Glutathione-S-transferase activity and its induction in several strains of l~'ibolium castaneum Ephraim Cohen Department of Entomology, The Hebrew University of Jerusalem, Faculty of Agriculture, Rehovot 76 100, Israel Keywords: Glutathione-S-transferase, hexachlorocyclohexane isomers, induction, 3-methylcholanthrene, pentobarbital, polychlorinated biphenyl, trans-stilbene oxide, Tribolium castaneum Abstract Similar levels of glutathione-S-transferase (GST) specific activities were detected in CSbb, Kano and CS + + strains of the flour beetle Tribo#um castaneum (Herbst) (Tenebrionidae, Coleoptera). The GST ac- tivity in the CTC-12 multi-resistant line was significantly higher compared with the above strains. The larval and adult stages are characterized by relatively high levels of GST activity. The enzyme activity was consider- ably reduced in pharate-pupae and pupae, and no activity was detected in eggs. The extent of inducibility of GST by various drugs and toxicants was dependent on the insect strain and physiological stage, and on the type of inducer. In general, the larval GST enzymes were strongly stimulated by pentobarbital as compared with the adult ones. Similar pattern of induction was observed with 3-methylcholanthrene and trans-stilbene oxide. The highest stimulation (up to 3.6 fold) of the transferase was detected in CTC-12 larvae treated with a polychlorinated biphenyl analog. The ~ and f hexachlorocyclohex- ane isomers are potent inducers of GST in the larval stage whereas the 6 isomer is far less effective. Introduction Glutathione-S-transferases (GST) are a family of widespread, soluble and non-specific enzymes known to catalyze the conjugation between reduced glutathione (GSH) and a variety of struc- turally diversed electrophilic substrates (Chas- seaud, 1973; Jakoby & Habig, 1980). Conjugated xenobiotics are inactivated and can be further transformed to yield mercapturic acids (Habig et al., 1974). The GST system has been detected in various in- sects and together with hydrolytic and oxidative systems, was implicated in conferring resistance in insects to insecticides (Oppenoorth & Wellings, 1976). GST as an important system of in vitro detoxication has been demonstrated for lindane (Tanaka et al., 1981) and for several or- ganophosphorus compounds (Motoyama & Dau- terman, 1980). Resembling other drug- Entomol. exp. appl. 41: 39-44, 1986 9 Dr W. Junk Publishers, Dordrecht - Printed in the Netherlands metabolizing enzymes such'as mixed function oxi- dases (MFO), esterases and epoxide hydrases, the GST enzyme is inducible in mammalian (Hales & Niems, 1977; Kuo et al., 1982; Hammock & Ota, 1983) and in insect species (Terriere, 1984) by a vari- ety of compounds. Phenobarbital is by far the com- monly used drug for demonstrating GST induction in insects (Ottea & Plapp, 1981; Hayaoka & Dauter- man, 1982). Secondary plant compounds (Yu, 1982) and the insecticides DDT and dieldrin (Hayaoka & Dauterman, 1982) were also reported to induce GST activity in insects. I examined and compared the ability of hexa- chlorocyclohexane (HCH) isomers, 3-methylcholan- threne (MC), trans-stilbene oxide (STO) and a poly- chlorinated biphenyl (PCB) analog, to induce GST activity in larvae and adults of various strains of the flour beetle Tribolium castaneum.
Transcript
Glutathione-S-transferase activity and its induction in several strains of l~'ibolium castaneum
Ephraim Cohen Department of Entomology, The Hebrew University of Jerusalem, Faculty of Agriculture, Rehovot 76 100, Israel
Similar levels of glutathione-S-transferase (GST) specific activities were detected in CSbb, Kano and CS + + strains of the flour beetle Tribo#um castaneum (Herbst) (Tenebrionidae, Coleoptera). The GST ac- tivity in the CTC-12 multi-resistant line was significantly higher compared with the above strains. The larval and adult stages are characterized by relatively high levels of GST activity. The enzyme activity was consider- ably reduced in pharate-pupae and pupae, and no activity was detected in eggs.
The extent of inducibility of GST by various drugs and toxicants was dependent on the insect strain and physiological stage, and on the type of inducer. In general, the larval GST enzymes were strongly stimulated by pentobarbital as compared with the adult ones. Similar pattern of induction was observed with 3-methylcholanthrene and trans-stilbene oxide. The highest stimulation (up to 3.6 fold) of the transferase was detected in CTC-12 larvae treated with a polychlorinated biphenyl analog. The ~ and f hexachlorocyclohex- ane isomers are potent inducers of GST in the larval stage whereas the 6 isomer is far less effective.
Introduction
Glutathione-S-transferases (GST) are a family of widespread, soluble and non-specific enzymes known to catalyze the conjugation between reduced glutathione (GSH) and a variety of struc- turally diversed electrophilic substrates (Chas- seaud, 1973; Jakoby & Habig, 1980). Conjugated xenobiotics are inactivated and can be further transformed to yield mercapturic acids (Habig et al., 1974).
The GST system has been detected in various in- sects and together with hydrolytic and oxidative systems, was implicated in conferring resistance in insects to insecticides (Oppenoorth & Wellings, 1976). GST as an important system of in vitro detoxication has been demonstrated for lindane (Tanaka et al., 1981) and for several or- ganophosphorus compounds (Motoyama & Dau- terman, 1980). Resembling other drug-
Entomol. exp. appl. 41: 39-44 , 1986 �9 Dr W. Junk Publishers, Dordrecht - Printed in the Netherlands
metabolizing enzymes such'as mixed function oxi- dases (MFO), esterases and epoxide hydrases, the GST enzyme is inducible in mammalian (Hales & Niems, 1977; Kuo et al., 1982; Hammock & Ota, 1983) and in insect species (Terriere, 1984) by a vari- ety of compounds. Phenobarbital is by far the com- monly used drug for demonstrating GST induction in insects (Ottea & Plapp, 1981; Hayaoka & Dauter- man, 1982). Secondary plant compounds (Yu, 1982) and the insecticides DDT and dieldrin (Hayaoka & Dauterman, 1982) were also reported to induce GST activity in insects.
I examined and compared the ability of hexa- chlorocyclohexane (HCH) isomers, 3-methylcholan- threne (MC), trans-stilbene oxide (STO) and a poly- chlorinated biphenyl (PCB) analog, to induce GST activity in larvae and adults of various strains of the flour beetle Tribolium castaneum.
40
Materials and methods
Insects. Four strains of the flour beetle were used. (1) CS+ + (Berkeley), a wild-type strain, and (2) CSbb, a black mutant, susceptible to insecticides of various groups, came from United States; (3) Kano, specifically resistant to malathion, and (4) CTC-12, a multi-resistant strain, were supplied from stock of Tribolium strains in the Department of Zoology, Tel-Aviv University, Israel. The insects were raised at 30~ on wheat flour supplemented with 5% brewers' yeast. Eggs were collected from stock cul- tures every 2 - 3 days and transferred to a fresh diet, to ensure continuous supply of insects at a desired chronological age and physiological stage.
Chemicals. 2,4-Dichloro-l-nitrobenzene (DCNB) and trans-stilbene oxide (STO) were obtained from Aldrich (Milwaukee, Wisc., U.S.A.). 1-Chloro-2,4- dinitrobenzene (CDNB), pentobarbital (sodium) from Merck (Darmstadt, Germany) and Serva (Heidelberg, Germany). Reduced glutathione (GSH) and 3-methylcholanthrene (MC) came from Sigma (St. Louis, Mo., U.S.A.).
Extraction o f GST enzyme. Insects were removed from cultures, cleaned and transferred to 5 volumes of 100 mM potassium (K)-phosphate buffer pH 7.4 containing 1.15% KC1 and 250 mM sucrose. After homogenization by Ultra Turrax, the mixture was filtered through four layers of gauze and cen- trifuged for 30 rain at 12000xg. The supernatant was filtered through glass-wool and subsequently centrifuged for 60 rain at 100000• The cytosol was used as the GST source. Protein levels were measured by the method of Bradford (1976) using bovine serum albumin as a standard.
GST assay. The enzymatic activity was determined spectrophotometrically by essentially following the method described by Habig et al. (1974). The reac- tion mixture in 1.0 ml volume consisted of 50 mM K-phosphate buffer pH 6.5 containing 0.58% KCI, 5 mM GSH and 1 mM of either CDNB or DCNB as electrophilic substrates. The enzymatic reaction at 25 ~ started by adding the enzyme preparation (20 ~1), and the absorbance of the CDNB and DCNB conjugates at 340 nm and 345 nm, respec- tively, was continuously monitored with Uvicon-80 double-beam spectrophotometer. The GST specific
activity was expressed as nmol conjugated substrate per min per mg protein, using the corresponding extinction coefficients of 9.6 cm lmM 1 and 10.0 c m - l m M -a for CDNB and DCNB.
Induction studies. The compounds tested as in- ducers of T. castaneurn GST were thoroughly mixed (W/W) with the insect diet at concentrations ranging from 0.01-0.5%. The dietary levels of the inducers were non-toxic to the insects. Fifty eggs were transferred from stock cultures to 2.0 g diet and either larvae or adults at specified age were re- moved for extraction of the GST enzyme.
Results
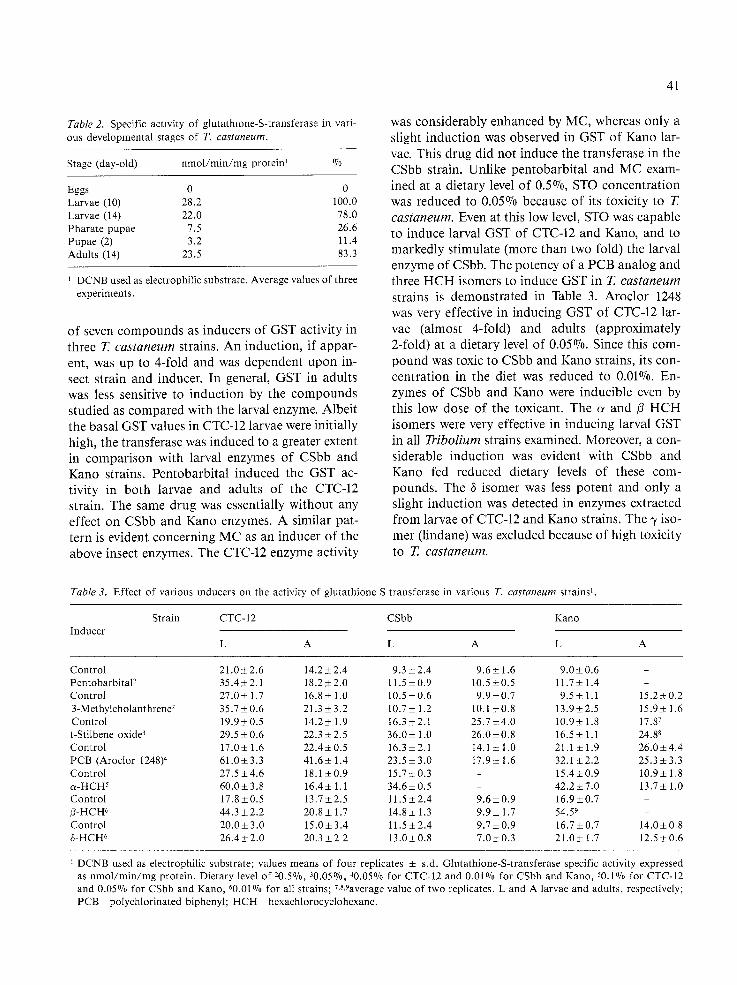
Table 1 summarizes the specific activities of GST in the cytosolic fractions obtained from larvae of several T. castaneum strains. The values of GST specific activities in the multi-resistant CTC-12 were higher than those of the three other strains, using either DCNB or CDNB as substrates. No ap- preciable differences among CSbb, Kano and CS+ + strains with respect to their GST activity were observed. The GST specific activities deter- mined at various physiological stages of T. castane- um (CTC-12) are presented in Table 2. Larvae and adults are characterized by high GST activity. The highest GST level was measured in 10-day-old lar- vae. No detectable GST activity could be found in eggs, and the enzyme activity dropped considerably in pharate-pupae and pupae by 61% and 89%, respectively. Apparently GST plays an important physiological role in mobile, feeding stages of the flour beetle.
Table 3 demonstrates the effects of dietary levels
Table 1. Glutathione-S-transferase activity in larvae of various T. eastaneum strains.
L DCNB used as electrophilic substrate. Average values of three
experiments.
of seven compounds as inducers of GST activity in three T. castaneum strains. An induction, if appar- ent, was up to 4-fold and was dependent upon in- sect strain and inducer. In general, GST in adults was less sensitive to induction by the compounds studied as compared with the larval enzyme. Albeit the basal GST values in CTC-12 larvae were initially high, the transferase was induced to a greater extent in comparison with larval enzymes of CSbb and Kano strains. Pentobarbital induced the GST ac- tivity in both larvae and adults of the CTC-12 strain. The same drug was essentially without any effect on CSbb and Kano enzymes. A similar pat- tern is evident concerning MC as an inducer of the above insect enzymes. The CTC-12 enzyme activity
41
was considerably enhanced by MC, whereas only a slight induction was observed in GST of Kano lar- vae. This drug did not induce the transferase in the CSbb strain. Unlike pentobarbital and MC exam- ined at a dietary level of 0.5%, STO concentration was reduced to 0.05% because of its toxicity to T. castaneum. Even at this low level, STO was capable to induce larval GST of CTC-12 and Kano, and to markedly stimulate (more than two fold) the larval enzyme of CSbb. The potency of a PCB analog and three H C H isomers to induce GST in T. castaneum
strains is demonstrated in Table 3. Aroclor 1248 was very effective in inducing GST of CTC-12 lar- vae (almost 4-fold) and adults (approximately 2-fold) at a dietary level of 0.05%. Since this com- pound was toxic to CSbb and Kano strains, its con- centration in the diet was reduced to 0.01%. En- zymes of CSbb and Kano were inducible even by this low dose of the toxicant. The o~ and/3 H C H isomers were very effective in inducing larval GST in all Tribolium strains examined. Moreover, a con- siderable induction was evident with CSbb and Kano fed reduced dietary levels of these com- pounds. The 6 isomer was less potent and only a slight induction was detected in enzymes extracted from larvae of CTC-12 and Kano strains. The -y iso- mer (lindane) was excluded because of high toxicity to T. castaneum.
Table 3. Effect of various reducers on the activity of glutathione-S-transferase in various T. castaneum strains ~.
DCNB used as electrophitic substrate; values means of four replicates _+ s.d. Glutathione-S-transferase specific activity expressed as n m o l / m i n / m g protein. Dietary level of 20.5%, 30.05~ 40.05% for CTC-12 and 0.01% for CSbb and Kano, 50.1% for CTC-12 and 0.05% for CSbb and Kano, 60.01~ for all strains; 7,8,9average value of two replicates. L and A larvae and adults, respectively; P C B - p o l y c h l o r i n a t e d biphenyl; HCH hexachlorocyclohexane.
42
Discussion
The ontogenetic pattern of the cytosolic GST ac- tivity is similar to the observed activities of fast migrating esterases (Cohen et al., 1977) and epoxide hydrase (Cohen, 1981) in T. castaneum. The high activity of these detoxication enzymes detected in the mobile, feeding stages (larvae and adults) might be related to the capacity for biotransforming xenobiotics present in ingested food or absorbed from the environment. Resting stages apparently do not require high levels of detoxifying enzymes including GST.
It is believed that GST confers resistance in con- junction with other detoxicating enzyme systems (Terriere, 1984). The T. castaneurn CTC-12 strain is characterized by resistance to diverse groups of in- secticides (Champ & Campbell-Brown, 1970). The polygenic nature of this resistance for malathion has been shown by Wool et al. (1982). Studies on various detoxifying enzyme systems have revealed that in comparison to other T. castaneurn strains, CTC-12 is characterized by high levels of microsomal oxidases (Cohen, 1982), enhanced ac- tivities of epoxide hydrase (Cohen, 1981) and GST (Table 1). Conceivably, the elevated level of activity of the above enzymes is closely associated with re- sistance of CTC-12 to a large number of insecti- cides. The high and specific resistance of Kano strain to malathion (Dyte & Rowland, 1968) might be related in part to higher levels of carboxylester- ase activity (Wool et al., 1982; Price, 1984). The specific activity of GST in this strain is similar to that detected in the susceptible CSbb line, and its epoxide hydrase activity is below that of CTC-12 (Cohen, 1981).
Various microsomal and cytosolic enzyme sys- tems associated with biotransformation of xenobi- otics including pesticides, are induced by a variety of drugs and toxicants. It has been demonstrated that insects treated with xenobiotics including secondary plant chemicals, were less sensitive to in- secticides (Yu, 1982). Phenobarbital is a potent well known inducer of inactivating enzyme systems both in insects and in mammals. Cytochromes b5 and P-450 (Cohen, 1981) and GST (Table 3) in T. castaneurn were induced by pentobarbital. The magnitude of GST induction in Tribolium was de- pendent on the insect strain, physiological stage and on type of the examined inducer. Hayaoka &
Dauterman (1982) have presented similar observa- tions regarding the stimulation of GST activity by phenobarbital in various housefly strains. The ex- tent of GST induction in Triboliurn did not exceed 4-fold and in this regard was similar to the housefly enzyme induced by phenobarbital. Although the GST basal specific activity in CTC-12 was rather high, it could be induced even to higher levels than the strains with initially lower activity. The oppo- site was reported for GST induction in houseflies (Hayaoka & Dauterman, 1982). It appears that al- most the full capacity of the enzyme activity had been already expressed in the resistant houseflies.
MC and STO were able to induce GST activity in mammalian systems (Baars et al., 1978; Seidegfird et al., 1979; Kuo et al., 1981). Also the inducibility of GST mRNA in rat liver by MC was reported (Pickett et al., 1982). The above compounds were effective as inducers of Tribolium GST, particularly in treated CTC-12 larvae (Table 3). STO was claimed to selectively induce GST and epoxide hy- drase enzymes in mammals (Kuo et al., 1982). Oth- er reports did not agree with the above claim in- dicating that also the MFO system was stimulated by this inducer (Seideggtrd et al., 1979; Meijer et al., 1982).
PCBs which are widespread pollutants of great hazard to the environment are regarded as potent inducers of microsomal oxidative enzymes in the insect Spodoptera eridania (Anderson, 1978) and in rat liver (Ahotupa, 1981). It has been demonstrated that the extent of inducibility was dependent on number and position of chlorines in the molecule. The PCB analog tested as a GST inducer in Triboli- urn was highly effective, mainly in stimulating CTC-12 larval enzyme. The potency of HCH isomers to induce GST was studied in a mam- malian system (Kraus et al., 1981). The c~ but not the/3 isomer was able to enhance GST levels in rat liver. The increase of GST activity (50-60%) was less than that observed in Tribolium strains (Ta- ble 3). The o~ and/3 isomers were almost equipotent in inducing GST activity in Tribolium. The 6 iso- mer was less active, indicating differences in magni- tude of induction among the HCH isomers.
The observations that diverse, non-specific detoxifying enzyme systems such as MFO, epoxide hydrase and GST in Triboliurn castaneum are indu- cible by a single compound (pentobarbital) and perhaps the high levels of these systems in the
multi-resistant CTC-12 strain, lend support to the notion (Ottea & Plapp, 1981) that they might be subjected to a common control mechanism. It is of interest that the genetic code in the housefly for GST and cytochrome P-450 proteins are located on the same chromosome (Motoyama et al., 1977). The inducibility of GST in Tribolium by diverse compounds adds to the accumulated vast body of information describing the induction phenomenon. However, the exact mechanism of induction at the cellular and biochemical levels is still poorly under- stood.
Acknowledgements
I thank Mrs. Batia Kaminsky for technical help and Dr. David Wool, Department of Zoology, Tel- Aviv University for providing the T. castaneum strains. Supported by a grant from the Israeli Academy of Sciences and Humanities.
R~sum~
L'activitd glutathione-S-transferase et son induction chez plusieurs lignges de Tribolium castaneum
Des niveaux voisins d'activitds sp6cifiques glutathione-S-transf6rase (GST) ont 6t6 mis en ~vi- dence chez les lign6es CSbb, Kano et CS+ + de Tribolium castaneum. Uactivit6 GST est sig- nificativement plus 616v6e chez la lign6e mul- tir6sistante CTC-12. Les stades larvaires et im- aginaux sont caractdris6s par des niveaux relativement 616vds d'activit6 GST. Uactivit6 en- zymatique 6tant consid6rablement r6duite chez les nymphes et les 'prd-imagos'; aucune activit6 n'a 6t6 ddcel6e dans les oeufs.
Les possibilitds d'induction de GST par diff6rentes, drogues et substances toxiques d6pend de l'6tat physiologique, de la lign6e de l'insecte et de la nature de l'inducteur. Gdn6ralement le pentobar- bital stimule plus fortement les enzymes larvaires qu'imaginales. Uinduction par 3-m6thylcholanthrbne et l'oxyde trans-stilbbne pr6sentait les m~mes traits. La stimulation de la transfdrase la plus 61evde (jusqu'/a 3,6 lois) a 6t6 dd- cel6e chez les les larves CTC-12 trait6es avec un ana- logue du biph6nyl polychlor6. Les isom~res c~ et
43
sont des inducteurs puissants de GST chez les larves tandis que l'isom6re A est de loin moins ef- ficace.
References
Ahotupa, M., 1981. Enhancement of epoxlde-metabolizing en- zyme activities by pure PCB isomers. Biochem. Pharmacol. 30: 1866-1869.
Anderson, R. S., 1978. Hydrocarbon hydroxylase induction in an insect Spodoptera erMama (Cramer) by polychlormated biphenyls (PCBs) Comp. Biochem. Physiol. 60C: 51-55.
Baars, A. J., M. Jansen & D. D. Breimer, 1978. The influence of phenobarbital, 3-methylcholanthrene and 2,3,7,8-tetrachloro- dibenzo-p-dioxin on glutathione-S-transferase activity of rat liver cytosol. Blochem. Pharmacol. 27: 2487-2494.
Bradford, M. M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein, utilizing the principle of protein-dye binding. Analyt. Biochem. 72: 248- 254.
Champ, B. R. & M. Campbell-Brown, 1970. Insecticide resis- tance in Australian Tribohum castaneum. II. Malathion resis- tance in Eastern Australia. J. Stored Prod. Res. 6: 111-131.
Chasseaud, L. E, 1973. The nature and distribunon of enzymes catalyzing the conjugation of glutathione with foreign com- pounds. Drug Metab. Rev. 2: 185-220.
Cohen, E., 1981. Epoxide hydrase activity in the flour beetle Tribolium castaneum (Coleoptera, Tenebriomdae). Comp. Biochem. Physiol. 69B: 29-34.
Cohen, E., 1982. Studies on several microsomal enzymes in two strains of Trtbolium castaneum (Tenebrionidae, Coleoptera). Comp. Biochem. Physiol. 71C: 123-126.
Cohen, E., E. Sverdlov & D. Wool, 1977. Expression of esterases during ontogenesis of the flour beetle Tribolium castaneum (Tenebrionidae: Coleoptera). Biochem. Genet. 15: 253-264.
Dyte, C. E. & D. G. Rowlands, 1968. The metabolism and syner- gism of malathion in resistant and susceptible strains of Tribolium castaneum. J. Stored Prod. Res. 4: 157-173.
HaNg, W. H., M. J. Pabst & W. B. Jacoby, 1974. Glutathione-S- transferase, the first enzymatic step in mercapturic acid for- mation. J. Biol. Chem. 249: 7130-7139.
Hales, B. E & A. H. Niems, 1977. Induction of rat hepatic glutathione-S-transferase B by phenobarbital and 3-methyl- cholanthrene. Biochem. Pharmacol. 26: 555-556.
Hammock, B. D. & K. Ota, 1983. Differential induction of cyto- solic epoxide hydrolase, microsomal epoxide hydrolase, and glutathione-S-transferase activities. Toxicol. Appl. Phar- macol. 71: 254-265.
Hayaoka, T. & W.C. Dauterman, 1982. Inducuon of glutathione-S-transferase by phenobarbital and pesticides in various house fly strains and its effect on toxicity. Pestic. Bi- ochem. Physiol. 17: 113-119.
Jakoby, W. B. & W. H. Habig, 1980. Glutathione transferases. In: W. B. Jakoby (ed.), Enzymatic Basis of Detoxification, Vol. II. Academic Press, New York: pp. 63-94.
Kno, C. H., J. B. Hook & J. Bernstein, 1981. Induction of drug- metabolizing enzymes and toxicity of trans-stilbene oxide in rat and kidney. Toxicology 22: 149-160.
44
Kraus, P., B. Gross & H. D. Kloft, 1981. The elevation of rat liver glutathione-S-trans ferase activity by alpha- hexachlorocyclohexane. Biochem. Pharmacol. 30: 355-361.
Meijer, J., A. Astrom, J. W. DePierre, F. P. Guengerich & L. Ernster, 1982. Characterization of the microsomal cytochrome P-450 species induced by trans-stilbene oxide. Bi- ochem. Pharmacol. 31: 3907-3916.
Motoyama, N. & W.C. Dauterman, 1980. Glutathione-S- transferases: Their role in metabolism of organophosphorus insecticides. Reviews in Biochemical Toxicology. Vol. II: 49- 69.
Motoyama, N., W. C. Dauterman & F. W. Plapp, Jr., 1977. Genetic studies on glutathione-dependent reactions in resis- tant strains of the housefly, Musca domestica L. Pestic. Bi- ochem. Physiol. 7: 443-450.
Oppenoorth, F. J. & W. Wellings, 1976. Biochemistry and phys- iology of resistance. In: C. E Wilkinson (ed.), Insecticide Bi- ochemistry and Physiology. Plenum Press, New York: 507-551.
Ottea, J. A. & E W. Ptapp, Jr., 1981. Induction of glutathione- S-aryl transferase by phenobarbital in the house fly. Pestic. Biochem. Physiol. 15: 10-13.
Pickett, C.B., C.A. Telakowski-Hopkins, A.M. Donohue, A. Y. H. Lu & B. F. Hales, 1982. Differential induction of rat hepatic cytochrome P-448 and glutathione-S-transferase B
Price, N. R., 1984. Carboxyesterase degradation of malathion m vitro by susceptible and resistant strains of Tribohum castaneum (Herbst) (Coleoptera, Tenebrionidae). Comp. Bi- ochem. Physiol. 77C: 95-98.
Seideg~rd, J., R. Morgenstern, J.W. DePierre & L. Ernster, 1979. Trans-stilbene oxide: A new type of inducer of drug- metabolizing enzymes. Biochim. Biophys. Acta 586: 10-21.
Tanaka, K., M. Nakajima & N. Kurihara, 1981. The mechanism of resistance to lindane and hexadeuterated lindane in the third Yumenoshima strain of house fly. Pestic. Biochem. Physiol. 16: 149-157.
Terriere, L. C., 1984. Induction of detoxication enzymes in in- sects. Ann. Rev. Entomol. 29: 71-88.
Wool, D., S. Noiman, O. Manheim & E. Cohen, 1982. Malathion resistance in Trtbohum strains and their hybrids: Inheritance patterns and possible enzymatic mechanisms (Coleoptera, Tenebrionidae). Biochem. Genet. 20: 621-636.
Yu, S.J., 1982. Host plant induction of glutathione-S- transferase in the fall armyworm. Pestic. Biochem. Physiol. 18: 101-106.