimed at avoidance of potential danger,without changing nociceptive motor responses [6,8]. However, theACC has also been implicated in the modulation of other responses toaversive emotional content, which demonstrates its broad role inmany types of emotional processing [27,49,50]. A study from our labo-ratory has shown that DLH microinjection in the guinea pig ACCdecreases the duration of episodes of the defensive behavior knownas tonic immobility [4]. This behavior is exhibited by some specieswhen in physical contact with a predator [51]. Thus, the ACC alsoseems to influence the behavioral strategy used during an aversivesituation.
In summary, the nociceptive circuitry in the ACC is not exclusivelymediated by glutamatergic neurotransmission via NMDA receptors,although its blockade prevents pronociception induced by DLH. TheGABAergic system appears to modulate ACC under physiologicalconditions. Even the exogenous administration of agonist was un-able to change the magnitude of vocalizations. However, inhibitionof this system with bicuculline exacerbates nociceptive responses,possibly by facilitating discharge of the excitatory ACC [32]. Fur-thermore, we showed the antinociceptive effect of muscimol onNMDA receptors pharmacologically blockaded with MK-801, whichallowed the prevalence of GABAergic neurotransmission during nox-ious stimulation.
Acknowledgments
We would like to thank Mrs. Mariulza Rocha Brentegani andMr. Rubens Fernando de Melo for technical assistance and histologicalprocessing. This work was supported by CAPES, CNPq and FAEPA.
References
[1] Bryant RA, Felmingham K, Whitford TJ, Kemp A, Hughes G, Peduto A, et al. Rostralanterior cingulate volume predicts treatment response to cognitive-behaviouraltherapy for posttraumatic stress disorder. J Psychiatry Neurosci 2008;33:142–6.
[2] Chua P, KramsM, Toni I, PassinghamR, Dolan R. A functional anatomy of anticipatoryanxiety. Neuroimage 1999;9:563–71.
[3] Frankland PW, Bontempi B, Talton LE, Kaczmarek L, Silva AJ. The involvementof the anterior cingulate cortex in remote contextual fear memory. Science2004;304(5672):881–3.
[4] Coutinho MR, Menescal-de-Oliveira L. Role of homocysteic acid in the guinea pig(Cavia porcellus) anterior cingulate cortex in tonic immobility and the influence ofNMDA receptors on the dorsal PAG. Behav Brain Res 2010;208:237–42.
[5] Johansen JP, Fields HL, Manning BH. The affective component of pain in rodents:direct evidence for a contribution of the anterior cingulate cortex. Proc Natl AcadSci U S A 2001;98:8077–82.
[6] Johansen JP, Fields HL. Glutamatergic activation of anterior cingulated cortex pro-duces an aversive teaching signal. Nat Neurosci 2004;7:398–403.
[7] Lei LG, Sun S, Gao YJ, Zhao ZQ, Zhang YQ. NMDA receptors in the anterior cingulatecortex mediate pain-related aversion. Exp Neurol 2004;189:413–21.
[8] Wang H, RenWH, Zhang YQ, Zhao ZQ. GABAergic disinhibition facilitates polysynap-tic excitatory transmission in rat anterior cingulate cortex. Biochem Biophys ResCommun 2005;338:1634–9.
[9] Zhang Z, Séguéla P. Metabotropic induction of persistent activity in layers II/III of an-terior cingulated cortex. Cereb Cortex 2010;20:2948–57.
[10] BozkurtA, Zilles K, Schleicher A, Kamper L, Arigita ES, UylingsHB.Distributions of trans-mitter receptors in the macaque cingulate cortex. Neuroimage 2005;25:219–29.
[11] LaGraize SC, Fuchs PN. GABAa but not GABAb receptors in the rostral anterior cingu-late cortex selectively modulate pain-induced escape/avoidance. Behav Exp Neurol2007;204:182–94.
[12] Menescal-de-Oliveira L, Lico MC. Pain modulation in the adrenergically stimulatedarea postrema in the alert guinea pig. Physiol Behav 1977;19:359–64.
[13] Menescal-de-Oliveira L, HoffmannA. The parabrachial region as a possible regionmod-ulating simultaneously pain and tonic immobility. Behav Brain Res 1993;56:127–32.
[14] Favaroni-Mendes LA, Menescal-de-Oliveira L. Role of cholinergic, opioidergic andGABAergic neurotransmission of the dorsal hippocampus in the modulation ofnociception in guinea pigs. Life Sci 2008;83:644–50.
[15] da Silva LFS, Coutinho MR, Menescal-de-Oliveira L. Opioidergic and GABAergicmechanisms in the rostral ventromedial medullamodulate the nociceptive responseof vocalization in guinea pigs. Brain Res Bull 2010;82:177–83.
[16] da Silva LFS, Coimbra NC, Menescal-de-Oliveira L. rostral ventromedial medullamodulates nociception and tonic immobility behavior through connections withthe A7 catecholaminergic region. Behav Brain Res 2012;233:422–7.
[17] Jürgens U, Pratt R. Role of the periaqueductal grey in vocal expression of emotion.Brain Res 1979;167:367–78.
[18] Borszcz GS. Increases in vocalization and motor reflex thresholds are influenced bythe site of morphine microinjection: comparisons following administration intothe periaqueductal gray, ventral medulla, and spinal subarachnoid space. BehavNeurosci 1995;109:502–22.
[19] Sanchéz C. Stress-induced vocalization in adult animals. A valid model of anxiety?Eur J Pharmacol 2003;463:133–43.
[20] Bandler R, Shipley MT. Columnar organization in the midbrain periaqueductal gray:modules for emotional expression? Trends Neurosci 1994;17:379–89.
[21] Borszcz GS. Pavlovian conditional vocalizations of the rat: a model system for ana-lyzing the fear of pain. Behav Neurosci 1995;109:648–62.
[22] Carrive P. The periaqueductal gray and defensive behavior: functional representa-tion and neuronal organization. Behav Brain Res 1993;58:27–47.
[23] Jürgens U. The neural control of vocalization in mammals: a review. J Voice2009;23:1–10.
[24] Zimmermann M. Ethical guidelines for investigations of experimental pain in con-scious animals. Pain 1983;16:109–10.
[25] Rössner W. Stereotaktischer hirnatlas vom meerschweinchen. Munich: PallasVerlag; 1965.
[26] Cao H, Gao YJ, Ren WH, Li TT, Duan KZ, Cui YH, et al. Activation of extracellularsignal-regulated kinase in the anterior cingulate cortex contributes to the inductionand expression of affective pain. J Neurosci 2009;29:3307–21.
[27] Donahue RR, LaGraize SC, Fuchs PN. Electrolytic lesion of the anterior cingulatecortex decreases inflammatory, but not neuropathic nociceptive behavior in rats.Brain Res 2001;897:131–8.
[28] Lee DE, Kim SJ, Zhuo M. Comparison of behavioral responses to noxious cold andheat in mice. Brain Res 1999;845:117–21.
[29] Kang SJ, LiuMG, Chen T, Ko HG, Baek GC, Lee HR, et al. Plasticity of metabotropic glu-tamate receptor-dependent long-term depression in the anterior cingulate cortexafter amputation. J Neurosci 2012;32:11318–29.
[30] Wu LJ, Zhao MG, Toyoda H, Ko SW, Zhuo M. Kainate receptor-mediated synaptictransmission in the adult anterior cingulate cortex. J Neurosci 2005;94:1805–13.
[31] Li TT, RenWH, Xio X, Nan J, Cheng LZ, Zhang XH, et al. NMDANR2A and NR2B recep-tors in the rostral anterior cingulate cortex contribute to pain-related aversion inmale rats. Pain 2009;146:183–93.
[32] Wei F, Li P, Zhuo M. Loss of synaptic depression in mammalian anterior cingulatecortex after amputation. J Neurosci 1999;19:9346–54.
[33] Calejesan AA, Kim SJ, Zhuo M. Descending facilitatory modulation of a behavioralnociceptive response by stimulation in the adult rat anterior cingulate cortex. Eur JPain 2000;4:83–96.
[34] López-Avila A, Coffeen U, Ortega-Legaspi JM, Del Angel R, Pellicer F. Dopamine andNMDA systems modulae long-term nociception in rat anterior cingulate córtex.Pain 2004;111:136–43.
[35] Harte SE, Spuz CA, Borszcz GS. Functional interaction between medial thalamus androstral anterior cingulate cortex in the suppression of pain affect. Neuroscience2011;172:460–73.
[37] Kardos J, Blaskó G, Kerekes P, Kovács I, Simonyi M. Inhibition of [3H] gaba binding torat brain synaptic membranes by bicuculline related alkaloids. Biochem Pharmacol1984;33:3537–45.
[38] Chebib M, Johnston GAR. The ‘ABC’ of GABA receptors: a brief review. Clin ExpPharmacol Physiol 1999;26:937–40.
[39] Xu H, Wu LJ, Wang H, Zhang X, Vadakkan KI, Kim SS, et al. Presynaptic and postsyn-aptic amplifications of neuropathic pain in the anterior cingulate cortex. J Neurosci2008;28:7445–53.
[40] GongKR, Cao FL, He Y, Gao CY,WangDD, Li H, et al. Enhanced excitatory and reducedinhibitory synaptic transmission contribute to persistent pain-induced neuronalhyper-responsiveness in anterior cingulate cortex. Neuroscience 2010;171:1314–25.
[41] Higashi H, Tanaka E, Nishi S. Synaptic responses of guinea pig cingulate cortical neu-rons in vitro. J Neurophysiol 1991;65:822–33.
[42] Zhuo M. Central inhibition and placebo analgesia. Mol Pain 2005;21:1–4.
14 J. Zugaib et al. / Physiology & Behavior 126 (2014) 8–14
[43] Zhang L, Zhang Y, Zhao ZQ. Anterior cingulate cortex contributes to the descendingfacilitatory modulation of pain via dorsal reticular nucleus. Eur J Neurosci2005;22:1141–8.
[44] Cohen S, Greenberg ME. Communication between the synapse and the nucleusin neuronal development, plasticity, and disease. Annu Rev Cell Dev Biol2008;24:183–209.
[45] Rebola N, Srikumar BN, Mulle C. Activity-dependent synaptic plasticity of NMDA re-ceptors. J Physiol 2010;588:93–9.
[46] Hara MR, Snyder SH. Cell signaling and neuronal death. Annu Rev Pharmacol Toxicol2007;47:117–41.
[47] Petralia RS. Distribution of extrasynaptic NMDA receptors on neurons. Sci World J2012;2012:1–11.
[48] Qu C, King T, Okun A, Lai J, Fields HL, Porreca F. Lesion of the rostral anterior cingu-late cortex eliminates the aversiveness of spontaneous neuropathic pain followingpartial or complete axotomy. Pain 2011;152:1641–8.
[49] Ortega-Legaspi JM,Avila AL, CoffeenU,Angel R, Pellicer F. Scopolamine into the anteriorcingulate cortex diminishes nociception in a neuropathic pain model in the rat: an in-terruption of “nociception-related memory acquisition”? Eur J Pain 2003;7:425–9.
[50] Zubieta JK, Ketter TA, Bueller JA, Xu Y, Kilbourn MR, Young EA, et al. Regulation ofhuman affective responses by anterior cingulate and limbic mu-opioid neurotrans-mission. Arch Gen Psychiatry 2003;60:1145–53.
[51] Leite-Panissi CR, Rodrigues CL, Brentegani MR, Menescal-de-Oliveira L. Endogenousopiate analgesia induced by tonic immobility in guinea pigs. Braz J Med Biol Res2000;34:245–50.