and Visual FunctionChloé Cartier1, Dave Saint-Amour1, 2, 3
1Département de psychologie, Université du Québec à Montréal, Montréal, Québec, Canada, 2Département d’ophtalmologie, Université de Montréal, Montréal, Québec, Canada, 3Centre de recherche, Centre hospitalier
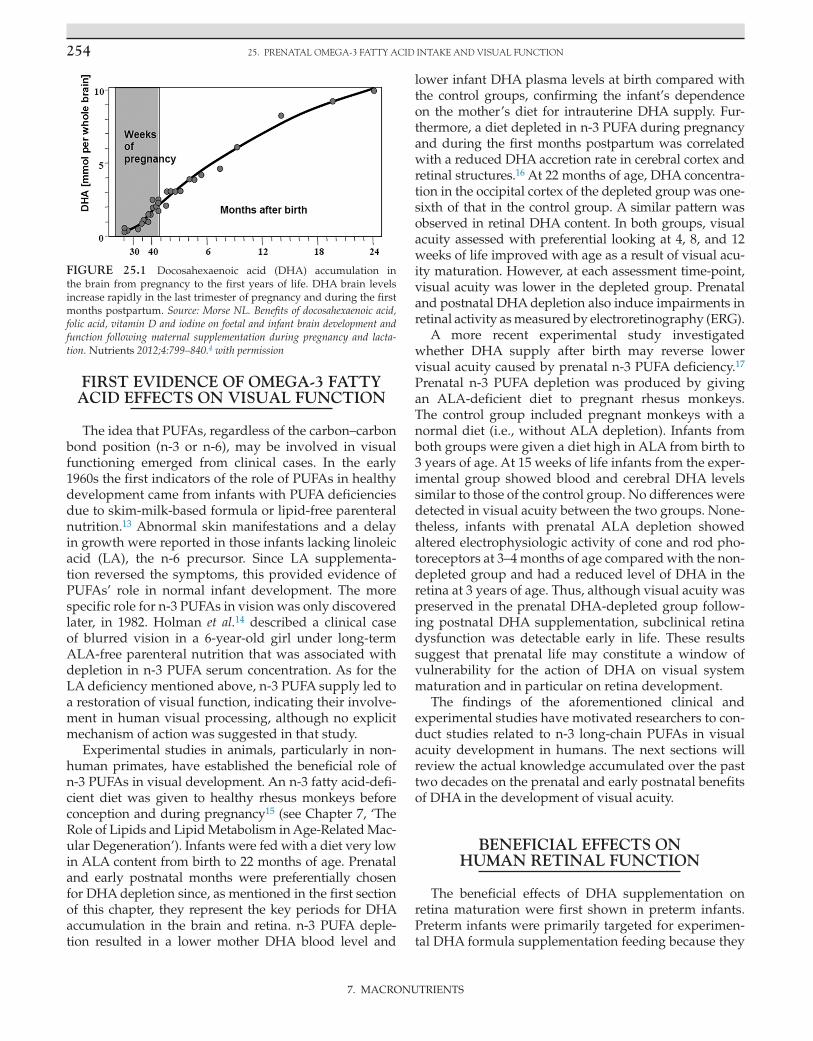
Lipids influence neuronal function by modifying the characteristics of the membrane, gene expression, and cell signaling, all of which play a critical role in neu-rotransmission, metabolism, growth, and cell differ-entiation.1 A considerable accumulation of long-chain polyunsaturated fatty acids (PUFAs) occurs in neural and retinal membranes during gestation, and it contin-ues throughout the first postnatal years, a period that is associated with a substantial growth of the brain2, 3 (Fig. 25.1). Inadequate intake of PUFAs can perturb cerebral maturational processes and thus have functional conse-quences on sensory and cognitive neurodevelopment at birth or later in life.
Docosahexaenoic acid (DHA) is the principal omega-3 (n-3) PUFA found in the mammalian central nervous system. It is highly concentrated in the gray matter of the cerebral cortex as well as in phospholip-ids of the photoreceptor outer segment membranes of the retina.5 Although DHA can be synthesized in the liver from its precursor, α-linolenic acid (ALA, 18:3n-3), only small amounts can be produced in humans. It is estimated that healthy adults can convert about 0.1% of ALA from their diet to DHA.6 Because of the low human capacity to synthesize DHA from its pre-cursors, in particular early in life,7 the source of DHA in the fetus and breastfed infants is highly dependent on maternal-to-fetal transfer before birth and breast milk postnatally. Indeed, there is a positive correlation between the mother’s circulating DHA level intakes
Handbook of Nutrition, Diet, and the Eyehttp://dx.doi.org/10.1016/B978-0-12-401717-7.00025-3
25
during pregnancy and lactation and infant DHA sta-tus at birth or in the few months postpartum. Maternal dietary intake of DHA is crucial for adequate DHA sup-ply to the fetus and infant.8–10 Pregnant women need to have a diet with enough DHA-containing products for their own health and to supply sufficient DHA to their developing infant. Because fetal DHA demands deplete maternal DHA stores during pregnancy, several factors may influence maternal availability to the fetus, such as the period (prenatal or postnatal), number of infants per pregnancy, and number of months since the last deliv-ery.11 Worldwide variations in dietary patterns as well as regional and personal dietary habits lead to impor-tant differences in DHA breast milk content between women and raise the question of whether DHA intakes may be sufficient in some populations with a Western diet to provide enough DHA to the developing baby. For example, women with higher dietary consumption of fish or seafood, found in Japan, the Philippines, and Chile, were reported to have two- to five-fold higher DHA milk contents than women with low fish product consumption12 (in Canada and the USA).
In recent decades, a growing body of evidence from epidemiologic and experimental studies has linked DHA prenatal exposure and DHA formula supplemen-tation with visual acuity in preterm or term infants. As described in the following sections, these studies have also provided a better understanding of the implications of DHA in visual processes and highlighted the necessity of an adequate DHA supply for optimal visual system development.
FIRST EVIDENCE OF OMEGA-3 FATTY ACID EFFECTS ON VISUAL FUNCTION
The idea that PUFAs, regardless of the carbon–carbon bond position (n-3 or n-6), may be involved in visual functioning emerged from clinical cases. In the early 1960s the first indicators of the role of PUFAs in healthy development came from infants with PUFA deficiencies due to skim-milk-based formula or lipid-free parenteral nutrition.13 Abnormal skin manifestations and a delay in growth were reported in those infants lacking linoleic acid (LA), the n-6 precursor. Since LA supplementa-tion reversed the symptoms, this provided evidence of PUFAs’ role in normal infant development. The more specific role for n-3 PUFAs in vision was only discovered later, in 1982. Holman et al.14 described a clinical case of blurred vision in a 6-year-old girl under long-term ALA-free parenteral nutrition that was associated with depletion in n-3 PUFA serum concentration. As for the LA deficiency mentioned above, n-3 PUFA supply led to a restoration of visual function, indicating their involve-ment in human visual processing, although no explicit mechanism of action was suggested in that study.
Experimental studies in animals, particularly in non-human primates, have established the beneficial role of n-3 PUFAs in visual development. An n-3 fatty acid-defi-cient diet was given to healthy rhesus monkeys before conception and during pregnancy15 (see Chapter 7, ‘The Role of Lipids and Lipid Metabolism in Age-Related Mac-ular Degeneration’). Infants were fed with a diet very low in ALA content from birth to 22 months of age. Prenatal and early postnatal months were preferentially chosen for DHA depletion since, as mentioned in the first section of this chapter, they represent the key periods for DHA accumulation in the brain and retina. n-3 PUFA deple-tion resulted in a lower mother DHA blood level and
FIGURE 25.1 Docosahexaenoic acid (DHA) accumulation in the brain from pregnancy to the first years of life. DHA brain levels increase rapidly in the last trimester of pregnancy and during the first months postpartum. Source: Morse NL. Benefits of docosahexaenoic acid, folic acid, vitamin D and iodine on foetal and infant brain development and function following maternal supplementation during pregnancy and lacta-tion. Nutrients 2012;4:799–840.4 with permission
D INTAKE AND VISUAL FUNCTION
UTRIENTS
lower infant DHA plasma levels at birth compared with the control groups, confirming the infant’s dependence on the mother’s diet for intrauterine DHA supply. Fur-thermore, a diet depleted in n-3 PUFA during pregnancy and during the first months postpartum was correlated with a reduced DHA accretion rate in cerebral cortex and retinal structures.16 At 22 months of age, DHA concentra-tion in the occipital cortex of the depleted group was one-sixth of that in the control group. A similar pattern was observed in retinal DHA content. In both groups, visual acuity assessed with preferential looking at 4, 8, and 12 weeks of life improved with age as a result of visual acu-ity maturation. However, at each assessment time-point, visual acuity was lower in the depleted group. Prenatal and postnatal DHA depletion also induce impairments in retinal activity as measured by electroretinography (ERG).
A more recent experimental study investigated whether DHA supply after birth may reverse lower visual acuity caused by prenatal n-3 PUFA deficiency.17 Prenatal n-3 PUFA depletion was produced by giving an ALA-deficient diet to pregnant rhesus monkeys. The control group included pregnant monkeys with a normal diet (i.e., without ALA depletion). Infants from both groups were given a diet high in ALA from birth to 3 years of age. At 15 weeks of life infants from the exper-imental group showed blood and cerebral DHA levels similar to those of the control group. No differences were detected in visual acuity between the two groups. None-theless, infants with prenatal ALA depletion showed altered electrophysiologic activity of cone and rod pho-toreceptors at 3–4 months of age compared with the non-depleted group and had a reduced level of DHA in the retina at 3 years of age. Thus, although visual acuity was preserved in the prenatal DHA-depleted group follow-ing postnatal DHA supplementation, subclinical retina dysfunction was detectable early in life. These results suggest that prenatal life may constitute a window of vulnerability for the action of DHA on visual system maturation and in particular on retina development.
The findings of the aforementioned clinical and experimental studies have motivated researchers to con-duct studies related to n-3 long-chain PUFAs in visual acuity development in humans. The next sections will review the actual knowledge accumulated over the past two decades on the prenatal and early postnatal benefits of DHA in the development of visual acuity.
BENEFICIAL EFFECTS ON HUMAN RETINAL FUNCTION
The beneficial effects of DHA supplementation on retina maturation were first shown in preterm infants. Preterm infants were primarily targeted for experimen-tal DHA formula supplementation feeding because they
OmEgA-3 FATTy ACId ExPOsuR
were hypothesized to have a higher risk of DHA defi-ciency than term infants, owing to their shorter intrauter-ine life and immature central nervous system. Very low birth weight healthy preterm newborns were assigned to receive different formulas with or without DHA supple-mentation, from 10 days to 57 weeks postconception.17–19 A breastfed group of preterm infants was also studied. Cone and rod photoreceptor activity was evaluated at 36 and 57 weeks postconception using ERG. Infants with-out supplementation presented a delay in retinal mat-uration relative to the two other groups. Furthermore, a higher luminance was necessary to elicit rod activa-tion in the no-supplementation group and amplitude responses remained lower than in infants in the DHA-supplemented and breastfed groups. Rod activity was similar in these last two groups. In all cases, n-3 PUFA plasma and erythrocyte levels were significantly asso-ciated with improved electrophysiologic rod response. No differences in ERG between groups were seen at 57 weeks postconception, however, suggesting that the ret-ina accumulated sufficient DHA to achieve normal func-tion with time, even on very low ALA intakes.
Preterm infants are known to be a population at risk of developing retinopathy of prematurity, an eye dis-ease characterized by abnormal retinal vessel growth (see Chapter 7, ‘The Role of Lipids and Lipid Metabo-lism in Age-Related Macular Degeneration’). Infants may recover spontaneously from the pathology or it can lead to important vision complications. Pawlik et al.20 conducted a clinical trial on preterm newborns under parenteral nutrition in which one group was given a classic fat emulsion and another group was given a fish oil emulsion containing DHA. While infants from both groups developed retinopathy in a similar way, a signifi-cantly better spontaneous recovery was associated with fish oil administration in the first days of life. Recent evi-dence suggests that the protective mechanism of action of the n-3 PUFA effect may be mediated by blood ves-sel growth. Indeed, 4-hydroxydocosahexaenoic acid (4-HDHA), a DHA metabolite, has been found to have antiangiogenic properties and to protect against abnor-mal retinal vessel proliferation.21
Retinal sensitivity has also been studied in the first days of life using ERG in healthy term infants from mothers supplemented with fish oil from 15 weeks of gestation until delivery.22 In contrast to studies con-ducted on preterm infants, no differences in retinal activity for either rod or cone responses were detected in these infants compared with placebo, evidence that adequate DHA for retinal development is accumulated in utero. However, subtle differences may still exist; for example, in both groups, DHA in cord blood correlated positively with retinal responses, reaffirming the neces-sity of an adequate DHA dietary intake during preg-nancy for optimal retina development. In addition to
7. MACRON
E And InFAnT VIsuAl ACuITy 255
their benefits in infants, n-3 PUFAs also constitute an essential nutrient for maintaining retinal functioning throughout life. Indeed, DHA dietary intakes have been found to provide protection against the risk of develop-ing age-related macular degeneration,23 the main cause of vision loss in people over 65 years of age (see Chapter 2, ‘Age-Related Macular Degeneration: An Overview’).
Different mechanisms have been proposed to explain why DHA deficiency may affect retinal function. It is likely that n-3 PUFA deficiency in the retina is mediated by an alteration of the photoreceptors per se, consider-ing the important role of n-3 fatty acids in membrane fluidity and phototransduction of rhodopsin in rods.24 Although the specific mechanisms of n-3 PUFA action on the retina remain to be defined, these studies provide evidence to promote n-3 PUFA intake, particularly from fish, to protect and optimize the eye structure and retinal function, especially in young and elderly populations.
OMEGA-3 FATTY ACID EXPOSURE AND INFANT VISUAL ACUITY
Acuity is the visual function most studied clinically in relation to n-3 PUFA exposure. Because visual acuity depends on the maturation and integrity of the retina, the thalamocortical pathway, and the primary visual cor-tex, it provides an excellent probe to assess the visual system.
Observational studies comparing infants fed human milk and formula and using both electrophysiologic and behavioral measurements of visual acuity have found improved performances in breastfed infants.25–27 Because human milk is known to contain DHA12 and since comparisons were made with non-DHA-supple-mented formula, it was suggested that this beneficial effect was, at least in part, a result of the action of DHA on visual pathways. To confirm this hypothesis, ran-domized clinical trials were conducted in term infants with n-3 PUFA postnatal supplementation. Using visual evoked potentials (VEPs), scalp recording, and the Teller Acuity Card behavioral procedure (Fig. 25.2), several studies have shown that postnatal DHA supplementa-tion improves visual acuity during the first months of life27,29–31 and at 12 months,32 while others have failed to demonstrate such an effect.27,33 Differential results may be due to heterogeneity between studies regarding sample sizes, confounding factors such as family stimu-lation, DHA sources, duration, and dose, as well as the methods used to assess visual function.
In contrast, studies conducted on preterm infants have provided more consistent results and nearly all have concluded that there is a beneficial effect of DHA postnatal supplementation on visual acuity develop-ment.18,25,34–36 In these studies, preterm newborns were
UTRIENTS
25. PRENATAL OMEGA-3 FATTY ACID INTAKE AND VISUAL FUNCTION256
randomly assigned to formula enriched with n-3 PUFAs or not. Visual acuity was measured between birth and 1 year of age with behavioral and electrophysiologic assessments. Using a behavioral assessment, Carlson and colleagues34,35 showed higher visual acuity in the preterm group supplemented with DHA from fish oil at 2 and 4 months of age, although this difference was not observed in the interval from 6.5 to 12 months. In O’Connor’s study,36 the researchers did not notice a sig-nificant effect of n-3 PUFA supplementation on visual acuity at 6 months of age using the same Teller Acuity Card procedure, but they found better visual acuity in supplemented infants when assessed with VEPs, evi-dence that the electrophysiologic measurement is more sensitive in detecting DHA-related beneficial effects on visual function.
As discussed above, the beneficial effects of n-3 PUFAs on visual acuity have mostly been detected before 6 months of age. Morale et al.37 reported that, in a large group of DHA-supplemented children (n = 243), the improved visual acuity observed with VEP record-ing during the first months of life was also present at 1 year. Thus, the fact that clear enhancements of visual acuity are not commonly detected after 6 months of age does not necessarily mean that they do not occur. First, as illustrated above, the method chosen to measure visual function (i.e., from behavior or brain activity) can make a difference. Second, since visual acuity develops rapidly during the first 6 months of life, it is possible that this function reaches a plateau, making it difficult to high-light differences between groups later in life because of ceiling effects.38
The fact that more robust DHA beneficial effects were reported with preterm infants suggests that the last months of gestation are a critical time window for DHA to affect visual acuity development. Accordingly, observational human studies have reported a signifi-cant relationship between prenatal n-3 PUFA exposure
7. MACRON
and visual acuity. Jacobson et al.39 studied DHA prena-tal exposure on visual acuity in term infants in an Inuit cohort from Nunavik, the northernmost region of Que-bec. Inuit individuals are chronically exposed to high amounts of n-3 PUFA because of their fish and marine mammal diet. Using the Teller Acuity Card, the authors found that cord DHA phospholipids were associated with better visual acuity at 6 months of life with no remaining effects detected at 11 months. To the present authors’ knowledge, three clinical trials have evaluated the effect of DHA supplementation during pregnancy on visual acuity development in infants using the same protocol, in which pregnant women were given DHA or placebo from midpregnancy until delivery.40–42 Behav-ioral assessments were conducted at birth and at 2, 4, and 6 months of age to evaluate the impact of DHA prenatal exposure on visual acuity development. Innis and collaborators40 reported better visual acuity in the supplemented group at 2 months of age compared with the placebo group. Judge et al.41 found the same positive effect of DHA at 4 months but no associations between DHA and visual acuity 2 months later. In Malcolm’s study,42 no difference in visual acuity performance was detected between the DHA-supplemented and the con-trol group at birth or at 50 and 66 weeks postconception. Nonetheless, for all infants, either in the placebo or in the DHA group, a significant positive association was found between the DHA level measured at birth and postnatal visual acuity performance.
In conclusion, while the beneficial effect of DHA on visual acuity maturation seems to be clearly demon-strated in preterm infants, the results obtained with pre-natal or postnatal DHA exposure in term populations are less consistent. Although some studies find enhanced visual acuity related to DHA prenatal exposure only in the first few months of life, this may be due to the fact that the behavioral assessment of acuity is not sensitive to longer term effects on visual acuity.
UTRIENTS
lOng-TERm BEnEFITs OF dEVElOPmEnTAl ExPOsu
7. MACRON
LONG-TERM BENEFITS OF DEVELOPMENTAL EXPOSURE
TO OMEGA-3 POLYUNSATURATED FATTY ACIDS
Although the role of n-3 PUFAs in visual acuity has been widely investigated, most of the studies focused only on early postnatal life, and it has not been estab-lished whether DHA exposure impacts other visual and cognitive functions.
In 2007, Birch et al.26 conducted a randomized clinical trial with 84 healthy term infants to evaluate the long-term beneficial effect of formula supplementation with DHA on visual acuity development. Infants were breast-fed, formula fed with DHA, or formula fed with no sup-plementation from birth to 17 weeks of age. At 4 years of age, visual acuity measured in the nonsupplemented group was found to be lower than in the supplemented and the breastfed groups. Although the difference was subtle and only detectable in the right eye, these results suggest that early postnatal DHA administration has long-term benefits for visual maturation.
In addition to standard visual acuity, some authors have included measurements of stereoacuity to evalu-ate the effect of DHA exposure on visual cortex matu-ration.29,43,44 Stereoacuity, or stereoscopic acuity, is the smallest detectable depth difference induced by binocu-lar disparity. In these studies, stereoacuity performance was compared between infants formula fed with DHA supplementation and infants formula fed without sup-plementation.29 Comparisons were also made between formula-fed and breastfed infants, and maternal intake of DHA during pregnancy was taken into account.43,44 At 4 months of age better stereoacuity was detected in DHA-supplemented infants compared with the non-supplemented group.29 At 3.5 years of age, stereoacuity was higher in breastfed children than in the formula-fed group.29 It is worth noting that better stereoacuity was observed at 3.5 years in children whose mothers had high fish oil intake during pregnancy compared with children whose mothers had low DHA intake. These results sug-gest a beneficial action of not only postnatal but also pre-natal exposure to DHA on visual brain maturation.
More recently, a VEP study was conducted in 136 school-aged (11-year-old) Inuit children in Nunavik to investigate the long-term beneficial effects of prenatal intake of n-3 PUFAs on visual functions.45 Because fish and marine mammals represent an important part of the Inuit diet, n-3 PUFA intake is substantially greater in this population than in southern Canada,46 although this population is concomitantly exposed to environmental contaminants such as methylmercury, polychlorinated biphenyls, and lead.47 In this study, several variables including contaminants were controlled statistically to isolate the effects of n-3 PUFAs on children’s visual
RE TO OmEgA-3 POlyunsATuRATEd FATTy ACIds 257
UTRIENTS
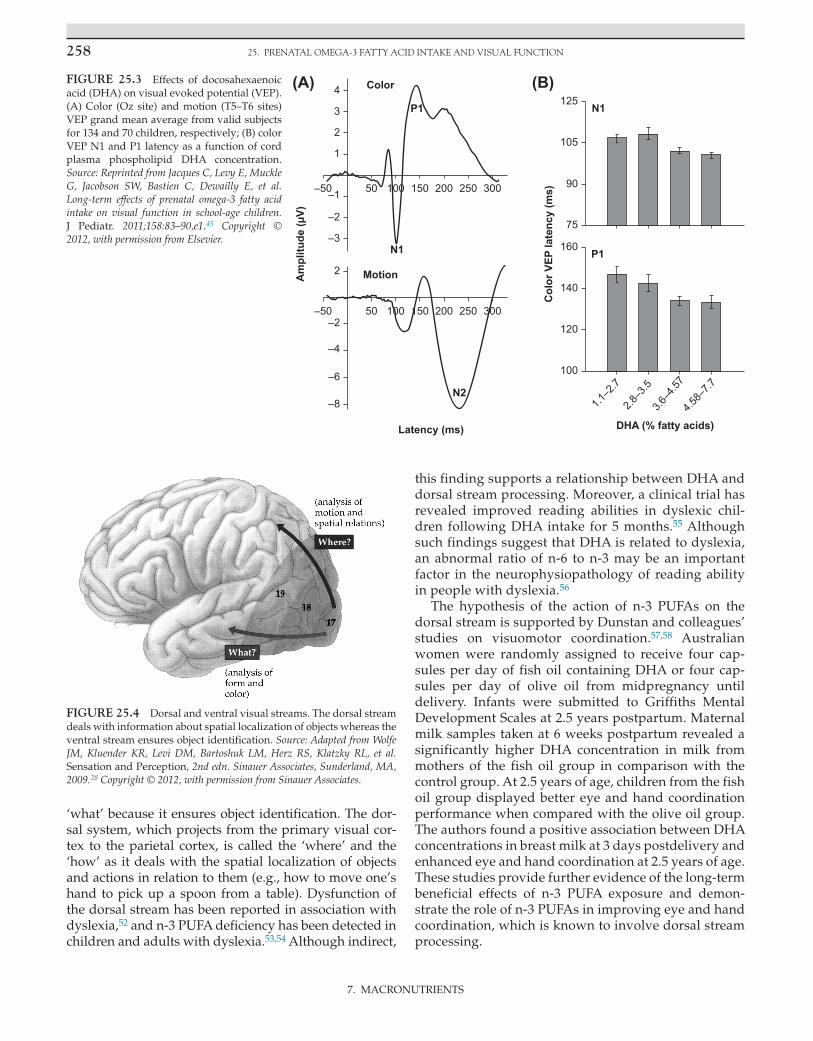
processing. DHA amounts were measured in umbili-cal cord blood samples taken at delivery as well as at the time of testing to control current exposure. Different VEP paradigms were used to assess parvocellular and magnocellular brain responses, two pathways that carry different types of visual information.48 The magnocel-lular pathway is optimally sensitive to low-to-medium spatial frequencies, low achromatic contrasts, and high temporal frequencies, while the parvocellular pathway is optimally sensitive to medium-to-high spatial fre-quencies, high contrast, and low temporal frequency. As a consequence, the magnocellular pathway is more sensitive to motion, whereas the parvocellular pathway plays a major role in processing stimulus detail and chromatic analysis. Considering that the parvocellular system mediates visual acuity, the authors hypothesized that the beneficial effects of prenatal n-3 PUFA intake on acuity observed in infancy39 continue to be evident in childhood, as revealed by parvocellular-related VEPs. Accordingly, the results showed a beneficial impact of prenatal exposure to n-3 PUFAs in school-aged children. Indeed, after adjustment for confounders, cord plasma DHA was associated with shorter latencies of the N1 and P1 components of the isoluminant pattern-reversal VEPs, whereas no effects were found for low contrasted motion-onset VEPs (Fig. 25.3). These findings support the notion that the beneficial effects of fetal DHA intake on visual development may persist into late childhood and suggest that this effect is specific to parvocellular function. This effect was subtle and subclinical, as it was not significantly related to behavioral measurement of visual acuity. As a result, the VEP findings in this study may be difficult to interpret in terms of clinical signifi-cance, but they are clearly non-negligible in terms of optimal visual function.
The aforementioned studies in Nunavik39,45 showed beneficial effects on visual function in relation to cord but not child plasma DHA. This suggests that DHA intake during the prenatal period plays a critical role in the early development of the visual system that may still be detectable at school age and during adulthood, although data are missing to support the latter hypoth-esis. Although DHA concentrations in the cord phos-pholipid plasma in Arctic Quebec are about three times higher than in southern Quebec,46 cord DHA levels in Nunavik are similar to those reported in several other Western countries, notably in Europe49 and in Massachu-setts in the USA.50
Recently, it has been proposed that the effects of n-3 PUFAs on visual cortical processing may preferen-tially involve the dorsal stream,51 which is often con-trasted with the so-called ventral visual streams as the two systems are anatomically and functionally segre-gated (Fig. 25.4). The ventral system, which relays pri-mary visual cortex input to temporal areas, refers to the
25. PRENATAL OMEGA-3 FATTY ACID INTAKE AND VISUAL FUNCTION258
‘what’ because it ensures object identification. The dor-sal system, which projects from the primary visual cor-tex to the parietal cortex, is called the ‘where’ and the ‘how’ as it deals with the spatial localization of objects and actions in relation to them (e.g., how to move one’s hand to pick up a spoon from a table). Dysfunction of the dorsal stream has been reported in association with dyslexia,52 and n-3 PUFA deficiency has been detected in children and adults with dyslexia.53,54 Although indirect,
this finding supports a relationship between DHA and dorsal stream processing. Moreover, a clinical trial has revealed improved reading abilities in dyslexic chil-dren following DHA intake for 5 months.55 Although such findings suggest that DHA is related to dyslexia, an abnormal ratio of n-6 to n-3 may be an important factor in the neurophysiopathology of reading ability in people with dyslexia.56
The hypothesis of the action of n-3 PUFAs on the dorsal stream is supported by Dunstan and colleagues’ studies on visuomotor coordination.57,58 Australian women were randomly assigned to receive four cap-sules per day of fish oil containing DHA or four cap-sules per day of olive oil from midpregnancy until delivery. Infants were submitted to Griffiths Mental Development Scales at 2.5 years postpartum. Maternal milk samples taken at 6 weeks postpartum revealed a significantly higher DHA concentration in milk from mothers of the fish oil group in comparison with the control group. At 2.5 years of age, children from the fish oil group displayed better eye and hand coordination performance when compared with the olive oil group. The authors found a positive association between DHA concentrations in breast milk at 3 days postdelivery and enhanced eye and hand coordination at 2.5 years of age. These studies provide further evidence of the long-term beneficial effects of n-3 PUFA exposure and demon-strate the role of n-3 PUFAs in improving eye and hand coordination, which is known to involve dorsal stream processing.
REFERE
7. MACRON
Animal and human studies have also explored the effect of DHA on visual attention.59–62 In humans, the Fagan Test of Infant Intelligence is commonly used to assess visual attention and recognition in infants. This test measures the infant’s natural tendency to spend more time looking at a novel stimulus than at a familiar stimulus. The infant is presented with two identical faces during a predetermined period. Then, the same items are presented a second time but each one is paired with a novel unfamiliar item. The total time the infant spends looking at each item is recorded. Using this test, two studies have reported shorter total looking duration in infants supplemented with DHA formula with no effects on visual recognition.59,61 According to the authors, this may reflect ‘more rapid visual information processes and a more mature attention’,59 including a better capac-ity to disengage or shift attention. Kannass et al.63 used a multiple object free-play task, in which children aged 12 and 18 months were presented with objects to explore, in order to assess attention in relation to DHA status at birth. Children were presented different toys at the same time. The time for which the children looked at each toy, the number of looks at the toys, and episodes of inatten-tion (i.e., not looking at the toys) were evaluated. While a shorter look duration is usually considered to reflect effi-cient visual processing during the first year of life, look duration typically increases after 1 year because of the time-course of attention development in order for the child to become able to maintain his or her attention on a task and to resist being distracted (sustained attention). Kannass et al. found a positive association between the mother’s DHA red blood cell phospholipid concentra-tion at delivery and the total duration of looking at the toys. They also found fewer episodes of inattention in children with high maternal DHA status at birth.63 These results show a positive effect of DHA exposure on visual attention development beyond the first year of life. The neural basis of the beneficial impact of DHA on attention is unknown, but the frontal cortex may be involved.64
All together, the studies discussed in this section sug-gest that n-3 PUFA preferentially affects the parvocellular system as far as subcortical and low-level visual structures are concerned and the dorsal stream at the cortical level. Of the two cortical visual streams, the dorsal one has been considered more ‘vulnerable’,65 and this may explain why the impact of n-3 PUFAs is more evident for dorsal visual function. However, the dorsal and ventral streams do interact, and normal visual perception is intrinsically dependent on both streams. It would be rather provoca-tive to claim that n-3 PUFAs do not influence ventral visual stream processing. Moreover, other cortical areas, such as the frontal cortex, are likely to be involved.64 There is a lack of data on the mechanisms underlying how n-3 PUFAs improve visual function for both prenatal and postnatal exposure. Further studies are therefore needed.
• The n-3 long-chain polyunsaturated fatty acids are highly concentrated in the gray matter of the brain and in photoreceptors of the retina.
• This localization has been studied to determine whether n-3 long-chain PUFAs are required for optimal visual acuity and cognitive development.
• Clinical cases and depletion experimental studies have highlighted the benefits of n-3 long-chain PUFAs for vision.
• Docosahexaenoic acid (DHA) enhances retinal function in humans and may protect against age-related eye disease.
• Supplementation with n-3 long-chain PUFAs has a positive effect on visual acuity development in preterm infants, and results are found more consistently than in term infants.
• A few studies have also found long-term beneficial effects of n-3 long-chain PUFAs on visual acuity and suggested that n-3 long-chain PUFAs act on specific visual systems (i.e., parvocellular pathway and dorsal pathway).
• Effects of n-3 long-chain PUFAs on visual attention and eye/hand coordination while requiring normal visual acuity are probably related to early mental and motor development, reflected in higher brain DHA accumulation.
AcknowledgmentsWe are very grateful to Susan Carlson for her comments in revising the draft of this chapter. This research was supported by the Canadian Institutes of Health Research and the Vision Health Research Network of the Fonds de recherche du Québec, awarded to Dave Saint-Amour.
References 1. Jump DB, Clarke SD. Regulation of gene expression by dietary fat.
Annu Rev Nutr 1999;19:63–90. 2. Clandinin MT, Chappell JE, Leong S, Heim T, Swyer PR, Chance GW.
Extrauterine fatty acid accretion in infant brain: implications for fatty acid requirements. Early Hum Dev 1980;4:131–8.
3. Martinez M. Tissue levels of polyunsaturated fatty acids during early human development. J Pediatr 1992;120(4 Pt 2):S129–38.
4. Morse NL. Benefits of docosahexaenoic acid, folic acid, vitamin D and iodine on foetal and infant brain development and function following maternal supplementation during pregnancy and lacta-tion. Nutrients 2012;4:799–840.
5. Giusto NM, Pasquare SJ, Salvador GA, Castagnet PI, Roque ME, Ilincheta de Boschero MG. Lipid metabolism in vertebrate retinal rod outer segments. Prog Lipid Res 2000;39:315–91.
6. Plourde M, Cunnane SC. Extremely limited synthesis of long chain polyunsaturates in adults: implications for their dietary essentiali-ty and use as supplements. Appl Physiol Nutr Metab 2007;32:619–34.
7. Salem Jr N, Wegher B, Mena P, Uauy R. Arachidonic and docosa-hexaenoic acids are biosynthesized from their 18-carbon precur-sors in human infants. Proc Natl Acad Sci USA 1996;93:49–54.
8. Dunstan JA, Mori TA, Barden A, Beilin LJ, Holt PG, Calder PC, et al. Effects of n-3 polyunsaturated fatty acid supplementation in pregnancy on maternal and fetal erythrocyte fatty acid composi-tion. Eur J Clin Nutr 2004;58:429–37.
9. Helland IB, Saugstad OD, Saarem K, Van Houwelingen AC, Nylander G, Drevon CA. Supplementation of n-3 fatty acids dur-ing pregnancy and lactation reduces maternal plasma lipid lev-els and provides DHA to the infants. J Matern Fetal Neonatal Med 2006;19:397–406.
10. Marc I, Plourde M, Lucas M, Sterescu A, Piedboeuf B, Dufresne A, et al. Early docosahexaenoic acid supplementation of mothers dur-ing lactation leads to high plasma concentrations in very preterm infants. J Nutr 2011;141:231–6.
11. Al MD, van Houwelingen AC, Hornstra G. Relation between birth order and the maternal and neonatal docosahexaenoic acid status. Eur J Clin Nutr 1997;51:548–53.
12. Yuhas R, Pramuk K, Lien EL. Human milk fatty acid composition from nine countries varies most in DHA. Lipids 2006;41:851–8.
13. Caldwell MD, Jonsson HT, Othersen Jr HB. Essential fatty acid de-ficiency in an infant receiving prolonged parenteral alimentation. J Pediatr 1972;81:894–8.
14. Holman RT, Johnson SB, Hatch TF. A case of human linolenic acid deficiency involving neurological abnormalities. Am J Clin Nutr 1982;35:617–23.
15. Neuringer M, Connor WE, Van Petten C, Barstad L. Dietary ome-ga-3 fatty acid deficiency and visual loss in infant rhesus monkeys. J Clin Invest 1984;73:272–6.
16. Anderson GJ, Neuringer M, Lin DS, Connor WE. Can prenatal n-3 fatty acid deficiency be completely reversed after birth? Effects on retinal and brain biochemistry and visual function in rhesus mon-keys. Pediatr Res 2005;58:865–72.
17. Birch DG, Birch EE, Hoffman DR, Uauy RD. Retinal development in very-low-birth-weight infants fed diets differing in omega-3 fatty acids. Invest Ophthalmol Vis Sci 1992;33:2365–76.
18. Hoffman DR, Birch EE, Birch DG, Uauy RD. Effects of supplemen-tation with omega 3 long-chain polyunsaturated fatty acids on reti-nal and cortical development in premature infants. Am J Clin Nutr 1993;57(Suppl. 5). 807–12S.
19. Uauy RD, Birch DG, Birch EE, Tyson JE, Hoffman DR. Effect of dietary omega-3 fatty acids on retinal function of very-low-birth-weight neonates. Pediatr Res 1990;28:485–92.
20. Pawlik D, Lauterbach R, Turyk E. Fish-oil fat emulsion supplemen-tation may reduce the risk of severe retinopathy in VLBW infants. Pediatrics 2011;127:223–8.
21. Sapieha P, Stahl A, Chen J, Seaward MR, Willett KL, Krah NM, et al. 5-Lipoxygenase metabolite 4-HDHA is a mediator of the anti-angiogenic effect of omega-3 polyunsaturated fatty acids. Sci Transl Med 2011;3:69ra12.
22. Malcolm CA, Hamilton R, McCulloch DL, Montgomery C, Weaver LT. Scotopic electroretinogram in term infants born of mothers supple-mented with docosahexaenoic acid during pregnancy. Invest Oph-thalmol Vis Sci 2003;44:3685–91.
23. Ho L, van Leeuwen R, Witteman JC, van Duijn CM, Uitterlinden AG, Hofman A, et al. Reducing the genetic risk of age-related mac-ular degeneration with dietary antioxidants, zinc, and omega-3 fatty acids: the Rotterdam study. Arch Ophthalmol 2011;129:758–66.
24. Kurlak LO, Stephenson TJ. Plausible explanations for effects of long chain polyunsaturated fatty acids (LCPUFA) on neonates. Arch Dis Child Fetal Neonatal Ed 1999;80:F148–54.
25. Birch EE, Birch DG, Hoffman DR, Uauy R. Dietary essential fatty acid supply and visual acuity development. Invest Ophthalmol Vis Sci 1992;33:3242–53.
INTAKE AND VISUAL FUNCTION
UTRIENTS
26. Birch EE, Garfield S, Castaneda Y, Hughbanks-Wheaton D, Uauy R, Hoffman D. Visual acuity and cognitive outcomes at 4 years of age in a double-blind, randomized trial of long-chain polyun-saturated fatty acid-supplemented infant formula. Early Hum Dev 2007;83:279–84.
27. Carlson SE, Ford AJ, Werkman SH, Peeples JM, Koo WW. Visual acuity and fatty acid status of term infants fed human milk and formulas with and without docosahexaenoate and arachidonate from egg yolk lecithin. Pediatr Res 1996;39:882–8.
28. Wolfe JM, Kluender KR, Levi DM, Bartoshuk LM, Herz RS, Klatzky RL, et al. Sensation and Perception. 2nd ed. Sunderland, MA: Sinauer Associates; 2009.
29. Birch EE, Hoffman DR, Castaneda YS, Fawcett SL, Birch DG, Uauy RD. A randomized controlled trial of long-chain polyunsaturated fatty acid supplementation of formula in term infants after wean-ing at 6 wk of age. Am J Clin Nutr 2002;75:570–80.
30. Birch EE, Castaneda YS, Wheaton DH, Birch DG, Uauy RD, Hoffman DR. Visual maturation of term infants fed long-chain polyunsaturated fatty acid-supplemented or control formula for 12 mo. Am J Clin Nutr 2005;81:871–9.
31. Hoffman DR, Birch EE, Birch DG, Uauy R, Castaneda YS, Lapus MG, Wheaton DH. Impact of early dietary intake and blood lipid composition of long-chain polyunsaturated fatty acids on later visual development. J Pediatr Gastroenterol Nutr 2000;31: 540–53.
32. Birch EE, Carlson SE, Hoffman DR, Fitzgerald-Gustafson KM, Fu VL, Drover JR, et al. The DIAMOND (DHA Intake And Mea-surement Of Neural Development) Study: a double-masked, ran-domized controlled clinical trial of the maturation of infant visual acuity as a function of the dietary level of docosahexaenoic acid. Am J Clin Nutr 2010;91:848–59.
33. Auestad N, Scott DT, Janowsky JS, Jacobsen C, Carroll RE, Montalto MB, et al. Visual, cognitive, and language assessments at 39 months: a follow-up study of children fed formulas containing long-chain polyunsaturated fatty acids to 1 year of age. Pediatrics 2003;112(3 Pt 1):e177–83.
34. Carlson SE, Werkman SH, Rhodes PG, Tolley EA. Visual-acuity de-velopment in healthy preterm infants: effect of marine-oil supple-mentation. Am J Clin Nutr 1993;58:35–42.
35. Carlson SE, Werkman SH, Tolley EA. Effect of long-chain n-3 fatty acid supplementation on visual acuity and growth of preterm in-fants with and without bronchopulmonary dysplasia. Am J Clin Nutr 1996;63:687–97.
36. O’Connor DL, Hall R, Adamkin D, Auestad N, Castillo M, Connor WE, et al. Growth and development in preterm infants fed long-chain polyunsaturated fatty acids: a prospective, randomized controlled trial. Pediatrics 2001;108:359–71.
37. Morale SE, Hoffman DR, Castaneda YS, Wheaton DH, Burns RA, Birch EE. Duration of long-chain polyunsaturated fatty acids availability in the diet and visual acuity. Early Hum Dev 2005;81: 197–203.
38. Cheatham CL, Colombo J, Carlson SE. n-3 fatty acids and cognitive and visual acuity development: methodologic and conceptual considerations. Am J Clin Nutr 2006;83(Suppl. 6): 1458–1466S.
39. Jacobson JL, Jacobson SW, Muckle G, Kaplan-Estrin M, Ayotte P, Dewailly E. Beneficial effects of a polyunsaturated fatty acid on infant development: evidence from the Inuit of Arctic Quebec. J Pediatr 2008;152:356–64.
40. Innis SM, Friesen RW. Essential n-3 fatty acids in pregnant women and early visual acuity maturation in term infants. Am J Clin Nutr 2008;87:548–57.
41. Judge MP, Harel O, Lammi-Keefe CJ. A docosahexaenoic acid-functional food during pregnancy benefits infant visual acuity at four but not six months of age. Lipids 2007;42:117–22.
42. Malcolm CA, McCulloch DL, Montgomery C, Shepherd A, Weaver LT. Maternal docosahexaenoic acid supplementation during preg-nancy and visual evoked potential development in term infants: a double blind, prospective, randomised trial. Arch Dis Child Fetal Neonatal Ed 2003;88:F383–90.
43. Birch E, Birch D, Hoffman D, Hale L, Everett M, Uauy R. Breast-feeding and optimal visual development. J Pediatr Ophthalmol Stra-bismus 1993;30:33–8.
44. Williams C, Birch EE, Emmett PM, Northstone K. Stereoacuity at age 3.5 y in children born full-term is associated with prenatal and postnatal dietary factors: a report from a population-based cohort study. Am J Clin Nutr 2001;73:316–22.
45. Jacques C, Levy E, Muckle G, Jacobson SW, Bastien C, Dewailly E, et al. Long-term effects of prenatal omega-3 fatty acid intake on visual function in school-age children. J Pediatr 2011;158: 83–90, e1.
46. Lucas M, Dewailly E, Muckle G, Ayotte P, Bruneau S, Gingras S, et al. Gestational age and birth weight in relation to n-3 fatty acids among Inuit (Canada). Lipids 2004;39:617–26.
47. Muckle G, Ayotte P, Dewailly E, Jacobson SW, Jacobson JL. Deter-minants of polychlorinated biphenyls and methylmercury expo-sure in Inuit women of childbearing age. Environ Health Perspect 2001;109:957–63.
48. Shapley R. Visual sensitivity and parallel retinocortical channels. Annu Rev Psychol 1990;41:635–58.
49. Krauss-Etschmann S, Shadid R, Campoy C, Hoster E, Demmel-mair H, Jimenez M, et al. Effects of fish-oil and folate supple-mentation of pregnant women on maternal and fetal plasma concentrations of docosahexaenoic acid and eicosapentaenoic acid: a European randomized multicenter trial. Am J Clin Nutr 2007;85:1392–400.
50. Donahue SM, Rifas-Shiman SL, Olsen SF, Gold DR, Gillman MW, Oken E. Associations of maternal prenatal dietary intake of n-3 and n-6 fatty acids with maternal and umbilical cord blood levels. Pros-taglandins Leukot Essent Fatty Acids 2009;80:289–96.
51. Molloy C, Doyle LW, Makrides M, Anderson PJ. Docosahexaenoic acid and visual functioning in preterm infants: a review. Neuropsy-chol Rev 2012;22:425–37.
52. Jednorog K, Marchewka A, Tacikowski P, Heim S, Grabowska A. Electrophysiological evidence for the magnocellular–dorsal path-way deficit in dyslexia. Dev Sci 2011;14:873–80.
7. MACRON
nCEs 261
53. Richardson AJ, Calvin CM, Clisby C, Schoenheimer DR, Montgomery P, Hall JA, et al. Fatty acid deficiency signs predict the severity of reading and related difficulties in dyslexic children. Prostaglandins Leukot Essent Fatty Acids 2000;63:69–74.
54. Taylor KE, Higgins CJ, Calvin CM, Hall JA, Easton T, McDaid AM, Richardson AJ. Dyslexia in adults is associated with clinical signs of fatty acid deficiency. Prostaglandins Leukot Essent Fatty Acids 2000;63:75–8.
55. Lindmark L, Clough PA. 5-month open study with long-chain polyunsaturated fatty acids in dyslexia. J Med Food 2007;10:662–6.
56. Cyhlarova E, Bell JG, Dick JR, Mackinlay EE, Stein JF, Richardson AJ. Membrane fatty acids, reading and spelling in dyslexic and non-dyslexic adults. Eur Neuropsychopharmacol 2007;17:116–21.
57. Dunstan JA, Mitoulas LR, Dixon G, Doherty DA, Hartmann PE, Simmer K, Prescott SL. The effects of fish oil supplementation in pregnancy on breast milk fatty acid composition over the course of lactation: a randomized controlled trial. Pediatr Res 2007;62:689–94.
58. Dunstan JA, Simmer K, Dixon G, Prescott SL. Cognitive assess-ment of children at age 2(1/2) years after maternal fish oil supple-mentation in pregnancy: a randomised controlled trial. Arch Dis Child Fetal Neonatal Ed 2008;93:F45–50.
59. Carlson SE, Werkman SH. A randomized trial of visual attention of preterm infants fed docosahexaenoic acid until two months. Lipids 1996;31:85–90.
60. Reisbick S, Neuringer M, Gohl E, Wald R, Anderson GJ. Visual at-tention in infant monkeys: effects of dietary fatty acids and age. Dev Psychol 1997;33:387–95.
61. Werkman SH, Carlson SE. A randomized trial of visual attention of preterm infants fed docosahexaenoic acid until nine months. Lipids 1996;31:91–7.
62. Colombo J, Kannass KN, Shaddy DJ, Kundurthi S, Maikranz JM, Anderson CJ, et al. Maternal DHA and the development of atten-tion in infancy and toddlerhood. Child Dev 2004;75:1254–67.
63. Kannass KN, Colombo J, Carlson SE. Maternal DHA levels and toddler free-play attention. Dev Neuropsychol 2009;34:159–74.
64. Makrides M, Neumann MA, Byard RW, Simmer K, Gibson RA. Fatty acid composition of brain, retina, and erythrocytes in breast- and formula-fed infants. Am J Clin Nutr 1994;60:189–94.
65. Braddick O, Atkinson J, Wattam-Bell J. Normal and anomalous development of visual motion processing: motion coherence and ‘dorsal-stream vulnerability’. Neuropsychologia 2003;41:1769–84.