,\ECOLOGICAL AND TAXONOMIC STUDIES OF '1'IE RUSSULACEAE AND OTIER ECTOMYCORRHIZAL BASIDIOMYCETE} IN TIE ‘ HIGIPELEVATION FORESTS OF TIE SOUTIERN APPALACHIANS bv ‘ Gerald F. Bills dissertation submitted to the Faculty of the — ‘ Virginia Polytechnic Institute and State University in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY in . - Botany APPROVED: ‘ 4 ” 6 · //5* ¤» /4//./ß’”)., '/’ * ’ ’— \·—L« Orson K. Miller, Jr. , Chairman Khidir W. Hilu 6 . A rx · e. 2 V Ä' M/u-} Golde I. Häzman W. Carter Jvnson I I 1Robert A. Peterson Jzy Stiges K May 24, 1985 Blackburg, Virginia
Transcript
,\ECOLOGICAL AND TAXONOMIC STUDIES OF '1'IE RUSSULACEAE
AND OTIER ECTOMYCORRHIZAL BASIDIOMYCETE} IN TIE ‘
HIGIPELEVATION FORESTS OF TIE SOUTIERN APPALACHIANS
bv‘ Gerald F. Bills
dissertation submitted to the Faculty of the —
‘ Virginia Polytechnic Institute and State University
in partial fulfillment of the requirements for the degree of
DOCTOR OF PHILOSOPHY
in .
- Botany
APPROVED: ‘
4 ” 6 ·//5*
¤» /4//./ß’”)., '/’*
’ ’— \·—L«
Orson K. Miller, Jr. , Chairman Khidir W. Hilu
6 . A rx ·
e. 2 VÄ' M/u-}
Golde I. Häzman W. Carter JvnsonI
I
1RobertA. Peterson Jzy Stiges K
May 24, 1985
Blackburg, Virginia
ECOLOGICAL AND TAXONOMIC STUDIES OF THE RUSSULACEAE
AND OTHR ECTOMYCORRHIZAL BASIDIOMYCETES IN THEHIGH-ELEVATION FORESTS OF TH SOUTHERN APPALACHIANS
éä n by
Gerald F. Bills
Orson K. Miller, Jr., Chairman
Botany
(ABSTRACT)A
Temperate and boreal fungal floras indicate that species of the
Russulaceae (the genera Rgggglg and Lggtßgjgg) are among the dominant
ectomycorrhizal fungi in forest communities. The contribution of the
Russulaceae to the communities of ectomycorrhizal Basidiomycetes fruiting
in red spruce and adjacent northern hardwood forests in.West Virginia was
evaluated and compared with other ectomycorrhizal Basidiomycetes
occupying the same habitats. The Russulaceae exhibited the greatest
species diversity of any family of ectomycorrhizal fungi fruiting in the
stands studied (44 X of the species in spruce, 39 X of the species in
hardwoods). Species of Lagtagigg and Ryggylg were among the most
productive in both forests.
Species diversity, productivity, and fruiting phenology of all
ectomycorrhizal Basidiomycetes were compared between red spruce and
northern hardwood stands for a 3-year period. Sporocarp numbers and
sporocarp frequency in 384 four mz quadrats in each forest type was used
to estimate productivity. Species richness was greater in hardwoods (36
species) than in spruce (27 species). Nine species were common to both
forests. Most productivity was coneentrated in a few species, while most
species were rare. Speeies·area curves were constructed for both forests.
Fungal species and tree species eomposition in both forests were compared
by principal component analysis.
Fungi in spruce forests were more produetive than in hardwood forests.
Productivity was highly variable among the three seasons studied because
of climatie variability. Sporocarp abundance and frequency wereV
positively correlated with basal area and density of mycorrhizal trees
and were negatively correlated with fern cover in hardwood forersts. .
Fruiting seasons extended from early July to late September or early .
October.
_ Numbers of species fruiting from the same four mz quadrats ranged from
0 to 7 in spruce forests and O to 5 in hardwood forests. Spatial patterns
of sporocarps of major species were eharaeterized by the variance·to-mean
ratio, mean erowding, patehiness, and spatial autocorrelation and were
found to exhiblt highly aggregated, contagious patterns. Interspecific
associations between pairs of major species were measured by 2 ¤ 2
eontingency tables and Cole's index of association.
A taxonomie and geographie survey of Rggsylg and Lggtggjygloeeurring
in both the quantitative study areas and in similar habitats in the
Southern Appalaehians was presented.
I would like to thank Dr. Orson K. Miller, Jr. for his teaching,
guidance, logistic support, and patience during the course of my graduates
studies. Drs. Golde I. Holtzman and W. Carter Johnson provided
indespensible assistance and guidance in the design, execution, and
analysis of the quantitative aspects of this study. The teaching,'
professional guidance, and personal advice of the other members of my
advisory comittee, Drs. Khidir W. Hilu, Robert A. Peterson, and R. Jay _
Stipes is greetly appreciated. The friendship, assistance, suggestions,”_
and logistic support of fellow graduate students, technical staff, and
faculty, past and present, at VPI & SU is also appreciated.
Financial support was provided by a graduate teaching assistantship
from the Department of Biology, VPI & SU. The Jeffress Memorial Trust
provided me with summer support and with a unique opportunity to learn
the higher fungi of Virginia. A Gertrude S. Burlingham Fellowship from
the New York Botanical Garden supported me during the spring of 1984 and
allowed me to study first-hand many of the Russulas and Lactarii described
by G. S. Burlingham, C. H. Peck, H. C. Beardslee, C. H. Keuffman, W. A.
Murrill, and R. Singer.
Finally, my education could not have been completed without moral and
financial support from my family and my wife, Nancy. Many thanks to all.
Acknowledgements iv
TABLE QB SQNIENIS
INTRODUCTION ........................... 1
CHAPTER 1. COMPARISON OF FRUITING OF ECTOMYCORRHIZAL BASIDIOMYCETES
BETWEEN RED SPRUCE AND NORTHERN HARDWOOD FORESTS ........ 4
Introduction ........................... 4
The study areas ......................... 8
Methods and materials ...................... 11
Results ............................. 14
Species diversity ....................... 14
Inter·plot relationships .................... 25
Sporocarp frequency and density ................ 30
Fruiting Phenology ....................... 33
Length of sampling period ................... 36
Discussion ............................ 37
CHAPTER 2. SPATIAL PATTERNS AND INTERSPECIFIC ASSOCIATIONS OF
ECTOMYCORRHIZAL BASIDIOMYCETES IN RED SPRUCE AND HARDWOOD FORESTS 47
Introduction ........................... 47
Methods .................... . ........ 49
Results ............................. 52
Sporocarp patterns ....................... 52
Interspecific associations ................... 60
Discussion ............................ 70
Table of Contents V
CHAPTER 3. SYNOPSIS OF RUSSULA IN THE HIGH-ELEVATION FORESTS OF THE
SOUTHRN APPALACHIANS ...................... 76
Introduction ........................... 76
Synopsis of Russula in the high·elevation forests ........ 79
CHAPTER 4. NOTES ON LACTARIUS IN THE HIGH-ELEVATION FORESTS OF THE
SOUTHRN APPALACHIANS ...................... 97
Introduction ........................... 97
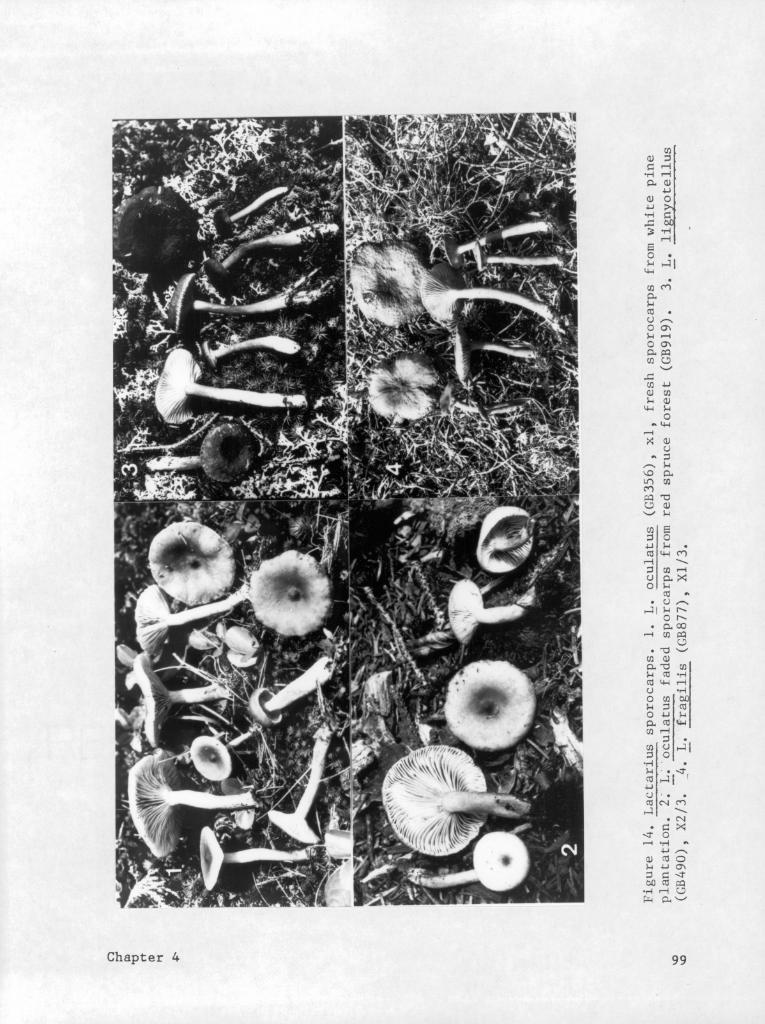
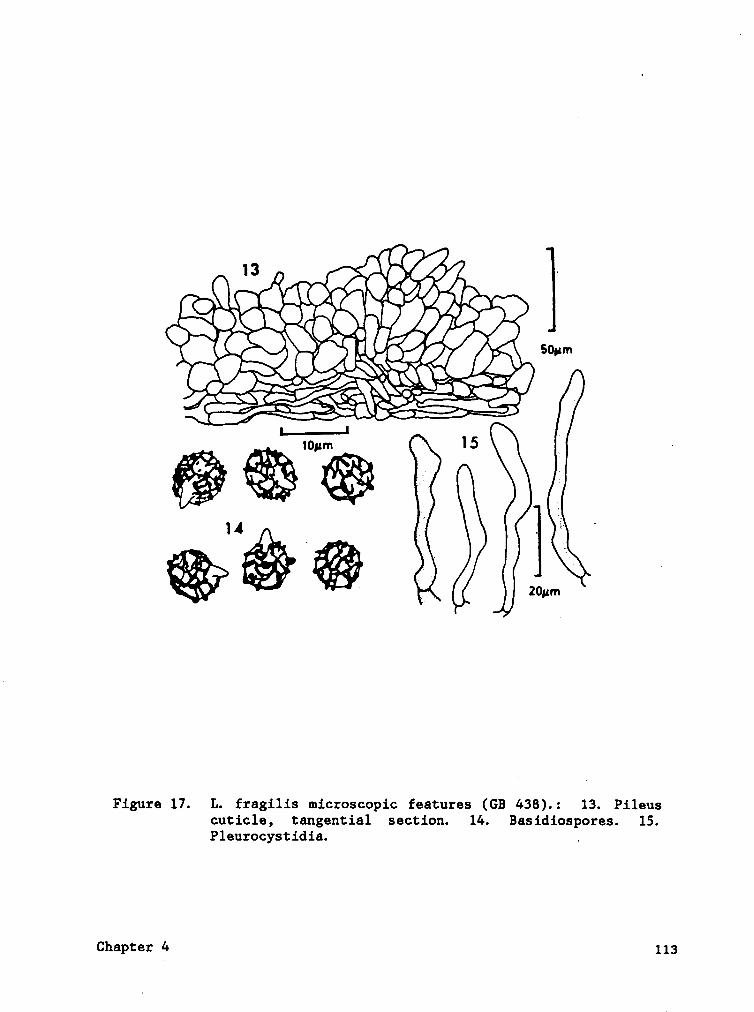
Descript ions of taxa .......................98
CHAPTER 5. DISTRIBUTION OF LACTARIUS IN THE HIGH-ELEVATION FORESTS
OF THE SOUTHRN APPALACHIANS ................. 118
Introduction .......................... 118
Habitats and distribution ................... 121
Discussion ........................... 124
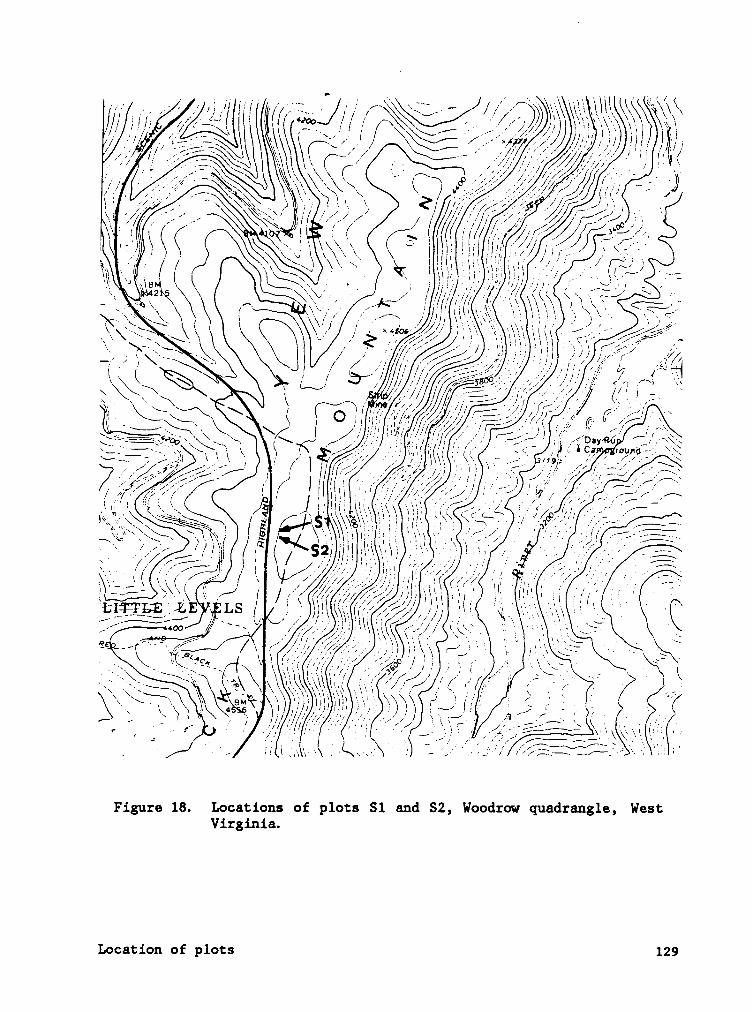
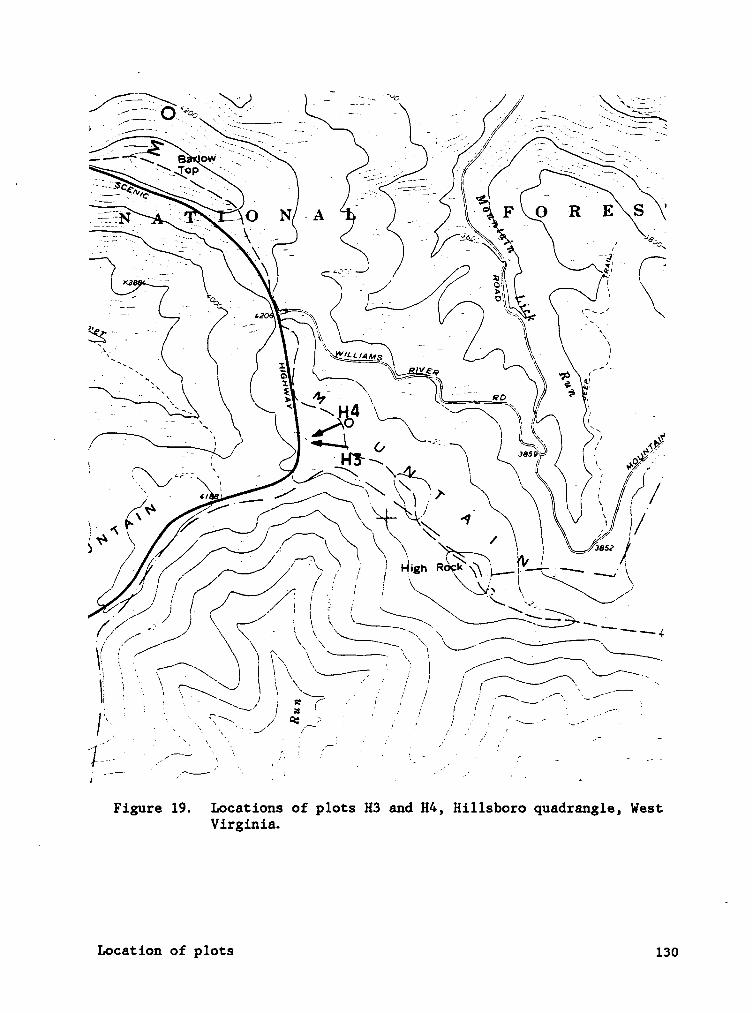
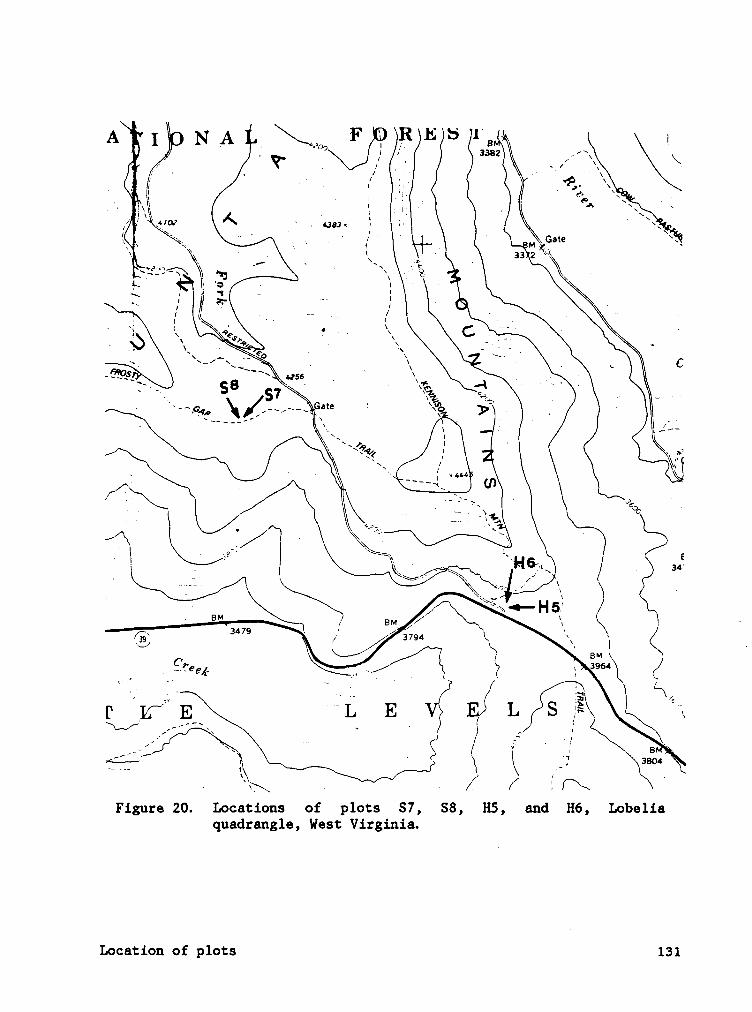
APPENDIX A. LOCATION OF SPRUCE AND HARDWOOD PLOTS, POCAHONTAS CO.,
WEST VIRGINIA. ........................ 128
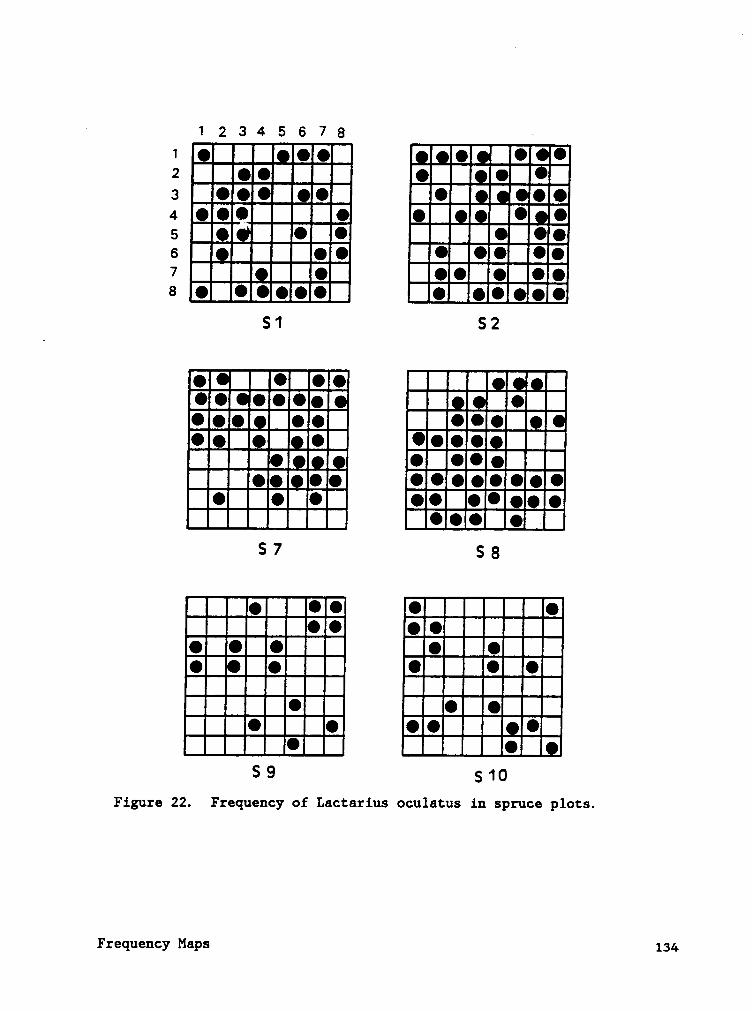
APPENDIX B. FREQUENCY MAPS OF MAJOR SPRUCE AND HARDWOOD
BASIDIOMYCETES. ........................ 133
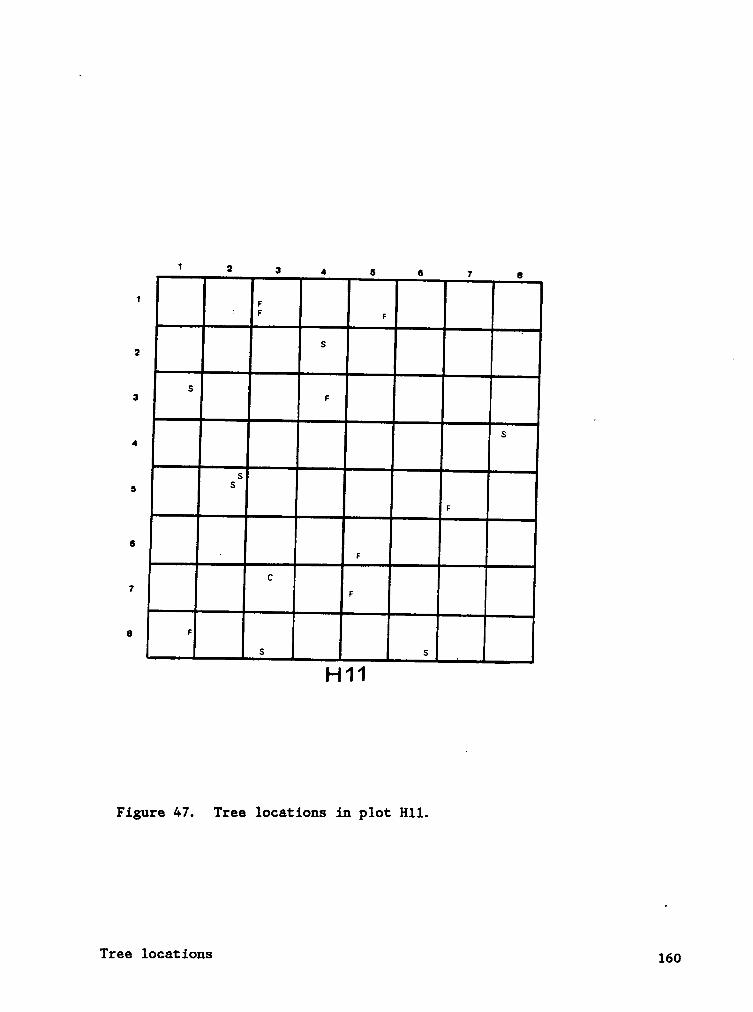
APPENDIX C. TREE (> 2 CM DBH) LOCATIONS IN HARDWOOD AND SPRUCE PLOTS. 149
APPENDIX D. RAW DATA 1981-83. ................. 162
Table of Contents Vi
BIBLIOGRAPHY .......................... 163
BIBLIOGRAPHY .......................... 164
VITA 173
Table of Contents v;|_;|_
LI§IQEFigure1. Red spruce and northern hardwood transition zone on thesouth end of Gauley Mt., Webster Co., West Virginia. . 6
Figure 2. Dominance·diversity curve for spruce ( ¤ ) and hardwood (u ) plots. ...................... 17
Figure 3. Species-area curve for spruce ( A ) and hardwood ( 0 )plots. ........................ 23
Figure 4. Ordination of spruce ( 0 ) and hardwood ( • ) plots byfungal frequency. ................... 29
Figure S. Ordination of spruce ( 0 ) and hardwood ( • ) plots by treebasal areas. ..................... 31
Figure 6. Sporocarp phenology of all species in hardwood and spruceplots. ........................ 39
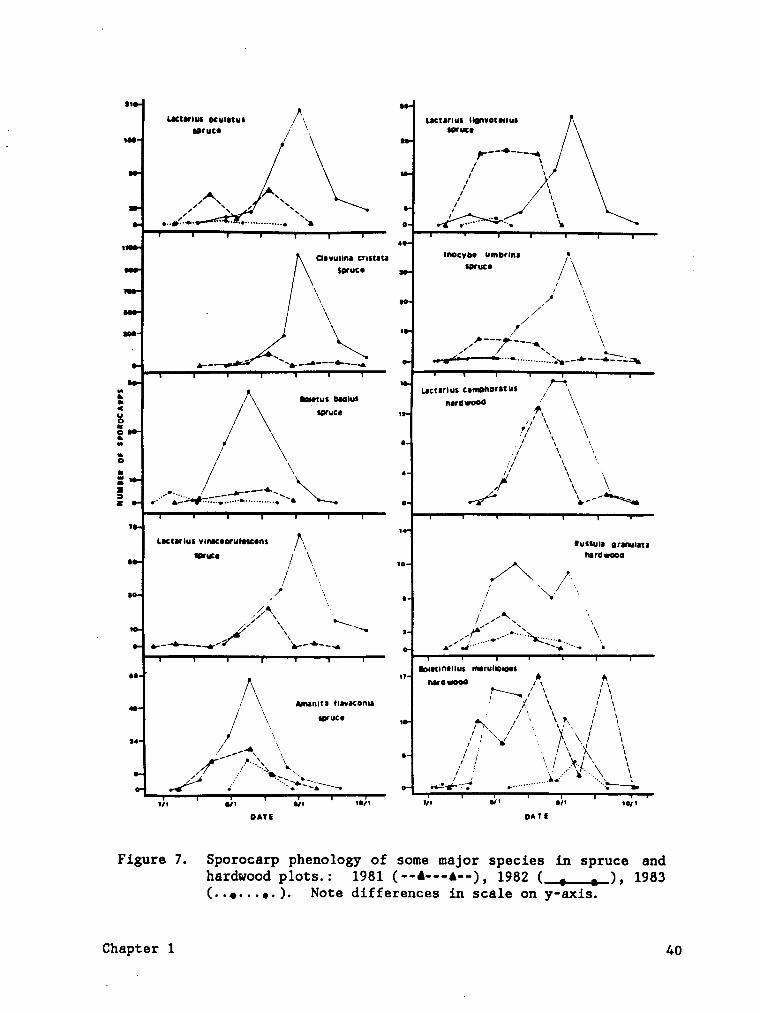
Figure 7. Sporocarp phenology of some major species in spruce andhardwood plots. .................... 40
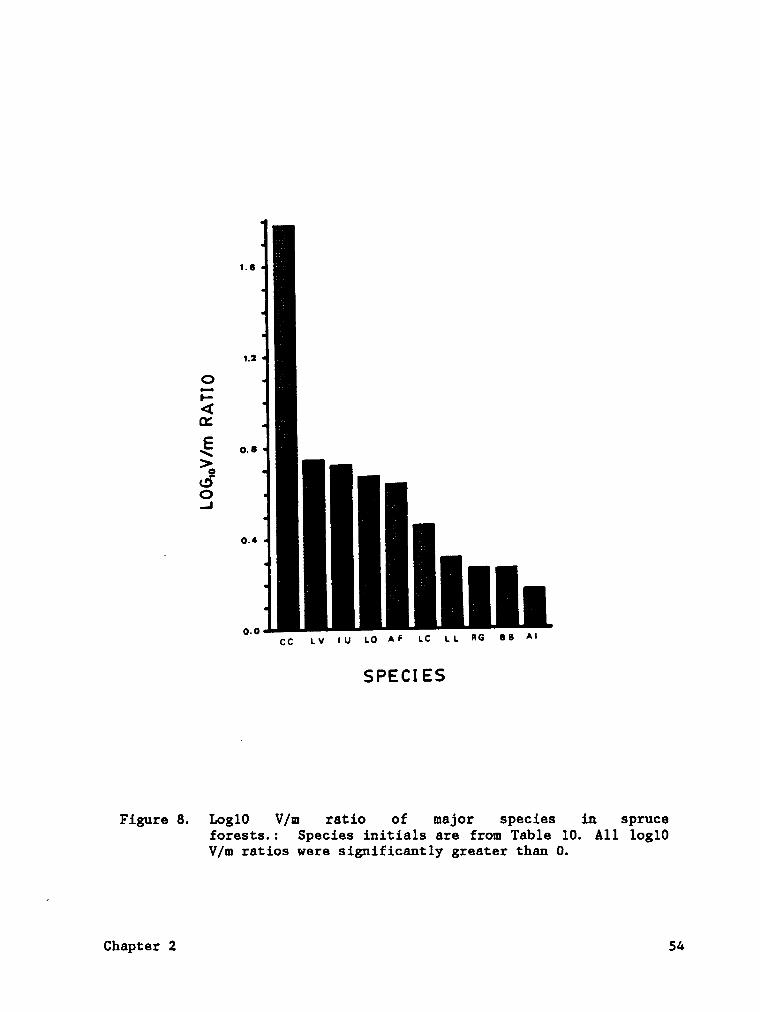
Figure 8. Log10 V/m ratio of major species in spruce forests. . . S4
Figure 9. Mean crowding of major species in spruce forests. . . . 55
Figure 10. Patchiness of major species in spruce forests. .... 56
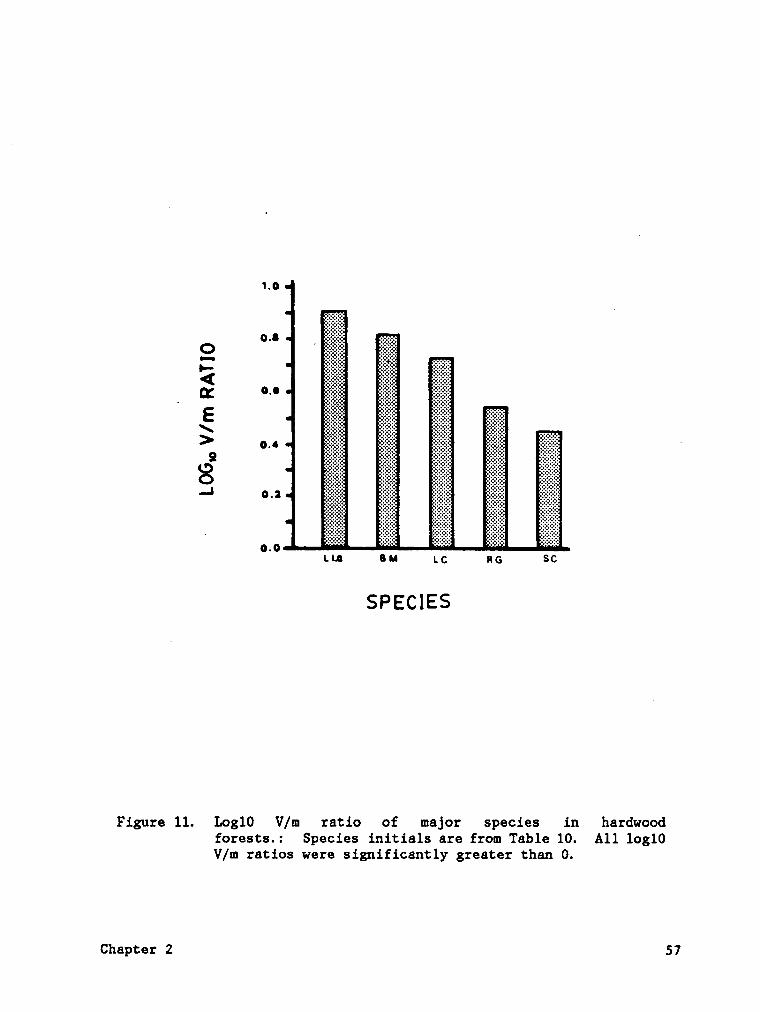
Figure 11. Log10 V/m ratio of major species in hardwood forests. . 57
Figure 12. Mean crowding of major species in hardwood forests. . . S8
Figure 13. Patchiness of major species in hardwood forests. . . . 59
Community studies of higher fungi have relied largely on quantitative
descriptions of fruiting. The limitations of studying fungal communities
based upon observation of sporocarps have been discussed by Hueck (1953),
Arnolds (1981), and Fogel (1981). Sporocarp productivity has been
estimated in one type of higher plant community (Richardson, 1970; Fogel,
1976) or has been compared among different plant communities (Lange, 1948;
Hering, 1966; Petersen, 1977; Wasterlund and Inglelog, 1981; Arnolds,
1981). Sporocarp presence, frequency, or productivity have been used
alone or in conjunction with the higher plants to compare vegetation
samples (Lange, 1948; Hering, 1966; Petersen, 1977; Arnolds, 1981). The
influence of precipitation and temperature on the phenology and
productivity of fruiting has been investigated to understand the
environmental conditions influencing the physiology of the fruiting
(wilkins and Patrick, 1940; Wilkins and Harris, 1946; Fogel, 1976).
Monitoring sporocarp productivity of different fungal species in a forest
dominated by a single ectomycorrhizal host has indicated possible
ectomycorrhizal associates of the dominant tree (Trappe, 1962;
Richardson, 1970; Fogel, 1976). Observation of the spatial pattern and
relative numbers of sporocarps in permanent reference areas over extended
time periods has elucidated the spatial pattern and relative abundance
Chapter 1 4
of vegetative mycelia in the forest rhizosphere (Last gg gl., 1983;
Newell, 1984; Cotter and Bills, in press).
_ Quantitative studies of macrofungal communities have usually focused
on sporocarp numbers or sporocarp biomass in standard reference areas.
The critical assumption of many of these studies was that the relative
productivity of sporocarps among fungal species in some way reflected
their relative dominance, mycelial biomass, or resource utilization.
Naturally occurring mycelia cannot be delimited in a direct manner, except
in rare cases. Presently there is no basis for correlating sporocarp
biomass or sporocarp numbers with mycelia biomass. However, sporocarp
density is a useful parameter because it can be applied objectively in
any study, and if sampling intervals and methods are comparable, sporocarp
densities can be compared among communities. This parameter is included
in this study to provide continuity with previous investigations. Rather
than emphasizing sporocarp numbers as a measure of mycelial activity, we
estimated the frequency with which sporocarps were present in relatively
small contiguous quadrats. lf the quadrats are small enough, frequency
' can show that sporocarps in widely separated quadrats at one sampling _
time may be part of a single zone of contiguous fruiting when sporocarps
are sampled in quadrats in intermediate positions. Therefore, frequency
provides an estimate of the spatial extent or ubiquity of the fungal
mycelium.
Species concepts of Basidiomycetes are based upon sporocarp
morphology, but rarely on habitat preferences. Statistical relationships
Chapter 1 5
between the distribution of fungal species and their associated higher
Vegetation may refine these concepts. In many "modern" taxonomic
treatments of Basidiomycetes, little attention is devoted to habitat
descriptions. Often fungi are described simply as fruiting "underA
conifers" or "in mixed woods". A species that appears to be "rare" or
is "poorly known" may be common locally in specialized habitats. This
local abundance may not be recognized by conventional collecting methods
but can be detected by periodic, systematic sampling of local communities_
(Arnolds, 1982; Fogel, 1982).
The ectomycorrhizal fungal community of pure red spruce forests was
chosen for study because the fungi could be assumed to form
ectomycorrhizae with only a single host and that niches for
ectomycorrhizal fungi would be limited by the availability of a singleA
tree species. Variations in tree age, root age, root size, litter and
soil depth, etc. could contribute, however, to niche diversification
among fungal species that occupied a similar substrate. To identify
properties of the community and species unique to the red spruce stands
we compared them to surrounding northern hardwood stands (Braun, 1950;
Whittaker, 1956; Core, 1966) of mixed ectomycorrhizal and endomycorrhizal
trees.
No comprehensive source of information on Basidiomycetes associated
with the red spruce forests of eastern North America exists, and few
mycorrhizal associates of red spruce are known (Treppe, 1962; Homola and
Mistretta, 1977). The stands of red spruce in this study are part of the
Chapter 1 7
disjunct southern extension of the boreal coniferous forest. The
structure and floristic composition of the Southern Appalachian and
Allegheny red spruce-Fraser fir-balsam fir forests are similar to those
in New York and New England (Oosting and Billings, 1951; Mclntosh and
Hurley, 1964; Stephenson and Clovis, 1983). Are the Basidiomycetes
associated with red spruce similar throughout its geographical range?
Presently, the only way to answer this question is by examining the
usually incomplete habitat descriptions of Basidiomycetes to determine
whether they were collected in the vicinity of red spruce.
The main objectives of this study were (1) to determine how species
composition, diversity, and density of ectomycorrhizal Basidiomycetes
fruiting in forest stands dominated by a single ectomycorrhizal host tree,
red spruce, differed from those of nearby forest stands of mixed
ectomycorrhizal and endomycorrhizal hardwood trees, (2) to describe the
inter-year variation in fruiting and sporocarp density in both forest
types and to characterize the fruiting phenology of major Basidiomycete
species of both forest types, and (3) to provide a basis for comparing
Basidiomycete communities in similar coniferous and hardwood forests in
eastern North America.
IHE.S§I1ZD.YAREA§
The study areas were located near the eastern edge of the unglaciated
Allegheny Plateau in Pocahontas Co., WV, within the Monongahela National
Chapter 1 8
Forest (boundaries 38° l7° N latitude, 38° 07° N latitude, 80° 22° W
longitude, 80° 12° W longitude). Sites were located on ridge crests
(elev. 1200-1350 m) on shallow, rocky well-drained, sandy-loam or
clay-loam soils. Within the spruce stands, soil pH (n = 23) ranged from
3.3 to 3.8 with an average of 3.5, and soil organic matter content was
12.3 % i 3.6 % S. D. Within the hardwood stands (n = 20), soil pH ranged
from 3.3 to 4.5 with an average of 3.6, and soil organic matter content
was 10.6 % i 3.4 % S. D.
Annual precipitation in the vicinity of the study sites during a
five-year period in 1967-72 ranged from 144 cm/yr to 160 cm/yr (Edens,
1973). Precipitation patterns for each growing season were estimated from
data collected by the U. S. Forest Service, Marlinton, WV, (elev. 650 m)
about 15-20 km west of the study areas (Table 1). Extended periods of
high humidity and fog are partially responsible for the persistance of
red spruce at these southern latitudes (Core, 1966). Frost-free periods
A range from 88 to 145 days (Edens, 1973).
Both spruce and hardwood stands were second-growth (55-75 years old).
Floristically and structurally the spruce stands were similar to stands
described by Stephenson and Clovis (1983). Spruce stands were nearly pure
red spruce with a sparse to dense shrub layer of ygggjgjgg
ggythgggggpgg, suppressed spruce seedlings, and ferns. An extensive
ground cover of bryophytes, especially the leafy liverwort ßgzzggjg
ggilgbgtg (L.) S. F. Gray, was often present. Composition of the hardwood
Chapter 1 9
Table 1. Biweekly summary of average (cm/day) and total (cm) rainfallfor growing seasons of 1981-3 based on U. S. Forest Servicedata, Marlinton, WV.
1981 1982 1983weeks average total average total average total
Jun 1-15 0.53 7.85 0.79 12.01 0.25 3.71
Jun 16-30 0.20 3.00 0.10 1.55 0.23 3.58
Jul 1-15 0.97 14.27 0.54 8.05 0.27 4.04
Jul 16-31 0.48 7.54 0.28 4.45 0.25 3.81
Aug 1-15 0.28 4.29 0.25 3.76 0.12 1.80
Aug 16-31 0.03 0.51 0.25 3.76 0.01 0.10
Sep 1-15 0.71 10.52 0.08 1.09 0.02 0.25
Sep 16-30 0.09 1.40 0.16 2.42 0.20 3.07
total (cm/season) 49.38 40.82 20.37
Chapter 1 1Q
stands are listed in Table 3. Herbaceous plants, including ferns, and
young Age; stems were abundant in the understories of the hardwood stands.
MEIHQQSANDHAIERIALS
This study was conducted during the growing seasons of l98l•3. Twelve
permanent 16 X 16 m (256 mz) plots were established on ridge crests of
three different mountains. On each mountain, two plots were located in
V a spruce forest and two in a hardwood forest (Table 2). Each plot was
subdivided into 64 2 X 2 m (4mz) quadrats.
Spruce plots were selected to exclude as many other woody species as
possible. Hardwood plots were selected to be as physically close to the
spruce plots on the same ridge crest, without including any red spruce.
In each plot, DBH (diameter at breast height) of all stems greater than
two cm was estimated and mapped, and fern, bryophyte, and spruce seedling
cover was estimated. Hardwood plot H3 was destroyed by a survey crew in
the spring of 1983. Sporocarp density and frequency estimates for 1983
were based only ou the five remaining hardwood plots.
Only sporocarps of fungi of families known to form ectomycorrhizae
(Watling, 1982; Miller, 1983) and some whose ecological role is uncertain
me;g1jgjgee, Eggglgge spp., and some Hyg;gphg;ee spp.) were counted.
Litter decomposers (e.g. Qgllyhie, Me;eemig;, or Mygege spp.) or
Chapter 1 11
Table 2. Location, forest type, and plot label.
forest typelocation spruce hardwood
Black Mountain Sl & S2 H3 & H4
Kennison Mountain S7 & S8 H5 8: H6
Rocky Knob S9 & S10 H11 & H12
Chapter 1 12
bryophyte-associated species (e.g. as Qglering spp.) were not counted.
Sporocarps of agarics and boletes were easily defined, but the coral
fungus, Qlgygliga ggistgtg, forms multiple, fused stems. A single
sporocarp of this fungus was defined as a separate stem completely
surrounded by the surface of the litter or the bryophyte layer, although
the stems may have been fused below ground. From examination of records
of sporocarp positions from previous sampling times, it was evident that
nearly all sporocarps deteriorated between sampling periods. Only
sporocarps that were long-lived and could have been counted twice (e.g.
ßglggggggmg gitringm, large Lggggrigg spp.), those needed for
identification, and those needed for Voucher specimens were removed from
the plots. Sporocarp numbers were prcbably underestimated because
sporocarps of fleshy fungi are usually short-lived (Richardson, 1970;
Lacy, 1984) and could have fruited and deteriorated between sampling
times. Representative Voucher specimens of all fungi are deposited at
VPI. Vascular plant nomenclature follows that of Strausbaugh and Core
(1978). Fungal nomenclature is listed in Table 4.
Individual sporocarps were counted within each 2 ¤ 2 quadrat at each
visit. Plots were visited eight to ten times per growing season at 7-
to 17-day intervals with the average time interval between visits 13.1 i
3.2 days S. D. (n = 23).
_ Density is the number of sporocarps per unit of area (either in all
plots of a forest type or in each plot). Frequency is the number of
quadrats in which a species occurred summed over the entire the study.
Chapter 1 13
Percent frequency is the percentage of the total number of quadrats (384)
in each forest type in which a species is present. The total number ofl
quadrats in which a species occurs in any given year is yearly frequency.
Frequency of a species is not the sum of its yearly frequencies.
BE§LJLI§
S.2EQIE§ IZIYERSJIY
Forests with several possible mycorrhizal hosts might have a greater
diversity of fungal associates than a forest composed of a single
mycorrhizal host. Two elements of species diversity are usually
considered, (1) species richness, the number of species, and (2)
equitability, the evenness of the contributions of different species to
the community. Fungal species richness was comparable between both forest
types (Tables 4, 5), but the two forest types exhibited little overlap
in fungal species composition (Tables 4, 7). A total of 54 species was
encountered over the three years, 27 species in the spruce plots, 36 in
the hardwood plots (Table 5). Nine (17 Z) species occurred in both forest
types. The family Russulaceae accounted for more species than any other
family in both forest types, 39 Z of the species in the hardwoods and 44 ‘
Z of the species in spruce. The total number of species occurring only
in one forest type or the other (19 only in spruce, 26 only in hardwoods) _
were not significantly different from what would be expected assuming the
two forest types were equally likely habitats (22.5 in each forest type),
Chapter 1 14
Tab1• 3. Basal ar••s lmz7ha) of tr•• sp•ci•s in spruco and
hardnood plots. Plots ar• ord•r•d from laft to ridut by ascanding _ ‘first principal couponant scoras. $p•ci•s ar• ordorad by dascandingfirst principal couponant scoros from a sp•ci•s ordination.Plot labals ar• indicatad in Tabla 2.
(Table 5). Likewise, the mean number of species per plot was not
. significantly different between spruce and hardwood plots (Table 5). The
similarity in slopes of the species-area curves, (Fig. 2) also indicated
comparability in species richness, although the initial slope of the
hardwood curve was slightly steeper because of the encounter of more
species in hardwood plots than in spruce plots.
Species equitability was greater in the hardwood plots than the spruce
plots. The Shannon-Wiener index (Shannon and Weaver, 1949), H'= -2 PiV
log2 Pi, was used to compare both species richness and equitability
between both forest types (Table 5). Where Pi is the probability of
sampling the ith species among all species. Shannon-Wiener diversity (H')
was greater for hardwood plots than spruce plots using both sporocarp
frequency and density (Table 5) because of the greater number of species
and greater equitability among species. Equitability or eveness of a
community (J') (Pielou, 1974, p. 300) can be measured directly from the
ratio of the observed species diversity (H') to the maximum value of H'
(H°max) in a completely equal community with the same number of species
(S). Therefore, equitability (J') is given by H'/ log 2 S. As expected,
higher (H') values were obtained using sporocarp density because density
measurements were more equally distributed among species. The
dominance-diversity curve (Greig-Smith, 1983), (Fig. 1) indicated that a
high proportion of the frequency in both forest types was concentrated
in relatively few species, that a high proportion of the species appear
rarely, and equitability was greater among the hardwood species. Greater
Chapter 1 16
III1• El
' "UIo
S :1 • §= o= - I5 NQIIOE
InA
5Q Ü u QE l • „„a C10 I 3S
••cx lu; 2nn g ¤¢
"' ¤ nun "‘CI II »
¤¤ =KIIIIII0 ¤¤¤¤¤¤¤¤Ü¤¤ 1:1:1 IIIIIIII 0• no II ao zu ao es
SPICIIS Stoutuct
Figure 2. Dominance-diversity curve for spruce ( ¤ ) and hardwood (• ) plots. : Most frequent species to least frequentspecies are ordered from left to right.
Chapter 1 17
T•b1• 4. ßasidianyeatss balonging ta •ata«•yearrhiz•1 f•••il.i•s aeaurringin spruea ls) and harchaad Ih! faust typ•s.
F••i1y• sp•ci•s faust typ•
IMANITACEAEAmnib ilmuszmin Atk. •
muib hlx: |$ah••ff.l •¤•«·•. •, InÄllüil S•¢•-. s, h
ßemuib sasiliu ts. a •«·.1¤••1mmib s.t¤n¤u1.•b Fr.• smsu sm. hgi}; gig}; lBu11.:Fr.) Vitt. h .
BOLETACEAEuisjimiisa mmlinidu «s¤+ui•n.1 Hurrill. hBzlßm !.f.f.i¤sa1 PK- 1 hklein Indian rr. •„ h
ggü IB. I C.) Seec. I1Enjglg IPR.) Seee. I1Enisism •¤-
h
HYGROPHORACEAE
|$cI·••ien.I Fr. I1Iknuff.) Smith I Heel. I1(Fr.) Fr. I1
(Fr.) Fr. I1Uamnanbzug sp- bTRICHOLGIATACEAE§.¥!$&r:gmi1¤§1im¤¤Fr•)F•y¤d •.h Ilgggjg lggg I$o¤p.ZFr.) Berk. I Bree. e, I1
SCLERODERHATICEAESslncszsang süzimn P••·••
h
19.Chapter 1
T•b1• 4. Cant.
Fnnily, •p•ci•• lplot
PAXILLACEAE
ggging; imgigg Ißatseh.) Fr. •
RUSSULACEAEFr. uh
Lasimim magnlixm Pk- • .Lasizcius simusn Pk- H
L1s$.u;iy1 m.¤.¤üi Pk- •• HLgsigdm ndmm Pk- H
lLasiuzisn limxsztnllus Smith I M••1- •
Lgjggigg gglgjgg IPR.) Burl.•
Imizciua mcdistn Pk- •
],;g_g;_;ig; ghgjgggig; IBu11.) Fr. h
Lasxgcisa Smith •
ßgggß; gg; L•C1•i•· •
· Bsasula slemtlma S•·¤v• •t
ßmub sanmm Pk- HB!-!.t§.ul.! mi.f.21il
$•¤r-• 3;:;;; Sh•ff••· •
ßggggi; IPR.) PR.•• h
ßggggi; Fr. ? h
Basale Iszzmäxzkii SH•f*••· Hauml: mm ¤•-•r1- Hmmala mlm ¤¤•·1- hBiggi]; ;j_],yj,gg_],; $h•ff•r ag h
eByssyla sgtatm Smith 1 H&;;_;g_],; gigggg I$eh••ff.) Fr. In
Chapter 1 20
Table 5. Comparison of species richness, species diversity, species
equitability, and species density of ectomycorrhizal
fungi between hardwood amd spruce plots
(n=6 plots/forest type). Shannon-Wiener index (H') and
z
equitability (J') calculated from sporocarp density and frequency.
spruce hardwood
number of species 27 36
number of species 188 27a
unique to each
mean i S. D. of
A
species/plot (256 mz) 15.7b i 2.2 13.2b i 4.3
' maximum number of
species/plot (256 mz) 19A 19
A
minimum number of I
species/plot (256 mz) 13 7
H'(frequency) 2.39 4.11
H'(density) 3.59 4.52
J'(frequency) 0.50 0.76
J'(density) 0.79 0.87 ·
a Not significantly dlfferent, P>0.10, xz = 1.8, 1 d.f.
b Not significantly different, P>O.10, T = 1.0, 10 d.f.
Chapter 1 21
equitabilty in the hardwood forests was indicated by direct measurement
of J!. .
The total number of species fruiting in a quadrat estimates the minimal'
number of ectomycorrhizal species that may be occupying the rhizosphere
within these small areas (Table 6). Species density was much higher in
the spruce plots than in the hardwoods. The proportion of the quadrats
with no sporocarps was much higher than those with sporocarps, 225 (59'%)
in the hardwoods compared to 59 (15 %) in the spruce. Two species
fruited in most (85 %) of the occupied spruce quadrats, and as many as
seven species fruited in a single quadrat. One species fruited in most
(25 X) of the occupied hardwood quadrats, and as many as five species
fruited in a single quadrat.
The concept of minimal area (Cain, 1938), or the smallest area in which
the species composition of a community is adequately represented, is
impossible to apply prior to sampling macrofungi because sporocarp
densities are in a state of continual flux. However, the possibility of
achieving a minimal sampling area was examined in retrospect by
constructing the species·area curve (Greig·Smith, 1983) (Fig. 2) from
accumulated data on sporocarp locations. One criterion for minimal area
(Mueller-Dombois and Ellenberg, 1974) is a sample size that contains 90
to 95 % of the maximum number of species encountered in the largest sample
unit. This criterion is impractical for fungi because the maximum number
· of species cannot be determined. Cain (1938) considered the minimal area
Chapter 1 22
'
35
m 30Lu A
{3 25 ,9
Eth 20 °U- AO
‘
O 15•
_ •Oz 10
• ‘
O‘A
5
0 zoo 600 600 000 1000 1200 v•0¤ 16¤¤
AREA MZ
Figure 3. Species-area curve for spruce ( A ) and hardwood ( • )plots.: The total number of species in the sample areaswas plotted against increasing areas of contiguousquadrats. When 256 mz, the size of one contiguous plot,was reached additional plots were chosen randomly andtheir species added.
Chapter l 23
Tabla 6. Distribution af nuebars af sp•ci••in 384
quadrafs af spruea and hardnaod plats.
na. af na. af quadrafa
spaciaa/qeadraf spruaa harcbaaad
0 54 2261 83 97
2 105 443 84 144 42 35 ll 16 4 07 l 0
fatal aeenpiad qnndrafs 330 158 ·uaan 2 S. 0. 2.4 rt 1.2 1.5 t 0.8
nurbar af sp•ei••/cpadraf
Chapter 1 24
to be the point on the species·area curve at which an increase of 10 %
in the sample area yields only 10 % more species of the total number
recorded. Using this criterion, the minimal area for the spruce forests
is approximately 250 mz and approximately 150 mz for the hardwood forests.
However, Rice and Kelting (1955) demonstrated that this 10 X point wlll
shift continually to the right as greater total areas are sampled.
To compare and to summarize variation in fungal and tree species
composition among the plots, principal component analysis (PCA) (Gauch,
1982, chap. 4) was used to order the plots x species data matrices for
fungi and trees separately. The hardwood plots appeared to be distinctly
different from the spruce plots because of the taxonomic discontinuity
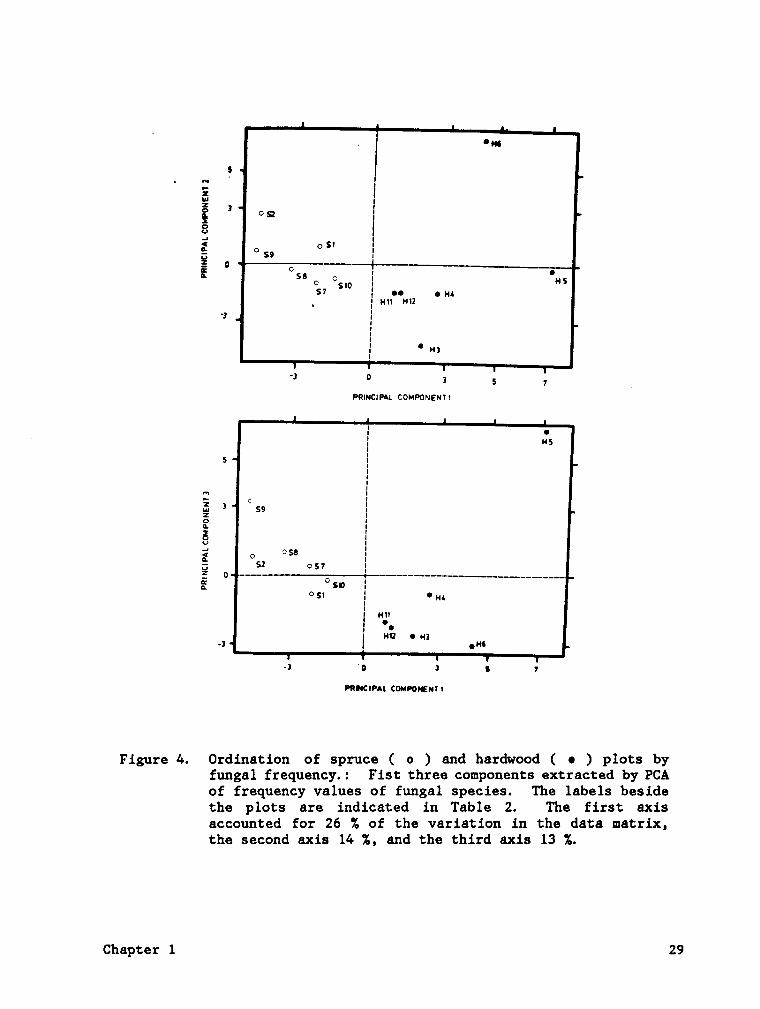
in fungal species between the two forest types (Fig. 4). Ordination
indicated a closer relationship among the fungal samples in spruce forests
than among hardwood forests. The spruce plots were relatively uniform
in fungal composition having nine fungal species in common. The wide
separation among hardwood samples along all three axes indicated a high
degree of taxonomic discontinuity. Hardwood plots H6 and H5 were the most
different from the rest, probably due to the high frequency of
ßglgtjggllgg mgggljgjggg in H6 and Lgg;g;1gs gigggggg in H5. In addition,
Hygggphgggg spp. and Engglgmg spp. were largely restricted to these two
plots (Table 7).
Chapter 1 25
T•b1•7. Pareant frnqaucy of basidionyuata sp•¤i•s in spruea and
Inrénood plots in southnastnrn Hast Virginia. Plots ars ordnrad fron1•ft to n·i¢1t by asoanding first principal cannvonnnts scoros. Spncix ·arn ordarnd by dasacnding first principal couponmt scoras fron a spnciasordirctiun. Plot labais are indieatad in Tsbla 2.
Figure /6. Ordination of spruce ( 0 ) and hardwood ( • ) plots byfungal frequency.: Fist three components extracted by PCAof frequency values of fungal species. The labels besidethe plots are indicated in Table 2. The first axisaccounted for 26 74 of the variation in the data matrix,the second axis 1/6 %, and the third axis 13 %.
Chapter 1 29
Some of the species common to both forests were among the most ubiquitous
(a-s-Las1:.a1:.1.u.1. Bsmsnlaxramuasa, Amanisalnmarasa, and
Lgggggjg lggggtg). These species are widely distributed in many forests
types and thought to have a broad ectomycorrhizal host ranges.
AND11EN.S.I1'X
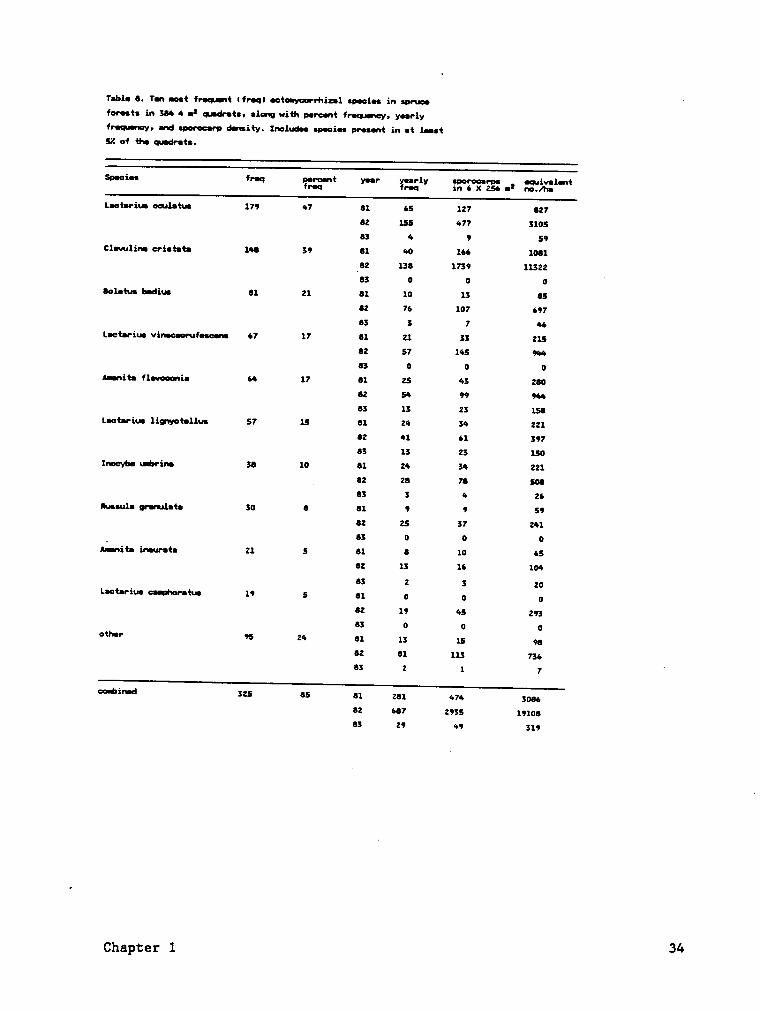
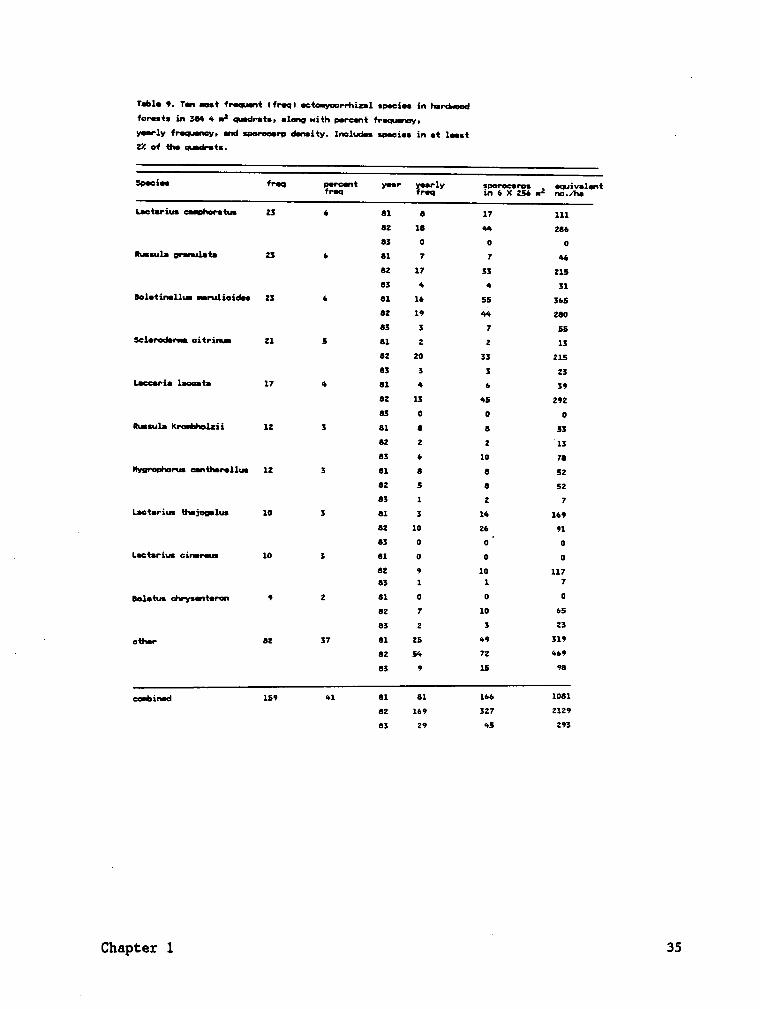
The frequency of major species is a measure of their relative ubiquity
within the plots (Tables 8, 9). These major species accounted for 88 Z
of the total frequency values, 96 Z of the total sporocarps in the spruce
plots, 66 Z of the total frequency values, and 75 Z of total sporocarps
in the hardwood plots during the three years.
Sporocarp frequency and density were highly variable among years.
Sporocarp frequencies and densities were consistently higher during the
second year for nearly all species, while the lowest values usually
occurred during the third year (Tables 8, 9; Fig. 5). Some species did
not fruit every year, especially the third year (Tables 8, 9; Fig. 6).
Species with the greatest sporocarp frequency or density were not the same
every year. Comparison of sporocarp frequency and density indicates that
the list of major species would be somewhat different if species were
ranked by density. For example, the sporocarp density of Qjgyyling
ggistgtg was greater than Lggtgrjgs ggglgtgs during the study, but L.
i
Chapter 1 30
E <
§E 0
OHLäE äbsr I
I
S7 %•"° I °°" „°¤s2 I
·2 0 3 5mmcnm commsurw
· II OH4
· II
·; IE I •HI2S20 S‘@°:$‘
MsIä ° S7 Sßsg
••_, • un}E
"‘I
ä'E OH)-a
I ~s‘ -2 0 a s
r¤••cum0¤u•=0••s»m
Figure S. Ordination of spruce ( 0 ) and hardwood ( • ) plots by treebasal areas.: The first three components extracted byPCA of basal area values of tree species. The labelsbeside the plots are indicated in Table 2. The first axisaccounted for 29 % of the variation in the data matrix,
I the second axis 22 %, and the third axis 16 %.
Chapter 1 31
ggglgtys was more frequently encountered and therefore was more
ubiquitous throughout the spruce plots.
Sporocarp frequency and density were much higher in both the spruce
and hardwood plots during 1981 and 1982. (Tables 8, 9; Fig. 5). Fruiting
was severely suppressed by the extreme drought conditions during July and
August of 1983 (Table 1). July and August of 1983 were the driest in
eastern North America in nearly 50 years (Wagner, 1984). Sporocarp
frequency and density was greater in spruce plots than hardwood plots
during the first two seasons but not the third.
The higher sporocarp frequency and density in the spruce plots may be
due partially to the greater number of trees providing a substrate for
ectomycorrhizal fungi. Spearman°s rank correlation coefficient (p) was
. used to measure the association of vegetation parameters with sporocarp
density and frequency. When all twelve plots were considered, the density
of ectomycorrhizal canopy trees (£;gxing; included to account for
frequency and density of ßglgtjngljgg mgrgljgjggg) was positively
correlated with sporocarp density (p = 0.69; P = 0.01), the basal area
of ectomycorrhizal canopy trees (lggxingg included) was positively —
correlated with sporocarp density (p = 0.75; P = 0.005), the density of
ectomycorrhizal canopy trees was positively correlated with sporocarp
frequency (p = 0.63; P = 0.006), and the basal area of ectomycorrhizal
canopy trees with sporocarp frequency (p = 0.73; P = 0.007). When the
six spruce plots or the six hardwood plots were analyzed separately,
Chapter 1 32
I
neither tree basal area nor tree density was correlated with sporocarp
density or sporocarp frequency. ‘
. Several investigators have observed an inhibitory effect of ferns and
herbaceous vegetation on sporocarp productivity (Wilkins and Harris,
1946; Wasterlund and Ingelog, 1981). This effect was observed in some
of the hardwood plots where fern cover ranged from 1.5-58 %. Within the
hardwood plots, there was a negative correlation between fern cover and
sporocarp density (p = -0.87; P = 0.05) and fern cover and sporocarp
frequency (p = -0.90; P = 0.04).
ERHIIINQThe
fruiting season for ectomycorrhizal fungi begins in early July and
extends into late September or early October for both forest types (Fig.
5). Sporocarps were observed for 89, 91, and 75 days for the three years,
respectively. The end of the first two fruiting seasons coincided with
·theadvent of heavy frosts or snowfall. In the third year, prolonged
drought in combination with cold temperature prematurely ended the
fruiting season. In both forests during the first and third years,
sporocarp density declined sharply after the first of August because of
late summer drought (Fig. 5). Both forests exhibited a strong peak of
sporocarp density in early September of the second season. During the
second half of August, 1982, rainfall was much greater than the same time
Chapter 1 33
T•b1•8. Tan nat faqnnt Ifr•q| •¤t¤¤yarrhi¤1 so•¤i••
in soruaf¤r••ts
in 386 6 6* ande-ots, dung with areunt fr•¤u•rn¤=y» y••r1y
and Kgggyjg (Bills and Miller, 1984).Lg_g_t_g;_j,g_5_was
described from a red spruce forest on Clingman°s Dome, Tennessee
(Hesler and Smith, 1979) and has not been reported elsewhere. The
remainder of the species in the spruce plots apparently never have been
reported fruiting in association with red spruce.
A high proportion of the species diversity in both the spruce and
hardwoods was attributable to species of the Russulaceae. This would
probably be the case in most boreal or temperate forests dominated by
ectomycorrhizal trees. Certain genera of ectomycorrhizal Basidiomycetes
commonly found in other coniferous forests (especially Ring; or Lggig)
were absent from the red spruce forests. Among these are species in the .
genera Irishclnma. Hxzmuhnmla (S¤<=ti¤¤¤
Hxzmnhoma.Chapter1 37
Qgmphjgjgg, Qhgggggmphgg, and Sgjllgs. There are no reports of hypogeous
Basidiomycetes in red spruce forests.
Most of the species of the hardwood plots are co«~on in the deciduous
forests of northeastern North America. Entglgmg lgggnigygtjg is known
only from North Carolina and Tennessee (Hesler, 1967). Prior to this
study, Kgsgglg gggglgns was known only from the type locality in Vermont
(Bills, 1984). Kg5;g1g.gpg;;5 is a poorly known species described from
northern hardwood forests of Vermont and may be synonymous with Buggy];
pggillg Peck (Singer, 1957). A few of the species from the hardwoods have
been reported to be associated with specific woody hosts; e.g.
ßglggjggllgg mgggligjggg with Egggjngs spp. (Snell and Dick, 1970),
Lastaxiuasinezmawith F.az¤1szz.a¤s1i.f.o.1i¤„ and Lastariusthsrinaalus with
ßgtyla spp. (Hesler and Smith, 1979). These associations were evident
in this study because plots with high density of these species either had
these trees in the plots or near the edge of the plots (Table 7).
§glg;ggg;mg giggjggm which fruits in association with many woody plants,
consistently produced sporocarps near Qggggyg gghgg in plot H4. Lgggggig
lgggggg, Lgggggigg ggmphgrgggs, and ßggsglg ggggylgtg had the widest
amplitudes in habitat of any of the major species, and both were found
fruiting in most plots.
The number of species found in spruce (27) and hardwoods (36) compares
favorably with numbers of species reported by other investigators. Hora
Chapter 1 38
120HARD? ·
100
I0
[Q .
Q soEä
’°g
0.V1ILO
g‘$°
svnucsIDg 12soZ
1000
7
500
250........ ....... \
07/1 8/1 9/1 10/1
DATE”
Figure 6. Sporocarp phenology of all species in hardwood and spruceplots.: 1981 (-4--+-), 1982 ( • • ), 1983(...•....•. ). Note differences in scales on y-axis.
Figure 7. Sporocarp phenology of some major species in spruce andhardwood plots.: 1981 (··l•--A--) , 1982 ( , , ), 1983(. .•. . . •. ). Note differences in scale on y-axis.
Chapter 1 40
(1959) reported 25 species and Richardson (1970) 12 species belonging to
ectomycorrhizal families fruiting in Scots pine plantations. Fogel
(1976) found 24 putatively ectomycorrhizal hypogeous fungi in a Douglas
fir stand. A Swedish stand of Norway spruce had 25 species fruiting
(Wasterlund and Ingelog, 1981). Stands of Qggrgggandin
Great Britain produced 4 to 11 species (Hering, 1966). In addition,
the numbers of epigeous species fruiting in various spruce stands is
comparable to the number of types of ectomycorrhizae observed on spruce.
Worjciechowska (1960) described 16 "form genera" of ectomycorrhizae on
Norway spruce within its northern range in Poland. Thirty·seven ”form
genera" of ectomycorrhizae were observed throughout the range of Norway
spruce in Poland (Dominik, 1961). Thomas gg gl. (1983) observed 25 types
of ectomycorrhizae in English Sitka spruce plantations.
_ We believe the stands described here are representative of other red
spruce and northern hardwood stands in the Southern Appalachians. Most
of the major species reported here have been observed fruiting abundantly
in other red spruce and northern hardwood stands in West Virginia,
Virginia, and North Carolina. However, many other Basidiomycetes
belonging to ectomycorrhizal families have been collected in spruce
andnorthernhardwood forests throughout the Southern Appalachians.
Therefore, my results do not represent the entire mycorrhizal fungal
flora.
Some doubt exists as to whether the concept of minimal area can be
applied to fungal communities (Christensen, 1981). An adequate
Chapter 1 41
species·area curve for macrofungi cannot be determined at a single point’
in time. The species·area curves derived from retrospective examination
of the frequency data resembles Christiansen°s (1981) species-isolates
curves for soil fungi. In her studies, repeated isolations of soil fungi
within one plant community continued to yield additional species with no
tendency for the species·area curve to level off. Fogel (1976) determined
the minimal sampling area for hypogeous fungi in a Douglas fir stand
during peak sporocarp production to be 100 mz. Arnolds (1981) concluded
that fungal species numbers continued to increase in grasslands at plot
sizes up to 400 mz and that a plot size of 1000 mz may be preferable to
ensure an adequate sample size.
The number of ectomycorrhizal species fruiting in a small area may
reflect the minimal number of available niches in the rhizosphere. But
few estimates of ectomycorrhizal species density in small (<10 mz) areas
are available. Several species of ectomycorrhizal fungi can occupy a very
small root surface area (Zak and Marx, 1964). Up to seven ectomycorrhizal
fungi have been isolated from a single four-year old Ring; glljggtji tree
(Zak and Marx, 1964). Deacon gt gl. (1983) reported at least five types
of ectomycorrhizae occurring within a seven m radius of a young birch.
These estimates of species density are within the range of the maximum
of seven species of fungi fruiting in a single 2 x 2 m quadrat reported
here.
Direct and indirect gradient analysis, and other multivariate
techniques could be adapted readily to the description of macrofungal
Chapter 1 42
communities. Ordination has been used to compare communities of yeasts
(Bow1es and Lachance, 1983) and soil fungi (Christensen, 1982). In this
study, PCA ordination clearly differentiated between the fungal
communities of spruce and hardwood forests, emphasized the similarities
in species composition among spruce plots and the dissimilarities in
fungal species composition among hardwood plots. In addition, it
demonstrated how strongly fungal species composition of the plots was
influenced by tree species composition.
Frequency estimates must be interpreted with caution. Frequency
depends upon the pattern and density of the individuals (or in this case
sporocarps), the size and shape of the quadrats (Greig-Smith, 1983), and
in this study the duration of the observations of the quadrats. The
resolution achieved by frequency can be increased or decreased by varying
the quadrat size. Sporocarp frequency cannot measure the true extent of
a mycelium or distinguish between sporocarps produced by a large
continuous mycelium or many cluster of small localized mycelia. Sporocarp
frequency does not provide information on fungi that do not produce
epigeous sporocarps. Frequency may give an impression of the horizontal
distribution of mycelia but indicates nothing about their vertical
distribution.
Sporocarp densities of the spruce plots were within the range for
sporocarp densities of ectomycorrhizal fungi in other coniferous forests.
Sporocarp density in the spruce plots ranged from 319 to 19,180 sporocarps
ha" yr". Richardson°s (1970) estimates ranged from 8750 to 20250
Chapter 1 43
sporocarps ha" yr°‘, and Fogel°s (1976) estimates ranged from 11052 to
16753 sporocarps ha"yr'“
The sporocarp densities of the hardwood plots
in this study were lower than for other coniferous forests. Sporocarp
density in the hardwood plot ranged from 352 to 1081 sporocarps ha°‘yr°‘.
An impression of the overall pattern of fruiting phenology is difficult
to gain from comparison of the three years. The late summer drought of
1981 and 1983 contributed to the high Variation in phenology. The 1982
season had adequate rainfall during July and August and presented a
phenology pattern similar to those described for other temperate and
boreal plant communities (Wilkins and Harris, 1946; Lange, 1948;
Richardson, 1970; Petersen, 1977). All these studies show the typical
strong peak of productivity near the end of the season.
_l
The length of the fruiting season for ectomycorrhizal fungi at
high-elevations in West Virginia appears to be relatively short compared
to fruiting seasons of ectomycorrhizal fungi at lower elevations or in
Amaritime climates but is commensurate with those in high-latitude,
low-elevation communities. Sporocarps of ectomycorrhizal fungi were
observed in both spruce and hardwood plots from July to early October
(75-91 days). Wilkins and Harris (1946) reported a fruiting season
lasting from August to November in an English pinewood and from June to
November in a beechwood. In Scotland, ectomycorrhizal fungi fruited from
June to December in a Scots pine plantation (Richardson, 1970). In.R1ggg
yiggigiggg stands in southwestern Virginia (elev. 450-750 m),
ectomycorrhizal fungi fruit from June to December (author°s unpublished
Chapter 1 44
data). The fruiting season is much shorter in Greenland tundra (Petersen,
1977), where ectomycorrhizal fungi fruit from July to mid-September.
Although only one additional species was found after the third year,
it is difficult to judge how many additional species would be found if
more years had been sampled. Fogel (1976) sampled 98 % of the
hypothetical number of hypogeous species of a Douglas fir forest in a
three year period. Arnolds (1981) observed most of his grassland sites
for three years but observed some selected sites up to six years. Based
upon these six year observation periods, he concluded that three years
of sampling yielded 75-92 % of the total species. However, during these
extended sampling periods, the Vegetation of these grasslands changed
significantly, which may have contributed to subsequent additions to the
mycoflora. Lange (1978) recorded 266 mycorrhizal species in a series of
Vegetation types over a ten-year period in the Beech Wood District of
Denmark. During any given season, 21-S9 % of these species were observed
to fruit.
In summary, this study provides baseline data on the diversity,
density, frequency, and phenology of ectomycorrhizal fungi fruiting in
red spruce and northern hardwood forest types. The fungal species
composition and sporocarp density of the two forest types were distinctly
different because of a strong dependence of fungi species composition on
the composition of tree present. Inter-plot similarities and differences
in fungal species composition and the dependence of fungal species
composition on the tree composition was emphasized by PCA, one of many
Chapter 1 45
ordination techniques which may be adapted to future studies of fungal
communities in response to vegetational or environmental gradients or
identifying fungal community structure in complex Vegetation landscapes.
Fungal species composition and relative abundance of fungal species in
other red spruce or northern hardwood communities may shift with changing
latitude, or with slight changes in tree composition.
Species species richness was not significantly different between
‘spruce and hardwood plots, the hardwood plots were more diverse because
of greater equitability among species. Evidence for this greater
diversity included the Shannon-Wiener index, the dominance diversity
curve, species area curve (indicating more species among hardwood plots,
and PCA ordination.
The results suggest that estimating sporocarp frequency in small
contiguous quadrats may be a more appropriate method than estimating
sporocarp density for comparing the the relative activity or ubiquity of
fungal species in plot studies. Fruiting phenology was comparable between
the two forest types, and the fruiting season was relatively short. The
fruiting season at this southern latitude was compressed because of the
short growing season at high-elevation. Variation in fruiting season is
assumed to occur in response gradients of altitude or latitude, but
quantitative, comparative studies are needed to measure the extent of this
Variation. _
Chapter 1 46
QHAHIEBL. S.BAI1AL2AIII":1RhJ§ANDQE
IHKE12$.BB§l£EAhIDIüBHW9QDEQRE§IS
Basidiomycetes usually fruit in spatially and temporally aggregated
patterns. Spatial aggregation is caused by multiple sporocarp production
by a single or a series of localized mycelia. Heterogeneity of habitat,
inoculum density, and non—random occurrence of host plants and substrates
contribute to the non-random patterns of mycelial occurrence. Temporal
aggregation is caused by repeated sporocarp production by perennating
mycelia. Basidiomycete fruiting is often described in the literature as
gregarious, clustered, in groups, or caespitose. The perennial nature
· of fruiting is best exemplified by the mushroom collector who returns
year-after-year to the same location for edible fungi. Perennial fruiting
has been expressed quantitatively as "constancy" (Lange, 1948) or as an
"index of specific fluctuation" (Arnolds, 1982). The spatially
aggregated, perennial, fairy ring fruiting pattern of certain grassland
fungi has been thoroughly investigated (Ingold, 1974; Smith, 1980;
Edwards, 1984). However, relatively little is known about the spatial
patterns of terrestrial sporocarps in forests, and variation of these
patterns within or among species (Fogel, 1981)
Observation of spatial patterns of sporocarps in reference plots
yields several kinds of information related to the biology of the fungus
Chapter 2 47
and its associated organisms. Sporocarps verify the presence of the
vegetative mycelium, and sometimes sporocarps numbers and their patterns
have indicated the spatial pattern and relative abundance of the
vegetative mycelia (Laiho, 1970; Thompson and Rayner, 1982; Last gg gl.,
1983; Edwards, 1984; Newell, 1984; Cotter and Bills, in press). In
addition, the spatial patterns of sporocarps may influence the spatial
patterns of animals that utilize sporocarps as feeding or breeding sites
(Shorrocks and Charlesworth, 1982; Ashe, 1984; Lacy, 1984). Knowledge
of spatial patterns of sporocarps is essential to the understanding
inoculum density and dispersal, and of the establishment of °
Basidiomycetes in forests.
How should fungal communities be sampled to yield maximum information?
Ideally, as in plant or animal communities, quadrat sizes and placement
would be determined by the species-area relationships within the
community, the relative sizes of the organisms, and the Variation in the
spatial patterns of the organisms. These critera are difficult to apply
to fungi because the composition of the community and its spatial pattern
based on sporocarps is continually varying (Fogel, 1981). Repeated and
prolonged observations of fungal communities are needed because of the
temporal Variation in their compositions. In addition, the lack of
knowledge of variation in sporocarp density and the unpredictable nature
of species density has prevented the application of consistent sampling
methods with subsequent difficulties in comparing and interpreting
results of different investigators (Hueck, 1953). Knowledge of spatial
Chapter 2 48
variation of fruiting patterns can be applied to further improve sampling
· methods for fungal communities.
Quantitative methods for describing spatial patterns of plants or
animals have rarely been applied to higher fungi (Fogel, 1981). Recently,
the spatial patterns of plant pathogenic fungi and diseased plants have
been investigated in several agricultural systems (Campell and
and Amggjgg igggrggg usually fruited singly or in small groups. High
patchiness was characteristic of species producing localized, dense
clusters of sporocarps with the clusters separated by large intervening
areas where either sporocarps were absent (e.g. Lgggggigg ggmphgggggg,
lgggyhg gghgjng) or uniformly present at low densities (e.g. Qlgggljgg
g;i;;g;g) (Figs. ll, 13). Species with a low uniform sporocarp density
(e.g. Amggjgg jggggggg) or high uniform density (e.g. Lgggggigg
ggglgggg) were characterized by low patchiness indices.
With the exception of Qlgggligg grjggggg, the V/m and mean crowding
of the major hardwood and spruce species were comparable. However, the
patchiness indices were much higher for all hardwood species because the
gregarious sporocarp production of Lgggggjg lgggggg, Rg;;g1g ggggglggg,
Lgggggjgg gggphg;g;g;, and Sglgrgggrmg gigrjggm was restricted to thel
rhizospheres of the few {ggg; and Qgggggg trees and ßglggjggllg;
mgggligigg; around the {rggiggg trees. Conversely, exclusion of
ectomycorrhizae from the areas dominated by endomycorrhizal tree species
and ferns may have contributed to the patchy distribution of sporocarps
in the hardwood forests.
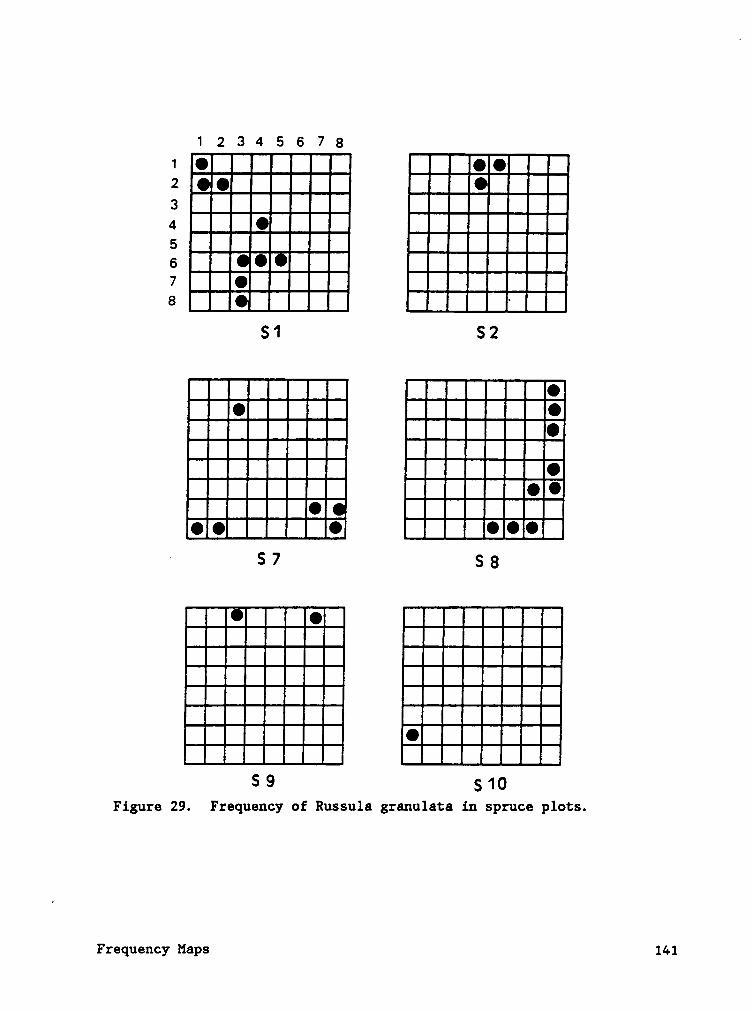
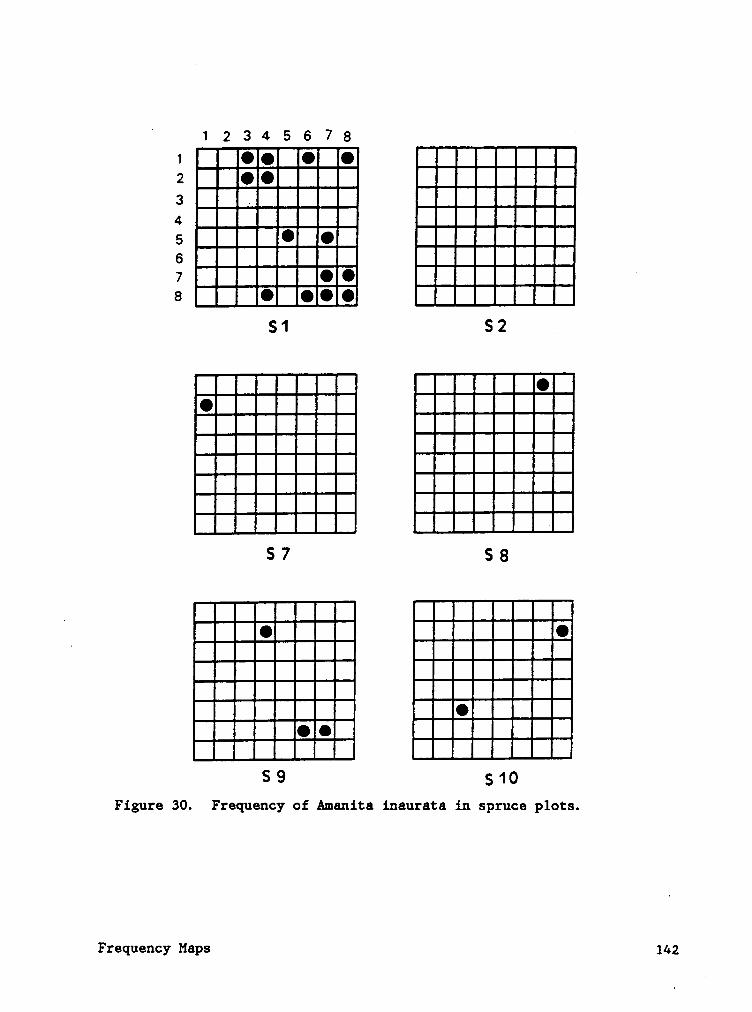
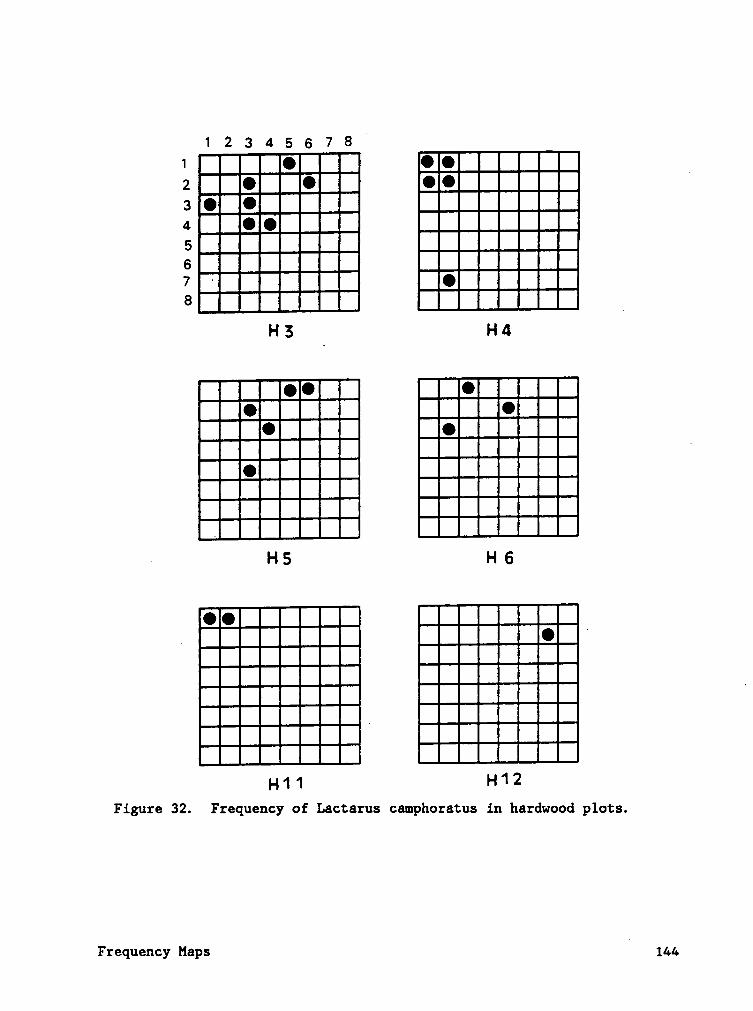
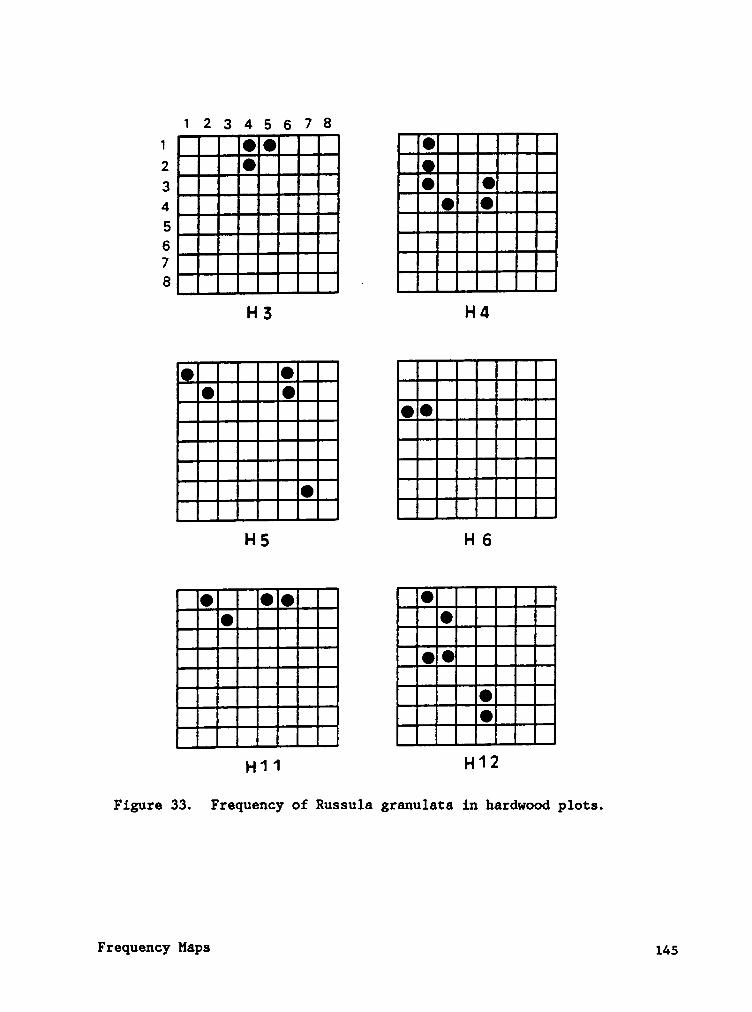
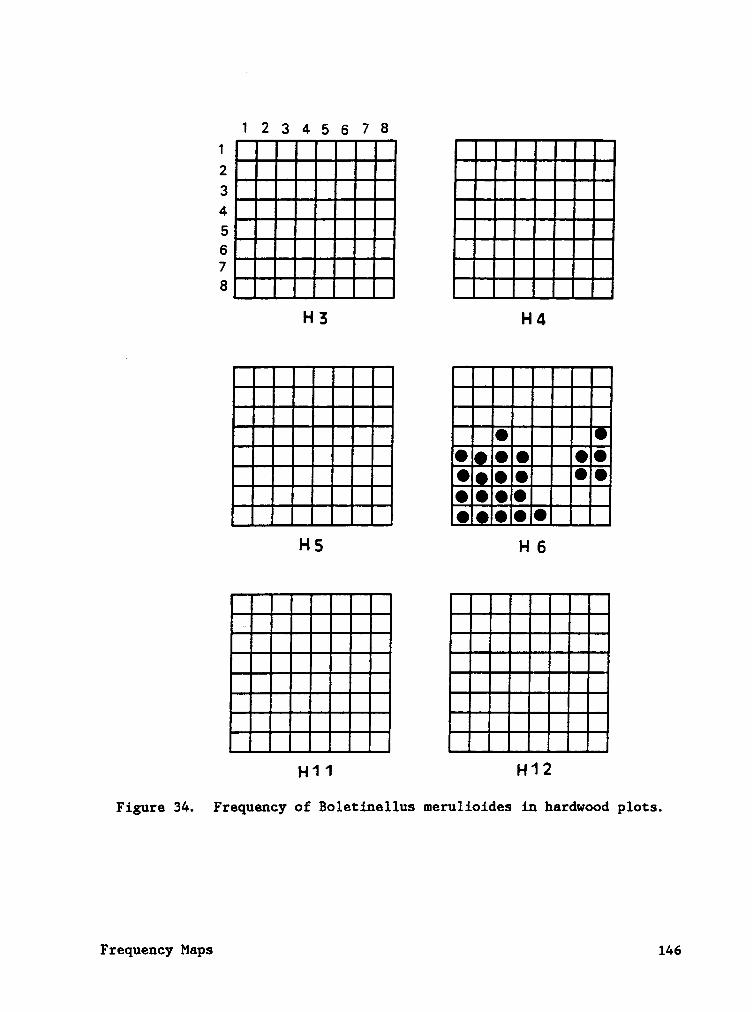
Frequency maps (Appendix B) indicate that species often occured in
contagious patterns. The contagious sporocarp frequency of ßglggjggllgg
ggggljgiggg (plot H6) was caused by two individual mycelial patches
producing sporocarps (Cotter and Bills, in press). Sporocarp density of
Chapter 2 53
1.6
1.2
OE<IZ
0.6>o6O.J
0.•
Orocc LV nu LO AF LC LL HG 88 ^*
SPECIES
Figure 8. Logl0 V/m ratio of major species in spruceforests.: Species initials are from Table 10. A11 log10V/m ratios were significantly greater than 0.
Chapter 2 S4
00
es
OEO2O so1UZ<LU2
es
V0‘ cc LO LV uu AF LC LL as nc; AI
SPECIES
Figure 9. Mean crowding of major species in spruceforests.: Species initials are from Table 10.
Chapter 2 55
II
10
U7V7 .UJZI
_ UI-g | .
0 LC nu cc LV AF R AI LL es LO
SPECIES
Figure 10. Patchiness of major species in spruce forests.: Speciesixxitials are from Table 10. .
Chapter 2 56
1.0
\‘
°_ 0 aésséaäsLw¤M LC nc Sc
SP ECIES
Figure 11. Logl0 V/m ratio of major species in hardwoodforests.: Species initials are from Table 10. All log10V/m ratios were significantly greater than 0.
Chapter 2W
57
s
z ’-,;;:;L-<L
Lg am L c n 6 s c
S PE C I E S
Figure 12. Mean crowding of major species in hardwoodforests. : Species initials are from Table 10.
Chapter 2 58
•c
E man‘°0•-¤
Eiägigä =E=i;E;E3E =E=E=E=E=· :i=i=i=E=Lg L c n 6 a M s 6SPECI ES
Figure 13. Patchiness of major species in hardwoodforests.: Species initials are from Table 10.
Chapter 2 59
ß. mgggljgiggs in one quadrat was clearly dependent on sporocarp density
in nearby quadrats. Although the mycelial pattern was not be directly
mapped for other species, the hypothesis that the occurrence of sporocarps
in a quadrat depended on the presence of sporocarps in nearby quadrats
rather than being randomly or regularly dispersed was tested using spatial
autocorrelation. Each major species, except ßglggggggm giggjggm, in both
hardwood and spruce forests was positively autocorrelated in at least one
of the plots in which it occurred (Tables 13, 15).
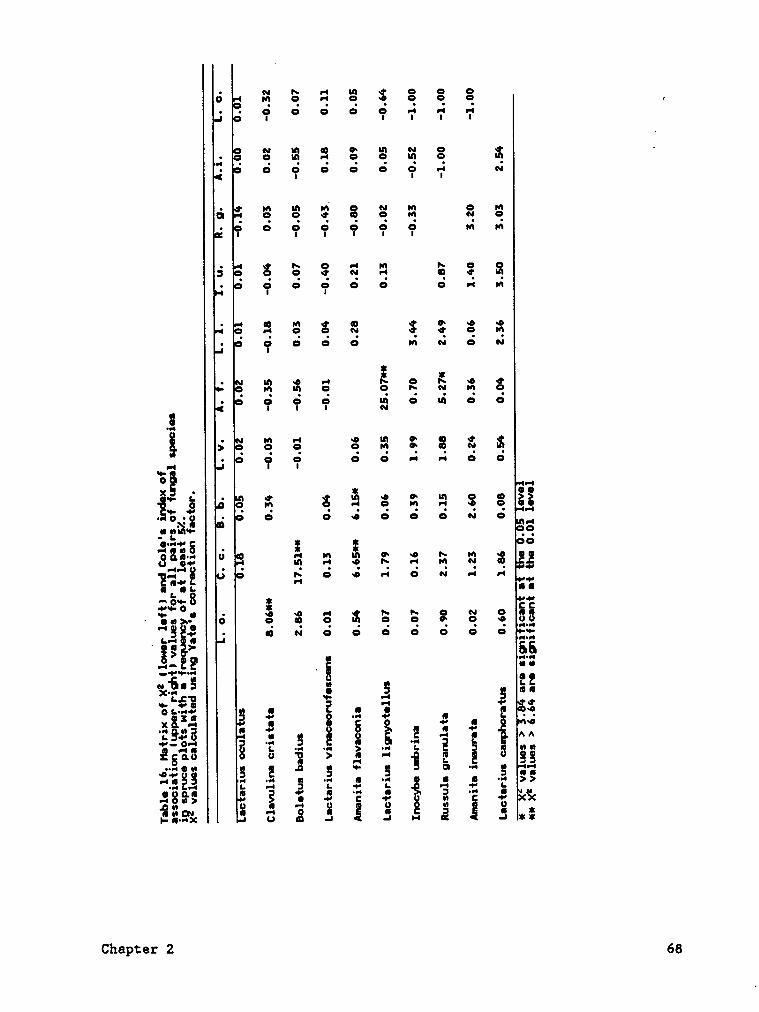
Most pairs of major species in the spruce forest were distributed
independently of each other as indicated by the X2 of the 2 ¤ 2
contingency tables and Cole's index (Table 16). However, :ignificantly
more co·occurrences than expected were detected in three speciespairsA.
flgyggggig). Significantly fewer co·occurrences than expected were
detected in three other species pairs (A. flgyggggjg - Q. ggjstggg, A.
b 0.001 < P =< 0.01 .c P =< 0.001· nof enoudu frequency classes for test
Chapter 2
i61
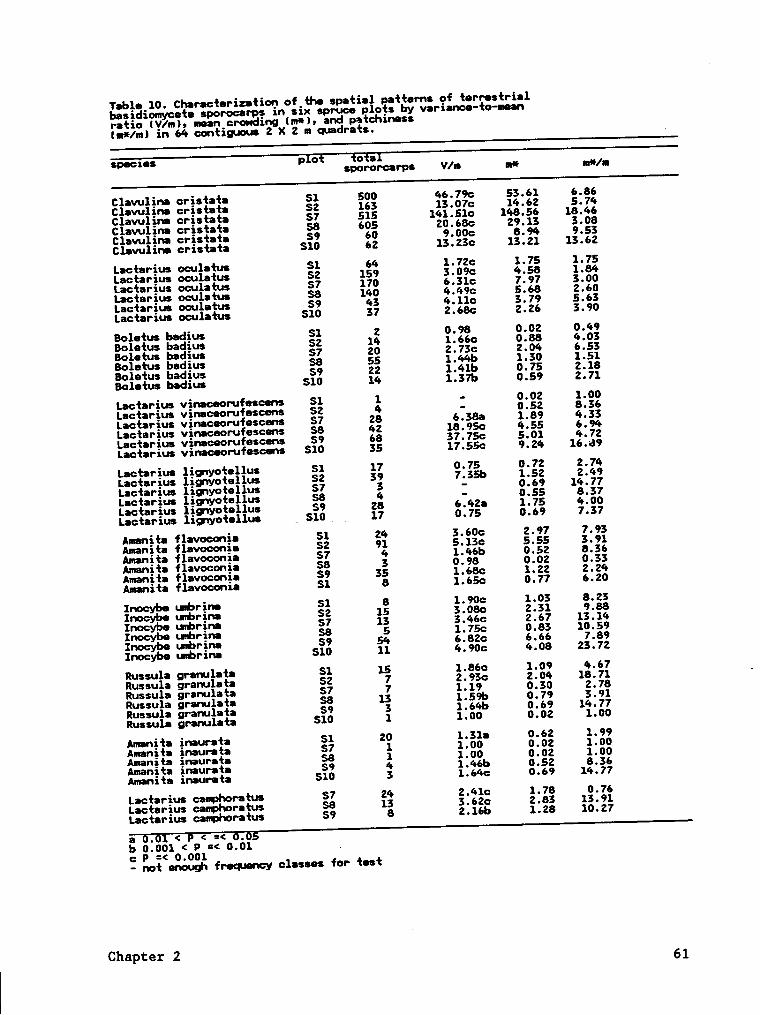
Tab1•_11. Charachrizafiop of_fh0 spafial Yntfems of_hrr•s·!ri•1bas;d1omyce‘l:• gporocarps xn sgx harduoogl p ots_b¥ yar~1;n0•-fo-mean·_ raho IV/nl, X ?o0<h•ss·ofyf1·I:_of ag Poxsson dxs rxbufxon _on {hs obs•rv0d_ raqusncy dgsirzbufxoq of sporocarpm mean crcwdxng(nä), and p•t0h1.n•ssI¤e*/ml xn 64 contxguous 2 X 2 n quadrats.
Table 12. Significance tests for spatial autocorrelation of- major basidiomycete species in spruce plots.
Frequency (freq) is the number of quadrats a species occupies.S.N.D. is the standard normal deviate.Occupied quadrats (BB), occupied and empty quadrats (BW),and empty quadrats (WW) were joined by queen°s moves.
species freq no. of S.N.D.plots BB BW WW
Lggtggigg ggglgggg 179 6 2.92c -3.19c 1.64a
Qlgygligg ggiggggg 148 6 4.46c -6.52c 4.81c
ßglgggg hggjgg 79 6 1.90a -4.32c 3.93c
Las:.a:.um 67 6 7-35<= ·S•21<= 1.916
Aggjtg flgyggggjg 64 6 2.44b -4.31c 3.64c
ljggygtgllgg 57 6 5.80c -2.51b -0.17
lgggygg gmbgjgg 38 6 4.28c -1.12 -0.36
Rgggglg gggglgtg 30 6 4.54c -5.56c 4.36cl
Amggita jgggggtg 21 5 1.90a -2.48a 1.89a
Lggtggigg ggmphgggggg 19 3 2.15b -2.39b 1.78a
a 0.05 < P S 0.10 ·b 0.01 < P S 0.05c P S 0.01
Chapter 2 63
Table 13. Significance tests for spatial autocorrelation ofmajor basidiomycete species in hardwood plots.Frequency is the number of quadrats a species occupies.S.N.D. is the standard normal deviate.Occupied quadrats (BB), occupied and empty quadrats (BW),and empty quadrats (WW) were joined by queen°s moves.
species freq no. of S.N.D.plots BB BW WW
Lggtggiggyggmphgggggg 23 6 4.69c -2.72c 1.57
Rgggglg ggggglggg 23 6 3.49c -0.91 0.01
ßglggjggllgg mg;g1;gi§g5 23 1 6.47c -7.92c 5.08c
Sslsmdema szitxinum 21 2 1- 47 -1- 20 0- 46
Lgggggjg lggggtg ° 17 6 4.13c -2.51b 1.54
a 0.05 < P S 0.10b 0.01 < P S 0.05c P S 0.01
Chapter 2’
64
Table 14. Significance tests for spatial autocorrelation ofmajor basidiomycete species in spruce plots.
_Frequency (freq) is the number of quadrats a species occupies.S.N.D. is the standard normal deviate.Occupied quadrats (BB), occupied and empty quadrats (BW),and empty quadrats (WW) were joined by the inverse of thedistance squared between their centers.
Table 15. Significance tests for spatial autocorrelation ofmajor basidiomycete species in hardwood plots.Frequency is the number of quadrats a species occupies.S.N.D. is the standard normal deviate.Occupied quadrats (BB), occupied and empty quadrats (BW),and empty quadrats (WW) were joined by the inverse of the ,
. distance squared between their centers.
species freq no. of S.N.D.plots BB BW WW
Lagtggigg ggmghgrgggs 23 6 3.75c -2.51c 1.58
Ryggylg ggggglgtg 23 6 2.71c -1.05 0.34
ßglgtjggllgg mgggligjjgg 23 1 6.47c •5.88c 3.76c
ßglgggggggg gjtgjggm 21 2 » 1.66a -0.64 -0.33
Lgggagjg lggggtg 17 6 3.19c 3.19c 1.34
a 0.05 < P S 0.10b 0.01 < P S 0.05c P S 0.01
Chapter 2 66
Only one species pair (L. gamphgrgtus - R. grgnylgtg) in the hardwood
forests was positively associated (Table 17). However, quadrats occupied
by L- samvlxuamu. E- z:.¤m1l¤.ta„ L- lasaeata. and S- sitrinum after:
occurred near each other in plots H4, H11, and H12 probably because their
sporocarps arose from mycelia associated with the same [ggg; and Qgggggg
trees. The lack of co-occurrence among these species might have been
because the quadrats were too small include more than one or two species.
When 2 x 2 quadrats were combined into 4 x 4 quadrats, the co-occurrence
of these species still did not deviate appreciably from expected values.
The statistical significance for co-occurrence could not be tested‘
because of low expected values obtained when the quadrats were combined
(1 to 3).
The hypotheses that all species of the spruce forest type (27) and all
the species of the hardwood forests type (36) were distributed
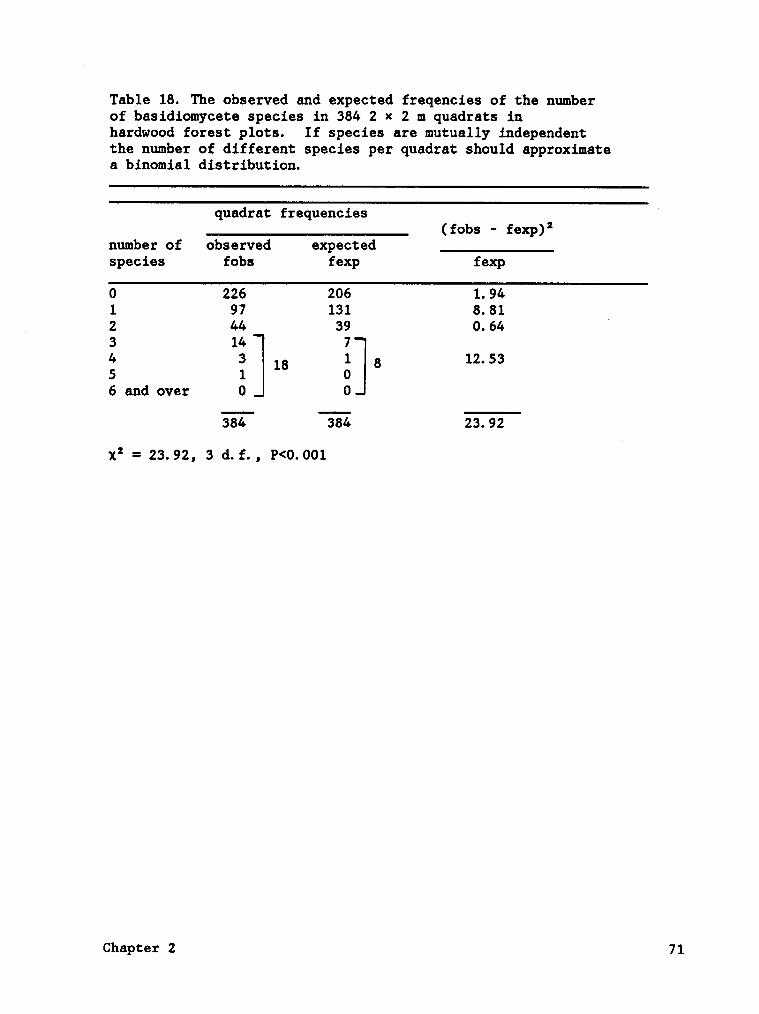
independently among the quadrats were rejected (Tables 18, 19). In the
spruce forest, fungi were absent in more quadrats (54) than expected under
the null hypothesis and fewer quadrats had four or more species than
expected (Table 18). The species of the hardwood forest type appeared
to be aggregated in certain locations and absent in others because no
fungi occurred in more quadrats than expected and two or more species
occurred more quadrats than expected (Table 19).
Chapter 2 67
°r· S'. '6 S ä Q
°
• • •
6
°
H,,‘
T, •
9Ü
°4 I I
_
N III ¢ Q2,0 In -1 Q , , . J1‘
6 => Q° 9 9
.Q NI. ··· ··· 2 S 2 2 Q QÜ Q Q • • •
6 Ü 44. ' Q 9 9 9 9 ¤I
A 0 ¤. S 'a S a' 2 Q Q Q¤ • , • •6
°
pl Ü. 9 ° 9
0 ~• ~0. 2 ä S 2 $ Q Q -2, • •° N
_ ' 6 0 0 ¤ '^ N QI
{ zu 0. Ih ~¤ •'*S Ä Z InIn G , • •
'°
, • • •In
°In Q. 9 Q Q u••
•••
Q. 0. an g S :2 °;QH. I I4-4gg
=· Q ää0 III ¢x_•_
c , g g 2 -1 *2 Q •-•°§,•_ ,3 •6 6 J
°Q Q N Ö
S•••¤•O
°••
.·•0:}+{ ä °
'.A M *2-2:*58 ' ·•
-^ Ä 3 M °f Q fg°¤_Q•« U gg H *0 , •*4 , • •
,4 rlQ-I3
· ' rl ¤ ·• "‘ Q Q ·‘°§.-IEgtäga0 N
°
$—•Q°6· _ ä ~e g 5 S '6 vg Q *2 S.;~ $0 - °Q Q
·‘
6 6 ¤° ° ..3-°•
MSE: ° ° N ° ° ssg.-I R«.••
' gß > ·g>-·:....§ ° Ü„
g ÜNan I ,2 •-I 4-• 0"°C.c"' [ ‘°••— ·•-.2
Q ·•Q •‘ .~Ä°"'?‘·-
Ü T- Ü § ° E Q 3 Ü ^^.5 0%
'l:
U E ••• Iä2 00
L 'V un U·
.;• L = 8+--0-** I. ·•• "‘> I-I*‘¤I
1 > Ü L ,.4••••°U:
·
,,-
·L
0-H „ .••Ü ‘:
*4, "I 2:
·
N-***7 L -·• 7 ...• • Q -•« X...• •-·• +• n 0 C
¤.•
"g • ·•·* ¢
" eo°
080 U
E Q I
—• ¢> Q •• ··• „ • 7< .; xx
JW g H ,4 H Ünum U -¤}.·••¤•X
68Chapter 2
Table};. Hafrix of $2 l•f$) •n•i|1Co1q'• igdex afl _ _-assocxa mn ¤.pp•r rx va ues or a aus o mga s usig hardnood {lots wifgia frequency of at IeasfpxX
values ca culated usmg Yate's correchon factor.
E. c. E. 9. E. nn. S. c. C. I.
Lachrius cauphoratus 0.21 -1.00 -1.00 -1.00
Russula grnnulata I3.96! -1.00 -1.00 0.02
Bolefimllus narulioides 2.90 2.90 -1.00 -1.00
$cl•r0der¤•• eifrimn 2.76 2.76 2.76 0.13 _
Laccaria laccata 2.52 0.29 2.52 2.94
·l Rz vaIu•s > 6.04••·•
$1§11¥l¢3h{ a¥ TF: 0.0I I•v•I
Chapter 2 69
Mosaics are patterns resulting from different areas of a plane having
different properties (Pielou, 1974). Continuous colonies of vegetatively
propagating plants are often considered as vegetation mosaics. The
spatial patterns of sporocarps can also be envisioned as species forming
overlapping mosaics of different sizes with different densities of
sporocarps within the mosaics. The presence and absence of a mushroom
species forms a two-phase mosaic while the combined presence and absence
of several species forms a multi-phase mosaic. The boundaries of the
mosaic phases are not well·defined because of the variation in sporocarp
densities and, as with the vegetative clones of higher plants, mosaics
of mushrooms were variable in size and shape. The clonal nature of
adjacent sporocarps was not obvious, however, because scattered, widely
separated individual sporocarps may be connected by a continuous
widespread mycelium or they may arise from small widely scattered mycelia.
Mosaic patterns may form in response to either the heterogeneity of
the biotic or abiotic environment. Abiotic factors might include
subsurface rock, soil depth, soil moisture, or canopy cover. Biotic
factors might include the availability of colonizable host tree roots,
physical exclusion by large tree roots or rhizomes of herbaceous plants,4
predation by soil invertebrates, or antagonistic or synergistic
interactions with other rhizosphere organisms. Measurement of the size
of the fruiting mosaics was often limited by plot boundaries.
Chapter 2 70
Table 18. The observed and expected freqencies of the numberof basidiomycete species in 384 2 ¤ 2 m quadrats inhardwood forest plots. If species are mutually independentthe number of different species per quadrat should approximatea binomial distribution.
quadrat frequencies '
(fobs - fexp):number of observed expectedspecies fobs fexp fexp
0 226 206 1. 941 97 131 8. 81
7 2 44 39 0. 64”
3 14 74 3 1 8 12.53S 1
180
6 and over 0 0
384 384 23.92
X2 = 23.92, 3 d. f. , P<0. 001
Chapter 2 71
Table 19. The observed and expected freqencies of the numberof basidiomycete species in 384 2 x 2 m quadrats inspruce forest plots. If species are mutually independentthe number of different species per quadrat should approximatea binomial distribution.
Copies of the raw data sets for fungal species and tree species are
obtainable from the author, Dr. Orson K. Miller, Jr. , and Dr. Golde I.
Holtzman. An additional set was deposited in the New York Botanical
Garden Archives.
Appendix D. Raw data 1981-83. 162
Bibliography163
I
__l____l_. 1939. Two new species of Rugsggle together with the sporeornamentation of some of our American Russulas. Myeglegie 31:490-498.
Cain, S. A. 1938. The species-area curve. Age;. Mid. Net. 19: 573-581.
Campbell, C. L. and S. P. Pennypacker. 1980. Distribution of hypocotylrct caused in snapbean by Bhizettenie selmi- 70:521-525.
Christensen, M. 1981. Species diversity and dominance in fungalcommunities. ln: Ihe fgngel eegggnjty. Eds D. T. Wicklow and G. C.Carroll. Marcell Dekker, Inc., New York. pp. 201-232.
Chui, W. F. 1945. The Russulaceae of Yunnan. Lleygie 8: 31-59.
Cliff, A. D. and J. K. Ord. 1973. Sgetjgl egtgee;;elet1g¤. Pion, London. ·175 p.
Cole, L. C. 1949. The measurement of interspecific association. Eeglegy30: 411-424.
Core, E. L. 1966. Yegetetjen gf lee; Y1;g1gie. McClain Printing Co.,Parsons. 217 p.
Cotter, H. V. T. and G. F. Bills. in press. Comparison of the spatialpatterns of the sexual and vegetative states of ßeletjgellge
Davis, M. B. 1983. Holocene vegetational history of the eastern UnitedStates- Pp- 166-181- In:. ef theunited Staus,. hl,. 2... Ihe Heletene- Ed- . H- C- Wright- Universityof Minnesota, Minneapolis.
Deacon, J. W., Donaldson, S. J., and F. T. Last. 1983. Sequences andinteractions of mycorrhizal fungi on birch. Ble; gg ßeil.71:257-262.
Dey, J. P. 1976. Phytogeographic relationships of the fructicose andfoliose lichens of the southern Appalachian Mountains. Pp. 398-416.In: hiuerx ef the hieta ef the sedthern
hrt lu- Algae end thai- Eds-. B- 0- Parker and M-K. Roane. University of Virginia, Charlottesville.
. 1978. Fructicose and foliose lichens of the high-mountain areasof the southern Appalachians. ß;gglegjet 81: 1-93.
Dominik, T. 1961. Studium nad mikotrofizmem swierka pospolitego -hp1.Pice excelsa (Lam.) Lk. w Polsce. lnete Bee, Leege £;„ 1961,209: 1-24.
Bibliography 165
\
Edens, D. L. 1973. The ecology and succession of Cranberry Glades, WestVirginia. Ph. D. Dissertation, North Carolina State University,Raleigh.
Edwards, P. J. 1984. The growth of fairy rings of Agggigg; ggggy;}; andtheir effect upon grassland vegetation and soil. J. Egg}. 72:505-513.
Fedde, G. F. 1973. Impact of the balsam wooly aphid (Homoptera:Phylloxeridae) on cones and seed produced by infested Fraser fir.Qgygg. Eyggmg}. 105: 673-680.
Fogel, R. 1976. Ecological studies of hypogeous fungi. II. Sporocarpphenology in a western Oregon Douglas Fir stand. Qgy. J. Egg. 54:1152-1162.
. 1981. Quantification of sporocarps produced by hypogeousfungi. }y Ihg fgygg} gggmgyigy. Ed., D and G. C. Carroll. Marcell
- Dekker, Inc., New York. pp. 553-568.
Gauch, H. G. 1982. Multivariate analysis in community ecology. CambridgeUniversity Press, New York. —
Giltrap, N. J. 1982. Production of polyphenol oxidase by ectomycorrhizalfungi with special reference to Lgggggig; spp. Iggy;. Egig. Mygg}.Egg. 78: 75-81.
Greig-Smith, P. 1983. Quantitative plant ecology. 3rd. Ed. University ofCalifornia Press. Los Angeles. 359p. ~
Hering, T. F. 1966. The terricolous higher fungi of four Lake Districtwoodlands. Iggy;. Eg. Mygg}. Egg. 49: 369-383.
Hesler, L. R. 1945a. Notes on Southern Appalachian fungi. VI. J. Igyyg;;ggAggg. Eg}. 20: 233-2
. 1945b. Notes on Southern Appalachian fungi. VII. J.Igyyg;;gg. Aggg. Eg}. 20: 363-372.
. 1960. Mushrooms of the Great Smokies. University ofTennessee Press, Knoxville. 289 p.
____________. 1960. A study of Eg;;g}g types. Mgm. Iggggy Egg. Q}gg 21:1-59.
. 1967. The genus Eygg}ggg in southeastern North America.B.e.ih- Nam Haduidaia 23= 1·196-
and A- H- Smith- 1979- Nnnth Anuisau sp.¢.¢.i¤ 9.£Lgggggig;. University of Michigan Press, Ann Arbor. 650 p.
Bibliography 166
Homola, R. L., and P. A. Mistretta. 1977. Ectomycorrhizae of Maine, Alisting of Boletaceae with associated hosts. Life Sciences and
A Agriculture Experiment Station Bulletin 735. Univ. Maine, Orono.
____________, and M. M. Czapowskyj. 1981. Ectomycorrhizae of Maine 2. Alisting of Lgggggigg with the associated host AgricultureExperiment Station Bulletin. 779. Univ. Maine, Orono.
Hongo, T. 1960. The Agaricales of Japan I-3. Russulaceae. AgggEhyggggxgg. Geobot. 18: 129-146.
_________, and K. Yokoyama. 1978. Mycofloristic ties of Japan to theA continents. Mgm. ßhigg Qgiy. Egg. Aggg ggi. 28: 76-80. 545-574.
_ Hora, F. B. 1959. Quantitative experiments on toadstool production inwoods. Igggg. E;. Myggl. ßgg. 42: 1-15.
___________. 1960. New checklist of British agarics and boleti part IV.Validations, new species and critical notes.I;ggg. ßgjg. Myggl.§gg. 43: 440-459.
Hueck, H. J. 1953. Myco-sociological methods of investigation. Vggggggig4: 84-101.
Ingold, C. T. 1974. Growth and death of a fairy ring. ßgll. ßgjgg Mgggl.Sgg. 8: 74.
Johnson, N. K. 1975. Controls of numbers of bird species on montaneislands in the Great Basin. Eyglggjgg. 29: 545-57
Kauffman, C. H. 1917. Tennessee and Kentucky fungi. Mggglggjg 9: 159-165.
Knudson, H., and T. Borgen. 1982. Russulaceae of Greenland. Pp. 216-243.lg: Agggjg ggg gjgjgg gygglggy. Eds., G. A. Laursen and J. A.Ammirati. University of Washington, Seattle.
Korhonen, M. 1984. Sgggggg gggkgg. Otava, Helsinki. 223 p.
Kornerup, A., and J. H. Wanscher. 1978. Mgghggg hggggggk gf gglgg;. 3rdEd. E. Methuen and Co., Ltd., London. 252 p.
Korstian, C. F. 1937. Perpetuation of spruce on cutover and burned lands1
in the higher southern Appalachians. Eggl. Mgggg;. 7: 125-167.
Kühner, R. 1975. Agaricales de la zone alpine. Genre Lgggggigg D. C. exS. F. Gray. ßgll. Sgg. Myggl. Eggggg 91: 5-69.
Bibliography 167
Lacy, R. C. 1984. Ecological and genetic response to mycophagy inDreeephilidee (ßiptere)- ln:
in reales! and e1o.1m.ie¤- Eds- , Q- Wheeler and M-Blackwell. Columbia University Press, New York. pp. 286-301.
Lange, M. 1948. The agarics of Maglemose. De. Bee. Agk. 13: 1-141.
_. 1978. Fungus flora in August, ten years observations :L¤ aDanish beech wood district. ße;. Ileeek;. 73: 21-54.
Last, F. T., P. A. Mason, J. Wilson, and J. W. Deacon. 1983. Fine rootsand sheathing mycorrhizae: their formation, function and dynamics.Elan; end Seil 71: 9-21-
Laursen, G. A. and J. A. Ammirati. 1982. Lactarii in Alaskan arctictundra. le: Ageele ege eleige myeelegy Eds., G. A. Laursen and J.A. Ammirati. University of Washington Press, Seattle. pp. 245-281.
MacArthur, R. H., and E. O. Wilson. 1967. Ihe eheegy ef lelegehlegeeggeehy. Princeton University, Princeton. 61: 909-916.
Mclntosh, R. P., and R. T. Hurley. 1964. The spruce-fir forests of theCatskill Mountains. Eeelegy 45: 314-326.
Miller, 0. K., Jr., 1983. Ectomycorrhizae in the Agaricales andGasteromycetes. Qeg. 1. ßee. 61: 909-916.
Muel1er·Dombois, D., and H. Ellenberg. 1974. Aims and methods ofvegetation study. Wiley, New York.
Neuheff, W- 1956- Die Milehlinze Laereriue)- Bed HeilbrrmnObb., Julius Klinkhardt. 248p.
Newell, K. 1984. lnteraction between two decomposer basidiomycetes and acollembolan under Sitka spruce: distribution, abundance andselective grazing. Sell ßlel. ßleehem. 16: 227-233.
Nicot, P. C., D. I. Rouse, and B. S. Yandall. 1984. Comparison ofstatistical methods for studying spatial patterns of soil bornRhyeeeeehelegy 74: 1399-1402.
Noy-Meir, I. 1973. Data transformation in ecological ordination. I. Someadvantages of non-centering. 1. Eeel. 61: 329-341.
Oosting, H. J., and W. D. Billings. 1951. A comparison of virginspruce-fir forests in the northern and southern Appalachian system.
_ Eeelegy 32: 84-103.
Peck, C. H. 1870 (1872). Report of the state botanist. Ageeel Bee. NegYegk Seeee Me;. 24: 41-108.
Bibliography 168
. 1884 (1885). New York species of Lgggggigg. Aggggl Egg. NggYggk Sgggg Mg;. 38: 111-133. -
Petersen, P. M. 1977. Investigations on the ecology and phenology of themacromycetes in the arctic. Mggg. Qgggg. 199: 1-72. 4
Petersen, R. H. 1976. The southern Appalachian Mountains as a refugiumfor tropical basidiomycetes. Pp. 287-295. lg: Qigggigggigggl
Alxs:2ggg fgggi. Ed. B. C. Parker and M. K. Roane. University ofVirginia, Charlottesville.
_____. 1979. Checklist of fungi of the Great Smoky Mountains„ National Park. Nat. Park Serv. Manage. Rep. No. 29.
Pielou, E. C. 1974. Rggglggigg ggg ggggggigy ggglggg. Gordon and Breach.New York. 424p.
Ramseur, G. S. 1960. The vascular flora of high mountain communities ofthe southern Appalachians. 1. Elighg Miggggll ßgi. ßgg. 76: 82-112.
Rice, E. L., and R. W. Kelting. 1955. The species•area curve. Egglggy 36:7-11.
Richardson, M. J. 1970. Studies on Egggglg gmggjgg and other agarics ina Scots pine plantation. Igggg. B;. Myggl. Sgg. 55: 217-229.
Ridgwey, R- 1912- 9.012: atandaxda and Publ- by the euther,Washington, D. C. 43 p., 53 pl.
”l
Remegnesi. H- 1967- Lea Knsaulsa dlE1::20.e at dLA£:i2¤2 du N2:d- Berdas,Paris. 998 p.
Schaeffer, J. 1952. Egggglg;Mgggg;gghig Bad Heilbrunn Obb., Juliusl
Klinkhardt. 295 p.
Shannon, C. E., and Weaver, W. 1949. Ihg ggghggggjggl ghgggy gfggggggggggjgg. University of Illinois Press, Urbana.
Shaffer, R. L. 1962. The subsection Qgggggggg of Egggglg. ßgjggggjg 14:254-284.
1964- The subseetieh ef Ruasnla- ¤y20.12z;La _56: 202-231.
______________. 1970a. Notes on the subsection Qggggggggigggjggg and otherspecies of Egggglg. Llgygig 33: 49-96. _
. 1970b. Cuticular terminology in Egggglg (Agaricales).ßgjggggig 22: 230-239.
Bibliography 169
. 1972. North American Russulas of the subsectionBzs.t.en1:.;L¤ae- Uxcelsxia 64= 1008-1053-
_____________. 1975. Some common North American species of Egggg}gsubsection Emg;}g}ggg. ßgih. Hggwjdgig 51: 207-237.
Shew, B. B., M. K. Beute, and C. L. Campbell. 1984. Spatial patterns ofsouthern stem rot caused by Sg}g;g;}gm-;g}f5}i in six North Carolinapeanut fields. Rhy;ggg;hg}ggy 74: 730-735.
Shorrocks, B. and P. Charlesworth. 1982. A field study of the associationbetween the stinkhorn Ehg}}gg imggdiggs Pers. and the Britishfungal-breeding D;g5ggh}}g. Big}. 1. Linnggg Sgg. 17: 307-318.
Simberloff, D. S. 1974. Equilibrium theory of island biogeography. Agggg}Egg. Egg}. Sys;. S : 161-182. .
Singer, R. 1939. Contribution a l°etude des Russules 4. Quelques Russulesamericaines et asiatiques. Eg}}. Sgg. Mygg}. Eggggg 55: 226-283.
. 1942. Type studies on basidiomycetes. I. Mygg}gg}g 34: 64-93.
_________. 1943. Type studies on basidiomycetes. II. Mygg}gg}g 35:142-163.
m.-1949-22:5-832.
tl. 1957. New and interesting species of basidiomycetes V. Sy_dg_;g;|,_a11: 141-272.
.....- 1962- The Azar.:I.s.a1s.a in modem Lazssnszmx- 2¤d Ed- J- Cramer.Vaduz. 915p.
m- 1975- The Axazisales in mmm taasmnemx- 3rd Ed- J- Cramer.Vaduz. 912p.
Sokal, R. R. and N. L. Oden. 1978. Spatial autocorrelation in biology.1. Methodology. Ejg}. 1. L}gnggg Sgg. 10: 199-228.
Smith, J. D. 1980. Is biological control of Mgggsmigs ggggggg fairy ringspossible? B}gg; Qiggggg 64: 348-354.
Strausbaugh, P. D., and E. L. Core. 1978. Ihg f}g;g gf Egg; V};g}g}g. 2nded. Seneca Books, Inc. Grantsville.
Stephenson, S. L., and J. F. Clovis. 1983. Spruce forests of the AlleghenyMountains in central West Virginia. Qgg;gggg 48: 1-12.
Takhtajau. A- L- 1969- Elmmrius p.1a¤.Ls.. mrigin ausl siispsmal-Smithsonian Institute Press. Washington, D. C. 310 p. '
Bibliography 170
Taylor, J. D., G. J. Griffen, and K. H. Garren. 1981. Inoculum pattern,inocculum density, disease incidence relationships, and population
. flucuations of of §y}igg;gg}gg}gg g;gyg}g;}gg microsclerotia inpeanut field soil. Bhyygggyhg}ggy 71: 1297-1302.
Thomas, G. W., D. Rogers, and R. M. Jackson. 1983. Changes in themycorrhizal status of Sitka spruce following outplanting. B}ggyggg Sgj} 71: 319-323.
Thompson, W. and A. D. M. Rayner. 1982. Spatial structure of a population¤f nlaaamhxlla in a w¤¤d1d¤d Sitd- Neu Bh1t.212zi¤.t1 92: 103-114.
Treppe, J. M. 1962. Fungus associates of ectotrophic mycorrhizae. Egy.Bgyg. 28: 538-606.
Van Steenis, C. G. G. 1972. Ngyhgfgggg, key genus to plant geography. Pp.275-288- In= and a12l11.t.i2n- Ed- , D- H-Valentine. Academic Press, London.
Vuilleumier, F. 1970. Insular biogeogrephy in continental regions. I. Thenorth Andes of America. Agg;. Ngyg;g}igy. 104: 373-388.
Wagner, A. J. 1984. Why it happened? yggyhgggigg 37: 12-17.
Wasterlund, I., and T. Ingelog. 1981. Fruit body production in some youngSwedish forests with special reference to logging waste. BggggyEgg}. Mggg. 3: 269-294.
Watling, R., 1982. Taxonomic status and ecological identity in thebasididmycetes- In thai: 212191:2 andggg}ggy. Eds., J. C. Frankland, J. N. Hedger, and M. J. Swift.Cambridge University Press, Cambridge. pp. 1-32.
White, P. S., R. I. Miller, and G. S. Ramseur. 1984. The species-arearelationship of the Southern Appalachian high peaks: vascularplant richness and rare plant distributions. gggygggg 49: 47-62.
Whittaker, R. H. 1956. Vegetation of the Great Smoky Mountains. Egg}.Mgggg. 26: l-80.
_______________. 1960. Vegetation of the Siskiyou Mountains, Oregon andCalifornia. Egg}. Mgggg;. 30: 279-338.
Wilkins, W. H., and S. H. M. Patrick. 1940. The ecology of larger fungi.IV. The seasonal frequency of grassland fungi with specialreference to the influence of environmental factors. Agg. Apg}.Big}. 27: 17-34.
, and G. C. M. Harris. 1946. The ecology of larger fungi.V. An investigation in the influence of rainfall and temperature
Bibliography 171
on the seasonal production of fungi in beechwood and pinewood.A33. Aggl. ßigl. 33: 179-188.
Wojciechowska, H. 1960. Studies on the mycotrophy of the spruce (Ejggggxgglgg Lk.) in its northern range with particular regard to themycotrophy of plant communities in the Forest District Sadlowo,Subdistrict Lipowo near Biskupiec Reszilski. Eglig Egg. Egl. Sgg.A. 2: 123-166. (translated from Polish).
Zak, B., and D. H. Marx. 1964. Isolation of mycorrhizal fungi from rootsof individual slash pines. Eggggg Sgj. 10: 214-221.