Proc. Natl. Acad. Sci. USA Vol. 77, No. 5, pp. 2465-2469, May 1980 Biochemistry Heterogeneity of apolipoprotein B: Isolation of a new species from human chylomicrons (lipoproteins/protein moiety/lipid transport) JOHN P. KANE*t, DAVID A. HARDMANt, AND HAROLD E. PAULUSt *Department of Medicine and tCardiovascular Research Institute, University of California, San Francisco, California 94143; and *Department of Medicine, University of California, Los Angeles, and Rehabilitation Center, 1000 Veteran Avenue, Los Angeles, California 90024 Communicated by John A. Clements, January 24,1980 ABSTRACT Low density lipoproteins and the triglycer- ide-rich lipoproteins of human serum each contain proteins of high molecular weight termed apolipoprotein B,which have previously been thought to be identical. We have isolated four species of apolipoprotein B with unique molecular weights and amino acid compositions. We have assigned numerical desig- nations to these species in a centile system based upon their relative apparent Mr in NaDodSO4. One which we term B-100, with an apparent Mr of 549,000 ± 7650 (SD) determined by NaDodSO4 gel electrophoresis, predominates in low density and very low density lipoproteins and is also present in chylomicrons from thoracic duct lymph or from plasma. Substantial amounts of two large proteins designated B-74 (apparent Mr 407,000 ± 5790) and B-26 (apparent Mr 144,500 + 8970), which appear to be complementary fragments or constituents of the B-100 pro- tein, are found in the low density lipoproteins of many indi- viduals. A distinct protein, B-48, with an apparent Mr of 264,000 i 8150 is a major and constant constituent of chylomicrons from thoracic duct lymph or from plasma. The lipoproteins of human serum contain at least 12 species of proteins' (apolipoproteins) (1). The most abundant of these, apolipoprotein B (apo B) shares with another species, apolipo- protein E, the ability to interact with high-affinity receptors on cell membranes (2). apo B is distinguished by its apparent inability to transfer among lipoprotein particles, by its extreme insolubility in aqueous buffers after the lipid moiety is removed, and by its insolubility in 4.2 M tetramethylurea (3). Studies of apo B solubilized with various denaturing solvents and am- phiphilic agents have yielded widely varying estimates of molecular weight (1, 4-7). We describe here the presence of four major molecular species of apo B. One of these is nearly the sole form in very low density lipoproteins (VLDL). It also appears in low density li- poproteins (LDL) and chylomicrons from thoracic duct lymph and plasma. Two others appear to be fragments or components of the former and appear only in LDL. The fourth, distin- guished by both its molecular weight and amino acid compo- sition, is found only in chylomicrons. MATERIALS AND METHODS Preparation of Lipoproteins. Serum was prepared from blood obtained from donors who had fasted for 12-16 hr. Li- poproteins of serum were separated by repetitive preparative ultracentrifugation in a 40.3 rotor, at 12°C, in a Beckman model L ultracentrifuge (8). VLDL were separated in the p < 1.006 g/cm3 fraction of serum from normolipidemic individuals after centrifugation for 18 hr. The ultracentrifugation was repeated once. Chylomicrons were judged to be absent from preparations of VLDL by the absence of a visible supernatant layer after refrigeration for 24 hr at 40C and by the absence of lipoproteins The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "ad- vertisement" in accordance with 18 U. S. C. §1734 solely to indicate this fact. 2465 in the application well after electrophoresis in agarose gel (9). Chylomicrons were prepared from the serum of individuals with mixed lipemia by centrifugation at 20,000 rpm in the 40.3 rotor for 20 min. Three milliliters of serum was layered under 0.15 M NaCl in each tube. Chylomicrons were recovered in the top 1.5 ml. The centrifugation was repeated once. LDL were isolated in parallel with VLDL. The p < 1.024 g/cm3 fraction was removed in the first ultracentrifugation (22 hr). The solvent density was then increased to 1.050 g/cm3. The supernatant fraction was removed after 22 hr. This solution was layered under 2 vol of KBr solution, 1.050 g/cm3, and recentrifuged for 22 hr to purify the LDL. Thoracic duct lymph was obtained from five normolipidemic patients undergoing lymphophoresis as an investigative inter- vention for rheumatoid arthritis and other presumed autoim- mune disorders. Chylomicrons were separated from lymph by centrifugation twice at p = 1.006 g/cm3 for 22 hr. Measures to Prevent Nonspecific Degradation of apo B. Gentamycin sulfate, 0.1 mg/ml (gift of Schering, Kenilworth, NJ), and nonesterified chloramphenicol, 0.05 mg/ml (gift of Parke, Davis), were added immediately to serum or lymph and were present in all buffers and ultracentrifugal media to pre- vent microbial degradation of apo B (10). Sodium azide (0.02%) was present in all solutions except the running gel buffer in the electrophoresis apparatus (azide inhibits polymerization of polyacrylamide). EDTA (1 mM, pH 8.6) was added to all so- lutions to prevent hydroperoxidative degradation of lipopro- teins. Preparation of lipoproteins was begun immediately upon obtaining serum or lymph and was completed without delay. The LDL were dialyzed against 0.15 M NaCl containing anti- biotics, azide, and EDTA for 22 hr at 40C before delipida- tion. Delipidation. For NaDodSO4 electrophoresis, all lipoproteins were delipidated by extraction twice with 20 vol of ethanol/ diethyl ether, 3:1 (vol/vol), for 20 hr. For preparative electro- phoresis of apolipoproteins from triglyceride-rich lipoproteins, the precipitate obtained as above was dispersed and extracted with 4.2 M tetramethylurea at 30°C (3) to remove proteins other than apo B. Analytical NaDodSO4 Gel Electrophoresis. Protein was dissolved in 50 mM sodium phosphate (pH 7.2) containing NaDodSO4 (10 mg/mg of protein) and 1% 2-mercaptoethanol (both from Pierce). An equal volume of 30% sucrose was added to the solution to facilitate layering on the gels. Electrophoresis was performed in 3% polyacrylamide gels by the method of Weber and Osborn (11). Samples containing 20 ,ug of apo B were applied to the gels. Gels were fixed in ethanol/water/ acetic acid, 9:9:2 by vol, for 2 hr, stained in 0.0025% Coomassie blue (Schwarz/Mann) in 10% acetic acid for 48 hr, and de- stained in 10% acetic acid. Photodensitometric scanning of the Abbreviations: LDL, low density lipoproteins; VLDL, very low density lipoproteins; apo B, apolipoprotein B.
Transcript
Proc. Natl. Acad. Sci. USAVol. 77, No. 5, pp. 2465-2469, May 1980Biochemistry
Heterogeneity of apolipoprotein B: Isolation of a new species fromhuman chylomicrons
(lipoproteins/protein moiety/lipid transport)
JOHN P. KANE*t, DAVID A. HARDMANt, AND HAROLD E. PAULUSt*Department of Medicine and tCardiovascular Research Institute, University of California, San Francisco, California 94143; and *Department of Medicine,University of California, Los Angeles, and Rehabilitation Center, 1000 Veteran Avenue, Los Angeles, California 90024
Communicated by John A. Clements, January 24,1980
ABSTRACT Low density lipoproteins and the triglycer-ide-rich lipoproteins of human serum each contain proteins ofhigh molecular weight termed apolipoprotein B,which havepreviously been thought to be identical. We have isolated fourspecies of apolipoprotein B with unique molecular weights andamino acid compositions. We have assigned numerical desig-nations to these species in a centile system based upon theirrelative apparent Mr in NaDodSO4. One which we term B-100,with an apparent Mr of 549,000 ± 7650 (SD) determined byNaDodSO4 gel electrophoresis, predominates in low density andvery low density lipoproteins and is also present in chylomicronsfrom thoracic duct lymph or from plasma. Substantial amountsof two large proteins designated B-74 (apparent Mr 407,000 ±5790) and B-26 (apparent Mr 144,500 + 8970), which appear tobe complementary fragments or constituents of the B-100 pro-tein, are found in the low density lipoproteins of many indi-viduals. A distinct protein, B-48, with an apparent Mr of 264,000i 8150 is a major and constant constituent of chylomicrons fromthoracic duct lymph or from plasma.
The lipoproteins of human serum contain at least 12 species ofproteins' (apolipoproteins) (1). The most abundant of these,apolipoprotein B (apo B) shares with another species, apolipo-protein E, the ability to interact with high-affinity receptorson cell membranes (2). apo B is distinguished by its apparentinability to transfer among lipoprotein particles, by its extremeinsolubility in aqueous buffers after the lipid moiety is removed,and by its insolubility in 4.2 M tetramethylurea (3). Studies ofapo B solubilized with various denaturing solvents and am-phiphilic agents have yielded widely varying estimates ofmolecular weight (1, 4-7).We describe here the presence of four major molecular
species of apo B. One of these is nearly the sole form in very lowdensity lipoproteins (VLDL). It also appears in low density li-poproteins (LDL) and chylomicrons from thoracic duct lymphand plasma. Two others appear to be fragments or componentsof the former and appear only in LDL. The fourth, distin-guished by both its molecular weight and amino acid compo-sition, is found only in chylomicrons.
MATERIALS AND METHODSPreparation of Lipoproteins. Serum was prepared from
blood obtained from donors who had fasted for 12-16 hr. Li-poproteins of serum were separated by repetitive preparativeultracentrifugation in a 40.3 rotor, at 12°C, in a Beckman modelL ultracentrifuge (8). VLDL were separated in the p < 1.006g/cm3 fraction of serum from normolipidemic individuals aftercentrifugation for 18 hr. The ultracentrifugation was repeatedonce. Chylomicrons were judged to be absent from preparationsof VLDL by the absence of a visible supernatant layer afterrefrigeration for 24 hr at 40C and by the absence of lipoproteins
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.
2465
in the application well after electrophoresis in agarose gel (9).Chylomicrons were prepared from the serum of individualswith mixed lipemia by centrifugation at 20,000 rpm in the 40.3rotor for 20 min. Three milliliters of serum was layered under0.15 M NaCl in each tube. Chylomicrons were recovered in thetop 1.5 ml. The centrifugation was repeated once. LDL wereisolated in parallel with VLDL. The p < 1.024 g/cm3 fractionwas removed in the first ultracentrifugation (22 hr). The solventdensity was then increased to 1.050 g/cm3. The supernatantfraction was removed after 22 hr. This solution was layeredunder 2 vol of KBr solution, 1.050 g/cm3, and recentrifugedfor 22 hr to purify the LDL.
Thoracic duct lymph was obtained from five normolipidemicpatients undergoing lymphophoresis as an investigative inter-vention for rheumatoid arthritis and other presumed autoim-mune disorders. Chylomicrons were separated from lymph bycentrifugation twice at p = 1.006 g/cm3 for 22 hr.
Measures to Prevent Nonspecific Degradation of apo B.Gentamycin sulfate, 0.1 mg/ml (gift of Schering, Kenilworth,NJ), and nonesterified chloramphenicol, 0.05 mg/ml (gift ofParke, Davis), were added immediately to serum or lymph andwere present in all buffers and ultracentrifugal media to pre-vent microbial degradation of apo B (10). Sodium azide (0.02%)was present in all solutions except the running gel buffer in theelectrophoresis apparatus (azide inhibits polymerization ofpolyacrylamide). EDTA (1 mM, pH 8.6) was added to all so-lutions to prevent hydroperoxidative degradation of lipopro-teins. Preparation of lipoproteins was begun immediately uponobtaining serum or lymph and was completed without delay.The LDL were dialyzed against 0.15 M NaCl containing anti-biotics, azide, and EDTA for 22 hr at 40C before delipida-tion.
Delipidation. For NaDodSO4 electrophoresis, all lipoproteinswere delipidated by extraction twice with 20 vol of ethanol/diethyl ether, 3:1 (vol/vol), for 20 hr. For preparative electro-phoresis of apolipoproteins from triglyceride-rich lipoproteins,the precipitate obtained as above was dispersed and extractedwith 4.2 M tetramethylurea at 30°C (3) to remove proteinsother than apo B.
Analytical NaDodSO4 Gel Electrophoresis. Protein wasdissolved in 50 mM sodium phosphate (pH 7.2) containingNaDodSO4 (10 mg/mg of protein) and 1% 2-mercaptoethanol(both from Pierce). An equal volume of 30% sucrose was addedto the solution to facilitate layering on the gels. Electrophoresiswas performed in 3% polyacrylamide gels by the method ofWeber and Osborn (11). Samples containing 20 ,ug of apo Bwere applied to the gels. Gels were fixed in ethanol/water/acetic acid, 9:9:2 by vol, for 2 hr, stained in 0.0025% Coomassieblue (Schwarz/Mann) in 10% acetic acid for 48 hr, and de-stained in 10% acetic acid. Photodensitometric scanning of the
Abbreviations: LDL, low density lipoproteins; VLDL, very low densitylipoproteins; apo B, apolipoprotein B.
Proc. Natl. Acad. Sci. USA 77 (1980)
_rn, ;viLJLJ
A B C D E F GFIG. 1. Comparative analytical NaDodSO4 gel electrophoresis
of apoproteins of serum and lymph. Gel A, lymph chylomicrons; gelB, VLDL; gel C, LDL; gel D, LDL from another donor; gels E, F, andG, reelectrophoresis of bands from donor D LDL. Lower bands in gelsA and B contain proline-rich apolipoprotein and apolipoproteins E,A, I, and C.
gels was performed at 602 nm with a scanning spectropho-tometer (model 740, Coming Medical Division, Medfield,MA).
Isolation of Components of apo B by Preparative Na-DodSO4 Gel Electrophoresis. Samples were prepared as foranalytical NaDodSO4 electrophoresis. Separation was accom-plished with a modified Shandon Southern preparative gelapparatus, MK II (Shandon Southern Instruments, Sewickley,PA).§ The buffer in the gel and in both tanks was 0.1 M sodiumphosphate (pH 7.2) with 0.1% NaDodSO4. Gentamycin sulfate(0.1 mg/ml) and sodium azide (0.02%) were added to the tankbuffer. The separating gel, 10 cm long, contained 2.3% poly-acrylamide. Separation of samples containing up to 4 mg of apoB was carried out at 25°C and 95 mA. Protein peaks were de-tected by absorbance at 280 nm with a flow-cell spectropho-tometer. Pooled samples of eluate taken from narrow intervalswithin each peak were lyophilized in 7.5 X 100mm glass tubesthat had been washed with chromic acid, rinsed repeatedly withglass-redistilled water, and then rinsed with 1% EDTA to re-move traces of chromate. The residue was extracted twice with2 ml of 20% trichloroacetic acid to remove NaDodSO4 andbuffer salts. After centrifugation (15,000 X g-min), the tri-chloroacetic acid was removed. The precipitate was washedtwice with acetone and dried under nitrogen.Amino Acid Aiialysis. Precipitates of apo B were hydrolyzed
at 105°C, under vacuum, with 1 ml of 6M HC1 (Ultrex, Baker,Phillipsburg, NJ), containing 0.05 mg of phenol. Portions ofeach sample were hydrolyzed for 22, 72, and 120 hr and ly-ophilized. The residues were dissolved in 0.2 M sodium citrate(pH 2.2) for analysis in triplicate with a two-column programon a Beckman model 121 M amino acid analyzer (13). Valuesfor threonine and serine were extrapolated to zero time, andvalues at 120 hr were used for the content of valine, isoleucine,
§ Modifications of instrument and technique are described in ref.12.
and leucine. Cysteine and methionine were measured by thetechnique of Hirs (14).
RESULTSWe have observed that the mobility of components of apo Bupon analytical NaDodSO4 gel electrophoresis is dependentupon the amount applied, perhaps accounting for some of thedisparate estimates of molecular weight that have been re-ported. Mobilities appear to become constant at minimal valueswhen 20 ,g or less is applied.On analytical NaDodSO4 gel electrophoresis, the apo B from
LDL of approximately half of our normal donors appeared asa single band corresponding to a Mr of 549,000 + 7650 (meanand SD of preparations from three separate donors) (Fig. 1, gelC). The LDL apo B of other apparently normal individualscontained three bands, one of which corresponded to the singleelement of Mr 549,000 described above and the other two ofMr 407,000 i 5790 and 144,500 + 8970 (mean and SD forpreparations from three separate donors) (Fig. 1, gel D). Thesethree elements are distinct from the numerous bands with Mrbetween 100,000 and 500,000, which appear with rigoroustreatment of apo B with denaturing solvents, with certain am-phiphiles, or upon protracted storage (4). They differ from thosebands by their unique and fixed apparent Mr and by the factsthat they are present in apo B from freshly prepared LDL andare unaltered in amount and mobilityby storage of either LDLor apo B. The apo B from LDL of different individuals maycontain only a trace of these additional bands or they maycomprise up to a third of the total mass. However, when the twosmaller elements are present, their relative absorbances suggestthat they are in equimolar amounts. To test this apparent stoi-chiometric relationship, we measured total recoveries of ami-noacyl mass in these peaks from two separate preparations ofLDL. We found relative masses of 73.8:26.2 and 75.0:25.0,indicating a precise 1:1 molar stoichiometry. Additional veryfaint bands might be seen in freshly prepared apo B (Fig. 1, gelD). One of these, which was not always present, had a mobility
1 ,000,00°r
0
c
0._
co00
100,000 -
B-100
B-74
B-48Myo~inHuman transferrin 2 x
B-26 BSA 2 x
Ovalbumin 3 xOvalbumin 2 xHuman transferrinBSA
Ovalbumin
1 u,uuu.I I I I I I0 0.2 0.4 0.6 0.8 1.0
RF (relative to NaDodSO4 micelles)FIG. 2. Comparison of electrophoretic mobilities of components
of apo B with protein standards in the preparative NaDodSO4 gelsystem (2.3% polyacrylamide/0.1 M phosphate buffer, pH 7.2/0.1%NaDodSO4). Proteins denoted 2x and 3X are putative dimers andtrimers; BSA is bovine, serum albumin. See subsection ProposedNomenclature in Results.
2466 Biochemistry: Kane et al.
--mmommom
S.I..- 11
Proc. Natl. Acad. Sc. USA 77 (1980) 2467
Table 1. Amino acid compositions of major apo B components of LDL (1.024 < p < 1.050 g/cm3)*Weighted
Amino sum, LDLacid B-74 B-26 B-74 and B-26 B-100 SEM
* Residues per 103 residues. Each value is the mean of data on three preparations, each of which rep-resents triplicate analyses at each of three different times of hydrolysis (22,72, and 120 hr)-i.e., eachvalue represents 27 determinations of amino acid composition. SEM, derived from pooled estimateof variance of all analyses, is provided as an index of precision of amino acid analysis.
t Denotes those amino acids for which there is no overlap in range between the B-74, B-26, and B-100proteins.
corresponding to that of the smaller species (apparent Mr264,000) of apo B seen in chylomicrons; however, its amino acidcomposition showed it was not the same protein (see below).After separation by preparative gel electrophoresis, all thesecomponents of LDL apo B retained their respective electro-phoretic mobilities (Fig. 1, gels E, F, and G). Photometric scansof the stained gels containing the isolated bands showed thatover 95% of the absorbance was in the primary bands.
Analytical electrophoretograms of human lymph chylomi-crons and serum chylomicrons contained a band with mobilityidentical to the large component found in apo B from LDL.However, they also contained a band corresponding to an ap-parent Mr of 264,000 + 8150 (mean and SD of values forpreparations from three individual donors) which was absentfrom apo B of LDL. As noted above, a trace of an element witha closely similar mobility was seen in apo B from some LDL,but its amino acid composition differed from the small elementfrom lymph chylomicrons. The overwhelmingly predominantelement of apo B in VLDL had a mobility corresponding to thelarge element in apo B from LDL. In addition to very faintminor bands which did not correspond to the three major bandsof apo B from LDL, some samples of VLDL appeared to con-tain a minor band with mobility corresponding to that of thesmall element seen in lymph chylomicrons. Calculated mo-lecular weights for the major elements of apo B, based on timeof elution from the preparative gel, were in each case within5% of the values obtained from relative mobilities in the ana-lytical system (Fig. 2).Proposed Nomenclature. Because of the wide range of ap-
parent Mr reported from different laboratories for apo B andthe heterogeneity described above, we propose a centile systemof nomenclature which uses the largest element of LDL apo Bas an internal reference standard in NaDodSO4 gel electro-phoresis. We assign the arbitrary value of 100 to this protein andexpress the apparent Mr of all other elements in relation toit-i.e., the three major components of LDL apo B becomeB-100, B-74, and B-26, and the major components of chylo-micron apo B become B-100 and B-48 (Fig. 2). This system of
nomenclature would be expected to normalize the effects ofmany of the differences in analytical technique among differentlaboratories and would allow the direct comparison of anymolecules derived from apo B by disaggregation, with the ap-parently naturally occurring forms of apo B describedabove.
Characterization of Major Elements of apo B. The aminoacid compositions of the B-100, B-74, and B-26 components ofLDL are compared in Table 1. We found consistent differencesin the content of histidine, arginine, aspartic acid, glutamic acid,proline, glycine, alanine, half-cystine, methionine, isoleucine,and phenylalanine between the B-74 and B-26 elements. Theamino acid compositions of both differed consistently from thatof the B-100 component. Because the sum of the apparent Mrof the B-74 and B-26 components equals that of the B-100protein (within the limits of resolution of the analytical tech-nique), it appeared that they might be derived from the B-100protein or, conversely, that they reflect incomplete assemblyof subunits of that protein. To test the relationship of B-74 andB-26 to the B-100 protein, we calculated a mass-weighted aminoacid composition for the combination of one particle each ofB-74 and B-26 by using their apparent Mr determined by Na-DodSO4 gel electrophoresis. The weighted composition ap-proaches that of B-100 protein as closely as replicate samplesof B-100 (Table 1).Major Components of apo B from Lymph Chylomicrons.
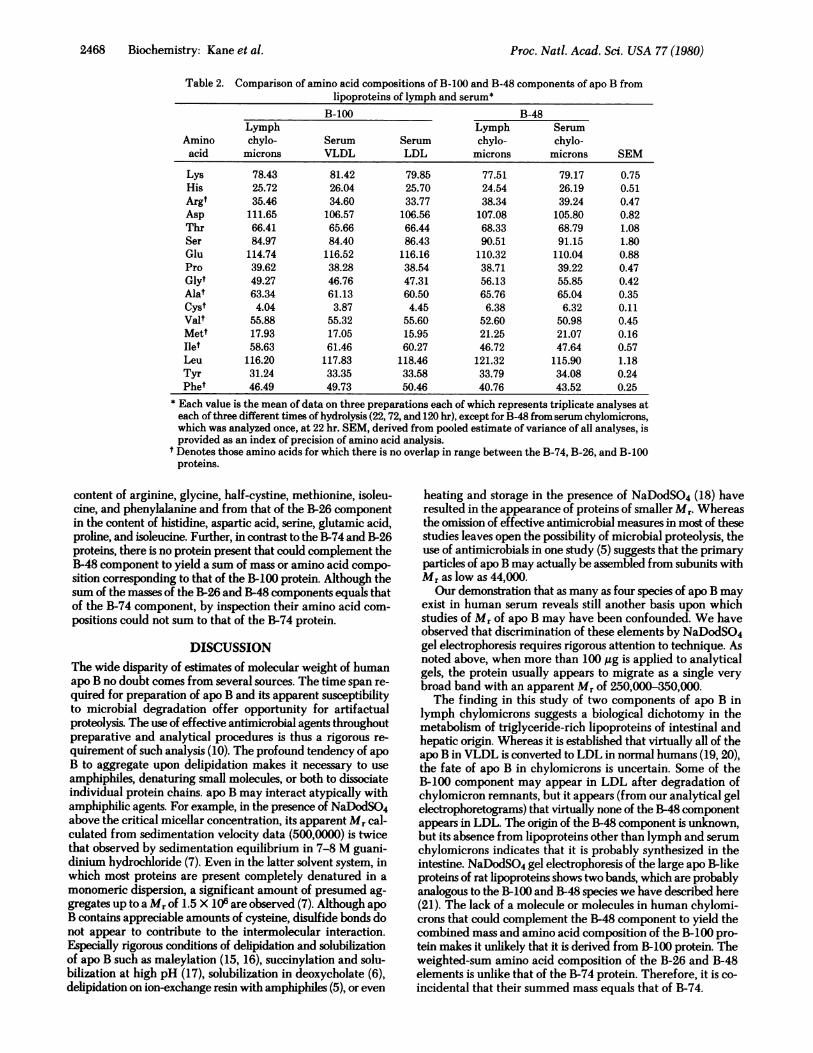
In Table 2 the amino acid compositions of the B-100 and B-48components of human lymph chylomicrons are compared withthe B-100 component of serum lipoproteins and with a singleanalysis of the B-48 component of serum chylomicrons. Dif-ferences in all amino acids between the various B-100 compo-nents, whether from lymph or serum lipoproteins, are nogreater than those observed in replicate analyses of any of theB-100 proteins, suggesting that the B-100 components of all thelipoproteins are identical. By contrast, the B-48 protein differsfrom the B-100 component in its content of glycine, half-cys-tine, methionine, isoleucine, and phenylalanine. Its amino acidcomposition also differs from that of the B-74 protein in its
Biochemistry: Kane et al.
Proc. Natl. Acad. Sci. USA 77 (1980)
Table 2. Comparison of amino acid compositions of B-100 and B-48 components of apo B fromlipoproteins of lymph and serum*
* Each value is the mean of data on three preparations each of which represents triplicate analyses ateach of three different times of hydrolysis (22,72, and 120 hr), except for B-48 from serum chylomicrons,which was analyzed once, at 22 hr. SEM, derived from pooled estimate of variance of all analyses, isprovided as an index of precision of amino acid analysis.
t Denotes those amino acids for which there is no overlap in range between the B-74, B-26, and B-100proteins.
content of arginine, glycine, half-cystine, methionine, isoleu-cine, and phenylalanine and from that of the B-26 componentin the content of histidine, aspartic acid, serine, glutamic acid,proline, and isoleucine. Further, in contrast to the B-74 and B-26proteins, there is no protein present that could complement theB-48 component to yield a sum of mass or amino acid compo-sition corresponding to that of the B-100 protein. Although thesum of the masses of the B-26 and B-48 components equals thatof the B-74 component, by inspection their amino acid com-positions could not sum to that of the B-74 protein.
DISCUSSIONThe wide disparity of estimates of molecular weight of humanapo B no doubt comes from several sources. The time span re-quired for preparation of apo B and its apparent susceptibilityto microbial degradation offer opportunity for artifactualproteolysis. The use of effective antimicrobial agents throughoutpreparative and analytical procedures is thus a rigorous re-quirement of such analysis (10). The profound tendency of apoB to aggregate upon delipidation makes it necessary to useamphiphiles, denaturing small molecules, or both to dissociateindividual protein chains. apo B may interact atypically withamphiphilic agents. For example, in the presence of NaDodSO4above the critical micellar concentration, its apparent Mr cal-culated from sedimentation velocity data (500,0000) is twicethat observed by sedimentation equilibrium in 7-8 M guani-dinium hydrochloride (7). Even in the latter solvent system, inwhich most proteins are present completely denatured in amonomeric dispersion, a significant amount of presumed ag-gregates up to a Mr of 1.5 X 106 are observed (7). Although apoB contains appreciable amounts of cysteine, disulfide bonds donot appear to contribute to the intermolecular interaction.Especially rigorous conditions of delipidation and solubilizationof apo B such as maleylation (15, 16), succinylation and solu-bilization at high pH (17), solubilization in deoxycholate (6),delipidation on ion-exchange resin with amphiphiles (5), or even
heating and storage in the presence of NaDodSO4 (18) haveresulted in the appearance of proteins of smaller Mr. Whereasthe omission of effective antimicrobial measures in most of thesestudies leaves open the possibility of microbial proteolysis, theuse of antimicrobials in one study (5) suggests that the primaryparticles of apo B may actually be assembled from subunits withMr as low as 44,000.Our demonstration that as many as four species of apo B may
exist in human serum reveals still another basis upon whichstudies of Mr of apo B may have been confounded. We haveobserved that discrimination of these elements by NaDodSO4gel electrophoresis requires rigorous attention to technique. Asnoted above, when more than 100 ,ug is applied to analyticalgels, the protein usually appears to migrate as a single verybroad band with an apparent Mr of 250,000-350,000.The finding in this study of two components of apo B in
lymph chylomicrons suggests a biological dichotomy in themetabolism of triglyceride-rich lipoproteins of intestinal andhepatic origin. Whereas it is established that virtually all of theapo B in VLDL is converted to LDL in normal humans (19,20),the fate of apo B in chylomicrons is uncertain. Some of theB-100 component may appear in LDL after degradation ofchylomicron remnants, but it appears (from our analytical gelelectrophoretograms) that virtually none of the B-48 componentappears in LDL. The origin of the B-48 component is unknown,but its absence from lipoproteins other than lymph and serumchylomicrons indicates that it is probably synthesized in theintestine. NaDodSO4 gel electrophoresis of the large apo B-likeproteins of rat lipoproteins shows two bands, which are probablyanalogous to the B-100 and B-48 species we have described here(21). The lack of a molecule or molecules in human chylomi-crons that could complement the B-48 component to yield thecombined mass and amino acid composition of the B-100 pro-tein makes it unlikely that it is derived from B-100 protein. Theweighted-sum amino acid composition of the B-26 and B-48elements is unlike that of the B-74 protein. Therefore, it is co-incidental that their summed mass equals that of B-74.
2468 Biochemistry: Kane et al.
Proc. Natl. Acad. Sci. USA 77 (1980) 2469
By contrast, the B-74 and B-26 proteins found in LDL are
precisely complementary. Their combined mass and *wightedamino acid composition are indistinguishable from those of theB-100 protein. The complementarity of B-74 and B-26 is un-derscored by our observation that they appear to be present inequimolar amounts. Whether they represent products ofcleavage of B-100 or incomplete assembly of that protein fromsmaller subunits awaits further analysis of their structure.However, if all LDL apo B does originate in VLDL, the absenceof B-74 and B-26 from VLDL favors the former. If B-100 is adimer in NaDodSO4 (7), then B-74 and B-26 may also be di-mers. The lack of correspondence of B-74 and B-26 to the bandsthat appear after rigorous treatment of apo B, such as boilingin NaDodSO4, suggests that they are not produced by similarmechanisms of dissociation. Their presence in LDL preparedfreshly in the presence of diisopropylfluorophosphate (10) andthe lack of change in these elements with storage stronglysuggest that they exist in circulating LDL. Also, their absencefrom all samples of VLDL that we have tested, including manyfrom individuals with three-component apo B in LDL, suggeststhat this is not an artifact of cleavage of B-100 in vitro, but thatB-74 and B-26 are formed from B-100 of LDL by some bio-logically mediated process in some individuals. This phenom-enon is observed in both males and females and it tends to bepresent in an individual's LDL for a period of at least severalmonths.The amino acid compositions of the four elements described
above permit certain conclusions about the structures of thoseproteins. If each is a single polypeptide chain, then the B-48protein of chylomicrons is distinct from the B-100 element. Itis probable that B-100 represents one protein found in VLDL,LDL, and chylomicrons. If B-48 and B-100 are assembled fromsmaller subunits, then the constituent monomers differ in eitherspecies or molar ratios in the two proteins. If B-100 is a singlechain, then the amino acid compositions of B-74 and B-26 are
compatible with production by cleavage at a single site. Thediscreteness and uniformity of the B-74 and B-26 proteinssuggest a single mechanism of cleavage. If B-100 is assembledfrom subunits, the amino acid compositions of B-74 and B-26indicate that more than one species of monomer must bepresent. Analysis of purified components of apo B thus becomes
an important step in the investigation of the subunit structureof these proteins.
This research was supported by SCOR Grant HL 14237 from theNational Heart Lung and Blood Institute and by the Dorothy andThomas Leavey Trust Foundation. J.P.K. was an Established Investi-gator of the American Heart Association during part of this work.
1. Kane, J. P. (1977) in Lipid Metabolism in Mammals, ed. Snyder,F. (Plenum, New York), pp. 209-257.
2. Innerarity, T. L. & Mahley, R. W. (1978) Biochemistry 17,1440-1447.
3. Kane, J. P., Sata, T., Hamilton, R. L. & Havel, R. J. (1975) J. CGn.Invest. 58, 1622-1634.
4. Morrisett, J. D., Jackson, R. L. & Gotto, A. M., Jr. (1975) Annu.Rev. Biochem. 44,183-207.
5. Socorro, L. & Camejo, G. (1979) J. Lipid Res. 20, 631-638.6. Chen, C-H. & Aladjem, F. (1978) Biochem. Med. 19, 178-
187.7. Steele, J. C. H., Jr. & Reynolds, J. A. (1979) J. Biol. Chem. 254,
1639-1643.8. Havel, R. J., Eder, H. A. & Bragdon, J. H. (1955) J. Clin. Invest.
34,1345-1353.9. Noble, R. P. (1968) J. Lipid Res. 9,693-700.
10. Chapman, M. J. & Kane, J. P. (1975) Biochem. Biophys. Res.Commun. 66, 1030-1036.
11. Weber, K. & Osborn, M. (1969) J. Biol. Chem. 244, 4406-4412.
12. Hardman, D. A. & Kane, J. P. (1980) Anal. Biochem., in press.13. Spackman, D. H. (1967) Methods Enzymol. 11, 3-15.14. Hirs, C. H. W. (1967) Methods Enzymol. 11, 197-199.15. Kane, J. P., Richards, E. G. & Havel, R. J. (1970) Proc. Nati. Acad.
Sci. USA 66, 1075-1082.16. Shore, V. & Shore, B. (1972) in Blood Lipids and Lipoproteins,
Quantitation, Composition and Metabolism, ed. Nelson, G. J.(Wiley, New York), pp. 789824.
17. Pollard, H., Scanu, A. M. & Taylor, E. W. (1969) Proc. Natl. Acad.Sci. USA 64,304-310.
18. Steele, J. C. H., Jr. & Reynolds, J. A. (1979) J. Biol. Chem. 254,1633-1638.
19. Bilheimer, D. W., Eisenberg, S. & Levy, R. I. (1972) Biochim.Biophys. Acta 260,212-221.
20. Sigurdsson, G., Nicoll, A. & Lewis, B. (1975) J. Clin. Invest. 56,1481-1490.
21. Krishnaiah, K. V., Ong, J., Walker, L. F., Orensztajn, J. & Getz,G. S. (1978) Fed. Proc. Fed. Am. Soc. Exp. Biol. 37,257.