Page 1
Basran, Harris, Sutcliffe and Scrutton 1
H-tunneling in the multiple H-transfers of the catalytic
cycle of morphinone reductase and in the reductive half-
reaction of the homologous pentaerythritol tetranitrate
reductase
Jaswir Basran1, Richard J. Harris1, Michael J. Sutcliffe1,2 and
Nigel S. Scrutton1
1Department of Biochemistry, University of Leicester, University Road, Leicester
LE1 7RH UK
2Department of Chemistry, University of Leicester, University Road, Leicester LE1
7RH UK.
_____________________________________________________________________
Corresponding author: Professor N. S. Scrutton. Telephone +44 116 223 1337; Fax,
+44 116 252 3369; email, [email protected]
Running title: Hydrogen tunneling in morphinone reductase and PETN reductase
This work was funded by grants from the BBSRC, the Wellcome Trust and the Lister
Institute of Preventive Medicine. N.S.S. is a Lister Institute Research Professor.
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on August 26, 2003 as Manuscript M305983200 by guest on A
ugust 3, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 2
Basran, Harris, Sutcliffe and Scrutton 2
ABBREVIATIONS
1PETN, pentaerythritol tetranitrate; MR, morphinone reductase; OYE, Old Yellow
Enzyme; KIE, kinetic isotope effect; SIE, solvent isotope effect.
2Appropriate fitting to the kinetic model was assessed using a number of criteria.
These involved: examination of the calculated spectra to ensure they made chemical
sense in terms of shape and sign (in this case negative absorption spectra were
predicted); the lack of systematic deviations of the residual plot; convergence to the
model obtained within a sensible number of iterative cycles (no more than 10);
confirmation that the number of significant singular values following singular value
decomposition are consistent with the fitted model.
3 An alternative explanation that we advanced in our previous work is that in
switching from a temperature independent to temperature dependent KIE thermal
energy is used to facilitate tunneling from excited vibrational modes where the
tunneling distance is expected to be less than for ground state tunneling reactions. In
principle, tunneling from an excited vibrational mode and/or the dominance of ‘gated’
dynamics over ‘Frank-Condon’ dynamics is consistent with the dissipative H-
tunneling model of Kuznetsov and Ulstrup (19), and studies of the temperature
dependence of the KIE cannot unequivocally disentangle the contribution of each to
the tunneling reaction.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 3
Basran, Harris, Sutcliffe and Scrutton 3
ABSTRACT
The mechanism of flavin reduction in morphinone reductase (MR) and pentaerythritol
tetranitrate (PETN) reductase, and flavin oxidation in MR, has been studied by
stopped-flow and steady-state kinetic methods. The temperature dependence of the
primary kinetic isotope effect for flavin reduction in MR and PETN reductase by
nicotinamide coenzyme indicates that quantum mechanical tunneling plays a major
role in hydride transfer. In PETN reductase, the KIE is essentially independent of
temperature in the experimentally accessible range, contrasting with strongly
temperature dependent reaction rates — consistent with a tunneling mechanism from
the vibrational ground state of the reactive C-H/D bond. In MR, both the reaction
rates and the KIE are dependent on temperature, and analysis using the Eyring
equation suggests that hydride transfer has a major tunneling component, which
unlike PETN reductase, is gated by thermally induced vibrations in the protein. The
oxidative half-reaction of MR is fully rate limiting in steady-state turnover with the
substrate 2-cyclohexenone and NADH at saturating concentrations. The KIE for
hydride transfer from reduced flavin to the α/β unsaturated bond of 2-cyclohexenone
is independent of temperature, contrasting with strongly temperature dependent
reaction rates — again consistent with ground state tunneling. A large solvent isotope
effect (SIE) accompanies the oxidative half-reaction, which is also independent of
temperature in the experimentally accessible range. Double isotope effects indicate
that hydride transfer from the flavin N5 atom to 2-cyclohexenone, and the protonation
of 2-cyclohexenone, are concerted and both the temperature independent KIE and SIE
suggest that this reaction also proceeds by ground state quantum tunneling. Our
results demonstrate the importance of quantum tunneling in the reduction of flavins
by nicotinamide coenzymes. This is the first observation of (i) three H-nuclei in an
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 4
Basran, Harris, Sutcliffe and Scrutton 4
enzymic reaction being transferred by tunneling and (ii) the utilisation of both passive
and active dynamics within the same native enzyme.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 5
Basran, Harris, Sutcliffe and Scrutton 5
INTRODUCTION
Enzymes are phenomenal catalysts that can achieve rate enhancements of up to ~21
orders of magnitude over the uncatalysed reaction rate (1). Our quest to understand
the physical basis of this catalytic power is challenging, and has involved sustained
and intensive research efforts by many workers in the physical and life sciences [for
recent reviews see (2-5)]. Recent years have witnessed new activity in this area, and
extended our theoretical understanding beyond the shortcomings of transition state
theory (6) to include roles for protein ‘motion’ (7,8), low barrier hydrogen bonds [e.g.
(9-11)], active site preorganisation [e.g. reviewed in (4,12)] and quantum mechanical
tunneling [for recent reviews see (13-15)]. New theoretical frameworks incorporating
quantum mechanical tunneling and protein motion are emerging to address the
catalytic potency of enzymes. These invoke motion in the protein and/or substrate to
drive the reaction (16-19). The reaction itself (i.e. the breaking and making of bonds)
is normally modelled using a hybrid quantum mechanical/molecular mechanical
(QM/MM) formulism, in which those atoms involved in the reaction are treated
quantum mechanically and the rest of the system treated classically using molecular
mechanics e.g. (20). An alternative approach, the “quantum Kramers” method
(15,21,22)—which treats the whole system using a simplified quantum mechanical
formulism—has been applied so far only to small organic systems. Additionally,
methodology has been developed for identifying computationally residues important
in creating reaction-promoting vibrations in enzymes (23). It has also been suggested
that dynamics of the enzyme should be divided into two types—passive
(reorganisation energy) and active (gating, or vibrational enhancement)—and that
tunneling is gated via active dynamics (i.e. a vibration modulating the hydrogen
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 6
Basran, Harris, Sutcliffe and Scrutton 6
transfer coordinate becomes thermally active resulting in increased tunneling
probability) (14,24).
Hydride transfer from a reducing nicotinamide coenzyme to a flavin cofactor
is a common reaction in biology, but the potential importance of H-tunneling in these
reactions has not been explored. H-tunneling has been characterised extensively in
NAD+-dependent alcohol dehydrogenases [e.g. (25-27)], which prompted us to study
more broadly a potential role for H-tunneling in flavoproteins that operate with
nicotinamide coenzymes. Herein, we have studied hydride and proton transfer in
morphinone reductase (MR)1 and pentaerythritol tetranitrate (PETN) reductase.
Crystallographic structures of these enzymes have established a close relationship to
Old Yellow Enzyme (OYE) (28-30), reflected also in the ability of the OYE family
members to reduce a number of 2-cyclic enones and to form complexes with steroids.
Like OYE, MR is a dimer, but the nature of the subunit interactions is different from
those seen in OYE. PETN reductase is a monomer and resembles in fold a single
subunit of OYE and MR, based on the archetypal eight-fold β/α barrel topology. The
active sites of all three enzymes are remarkably conserved despite differences in their
catalytic properties. NADPH-dependent PETN reductase was purified and cloned
from a strain of Enterobacter cloacae (strain PB2), which was isolated on the basis of
its ability to utilise nitrate ester explosives such as PETN and glycerol trinitrate
(GTN) as a sole nitrogen source (31,32). MR was identified in a strain of
Pseudomonas putida (strain M10) isolated from industrial waste liquors (33). MR
catalyses the NADH-dependent saturation of the carbon-carbon bond of both
morphinone and codeinone to produce hydromorphone (a powerful analgesic) and
hydrocodone (an antitussive), which are valuable semi-synthetic opiate drugs (34,35).
The half-reactions of MR and PETN reductase have been investigated using stopped-
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 7
Basran, Harris, Sutcliffe and Scrutton 7
flow methods (29,36,37), enabling elucidation of the kinetic mechanisms for each
half-reaction. We demonstrate in this paper that hydride transfer from nicotinamide
coenzyme to flavin in both enzymes occurs by quantum tunneling, but that the nature
of the tunneling reaction is different. Despite the similar active site architectures, this
likely reflects differences in the dynamics of the enzyme scaffold in MR and PETN
reductase. We show additionally that hydride transfer from reduced flavin to the
substrate 2-cyclohexenone in MR also occurs by tunneling, and that this reaction is
concerted with proton transfer from an unidentified active site acid to the substrate
unsaturated bond.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 8
Basran, Harris, Sutcliffe and Scrutton 8
EXPERIMENTAL PROCEDURES
Chemicals and enzymes—Complex bacteriological media were from Oxoid. Mimetic
Orange 2 and Mimetic Yellow 2 affinity chromatography resins were from Affinity
Chromatography Ltd. Q-Sepharose resin was from Pharmacia. PETN reductase was
prepared from E. coli JM109/pONR1 and purified as described (32), but also
incorporating a final chromatographic step using Q-Sepharose (28). MR was purified
from a recombinant strain of E. coli transformed with plasmid pMORB2, which
expresses the enzyme from the cloned mor B gene as described previously (33), but
also incorporating a final chromatographic step using Q-sepharose (29). NADPH and
NADH were from Sigma. 2-cyclohexenone was from Acros Organics. Deuterium
oxide (99.9 % deuterium) was from Goss Scientific Instruments Ltd. The following
extinction coefficients were used to calculate the concentration of substrates and
enzymes: NAD(P)H (ε340 = 6.22 x 103 M-1 cm-1); PETN reductase and MR (ε464 =
11.3 x 103 M-1 cm-1); 2-cyclohexenone (ε232 = 11.0 x 103 M-1 cm-1).
Deuterated compounds—A-side NAD2H was synthesised enzymatically as described
previously (38) and ethanol precipitated by the method of Pollock and Barber (39). A
further purification step was performed using a Q-Sepharose column. The column was
equilibrated with 10 mM ammonium hydrogen carbonate, pH 9 (buffer A) and A-side
NAD2H (approximately 20 mg) applied to the column. The column was washed with
buffer A and then developed with a linear gradient of 10 mM – 400 mM ammonium
hydrogen carbonate, pH 9; NAD2H eluted at approximately 300 mM ammonium
hydrogen carbonate. The A260 / A340 ratio of nucleotide-containing fractions was
determined and those fractions with a ratio of ≤ 2.3 were deemed pure (39), pooled
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 9
Basran, Harris, Sutcliffe and Scrutton 9
and freeze-dried. Purified NAD2H was stored at –80 °C and dissolved in 20 mM Tris
buffer pH 8.5 prior to use in kinetic experiments.
The synthesis and purification of NADP2H was as for NAD2H but with the
following modifications: NADP+-specific T. brockii alcohol dehydrogenase was used
in the enzymatic synthesis of NADP2H. NADP2H was eluted from the Q-Sepharose
column with 400 mM ammonium hydrogen carbonate, pH 9.
Preparation of anaerobic samples—Buffers were made anaerobic by bubbling argon
gas through solutions for ~2 hours. Solutions were then placed in an anaerobic glove
box (Belle Technology) overnight to remove any residual traces of oxygen. Protein
samples were made anaerobic by passing them through a small gel filtration (Bio-Rad
10 DG) column housed in the glove box, which had been pre-equilibrated with
anaerobic buffer. Coenzyme solutions were made by adding the solid to anaerobic
buffer. Solutions of 2-cyclohexenone were made by dilution of a 10 M stock into
anaerobic buffer.
Solvent isotope effect experiments—For experiments conducted in 2H2O, all buffer
components and substrates were dissolved in 2H2O and the pD of the solution was
calculated by the addition of 0.4 to the pH meter reading to correct for the isotope
effect on the electrode. Stock solutions of MR were exhaustively dialysed against 50
mM potassium phosphate buffer, pD 7.0. Preparation of anaerobic samples was as
outlined above.
Kinetic measurements— To prevent the oxidase activities of MR and PETN
reductase, all kinetic studies were performed under strict anaerobic conditions (<5
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 10
Basran, Harris, Sutcliffe and Scrutton 10
ppm O2) within a glove box environment (Belle Technology). Experiments were
performed in 50 mM potassium phosphate buffer, pH 7.0, at the stated temperatures.
Rapid reaction kinetic experiments were performed using an Applied Photophysics
SX.18MV-R stopped-flow spectrophotometer contained within the glove box.
Spectral changes accompanying flavin reduction and reoxidation in both enzymes
were monitored by rapid-scanning stopped-flow spectroscopy using a photodiode
array detector and X-SCAN software (Applied Photophysics). Spectral deconvolution
was performed by global analysis and numerical integration methods using PROKIN
software (Applied Photophysics). For single wavelength studies, data collected at 464
nm (flavin reduction and reoxidation) were analysed using nonlinear least squares
regression analysis on an Acorn Risc PC microcomputer using Spectrakinetics
software (Applied Photophysics). In the reductive half-reaction, experiments were
performed by mixing enzyme in the appropriate buffer with an equal volume of
reducing cofactor in the same buffer at the desired concentration. For studies of the
oxidative half-reaction, the sequential mixing mode of the stopped-flow apparatus
was used. Enzyme was rapidly mixed with a stoichiometric amount of reducing
cofactor to enable reduction of the enzyme-bound FMN and after a suitable pre-
determined ageing period, the reduced enzyme solution was rapidly mixed with 2-
cyclohexenone and reoxidation monitored at 464 nm. In both half-reactions, the
concentration of substrate was always at least 10-fold greater than that of enzyme,
thereby ensuring pseudo-first-order conditions. For each substrate concentration, at
least five replica measurements were collected and averaged.
The kinetics of the reductive half-reaction of MR and PETN reductase were
investigated at 464 nm, essentially as described previously (36,37). Observed rate
constants for flavin absorption changes occurring in the reductive half-reactions of
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 11
Basran, Harris, Sutcliffe and Scrutton 11
both enzymes were obtained from fits of the data to a standard double exponential
expression, where kobs1 (95 % total absorption change) and kobs2 (5 % total absorption
change) are observed rate constants for fast and slow phases, respectively. The faster
of the two phases is attributed to flavin reduction, as indicated by photodiode array
experiments of the reductive half-reaction. The slow phase represents a minor spectral
change, the origin of which is uncertain. Observed rates for the fast phase (kobs1) for
both half-reactions were fitted using the general hyperbolic expression (Eq. 1),
consistent with the kinetic schemes presented in Results and with previous reported
studies (see (36,37) for further details).
]S[]S[lim
obs +=
Kkk
Eq. 1
In equation 1, klim is the limiting rate for flavin reduction (reductive half-reaction) or
flavin oxidation (oxidative half-reaction). At low temperatures (5 °C), flavin
reduction was essentially independent of reducing nicotinamide coenzyme
concentration, consistent with previous studies (36,37). The pH dependence of the
rate of flavin reduction was measured in H2O in the range pH 5.5 – pH 9 using KMB
buffer (55 mM MES, 25 mM Tris, 25 mM ethanolamine).
In the oxidative half-reaction of MR, absorption changes at 464 nm reported
on flavin oxidation by 2-cyclohexenone as described previously (29). Transients were
biphasic, with the fast phase (kobs1; 90 % of the total absorption change) reporting on
flavin oxidation. The origin of the slow phase (kobs2; 10 % of the total absorption
change) is uncertain. Observed rates for the fast phase were hyperbolically dependent
on substrate concentration and data were fitted using equation 1.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 12
Basran, Harris, Sutcliffe and Scrutton 12
Steady-state kinetic analysis—Steady-state kinetic measurements were performed
using a Jasco V530 UV/VIS spectrophotometer with a 1 cm light path. Assays were
conducted in 50 mM potassium phosphate buffer, pH 7.0, at 25 °C in a total volume
of 1 mL. For determination of the kinetic parameters (apparent Km and kcat) for 2-
cyclohexenone, the reaction mixture contained 150 µM NADH, 250 nM MR and the
concentration of 2-cyclohexenone was varied. Initial velocity data as a function of 2-
cyclohexenone were analyzed by fitting to the standard Michaelis-Menten rate
equation. The pH dependence of the steady-state reaction was measured in H2O and
2H2O in the range pH 5.5 – pH 9 using KMB buffer (55 mM MES, 25 mM Tris, 25
ethanolamine). In both the pH and temperature dependence studies the concentration
of 2-cyclohexenone was kept constant (and saturating) at 50 mM. MR activity was
measured by following the decrease in absorption at 350 nm due to oxidation of
NADH and initial rates of reaction were calculated using an ε = 5650 M-1 cm-1.
Enzyme monitored turnover—Steady-state measurements were also performed using
the enzyme monitored turnover method as described by Gibson et al for reactions
catalysed by glucose oxidase (40). Reactions were performed in an Applied
Photophysics SX18-MVR reaction analyzer. Solution conditions are described in
Results. Data analysis was essentially as described elsewhere (40).
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 13
Basran, Harris, Sutcliffe and Scrutton 13
RESULTS
Temperature dependence of the reductive half-reaction of MR and kinetic isotope
effects—The mechanism of flavin reduction in MR was determined previously and
shown to involve the rapid formation of an E-NADHCT charge-transfer intermediate
prior to flavin reduction [(36); Scheme 1]. Formation and decay of the charge-transfer
species can be monitored at 552 nm, and flavin reduction at 462 nm. The decay of the
charge-transfer species is kinetically indistinguishable from flavin reduction,
indicating the two processes are linked (36). Spectral changes observed in the
reductive half-reactions of MR with a 10-fold excess of NADH were previously
shown to fit to a two-step kinetic model: A → B → C, consistent with these
assignments, where A is the oxidised enzyme, B is an enzyme-coenzyme charge-
transfer intermediate and C is enzyme containing the reduced (dihydroflavin) form of
the flavin cofactor (36), although fitting to more complex reversible kinetic models
was not explored. In performing more extensive kinetic studies to search for tunneling
regimes in the hydride transfer reaction from NADH to FMN it is essential to know if
the measured rate constants in stopped-flow experiments report on (i) the approach to
an equilibrium position for a reversible chemical step or (ii) an essentially irreversible
reaction. With this in mind, we have conducted additional stopped-flow
measurements using photodiode array spectroscopy and analysed globally the spectral
changes by fitting to reversible kinetic models.
Data were collected for reactions of MR with protiated and deuterated
coenzyme at both 5 °C and 36 °C (i.e. the extremes of the temperature range used in
analysis of the temperature dependence of the reductive half-reaction described
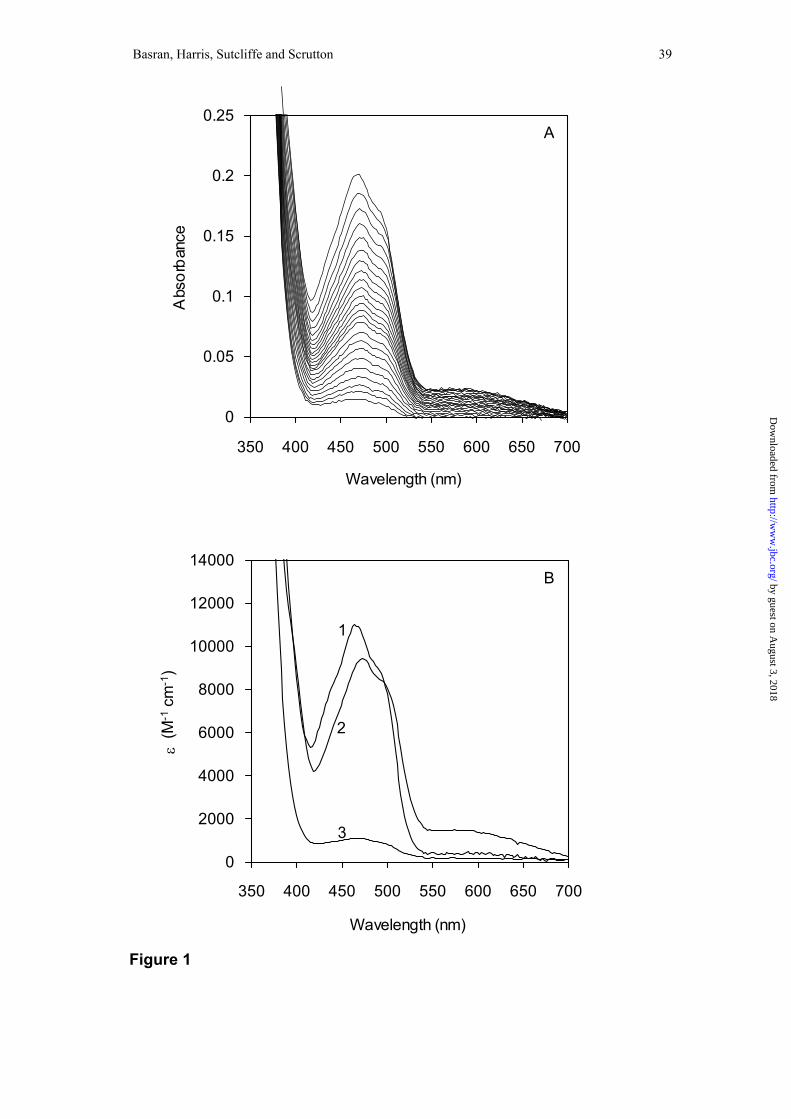
below). A typical dataset is shown in Figure 1 for the reaction of 20 µM MR with 200
µM NADH at 5 °C. Attempts to fit to a fully reversible model, A ↔ B ↔ C were
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 14
Basran, Harris, Sutcliffe and Scrutton 14
unsuccessful based on a number of criteria2. However, the data were readily fitted to
the A → B → C kinetic model (rate constant for A → B is 154 s-1; rate constant for B
→ C is 14.8 s-1) as described previously by Craig et al (36) and the determined rate
constants agree closely with those determined from single wavelength analysis of
kinetic transients at 462 nm (flavin reduction) and 552 nm (charge transfer formation
and decay). For the fully reversible kinetic model A ↔ B ↔ C the criteria used to
assess the fit2 could only be satisfied when the rate constant for the conversion of C
→ B was initially estimated as a very small value (i.e. <0.1 s-1); the final rate
constants are A → B (149 s-1), B → C (15.2 s-1), B → A (4.5 s-1) and C → B (0.1 s-1)
(Figure 1), again consistent with studies performed at single wavelength (36). The
predicted spectra for the enzyme forms (Figure 1B) obtained by fitting to a reversible
scheme in which the rate of conversion of C → B is very small (0.1 s-1) are essentially
identical to those obtained when fitting to the A → B → C model (36). This indicates
that the rate of reverse hydride transfer from FMNH2 to NAD+ is negligible.
Qualitatively, similar results were obtained for reactions performed at 36 °C and in
studies with NAD2H. These observations are consistent with the known reduction
potential of NADH (-320 mV) and MR (-237 mV; (41)), and they indicate that
reductive transients measured under single wavelength conditions at 464 nm report on
essentially irreversible reduction of the FMN by NADH. More recent studies with the
C191A mutant of MR have provided evidence for an additional E-NADH
intermediate that accumulates in the dead time (~1 ms) of the stopped-flow
instrument, prior to the formation of the E-NADHCT charge-transfer complex, giving
rise to Scheme 2 for the reductive half-reaction of MR (29). This modified scheme for
the reductive half-reaction of MR is consistent with work on Old Yellow Enzyme (42)
and estrogen binding protein (43), and probably holds also for wild-type MR
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 15
Basran, Harris, Sutcliffe and Scrutton 15
[although unequivocal evidence is lacking; (29)]. Notwithstanding the increased
complexity of this scheme, measurements of flavin reduction at 464 nm would still
report on the essentially irreversible rate of reduction of the FMN cofactor by NADH.
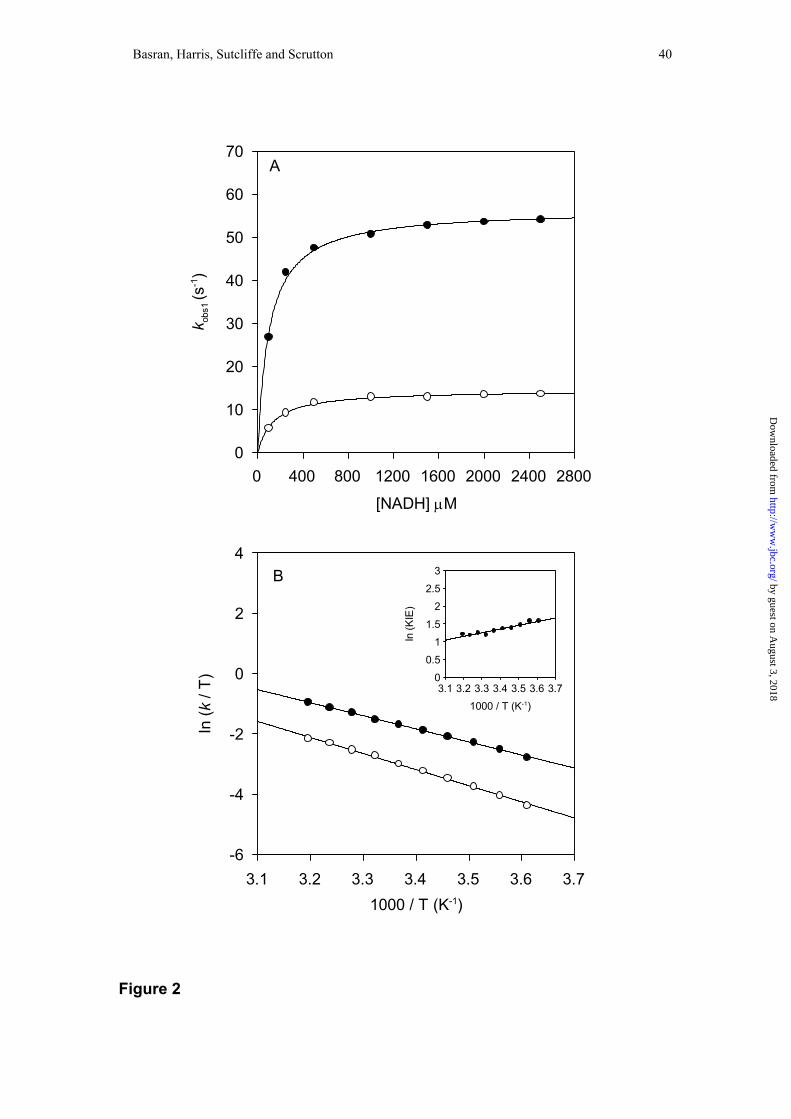
For wild-type MR at 5 °C, the rate of flavin reduction is independent of
NADH concentration in the range 100 to 1100 µM coenzyme (36), but at higher
temperatures a hyperbolic dependence of the rate of flavin reduction on NADH
concentration is observed (Figure 2). The value of K is dependent on temperature (45
± 7 at 15 °C; 101 ± 7 at 25 °C and 320 ± 24 at 35 °C). In fitting data, it was assumed
that flavin reduction is essentially irreversible (i.e. the ordinate intercept in Figure 2
approximates to zero). This is consistent with global fitting of photodiode array for
the reductive half-reaction (see above). The limiting rate constant, klim, for flavin
reduction in MR is independent of solution pH between the range pH 5.5 – 9.0. A
primary KIE of 3.9 ± 0.1 (25 °C) is observed for the limiting rate of flavin reduction
calculated by fitting to Eq. 1 (Figure 2A) and there is no significant solvent isotope
effect on flavin reduction (SIE = 1.05 ± 0.02). These results suggest that the solvent
isotope effect observed in steady-state turnover of MR (see below) must arise from
effects on step(s) that take place after flavin reduction (i.e. in the oxidative half-
reaction).
Eyring plots for the limiting rate of flavin reduction, klim, in protiated solvent
were constructed by performing stopped-flow studies at each temperature with 5 mM
coenzyme (Figure 2B), thus ensuring that the NADH concentration was always at
least 10-fold greater than the value of K. Data were fitted to the Eyring equation (Eq.
3), and the parameters ∆H ‡ and A’H:A’D obtained.
( ) RTHRShkTk ///ln/ln ‡‡Blim ∆−∆+= Eq. 3
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 16
Basran, Harris, Sutcliffe and Scrutton 16
The definitions of the terms A’H and A’D are given in our previous work (44). The
data indicate that the KIE is dependent on temperature, with ∆∆H‡ (∆H‡D-∆H‡H) =
(43.5 – 35.3 kJ mol-1) = 8.2 ± 0.27 kJ mol-1. The value of A’H:A’D < 1 (= 0.126 ±
0.005), consistent with a tunneling mechanism for transfer of the hydride ion from
NADH to FMN that is gated by vibrations coupled to the reaction coordinate (see
Discussion) (45).
Temperature dependence of the reductive half-reaction of PETN reductase and kinetic
isotope effects—The mechanism of action of PETN reductase is similar to that of MR,
and detailed stopped-flow studies with NADPH have revealed the existence of an E-
NADPHCT charge-transfer species prior to flavin reduction (37). The kinetic model
shown in Scheme 1 (but with NADPH as coenzyme) is consistent with the published
kinetic data. Given the potential importance of tunneling in the reductive half-reaction
of MR, we also investigated the temperature dependence of this half-reaction in
PETN reductase. Like with MR, charge-transfer formation and decay can be observed
at long wavelength (560 nm), and flavin reduction is monitored conveniently at 464
nm. As with MR, fitting of spectral datasets for the reductive half-reaction (20 µM
PETN reductase mixed with 200 µM NADPH at 5 °C) to a fully reversible kinetic
model A ↔ B ↔ C indicated that hydride transfer is essentially irreversible (i.e. C →
B, 0.1 s-1) to satisfy the fitting criteria2 (data not shown). All other microscopic rate
constants determined from this fit (A → B, 109 s-1; B → C 11 s-1; B → A, 2 s-1) are
consistent with data obtained from single wavelength studies (37). Our analyses again
demonstrate that hydride transfer is essentially irreversible, which is consistent with
the known redox potential of PETN reductase (37). These data also confirm that
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 17
Basran, Harris, Sutcliffe and Scrutton 17
single wavelength studies of the reductive half-reaction performed at 464 nm report
on the essentially irreversible rate of hydride transfer from NADPH to FMN.
As with MR, decay of the charge-transfer species monitored at 560 nm is
linked kinetically to flavin reduction at 464 nm (37). Our previous studies also
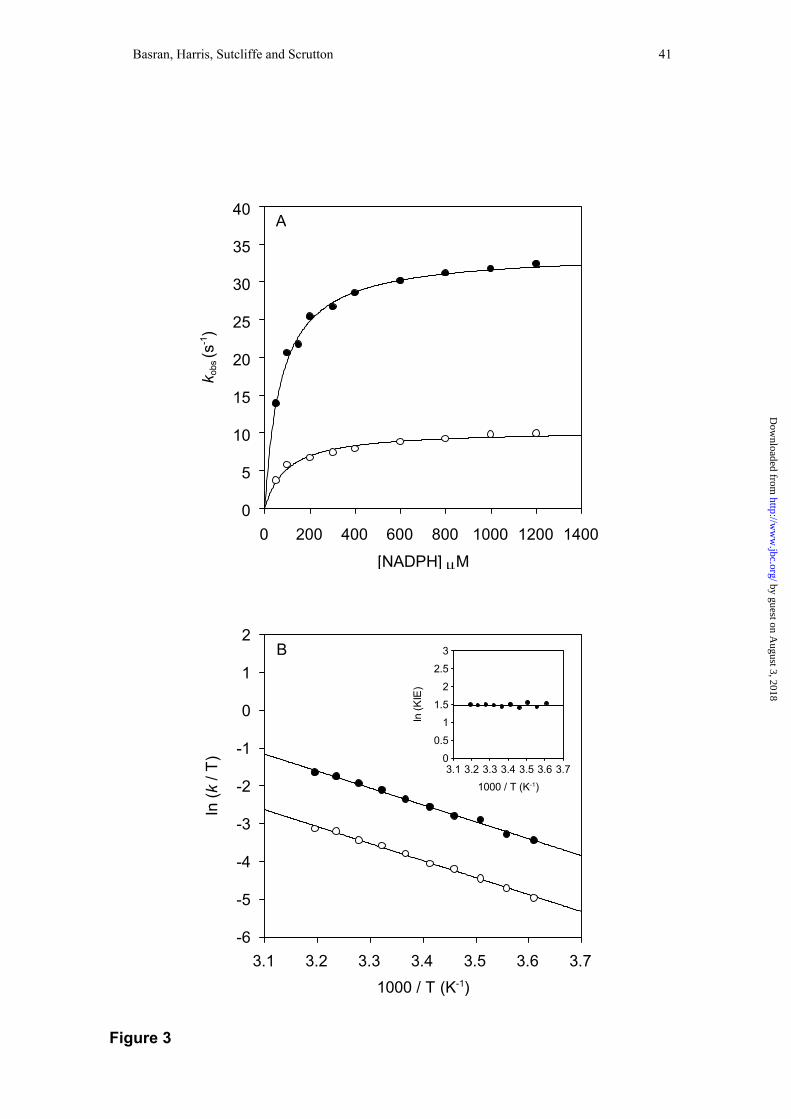
indicated that the flavin reduction rate is independent of NADPH concentration at 5
°C (37), but as with MR we have shown herein that at higher temperatures a
hyperbolic dependence is observed (Figure 3). The value of K determined by fitting to
Eq. 1 is dependent on temperature (27 ± 3 at 15 °C; 73 ± 4 at 25 °C and 186 ± 14 at
35 °C). As with MR, the limiting rate constant for flavin reduction is independent of
solution pH between the pH values 5.5 and 9. An Eyring plot of the limiting rate of
flavin reduction indicates that the KIE (~4.1) for hydride transfer from coenzyme to
FMN is essentially independent of temperature (A’H:A’D = 4.1 ± 0.3; ∆∆H‡ = 0.20 ±
0.01 kJ mol-1); fitting to the Eyring equation yielded values for ∆H‡D and ∆H‡H of
36.6 ± 0.9 and 36.4 ± 0.9 kJ mol-1, respectively. The data is strikingly similar to that
obtained for C-H bond breakage catalysed by a number of amine oxidising enzymes
in which reaction rates are strongly dependent on temperature, but the KIE is
independent of temperature over the experimentally accessible range (44,46-48). H-
transfer in these enzymes occurs by a quantum tunneling mechanism, probably from
the vibrational ground state of the reactive C-H/C-D bond (see (13) for a recent
review), and this interpretation is consistent with hybrid quantum
mechanical/molecular mechanical simulations of these reactions (49).
Stopped-flow methods for the kinetics of hydride transfer from FMNH2 to 2-
cyclohexenone in the oxidative half-reaction of MR—The kinetics of the oxidative
half-reaction of MR determined at 25 °C have been reported elsewhere (29). Also,
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 18
Basran, Harris, Sutcliffe and Scrutton 18
using the fitting criteria discussed for the reductive half-reaction, fitting of spectral
data associated with the oxidative half-reaction to a fully reversible model indicates
that the reverse rate of hydride transfer is at least ~150-fold slower than the forward
rate. This is consistent with the lack of an ordinate intercept when fitting data in plots
of observed rate of flavin oxidation versus 2-cyclohexenone concentration to equation
1 (29). Thus, the transfer of a hydride ion from FMN to 2-cyclohexenone can be
analysed essentially as an irreversible reaction.
Herein, our studies were extended over the accessible temperature range (4 to
40 °C) to enable comparison with steady-state turnover data (see below), and to assess
the effect of temperature on the binding of 2-cyclohexenone. In these experiments,
enzyme (5 µM) was reduced with a stoichiometric amount of NADH, and following
an appropriate time to effect full reduction, the reduced enzyme was mixed with 2
cyclohexenone in a double mixing sequential stopped-flow experiment. The
experimental approach is different to that reported previously, in which the kinetics of
the oxidative half-reaction were analysed by single mixing of MR (which had been
titrated with sodium dithionite) with 2-cyclohexenone (29). Notwithstanding, at a
range of temperatures studied in the double mixing method described in this
manuscript, the dependence of the observed flavin reoxidation rate on 2-
cyclohexenone concentration is hyperbolic (Figure 4), consistent with our previous
findings at 25 °C (29). Limiting rate constants, klim, and reduced enzyme-substrate
dissociation constants, K, at 4 and 40 °C are given in the legend to Figure 4. The lack
of major change in K over the accessible temperature range established that a study of
the temperature dependence of the limiting rate of flavin reduction is possible at a 2-
cyclohexenone concentration of 50 mM throughout the temperature range.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 19
Basran, Harris, Sutcliffe and Scrutton 19
In the oxidative half-reaction of MR a hydride ion is transferred from the N5
atom of FMN to 2-cyclohexenone (Figure 5). Saturation of the double bond also
requires a proton, but the identity of the proton donor in MR is as yet uncertain (29).
The exchange of protium or deuterium on the flavin N5 atom with protons from bulk
solvent is a potential problem one encounters in studies of the oxidative half-reaction
that employ KIEs as probes of H-transfer. A series of double mixing stopped-flow
studies were performed to investigate how rapidly deuterium at the N5 position of
enzyme-bound FMN exchanges with protons in bulk solvent. MR (5 µM) was mixed
with 5 µM NADH (NAD2H) and the time required to fully reduce the flavin was
determined from absorption changes at 464 nm in single mix experiments at 4, 24 and
40 °C (Figure 6). Using the sequential mixing mode of the stopped-flow apparatus,
and following a suitable aging time to effect complete reduction of the enzyme, the
kinetics of the OHR were then followed by rapidly mixing the reduced enzyme with
50 mM 2-cyclohexenone (Figure 6). In a series of ‘wash-out’ experiments, the
reduced enzyme was also allowed to age for increasing lengths of time to allow
exchange of deuterium on the flavin N5 atom, before enzyme was mixed with 2-
cyclohexenone. These experiments indicated that the KIE value remained constant up
to an aging time of ~100 s at 24 °C. Thereafter, the KIE diminished as the aging time
was extended beyond 100 s, reflecting exchange of deuterium with protons from bulk
solvent. At 24 °C and with an aging time of 10 s, the KIE measured for the oxidative
half-reaction was 3.8 ± 0.4. Although the exchange kinetics were faster at higher
temperatures, an aging time of 10 s did not lead to significant exchange with protons
from bulk solvent. Using this approach, a KIE of 3.7 ± 0.3 was measured at 4 °C
(Figure 6). The sequential mixing method provides a guide as to the value of the KIE
for hydride transfer in the oxidative half-reaction at different temperatures, and serves
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 20
Basran, Harris, Sutcliffe and Scrutton 20
to illustrate that deuterium is not rapidly lost from the N5 atom of the flavin following
reduction by coenzyme. However, the method is not sufficiently robust to provide
accurate and highly reproducible rates for hydride/deuteride transfer as a function of
temperature (and thus as a probe of tunneling). For this reason, we chose to study the
oxidative half-reaction under steady-state conditions.
Steady-state analysis of MR, double isotope effects and temperature dependence of
kinetic isotope effects— Steady-state assays of MR have established that the limiting
rate of hydride transfer in the oxidative half-reaction is comparable to kcat, suggesting
this is the overall rate-limiting step in steady-state turnover (29). The enzyme
monitored turnover method (40) using diode array and single wavelength detection in
the stopped-flow instrument was used to confirm further that the oxidative half-
reaction is rate limiting. In enzyme monitored turnover experiments, the reduction
level of the flavin is monitored prior to, during and after the steady-state phase by
absorption measurements at 464 nm. In these experiments there is a rapid and almost
complete bleaching of the flavin absorption at 464 nm on mixing enzyme with NADH
and 2-cyclohexenone (Figure 7A). This is followed by a steady-state phase and then
finally an increase in absorbance as the oxidised enzyme is regenerated owing to
depletion of the reducing cofactor. The spectral forms of MR obtained at different
time points during the course of this reaction are shown in the inset of Figure 7A.
These confirm that the reduced form of the enzyme is the predominant species under
steady-state turnover conditions. At the start of data acquisition (point 1 on the trace;
3.8 ms after mixing) the oxidised E-NADHCT charge transfer species has already
formed owing to the relatively high concentration of NADH used (formation of the E-
NADHCT charge transfer species is second order with respect to NADH
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 21
Basran, Harris, Sutcliffe and Scrutton 21
concentration) and the relatively long time delay from mixing to data acquisition in
this experiment. Following rapid reduction of the flavin by NADH, a steady-state
phase is established during which the predominant species is the 2 electron form of
MR (point 2 on the trace). As NADH is depleted, the oxidised enzyme is formed once
again (point 3 on the trace).
The data were analysed by the method of Gibson et al (40) and a series of
parallel lines were obtained when the reciprocal of the turnover number was plotted
versus reciprocal of the NADH concentration (Figure 7B), consistent with a ping-
pong reaction that is the result of shared binding sites for NADH and 2-
cyclohexenone in the OYE family of enzymes (28-30). The inset to Figure 7B shows
a secondary plot of the ordinate intercept versus 2-cyclohexenone concentration. The
true turnover number (kcat = 2.5 ± 0.1 s-1) for the MR catalysed reaction is obtained
from the ordinate intercept of this secondary plot and the true Km for 2-cyclohexenone
(3.0 ± 0.2 mM) is derived from this plot by dividing the value of the gradient by the
ordinate intercept. The true Km for NADH (6.2 ± 1.4 µM) was calculated by dividing
the slope of any line in Figure 7B by the intercept of that line. The kinetic parameters
measured using the enzyme monitored turnover method are similar to apparent values
published previously for MR obtained by conventional enzyme assay (29) using the
initial rate method. In addition, the limiting rate for flavin oxidation by 2-
cyclohexenone (2.9 s-1) measured in the stopped-flow apparatus is similar to the
turnover number (2.5 s-1) indicating that flavin oxidation is rate-limiting in steady-
state turnover.
Having established that flavin oxidation is rate limiting in steady-state
turnover, we performed a detailed study of the oxidative half-reaction as a function of
temperature and isotopic substitution to (i) demonstrate that hydride transfer in the
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 22
Basran, Harris, Sutcliffe and Scrutton 22
oxidative half-reaction is fully rate limiting, (ii) probe for tunneling in this reaction
and (iii) obtain evidence for a concerted hydride and proton transfer in the oxidative
half-reaction. The oxidative half-reaction of PETN reductase is not rate limiting in
steady-state turnover (37), and thus our studies were restricted to MR.
Steady-state assays were initially performed at 25 °C using NADH or NAD2H
as reducing coenzyme, and in protiated and deuterated solvent. Plots of initial velocity
versus 2-cyclohexenone concentration (range 0 to 30 mM) at a fixed coenzyme
concentration (150 µM) were hyperbolic, and fitting to the Michaelis-Menten
equation yielded apparent Km values for 2 cyclohexenone of 5.5 ± 0.1 mM (for
NADH in protiated solvent), 4.3 ± 0.5 mM (for NAD2H in protiated solvent), 5.4 ±
0.4 mM (NADH in deuterated solvent) and 3.5 ± 0.4 mM (NAD2H in deuterated
solvent). Turnover numbers were then determined using a fixed concentration of 2-
cyclohexenone (50 mM) and reducing coenzyme concentration (150 µM) to obtain
values for the primary KIE for hydride transfer from the flavin N5 atom to 2-
cyclohexenone, the SIE for protonation of 2-cyclohexenone and the double isotope
effect for the oxidative half-reaction (Table 1). The primary KIE observed for hydride
transfer from the N5 atom of the reduced flavin to 2-cyclohexenone, and the values of
the turnover numbers obtained compared with flavin reoxidation rates measured in the
stopped-flow, is consistent with this reaction being fully rate limiting in multiple
turnover assays. Moreover, a large SIE is seen on the turnover number indicating that
a protonation event accompanies reduction of 2-cyclohexenone, consistent with the
chemical scheme shown in Figure 5. The double isotope effect method is useful in
demonstrating that two H-transfer reactions are concerted, i.e. if the chemical step is
fully rate limiting (as is the case for MR) and the two isotopes are on the same step,
the second isotope effect should remain unchanged. A more detailed treatment is
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 23
Basran, Harris, Sutcliffe and Scrutton 23
given in ref (50). This prediction holds (within error) for the oxidative half-reaction of
MR, consistent with a concerted reaction involving the simultaneous transfer of a
hydride and proton (Table 1).
The temperature dependence of the KIE values in protiated solvent indicate
that the KIE is essentially independent of temperature (∆∆H‡ = -0.52 ± 0.05 kJ mol-1)
over the accessible temperature range (Figure 8), and the A’H:A’D ratio (3.7 ± 2.1) is
greater than unity. The data are consistent with a concerted hydride and proton
transfer reaction that occurs by quantum tunneling from the vibrational ground states
of the reactive bonds, in a reaction that is not gated by vibrations coupled to the
reaction coordinate (see Discussion). Comparable studies with deuterated solvent and
NAD2H were not performed owing to the very slow turnover rates with NAD2H at
low temperature, but temperature dependent studies with NADH in deuterated solvent
indicate that the solvent isotope effect is also independent of temperature (∆∆H‡ = -
0.90 ± 0.05 kJ mol-1), with the A’H:A’D ratio (3.1 ± 2.0) greater than unity, again
consistent with a tunneling mechanism. Steady-state pH dependence studies
demonstrated that the oxidative half-reaction in both H2O and 2H2O is independent of
solution pH (in the pH range pH 7 – pH 9) and that the solvent isotope effect remains
constant (~ 2) in this pH range. Also, the turnover number of MR was independent
of solution viscosity in reactions performed over a range of glycerol concentrations (1
to 35% w/v), indicating that the increased viscosity of the deuterated solvent used in
these studies is not responsible for the measured SIE.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 24
Basran, Harris, Sutcliffe and Scrutton 24
DISCUSSION
Our earlier studies with methylamine dehydrogenase established that enzymic H-
transfer can proceed solely by quantum tunneling (44), without the need to (partially)
ascend the energy barrier separating reactants from products. The reaction of
methylamine dehydrogenase with methylamine was originally modelled using the
vibrationally enhanced ground state tunneling (VEGST) model of Bruno and Bialek
(16) at a time when the environmentally coupled hydrogen tunneling model (19),
which explicitly recognises reorganisation energy (so-called ‘passive dynamics’) and
active dynamics (gating motion) (24), was not available. We have continued to
provide (along with others) experimental evidence for enzyme catalysis based on
dissipative tunneling models in which H-transfer occurs entirely by quantum
mechanical tunneling [e.g. (24,46-48,51)], and these models are distinct from the
earlier tunnel-correction models (52) used to interpret anomalous kinetic isotope
effect data with other enzyme systems (see (53) for a review).
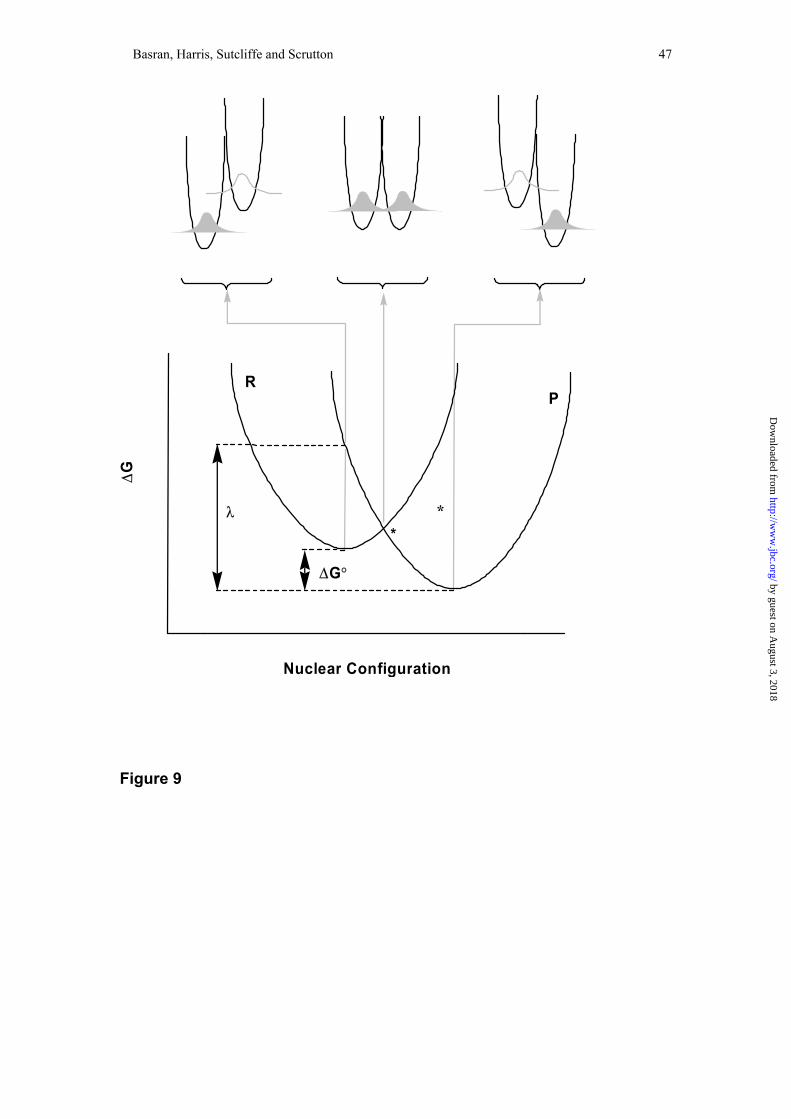
A major step forward has been the realisation that tunneling is driven by
thermally induced vibrations in the protein scaffold (a thermally fluctuating energy
surface), as described by the theoretical model of Kuznestov and Ulstrup (19), and
illustrated in Figure 9. This model showed that (24):
( ) ( )TermCFRT
Gconstktunnel ..4
exp.)(2
×
+°∆−
×=λ
λ Eq. 3
Here, ktunnel is the tunneling rate constant; const. an isotope-independent term; the
term in square parentheses is an environmental energy term relating the driving force
of the reaction, °∆G , to the reorganisational energy, λ (R is the gas constant and T the
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 25
Basran, Harris, Sutcliffe and Scrutton 25
temperature in K); the F.C. Term is the Frank-Condon nuclear overlap along the
hydrogen coordinate and arises from the overlap between the initial and the final
states of the hydrogen’s wavefunction. In the simplest limit, in which only the lowest
vibrational level is occupied, the F.C. Term will be temperature independent;
otherwise, it will be temperature dependent. Temperature dependent ‘gating’
dynamics can modulate the tunneling overlap and the KIE is dependent on the
energetic cost of gating—the KIE (equation 4) can be derived from equation 3 (24):
( ) ( )
( ) ( )∫∫
−−
−−==
0
1
0
1
d/exp2/exp
d/exp2/expKIE
BX2
DDD
BX2
HHH
D
Hr
r
r
r
XTkErm
XTkErm
F.C.TermF.C.Term
h
h
ω
ω Eq. 4
Here, kB is Boltzmann’s constant, ro is the equilibrium separation, r1 is the final
separation, ωH and ωD the frequencies of the reacting bond, and mH and mD the masses
of the transferred particle. The energetic cost of gating (Ex) is given by (14):
2X
2XX
2XX 2
121 rmXE ωω == h Eq. 5
Here the gating coordinate (X) is related to the gating oscillation (ωX), the distance the
gating ‘unit’ moves (rX) and its mass (mX) as follows:
xxX hωmrX = Eq. 6
The model predicts that if the gating term dominates (i.e. ħωx<kBT), the observed KIE
will be temperature dependent, since this leads to different transfer distances for the
heavy and light isotope. However, if the Frank-Condon term dominates the KIE will
either be temperature independent or, if excited vibrational levels are occupied, there
will be some temperature dependence. In the regime where ħωx ~ kBT gating plays
some role in modulating the tunneling probability, temperature dependent KIEs are
observed and the AH/AD values decrease (compared with the regime where the Frank-
Condon dynamics dominate), and may approach unity (14). Our previous studies with
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 26
Basran, Harris, Sutcliffe and Scrutton 26
wild-type trimethylamine dehydrogenase and the Y169F mutant (48), and reactions of
methylamine dehydrogenase with methylamine and ethanolamine (44,47) have
demonstrated both temperature independent and temperature dependent KIEs,
suggesting that tunneling can proceed in reactions dominated either by ‘gated’ or
‘Frank-Condon’ dynamics in both enzymes3. Similar explanations have been
advanced to explain temperature dependent KIEs in wild-type and mutant forms of
soybean lipoxygenase-1 (24). The experimental evidence with these enzymes is
therefore consistent with H-transfer by environmentally coupled hydrogen tunneling
and can be modelled satisfactorily using the theoretical framework of Kuznetsov and
Ulstrup (19). Interestingly, the temperature dependent KIE for MR versus the
temperature independent KIE for PETN reductase might be explained in these very
terms. One possibility is that PETN reductase is relatively more rigid compared with
MR, suggesting gating is less dominant in PETN reductase, which in turn predicts that
the KIE would be more temperature dependent in MR than in PETN reductase. Also,
the active site of PETN reductase might be more optimally configured for hydride
transfer than that of MR, thus requiring little (or no) vibrational assistance through
gated motion.
We have compared the high resolution crystal structures of MR (29) and
PETN reductase (28) in an attempt to provide insight into why gating is potentially
more important in MR (a more detailed analysis in the future will involve QM/MM,
variational transition state theory and molecular dynamics studies). A key factor
could be double stranded anti-parallel β-sheet D, against which the NAD(P)H
coenzyme is thought to bind (29), which harbours arginine residues important in the
recognition of the 2’phosphate of NADPH (PETN reductase) and a glutamate residue
required to form a H-bond with the 2’OH group of NADH (MR). These types of
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 27
Basran, Harris, Sutcliffe and Scrutton 27
sidechain-coenzyme interactions are consistent with work on other nicotinamide-
dependent enzymes, e.g. (54-56). The position of this sheet diverges at Leu-133
(PETN reductase)/Val-138 (MR) and converges again at Ile-141 (PETN
reductase)/Gly-146 (MR). Another difference in this region is the insertion of a
glycine residue (Gly-133) in MR immediately before the start of β-sheet D. Taken
together, these differences suggest that MR might be more mobile at physiological
temperatures in this region than PETN reductase—thus, active dynamics (gating) is
more likely an important feature in MR than in PETN reductase. This is consistent
with the temperature factors for MR (all Cα temperature factors > 40; pdb accession
code 1GWJ) and PETN reductase (all Cα temperature factors <20; pdb accession code
1GVQ) in this region. Of course, caution must be exercised in interpreting the
experimental results in the light of crystal structures, which do not contain the
nicotinamide coenzyme. Structural studies with coenzyme bound are now a priority
for future work, and these will form a platform for more detailed computational
analysis using hybrid QM/MM and related methods.
Our kinetic studies have additionally indicated that hydride transfer from
reduced flavin to the substrate 2-cyclohexenone in MR also occurs by tunneling. The
KIE for this reaction is temperature independent, consistent with the lack of
vibrational assistance. This reaction is concerted with proton tunneling from an
unidentified active site acid to the substrate unsaturated bond. Thus, MR invokes
both active (i.e. vibrationally gated) hydride transfer (reductive half-reaction) and
passive (i.e. no vibrational assistance) hydride and proton tunneling (oxidative half-
reaction). This illustrates, for the first time, how both active and passive tunneling can
be invoked within the same enzyme.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 28
Basran, Harris, Sutcliffe and Scrutton 28
Concluding Remarks—Hydride transfer from a reducing nicotinamide coenzyme to a
flavin cofactor is a common reaction in biology—examples of such systems include:
adrenodoxin reductase, cytochrome P450 reductase, ferredoxin reductase, nitric oxide
synthase and sulphite reductase. Despite their generality, the potential importance of
H-tunneling in these reactions has not been explored hitherto. Our studies have shown
that two such systems, PETN reductase and MR, invoke H-tunneling. Despite the
similar overall architecture of the active sites, hydride transfer in the reductive half-
reaction of MR, but not in that of PETN reductase, is gated by protein dynamics.
Also, hydride transfer from reduced flavin to the substrate 2-cyclohexenone in MR
occurs by tunneling with no significant gating component. Furthermore, this reaction
is concerted with proton tunneling from an unidentified active site acid to the
substrate unsaturated bond. This is the first observation of (i) three H-nuclei in an
enzymic reaction all being transferred by quantum mechanical tunneling and (ii) both
passive and active dynamics associated with H-tunneling in the same native enzyme
[although both tunneling regimes have been identified in wild-type versus mutant
enzymes, e.g. (14)]. More generally, our work reinforces the key role of quantum
tunnelling reactions in enzymic H-transfer.
ACKNOWLEDGMENTS
The work described in this article was funded by the UK Biotechnology and
Biological Sciences Research Council, the UK Engineering and Physical Sciences
Research Council, the Wellcome Trust and the Lister Institute of Preventive
Medicine. NSS is a Lister Institute Research Professor.
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 29
Basran, Harris, Sutcliffe and Scrutton 29
REFERENCES
1. Lad, C., Williams, N. H., and Wolfenden, R. (2003) Proc. Natl. Acad. Sci. U S
A 100, 5607-5610.
2. Neet, K. E. (1998) J. Biol. Chem. 273, 25527-25528
3. Cannon, W. R., and Benkovic, S. J. (1998) J. Biol. Chem. 273, 26257-26260
4. Warshel, A. (1998) J. Biol. Chem. 273, 27035-27038
5. Cleland, W. W., Frey, P. A., and Gerlt, J. A. (1998) J. Biol. Chem. 273,
25529-25532
6. Kraut, J. (1988) Science 242, 533-540
7. Cameron, C. E., and Benkovic, S. J. (1997) Biochemistry 36, 15792-15800
8. Rajagopalan, P. T., Lutz, S., and Benkovic, S. J. (2002) Biochemistry 41,
12618-12628
9. Frey, P. A., Whitt, S. A., and Tobin, J. B. (1994) Science 264, 1927-1930
10. Gerlt, J. A., and Gassman, P. G. (1993) Biochemistry 32, 11943-11952
11. Cleland, W. W., and Kreevoy, M. M. (1994) Science 264, 1887-1890
12. Cannon, W. R., Singleton, S. F., and Benkovic, S. J. (1996) Nat. Struct. Biol.
3, 821-833
13. Sutcliffe, M. J., and Scrutton, N. S. (2002) Eur. J. Biochem. 269, 3096-3102
14. Knapp, M. J., and Klinman, J. P. (2002) Eur. J. Biochem. 269, 3113-3121
15. Antoniou, D., Caratzoulas, S., Kalyanaraman, C., Mincer, J. S., and Schwartz,
S. D. (2002) Eur. J. Biochem. 269, 3103-3112
16. Bruno, W. J., and Bialek, W. (1992) Biophys. J. 63, 689-699
17. Borgis, D., and Hynes, J. T. (1996) J. Phys. Chem. 100, 1118-1128
18. Antoniou, D., and Schwartz, S. D. (1997) Proc. Natl. Acad. Sci. U. S. A. 94,
12360-12365
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 30
Basran, Harris, Sutcliffe and Scrutton 30
19. Kuznetsov, A. M., and Ulstrup, J. (1999) Can. J. Chem. 77, 1085-1096
20. Gao, J., and Thompson, M. (eds) (1998) Methods and Applications of
Combined Quantum Mechanical and Molecular Mechanical Methods,
American Chemical Society, Washington DC
21. Antoniou, D., and Schwartz, S. D. (1999) J. Chem. Phys. 110, 7359-7364
22. Antoniou, D., and Schwartz, S. D. (1999) J. Chem. Phys. 110, 465-472
23. Mincer, J. S., and Schwartz, S. D. (2003) J. Phys. Chem. B 107, 366-371
24. Knapp, M. J., Rickert, K., and Klinman, J. P. (2002) J. Am. Chem. Soc. 124,
3865-3874
25. Kohen, A., Cannio, R., Bartolucci, S., and Klinman, J. P. (1999) Nature 399,
496-499
26. Bahnson, B. J., Colby, T. D., Chin, J. K., Goldstein, B. M., and Klinman, J. P.
(1997) Proc. Natl. Acad. Sci. U S A 94, 12797-12802
27. Bahnson, B. J., Park, D. H., Kim, K., Plapp, B. V., and Klinman, J. P. (1993)
Biochemistry 32, 5503-5507
28. Barna, T. M., Khan, H., Bruce, N. C., Barsukov, I., Scrutton, N. S., and
Moody, P. C. (2001) J. Mol. Biol. 310, 433-447
29. Barna, T., Messiha, H. L., Petosa, C., Bruce, N. C., Scrutton, N. S., and
Moody, P. C. (2002) J. Biol. Chem. 277, 30976-30983
30. Fox, K. M., and Karplus, P. A. (1994) Structure 2, 1089-1105
31. Binks, P. R., French, C. E., Nicklin, S., and Bruce, N. C. (1996) Appl.
Environ. Microbiol. 62, 1214-1219
32. French, C. E., Nicklin, S., and Bruce, N. C. (1996) J. Bacteriol. 178, 6623-
6627
33. French, C. E., and Bruce, N. C. (1994) Biochem. J. 301, 97-103
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 31
Basran, Harris, Sutcliffe and Scrutton 31
34. Melmon, K., and Morrelli, H. (1972) Clinical pharmacology: basic principles
in therapeutics, Macmillan Publishing Co., New York
35. Moffat, A., Jackson, J., Moss, M., and Widdop, B. (1986) Clarke's isolation
and identification of drugs, The Pharmaceutical Press, London
36. Craig, D. H., Moody, P. C. E., Bruce, N. C., and Scrutton, N. S. (1998)
Biochemistry 37, 7598-7607
37. Khan, H., Harris, R. J., Barna, T., Craig, D. H., Bruce, N. C., Munro, A. W.,
Moody, P. C., and Scrutton, N. S. (2002) J. Biol. Chem. 277, 21906-21912
38. Viola, R. E., Cook, P. F., and Cleland, W. W. (1979) Anal. Biochem. 96, 334-
340
39. Pollock, V. V., and Barber, M. J. (2001) Biochemistry 40, 1430-1440
40. Gibson, Q. H., Swoboda, B. E. P., and Massey, V. (1964) J. Biol. Chem. 239,
3927-3934
41. Craig, D. H., Barna, T., Moody, P. C., Bruce, N. C., Chapman, S. K., Munro,
A. W., and Scrutton, N. S. (2001) Biochem. J. 359, 315-323
42. Massey, V., and Schopfer, L. M. (1986) J. Biol. Chem. 261, 1215-1222
43. Buckman, J., and Miller, S. M. (1998) Biochemistry 37, 14326-14336
44. Basran, J., Sutcliffe, M. J., and Scrutton, N. S. (1999) Biochemistry 38, 3218-
3222
45. Jonsson, T., Glickman, M. H., Sun, S. J., and Klinman, J. P. (1996) J. Am.
Chem. Soc. 118, 10319-10320
46. Harris, R. J., Meskys, R., Sutcliffe, M. J., and Scrutton, N. S. (2000)
Biochemistry 39, 1189-1198
47. Basran, J., Patel, S., Sutcliffe, M. J., and Scrutton, N. S. (2001) J. Biol. Chem.
276, 6234-6242
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 32
Basran, Harris, Sutcliffe and Scrutton 32
48. Basran, J., Sutcliffe, M. J., and Scrutton, N. S. (2001) J. Biol. Chem. 276,
24581-24587.
49. Faulder, P. F., Tresadern, G., Chohan, K. K., Scrutton, N. S., Sutcliffe, M. J.,
Hillier, I. H., and Burton, N. A. (2001) J. Am. Chem. Soc. 123, 8604-8605
50. Cleland, W. W. (1991) in Enzyme mechanism from isotope effects (Cook, P.
F., ed), pp. 247-265, CRC Press, Boca Raton
51. Kohen, A., Cannio, R., Bartolucci, S., and Klinman, J. P. (1999) Nature 399,
496-499
52. Bell, R. P. (1980) in The Tunnel Effect in Chemistry, pp. 51-140, Chapman
and Hall, London
53. Bahnson, B. J., and Klinman, J. P. (1995) Methods Enzymol. 249, 373-397
54. Scrutton, D. R., Robson, P., and Davies, R. M. (1967) Nature 213, 950-952
55. Bocanegra, J. A., Scrutton, N. S., and Perham, R. N. (1993) Biochemistry 32,
2737-2740
56. Scrutton, N. S., Berry, A., and Perham, R. N. (1990) Nature 343, 38-43
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 33
Basran, Harris, Sutcliffe and Scrutton 33
TABLES
Table 1. Isotope effects obtained from steady-state reactions of MR with 2-
cyclohexenone. Reaction conditions: 50 mM potassium phosphate buffer pH 7.0; 25
°C. Turnover numbers were measured using NADH (NAD2H) and 2-cyclohexenone
at concentrations of 150 µM and 50 mM respectively. Absorbance changes were
monitored at 350 nm and reaction rates calculated using an ε = 5650 M-1 cm-1.
kcat (s-1) H2O kcat (s-1) 2H 2O
NADH NAD2H NADH NAD2H a2.78 ± 0.11
(n = 8) b0.79 ± 0.02
(n = 8) c1.22 ± 0.086
(n = 8) d0.34 ± 0.043
(n = 10)
KIE NADH/NAD2H; Protiated solvent (a/b)
KIE NADH/NAD2H; Deuterated solvent (c/d)
SIE H2O/2H2O; NADH (a/c)
SIE H2O/2H2O; NAD2H (b/d)
Double isotope effect (a/d)
3.52 ± 0.23 3.56 ± 0.7 2.28 ± 0.25 2.32 ± 0.35 8.18 ± 1.36
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 34
Basran, Harris, Sutcliffe and Scrutton 34
SCHEMES
Scheme 1.
Scheme 2
NADHCT.E EH2 + NAD+E + NADH . NAD+EH2k-1
k1 k2 k3
k-2 k-3
NADHE. NADHCT.E EH2 + NAD+E + NADH . NAD+EH2
k1
k-2
k2 k3 k4
k-3k-1 k-4
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 35
Basran, Harris, Sutcliffe and Scrutton 35
FIGURE LEGENDS
Figure 1. Spectral changes accompanying the reduction of MR by NADH.
Conditions: 50 mM potassium phosphate buffer, pH 7.0; 5 °C; MR (20 µM) was
mixed with NADH (200 µM). Panel A, spectral changes captured using a photodiode
array device showing the bleaching of flavin absorbance resulting from reduction by
NADH. Panel B, deconvoluted spectra of reaction intermediates obtained by fitting to
a reversible kinetic scheme A ↔ B ↔ C. Spectrum 1, the oxidised enzyme (species
A); spectrum 2, the E.NADPHCT intermediate (species B); spectrum 3, the
dihydroflavin form of MR (species C).
Figure 2. Reductive half-reaction of MR. Panel A, plot of observed flavin reduction
rate versus coenzyme concentration for the reductive half-reaction of MR. Conditions:
50 mM potassium phosphate buffer, pH 7.0; 25 °C. Filled circles, data for NADH;
fitting to Eq. 2 yields values for klim (56 ± 0.6 s-1) and K (101 ± 7 µM). Open circles,
data for NAD2H; fitting to Eq. 2 yields values for klim (14.4 ± 0.2 s-1) and K (143 ± 10
µM). Panel B, Eyring plots for the limiting rate of flavin reduction in MR. Reactions
with NADH (filled circles) and NAD2H (open circles). ln(A’H) = 12.9 ± 0.2, ln(A’D) =
15 ± 0.3, ∆H‡H = 35.3 ± 0.5 kJ mol-1, ∆H‡D = 43.5 ± 0.8 kJ mol-1. Inset: plot of
ln(KIE) versus 1/T. Rate constants are observed rate constants measured at 5 mM
coenzyme. Each data point is the average of at least five measurements. All errors for
temperature dependence plots are ≤ 5 % of the measured value.
Figure 3. Panel A: Plots of observed flavin reduction rate versus coenzyme
concentration for the reductive half-reaction of PETN reductase. Conditions: 50 mM
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 36
Basran, Harris, Sutcliffe and Scrutton 36
potassium phosphate buffer, pH 7.0; 25 °C. Filled circles, data for NADPH; fitting to
Eq. 2 yields values for klim (34 ± 0.4 s-1) and K (73 ± 4 µM). Open circles, data for
NADP2H; fitting to Eq. 2 yields values for klim (10 ± 0.3 s-1) and K (98 ± 12 µM).
Panel B: Eyring plot for the limiting rate of flavin reduction in PETN reductase.
NADPH (filled circles) and NADP2H (open circles). ln(A’H) = 12.7 ± 0.4, ln(A’D) =
11.3 ± 0.4, ∆H‡H = 36.4 ± 0.9 kJ mol-1, ∆H‡D = 36.6 ± 0.9 kJ mol-1. Inset: plot of
ln(KIE) versus 1/T. Rate constants are observed rate constants measured at 5 mM
coenzyme. Each data point is the average of at least five measurements. All errors for
temperature dependence plots are ≤ 5 % of the measured value. Owing to the
temperature dependence of K, klim values measured above 30 °C were obtained by
fitting plots of the rate of flavin reduction versus substrate concentration to Eq. 1.
Below this temperature, klim values were obtained by performing reactions with
saturating coenzyme (5 mM NADPH).
Figure 4. Plot of kobs1 versus 2-cyclohexenone concentration for the oxidative half-
reaction of MR at 40 °C determined from a double mixing stopped-flow experiment.
Fitting to Eq. 1 yields a K of 1.9 ± 0.1 mM and a klim of 4.0 ± 0.1 s-1. Comparable data
collected at 4 °C yields a K of 2.5 ± 0.2 mM and a klim of 0.92 ± 0.01 s-1.
Figure 5. Proposed scheme for the oxidative half-reaction of MR. The identity of the
proton donor in the oxidative half-reaction is not known.
Figure 6. Sequential stopped-flow absorption transients showing reduction of MR
with NADH and NAD2H and subsequent oxidation by 2-cyclohexenone. Transients
measured in 50 mM potassium phosphate buffer, pH 7.0; 4 °C; 464 nm. Panel A: the
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 37
Basran, Harris, Sutcliffe and Scrutton 37
reductive half-reaction; trace 1, 10 µM MR mixed with 10 µM NADH (kobs1 = 6 ±
0.09 s-1); trace 2, 10 µM MR mixed with 10 µM NAD2H. (kobs1 = 1.36 ± 0.02 s-1). KIE
= 4.4 ± 0.13. Panel B: the oxidative half-reaction; trace 1, 5 µM MR reduced with
5µM NADH and then rapidly mixed with 50 mM 2-cyclohexenone (aging time = 10
s, kobs1 = 1.18 ± 0.005 s-1); trace 2, 5 µM MR reduced with 5µM NAD2H and then
rapidly mixed with 50 mM 2-cyclohexenone (aging time = 30 s, kobs1 = 0.32 ± 0.003 s-
1).
KIE = 3.7 ± 0.05.
Figure 7. Enzyme monitored turnover of MR. Panel A; 10 µM MR was rapidly
mixed with 2 mM NADH and 50 mM 2-cyclohexenone in the stopped-flow apparatus
(reactions were performed in 50 mM potassium phosphate buffer, pH 7.0; 25 °C). The
reaction was monitored at 464 nm as a function of time. Inset: spectra of the enzyme
recorded with a photodiode detector at time points 1, 2 and 3 shown in the main panel.
Panel B; double reciprocal plots for data obtained as described in Gibson et al (40)
collected at 1.5 mM (filled circles), 2 mM (open circles), 7.5 mM (filled squares), 15
mM (open squares), 20 mM (filled triangles), 2-cyclohexenone concentration. For
clarity, only selected plots are shown. Inset: A secondary plot of the ordinate
intercepts obtained from the reciprocal plots as a function of 2-cyclohexenone
concentration.
Figure 8. Eyring plots for steady-state reactions of MR in protiated and deuterated
solvent. MR with NADH (closed circles) and NAD2H (open circles) in protiated
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 38
Basran, Harris, Sutcliffe and Scrutton 38
solvent; ln(A’H) = 2.3 ± 0.4, ln(A’D) = 1.0 ± 0.4, ∆H‡H = 17.6 ± 0.9 kJ mol-1, ∆H‡D =
17.1 ± 0.9 kJ mol-1. MR with NADH in deuterated solvent (filled triangles); ln(A’H) =
1.4 ± 0.3; ∆H‡H = 16.7 ± 0.7 kJ mol-1. Inset: plot of ln(KIE) versus 1/T. Conditions:
50 mM potassium phosphate buffer, pH 7.0; 0.2 µM MR; 150 µM coenzyme; 50 mM
2-cyclohexenone. All errors for temperature dependence plots are ≤ 5 % of the
measured value.
Figure 9. Environmentally coupled tunneling model of enzyme-catalysed reactions.
Reactant (R = ES) and product (P = EP) energy curves for distortion of the protein
structure. The tunneling does not occur until the geometry of the protein is distorted,
so that the R and P curves intersect – the intersection point (*) is the optimum
geometry required for the reaction to occur. Thus, thermally induced conformational
change in the protein is a prerequisite for the tunneling reaction. Top: Hydrogen
Gibbs free energy surface at different positions on the R and P curves—left panel:
reactants’ equilibrium configuration; centre panel: transition configuration; right
panel: products’ equilibrium configuration. (Adapted from (19) and (14).)
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 39
Basran, Harris, Sutcliffe and Scrutton 39
Wavelength (nm)
350 400 450 500 550 600 650
Abs
orba
nce
0
0.05
0.1
0.15
0.2
0.25
Wavelength (nm)
350 400 450 500 550 600 650
ε (M
-1 c
m-1
)
0
2000
4000
6000
8000
10000
12000
14000
A
B
3
2
1
Figure 1
700
bhttp://w
ww
.jbc.org/D
ownloaded from
y guest on A
ugust 3, 2018
700
Page 40
Basran, Harris, Sutcliffe and Scrutton 40
3.1
4
2
0
-2
-4
-6
In (k
/ T)
0
70
60
50
40
30
20
10
0
k obs
1 (s
-1)
A
Figure 2
3.73.63.53.43.33.21000 / T (K-1)
3.73.63.53.43.33.23.1
3
2.5
2
1.5
1
0.5
0
1000 / T (K-1)
In (K
IE)
28002400200016001200800400
[NADH] µM
B by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 41
Basran, Harris, Sutcliffe and Scrutton 41
3.1
2
1
0
-1
-2
-3
-4
-5
-6
In (k
/ T)
0
40
35
30
25
20
15
10
5
0
k obs
(s-1)
A
Figure 3
3
140012001000800600400200
[NADPH] µM
B
by guest on Augu
http://ww
w.jbc.org/
Dow
nloaded from
3.73.63.53.43.33.21000 / T (K-1)
3.73.63.53.43.33.23.1
2.5
2
1.5
1
0.5
0
1000 / T (K-1)
In (K
IE)
st 3, 2018
Page 42
Basran, Harris, Sutcliffe and Scrutton 42
403020100
5
4
3
2
1
0
[2-cyclohexenone] (mM)
k obs
1 (s-1
)
Figure 4
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 43
Basran, Harris, Sutcliffe and Scrutton 43
Figure 5
N
N
N
N
H
O
OCH3
CH3
R
H
H
O
NH2
Asn 189
H A
Oδ -
δ+
NH
N
His 186+
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 44
Basran, Harris, Sutcliffe and Scrutton 44
0
0.1
0.08
0.06
0.04
0.02
0
A46
4
0
0.05
0.04
0.03
0.02
0.01
0
A46
4
A
B
Figure 6
30252015105
Time (s)
2
1
by guehttp://w
ww
.jbc.org/D
ownloaded from
1
st on August
4
2
3, 2018
2016128
Time (s)
Page 45
Basran, Harris, Sutcliffe and Scrutton 45
0.50 30025020015010050
0.1
0.08
0.06
0.04
0.02
Time (s)
A 464
806040200
3.5
3
2.5
2
1.5
1
0.5
0
1 / [NADH] (mM-1)
1 / r
ate
(s)
0.80.60.40.20
2
1.5
1
0.5
0
1 / [2-cyclohexenone] (mM-1)
Inte
rcep
t (s)
700600500400
0.1
0.08
0.06
0.04
0.02
0
Wavelength (nm)
Abs
orba
nce
A
B
1
2
3
12 3
Figure 7
350
by http://w
ww
.jbc.org/D
ownloaded from
100
guest on August 3, 2018
Page 46
Basran, Harris, Sutcliffe and Scrutton 46
3.73.63.53.43.33.2
-2
-3
-4
-5
-6
-7
-8
1000 / T (K-1)
In (v
i / T
)
3.73.63.53.43.33.2
3
2.5
2
1.5
1
0.5
0
1000 / T (K-1)
In (K
IE)
Figure 8
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 47
Basran, Harris, Sutcliffe and Scrutton 47
Figure 9
Nuclear Configurati
∆G
R
*
P
∆G°
λ
http://D
ownloaded from
*
on
by guest on August 3, 2018
ww
w.jbc.org/
Page 48
Jaswir Basran, Richard J. Harris, Michael J. Sutcliffe and Nigel S. Scruttontetranitrate reductase
reductase and in the reductive half-reaction of the homologous pentaerythritol H-tunneling in the multiple H-transfers of the catalytic cycle of morphinone
published online August 26, 2003J. Biol. Chem.
10.1074/jbc.M305983200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on August 3, 2018
http://ww
w.jbc.org/
Dow
nloaded from