ORIGINAL ARTICLE High expression of GCLC is associated with malignant melanoma of low oxidative phenotype and predicts a better prognosis Dimitrios Mougiakakos & Riki Okita & Takashi Ando & Christoph Dürr & Jules Gadiot & Jiro Ichikawa & Robert Zeiser & Christian Blank & C. Christian Johansson & Rolf Kiessling Received: 7 September 2011 / Revised: 20 December 2011 / Accepted: 2 January 2012 / Published online: 17 January 2012 # Springer-Verlag 2012 Abstract Reactive oxygen species (ROS) are strongly im- plicated in melanoma development, and treatment with anti- oxidants has shown efficacy in suppressing malignant transition and progression. Here, we investigated the signif- icance of the glutamate-L-cysteine ligase catalytic subunit (GCLC) expression, a key regulator of glutathione synthe- sis, for malignant melanoma. A large set of melanoma cell lines (n 0 36) was analyzed, and higher GCLC levels were associated with lower presence of intracellular ROS and interestingly also lower rates of cell proliferation. Moreover, treatment with the antioxidant N-acetylcysteine efficiently reduced the growth speed of several investigated malignant cells. In addition GCLC expression was significantly linked to a prominent set of cellular antioxidants, accounting for the observed lower basal levels of oxidative stress and higher antioxidative capacity. Key attributes defining the malignant phenotype of melanoma cells including survival, invasiveness, and switch from E-cadherin to N-cadherin expression were more prominent in cells with lower GCLC expression. Our findings were further corroborated by observations in Rag2 −/− γc −/− mice, in which melanoma cells with lower GCLC expression depicted a dramatically stronger tumor growth. Furthermore, prognostic significance of GCLC expression was investigated in patients (n 0 28) with advanced malignant melanoma. High tumor immunoreactivity for GCLC was a significant determinant for better 5-year overall survival. Conclusively, we show for the first time that GCLC may serve a dual role, as a surrogate marker for cellular redox Dimitrios Mougiakakos, Riki Okita, C. Christian Johansson, and Rolf Kiessling contributed equally to this work. Electronic supplementary material The online version of this article (doi:10.1007/s00109-012-0857-4) contains supplementary material, which is available to authorized users. D. Mougiakakos : R. Okita : C. C. Johansson : R. Kiessling Department of Oncology and Pathology, Cancer Center Karolinska, Karolinska Institutet, Stockholm, Sweden D. Mougiakakos (*) Department of Internal Medicine 5, Hematology and Oncology, University of Erlangen, Glückstrasse 4a, Erlangen, Germany e-mail: [email protected]T. Ando Department of Orthopaedic Surgery, University of Yamanashi, Yamanashi 409-3898, Japan C. Dürr : R. Zeiser Division of Hematology and Oncology, Department of Medicine, Freiburg University Medical Center, Albert-Ludwigs-University, Freiburg, Germany J. Gadiot : C. Blank Department of Medical Oncology and Division of Immunology, The Netherlands Cancer Institute-Antoni van Leeuwenhoek Hospital (NKI-AVL), Amsterdam 1066 CX, The Netherlands J. Ichikawa Department of Orthopaedics and Pharmacology, Vanderbilt University, Nashville, TN, USA J Mol Med (2012) 90:935–944 DOI 10.1007/s00109-012-0857-4
Transcript
ORIGINAL ARTICLE
High expression of GCLC is associated with malignantmelanoma of low oxidative phenotype and predictsa better prognosis
Received: 7 September 2011 /Revised: 20 December 2011 /Accepted: 2 January 2012 /Published online: 17 January 2012# Springer-Verlag 2012
Abstract Reactive oxygen species (ROS) are strongly im-plicated in melanoma development, and treatment with anti-oxidants has shown efficacy in suppressing malignanttransition and progression. Here, we investigated the signif-icance of the glutamate-L-cysteine ligase catalytic subunit(GCLC) expression, a key regulator of glutathione synthe-sis, for malignant melanoma. A large set of melanoma celllines (n036) was analyzed, and higher GCLC levels wereassociated with lower presence of intracellular ROS andinterestingly also lower rates of cell proliferation. Moreover,treatment with the antioxidant N-acetylcysteine efficientlyreduced the growth speed of several investigated malignantcells. In addition GCLC expression was significantly linkedto a prominent set of cellular antioxidants, accounting for
the observed lower basal levels of oxidative stress andhigher antioxidative capacity. Key attributes defining themalignant phenotype of melanoma cells including survival,invasiveness, and switch from E-cadherin to N-cadherinexpression were more prominent in cells with lower GCLCexpression. Our findings were further corroborated byobservations in Rag2−/−γc−/−mice, in which melanoma cellswith lower GCLC expression depicted a dramatically strongertumor growth. Furthermore, prognostic significance of GCLCexpression was investigated in patients (n028) with advancedmalignant melanoma. High tumor immunoreactivity forGCLC was a significant determinant for better 5-year overallsurvival. Conclusively, we show for the first time that GCLCmay serve a dual role, as a surrogate marker for cellular redox
Dimitrios Mougiakakos, Riki Okita, C. Christian Johansson, and RolfKiessling contributed equally to this work.
Electronic supplementary material The online version of this article(doi:10.1007/s00109-012-0857-4) contains supplementary material,which is available to authorized users.
D. Mougiakakos :R. Okita :C. C. Johansson : R. KiesslingDepartment of Oncology and Pathology, Cancer CenterKarolinska, Karolinska Institutet,Stockholm, Sweden
D. Mougiakakos (*)Department of Internal Medicine 5, Hematology and Oncology,University of Erlangen,Glückstrasse 4a,Erlangen, Germanye-mail: [email protected]
T. AndoDepartment of Orthopaedic Surgery, University of Yamanashi,Yamanashi 409-3898, Japan
C. Dürr : R. ZeiserDivision of Hematology and Oncology, Department of Medicine,Freiburg University Medical Center, Albert-Ludwigs-University,Freiburg, Germany
J. Gadiot :C. BlankDepartment of Medical Oncology and Division of Immunology,The Netherlands Cancer Institute-Antoni van LeeuwenhoekHospital (NKI-AVL),Amsterdam 1066 CX, The Netherlands
J. IchikawaDepartment of Orthopaedics and Pharmacology,Vanderbilt University,Nashville, TN, USA
J Mol Med (2012) 90:935–944DOI 10.1007/s00109-012-0857-4
state as well as malignant potential of melanoma cells. Thesepromising results regarding its prognostic significance as wellas its potential as a pharmacological target require further in-depth investigations.
Malignant melanoma has dramatically increased in incidenceduring the past two decades, and therapeutic outcome remainsunsatisfactory [1]. Oxidative stress, as induced by UV irradi-ation, has been implicated in the carcinogenic process, whichis mainly due to oncogenic DNA damages [2]. In addition,enhanced production of reactive oxygen species (ROS) hasbeen linked to disease progression by promoting survivability,proliferation, and even horizontal cell growth [3, 4]. Notice-able, elevated ROS production can already be observed inpremalignant lesions, and importantly, gender-specific differ-ences in ROS production have been suggested as contributorsto better survival of female patients [5]. Endogenous antiox-idants are found at reduced levels in melanocytes from mela-noma patients, which is in line with observations of increasedROS levels in melanoma cell lines of more aggressive pheno-types [6–8]. Overexpression of superoxide dismutase andtreatment with antioxidants resulted in growth arrest of mela-noma cells [4, 9, 10]. In animal as well as in patient studies,the application of the antioxidant N-acetylcysteine (NAC)delayed tumor development and protected melanocytic naeviagainst UV-induced oxidative stress, respectively [11, 12].
Glutamate-L-cysteine ligase (GCL) catalyzes the rate-limiting step of glutathione [L-g-glutamyl-cysteinyl-glycine(GSH)] synthesis, the key cellular antioxidant. As a hetero-dimer, it is composed of the catalytically active heavy sub-unit (GCLC), which contains all the substrate-binding sites,and a regulatory subunit (GCLM) that modulates the GCLaffinity for glutamate. GCLC is (over-) expressed in severaltypes of tumors including mesothelioma [13], ependymoma[14], and breast cancer [15]. Despite the observations that anelevated GSH synthesis can be associated with the increasedradiochemoresistance and self-renewal of certain tumortypes, GCLC expression has been reported to correlatepositively with low-grade ependymomas and a better sur-vival in breast cancer [13–17]. These findings emphasize thecomplex role of oxidative stress in cancer, ranging frompromotion of progression to induction of apoptosis.
In our present study, we investigated the GCLC expres-sion in a large number of melanoma cell lines from theEuropean Searchable Tumour Cell Line and Data Bank(ESTDAB) [18]. As GSH holds a central role in cell phys-iology, we were interested in whether GCLC expression isassociated with the general antioxidative properties as well
as the malignant potential of melanoma cells. Our data showthat high expression of GCLC may be a promising surrogatemarker for low inherent oxidative stress, increased expres-sion of several antioxidant enzymes as well as low growthspeed and invasiveness, which altogether define a less ma-lignant phenotype. In line with our in vitro and in vivoobservations, advanced melanoma patients with high GCLCexpression within the primary tumor tissue had a better 5-year overall survival.
These results indicate that GCLC may represent a novelbiomarker of prognostic relevance and motivate the use ofROS-neutralizing agents in melanoma, especially in combi-nation with targeted (immunological) interventions.
Material and methods
Reagents DMEM, RPMI 1640 cell medium, fetal bovineserum (FBS), and trypan blue (0.4%) were purchased fromInvitrogen (Carlsbad, CA, USA); dimethyl sulfoxide, dacar-bazine (DTIC), hydrogen peroxide (H2O2), penicillin–strep-tomycin, Tween 20, and trypsin–EDTA solution wereobtained from Sigma-Aldrich (St. Louis, MO, USA) andIFN-γ from Boehringer Ingelheim (Ingelheim, Germany).
Cell lines All human melanoma cell lines (SupplementalTable 1) utilized in this study were obtained from the EST-DAB repository (http://www.ebi.ac.uk/ipd/estdab/). The ge-notype was identified using a STR Identifier kit (AppliedBiosystems) (Supplemental Table 1). All the cell lines werepositively authenticated matching in more than 80% of thealleles and/or showing evaluation values greater than 0.8 withthe previously reported STR profiles. Cells were cultured inRPMI 1640medium supplemented with 10% heat-inactivatedFBS, 2mML-glutamine, and 50U/ml penicillin–streptomycinat 37°C in a humidified atmosphere of 5% CO2.
Mice Rag2-/-γc-/- mice were purchased from the localstock of the animal facility at Freiburg University (Ger-many). The animal protocol (G08-8) was approved by theRegierungspräsidium Freiburg (local animal committeeFreiburg). Mice were injected with 2×106 melanoma cellssubcutaneously. The tumor size (millimeters) was retracedby measuring the diameter of the tumor with a micrometer(Mitutoyo Europe, Neuss, Germany), and the average of twomeasurements at each time point was taken. The animals weresacrificed when the tumor size reached a diameter above7 mm or showed ulceration. Tissues were fixed using 4%paraformaldehyde for at least 48 h and later embedded inparaffin.
Patient material Our study included formaldehyde-fixed,paraffin-embedded primary cutaneous malignant melanoma
specimens from 28 stage III–IV patients with completeclinicopathological and follow-up data. Patients were trea-ted at the Department of Medical Oncology of the Nether-lands Cancer Institute during the period 2000–2009. Thestudy was conducted according to the Declaration ofHelsinki, and an ethical approval was obtained from thecentral medical ethic committee of the NKI-AVL, Amsterdam.The patients' clinicopathological characteristics are summa-rized in Table 1.
Western blot analysis Cells were lysed in a CelLytic (Sig-ma-Aldrich) buffer containing Protease Inhibitor Cocktail(Sigma-Aldrich) and Phosphatase Inhibitor Cocktail 2 (Sig-ma-Aldrich). The protein concentration of cell lysates wasdetermined by a BCA Protein assay (Thermo Scientific,Rockford, IL, USA). Proteins (3–5 mg/lane) were size frac-tioned by 4–12% NuPage Bis-Tris gels (Invitrogen) andtransferred onto polyvinylidene difluoride membranes(Immobilon-P, Millipore, Bedford, MA, USA). All mem-branes were blocked in PBS/0.05% Tween 20/2% milkpowder for 30 min and washed prior to overnight incubationwith primary antibodies against GCLC (sc-22755, SantaCruz Biotech, San Diego, CA, USA), AKT (#9272, CellSignaling, Frankfurt, Germany), pAKT (#9271, Cell Sig-naling), ERK1/2 (#9102, Cell Signaling), pERK1/2(#9101, Cell Signaling), and β-actin (Sigma-Aldrich).After rinsing in PBS/0.05% Tween 20, membranes wereincubated with HRP-conjugated goat anti-mouse (CellSignaling) or anti-rabbit (Cell Signaling) secondary anti-bodies for 1 h at room temperature. Membranes weredeveloped using ECL plus detection systems (GE Health-care, Fairfield, CT, USA). Semiquantitative protein
quantification was performed with Image J 1.42q soft-ware (http://rsb.info.nih.gov/ij).
Real-time PCR Total RNA was extracted using an RNeasymini kit (Qiagen, Hilden, Germany) and transcribed (re-verse) to cDNA (Bio-Rad, Hercules, CA, USA). MessengerRNA (mRNA) was quantified by real-time PCR (SYBRGreen Supermix, Applied Biosystems) on an ABI 7500Detection System (Applied Biosystems). Relative gene ex-pression was determined by normalizing the gene expres-sion of each target gene to the β-actin housekeeping geneexpression. Gene-specific primer sequences are listed inSupplemental Table 2.
Flow cytometry The fluorochrome-conjugated mouse anti-human antibodies CD44-APC (IM7), ICAM-1-FITC(23C6), N-Cadherin-PE (8C11), and MHCI-PE (W6/32)were purchased from eBioscience (San Diego, CA, USA),CD54-APC (HCD54) from Biolegend (San Diego, CA,USA), E-Cadherin-PE (180224) from R&D Systems (Min-neapolis, MN, USA), and MHCII-FITC from BD Bioscien-ces (San Diego, CA, USA). All cell stainings wereperformed according to the manufacturers' recommenda-tions and cells analyzed using a LSRII flow cytometer(BD Biosciences) and FlowJo 9.0.2 software (Treestar, Ash-land, OR).
Cell-doubling calculation 1×106 cells were seeded in T-25flasks, and cell number was determined after 48 h mi-croscopically following trypan blue staining. The growthrate for each cell line was assessed at least twice, andthe results were found to be reproducible. Doublingtimes derived from the exponential growth phase werecalculated as follows:
DT¼ 48�log 2ð Þ=log cell number after 48�h culture=seeded cell numberð Þ
Oxidative stress measurements The levels of intracellularoxidation were assessed by flow cytometry after thecells were labeled with 5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein-diacetate-acetyl-ester (CM-H(2)-DCFDA) (Invitrogen) which is metabolized to fluo-rescent (FL-1) 2′,7′-dichlorofluorescein (DCF) upon in-tracellular oxidation.
Measuring cell surface thiols Surface thiol contentwas assessed by flow cytometry upon staining withAlexa Fluor 633-coupled maleimide (ALM-633;Invitrogen).
Cell death assays Cell death was determined by flowcytometry upon 7-AAD (BD Biosciences) and Annexin V-
Table 1 The patients' (n028) clinicopathological characteristics
Age (years, mean, range) 56.7 (21–90)
Gender (n and %)
Male 16 57%
Female 12 43%
Breslow index (mm, mean range) 4.3 (1.6–12.5)
Ulceration Yes 17 61%
No 5 18%
Unknown 6 21%
Histology SSM 9 32%
NM 15 54%
Unclear 4 14%
AJCC stage at time of sampling
III 15 54%
IV 13 46%
n number of patients, mm millimeters, SSM superficial spreadingmelanoma, NM nodular melanoma, AJCC American Joint Committeeon Cancer
FITC (eBioscience) co-staining. The percentage of specificcell death was calculated according to the following formula:
Specific cell death %alð Þ ¼ 100� %dead cells�%baseline dead cellsð Þ= 100%�%baseline dead cellsð Þ:
Determination of intracellular glutathione levels The intra-cellular content of reduced (GSH), oxidized (GSSG), andtotal glutathione was measured by a Glutathione Assaykit (Biovision, Mountain View, CA, USA). Briefly, sam-ples were homogenized and prepared according to themanufacturer's protocol for determination of GSH andGSSG. A reducing agent mix converted GSSG to GSH,and o-phtalaldehyde was added for 40 min at roomtemperature. Subsequently, samples were read on a fluo-rescence plate reader (Wallac 1420 Victor 2, PerkinElmer, Waltham, MA, USA) and GSH/GSSG concentra-tions calculated.
Cell invasion assay Cell invasion was assessed by usinga QCM cell invasion assay (Millipore, Frederik, MD,USA). Briefly, cells were loaded into the chambers of a96-well plate with pores (8 mm) coated with a thinlayer of ECM matrix. Upon overnight culturing, inva-sive cells migrated through this matrix and attached tothe bottom of the membrane. Following detachmentfrom the membrane by a Cell Detachment Buffer, cellswere lysed and stained with CyQuant GR Dye, and cellnumber was determined by a fluorescence plate readerfollowing the manufacturer's instructions (Wallac 1420Victor 2).
Immunohistochemistry Tumor tissue derived from patientsand mice was stained for GCLC expression and tumor-infiltrating lymphocytes as described in detail within thesection of supplemental information. Staining assessmentwas performed according to previous publications [15, 19].Two variables, number of GCLC positive cells and overallintensity of immunoreactivity, were scored independently.Briefly, the scoring for number of positive tumor cells wasdefined as follows: “0,” no positive cells; “1,” <10% posi-tive cells; “2,” 10–30% positive cells; “3,” 30–60% positivecells; and “4,” >60% positive cells. The intensity scoringwas defined as follows: “0,” no staining; “1,” weak stain-ing; “2,” moderate staining; “3,” strong staining; and“4,”very strong staining. Both variables multiplied by eachother yield a product (score) that allows categorization ofgroups with negative/low (score 0–4) and high (score >4)immunoreactivity, respectively. Infiltrating T cells (sum ofCD4± and CD8± cells) were scored as the percentage oftumor cells: “0,” ≤1% positive cells; “1,” 1–5% positivecells; “2,” 5–30% positive cells; and “3,” ≥30% positivecells.
Statistics According to the distribution levels, differences inmeans and correlation analyses were evaluated with para-metric (two-tailed Student's or paired t test and Pearson'stest) or nonparametric (Mann–Whitney U or Wilcoxon andSpearman's ρ test) tests. Cumulative survival time was cal-culated by the Kaplan–Meier method and analyzed by log-rank testing. All analyses were performed at a significancelevel of 5% (p<0.05) using SPSS version 16.0 (SPSS Inc.,IL, USA).
Results
Correlation of GCLC expression, cell growth,and intracellular ROS
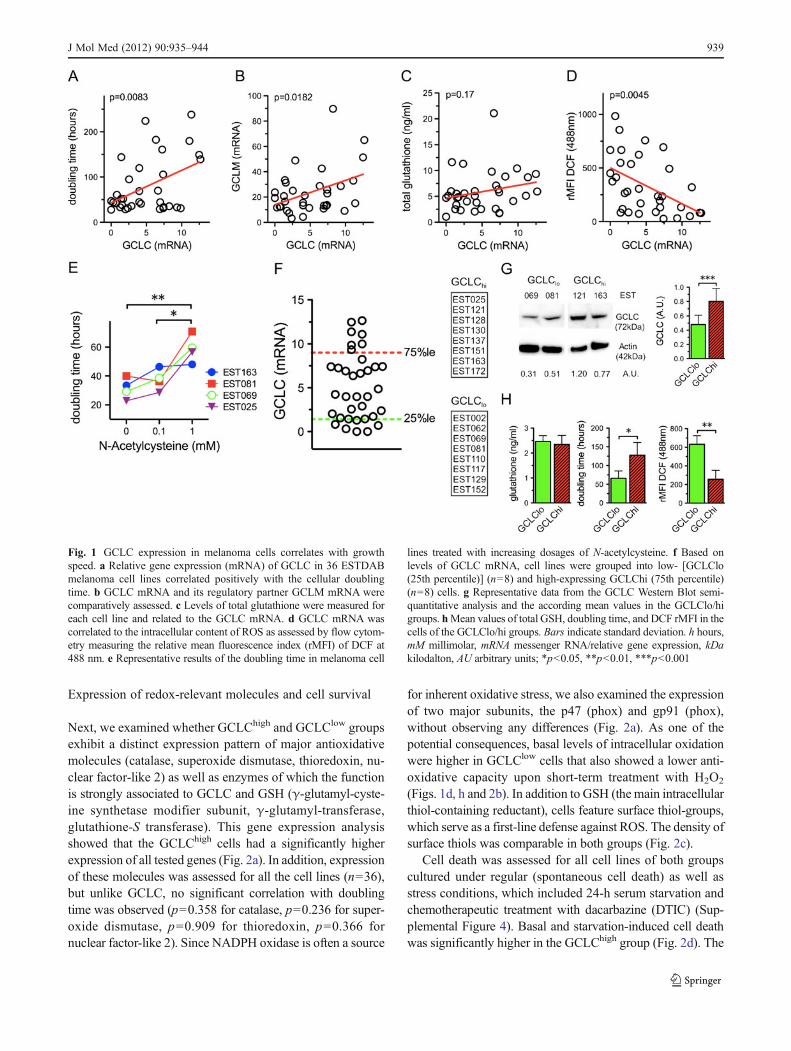
Levels of GCLC mRNA correlated positively with an in-creased doubling time of melanoma cell lines (Fig. 1a). GCLCand GCLM form GCL, which mediates the rate-limiting stepof the cellular GSH synthesis. Albeit relative gene expressionof both molecules correlated significantly (Fig. 1b), higherGCLC levels were not linked to an increased intracellularcontent of total glutathione (Fig. 1c), the reduced (GSH) oroxidized (GSSG) form, as well as the GSH/GSSG ratio (Sup-plemental Figure 1). However increasing GCLC expressionwas associated with reduced intracellular ROS levels(Fig. 1d). Treatment with the thiol-containing antioxidantNAC led to a growth speed reduction in tested melanoma celllines (Fig. 1e) similar to previous reports [4] and without anynegative impact on their viability (Supplemental Figure 2). Asexpected, NAC replenished the cellular glutathione levels, butwe could not observe any disproportionate increase of GSH orGSSG (Supplemental Figure 3).
For further analyses, we grouped the cell lines of the 25thand 75th percentile (based on their GCLC mRNA expres-sion) into a GCLClow and GCLChigh group (Fig. 1f). Inaccordance with the data from the gene expression analysis,GCLC protein levels were significantly distinct betweenthese two groups (GCLClow, 0.52±0.16; GCLChigh, 0.78±0.18; p00.003) (Fig. 1g). These results suggested thatGCLC expression is primarily regulated at the mRNA levelby either transcriptional and/or post-transcriptional mecha-nisms. As anticipated and based on the aforementionedresults, the cells in both groups exhibited similar glutathionelevels but a distinct growth speed and intracellular ROS yield(Fig. 1h).
938 J Mol Med (2012) 90:935–944
Expression of redox-relevant molecules and cell survival
Next, we examined whether GCLChigh and GCLClow groupsexhibit a distinct expression pattern of major antioxidativemolecules (catalase, superoxide dismutase, thioredoxin, nu-clear factor-like 2) as well as enzymes of which the functionis strongly associated to GCLC and GSH (γ-glutamyl-cyste-ine synthetase modifier subunit, γ-glutamyl-transferase,glutathione-S transferase). This gene expression analysisshowed that the GCLChigh cells had a significantly higherexpression of all tested genes (Fig. 2a). In addition, expressionof these molecules was assessed for all the cell lines (n036),but unlike GCLC, no significant correlation with doublingtime was observed (p00.358 for catalase, p00.236 for super-oxide dismutase, p00.909 for thioredoxin, p00.366 fornuclear factor-like 2). Since NADPH oxidase is often a source
for inherent oxidative stress, we also examined the expressionof two major subunits, the p47 (phox) and gp91 (phox),without observing any differences (Fig. 2a). As one of thepotential consequences, basal levels of intracellular oxidationwere higher in GCLClow cells that also showed a lower anti-oxidative capacity upon short-term treatment with H2O2
(Figs. 1d, h and 2b). In addition to GSH (the main intracellularthiol-containing reductant), cells feature surface thiol-groups,which serve as a first-line defense against ROS. The density ofsurface thiols was comparable in both groups (Fig. 2c).
Cell death was assessed for all cell lines of both groupscultured under regular (spontaneous cell death) as well asstress conditions, which included 24-h serum starvation andchemotherapeutic treatment with dacarbazine (DTIC) (Sup-plemental Figure 4). Basal and starvation-induced cell deathwas significantly higher in the GCLChigh group (Fig. 2d). The
Fig. 1 GCLC expression in melanoma cells correlates with growthspeed. a Relative gene expression (mRNA) of GCLC in 36 ESTDABmelanoma cell lines correlated positively with the cellular doublingtime. b GCLC mRNA and its regulatory partner GCLM mRNA werecomparatively assessed. c Levels of total glutathione were measured foreach cell line and related to the GCLC mRNA. d GCLC mRNA wascorrelated to the intracellular content of ROS as assessed by flow cytom-etry measuring the relative mean fluorescence index (rMFI) of DCF at488 nm. e Representative results of the doubling time in melanoma cell
lines treated with increasing dosages of N-acetylcysteine. f Based onlevels of GCLC mRNA, cell lines were grouped into low- [GCLClo(25th percentile)] (n08) and high-expressing GCLChi (75th percentile)(n08) cells. g Representative data from the GCLC Western Blot semi-quantitative analysis and the according mean values in the GCLClo/higroups. hMean values of total GSH, doubling time, and DCF rMFI in thecells of the GCLClo/hi groups. Bars indicate standard deviation. h hours,mM millimolar, mRNA messenger RNA/relative gene expression, kDakilodalton, AU arbitrary units; *p<0.05, **p<0.01, ***p<0.001
J Mol Med (2012) 90:935–944 939
susceptibility to chemotherapy-induced cell death was signif-icantly higher (p00.04) in the GCLChigh group at a dosage of250 μg/ml of DTIC and showed a similar tendency (p00.06)at 100 μg/ml (Fig. 2d).
Expression of surface molecules associated with metastaticpotential and/or immunological recognition of melanomacells
Several surface molecules have been found to be positivelyor negatively involved in the invasive and/or metastatic
potential of melanoma cells. Among the tested negativeprognostic markers, ICAM-1 and N-cadherin were bothsignificantly (p00.004 and p00.025, respectively) higherexpressed in the GCLClow group (Fig. 3a, SupplementalFigure 5). In contrast, E-cadherin, which is considered apositive prognostic marker, was found at significantly(p00.017) higher levels in the GCLChigh group (Fig. 3a,Supplemental Figure 5). Successful silencing of the GCLCgene (Supplemental Figure 6) did not lead to any substantialreversal of the phenotype (Supplemental Figure 7) implyingno direct GCLC-dependent regulation. Alterations in ROS-sensitive kinase activities and in particular AKT (proteinkinase B) as well as ERK1/2 (extracellular signal-regulatedkinases) have been linked to progression and survivabilityof melanoma cells. No differential expression and/or activa-tion were observed (Fig. 3b, Supplemental Figure 8). Fur-thermore, in vitro invasiveness was tested for both groups,and a significantly (p00.013) higher cell transmigration wasseen for the GCLClow group (Fig. 3c). As it is well describedthat antigen presentation through MHCs can largely beaffected in melanoma cells, we analyzed both groups withrespect to basal as well as IFN-γ-induced MHC class I/IIexpression. We could not find any differences between bothgroups (Fig. 3d and e).
In vivo tumorigenicity
Two randomly chosen cell lines from each group (GCLClow
EST067/081, GCLChigh EST128/163) were injected subcu-taneously into Rag2-/-γc-/- mice. The in vivo tumor growthstrongly correlated with the GCLC levels (Fig. 4a, b). Micereceiving GCLClow cells had to be sacrificed at a muchearlier time point due to the rapid tumor growth (p00.014)(Fig. 4c).
Fig. 2 Cells with high GCLC expression have lower levels of inherentoxidative stress, higher antioxidative capacity, and decreased surviv-ability. a The relative gene expression of the antioxidative enzymescatalase (cat), superoxide dismutase (sod), thioredoxin-1 (trx1) andnuclear related factor 2 (nrf2), the GCLC/GSH-related enzymesglutamate-L cysteine ligase modifier subunit (gclm), γ-glutamyl-transferase (ggt), glutathione-S transferase (gst), and of the two sub-units (p47 and gp91) of the reactive oxygen species producing NADPHcomplex was determined by real-time PCR in the cells of the GCLClo/hi groups. b Representative data of two of the tested cell lines(EST081; GCLClo) and (EST025; GCLChi), respectively, labeled withCM-H2DCFDA for the detection of basal intracellular levels of oxi-dative stress (DCF) and upon treatment with 100 μM H2O2 as well asthe according mean values of the GCLClo/hi groups. c Surface thiolswere quantified by staining the GCLClo/hi cells with Alexa Fluor 633coupled maleimides (ALM). d Mean values of spontaneous, (serum-)starvation (24 h) as well as dacarbazine (DTIC)-induced cell death asassessed by flow cytometry in the GCLClo/hi groups. rMFI relativemean fluorescence index. *p<0.05, **p<0.01, ***p<0.001
R
940 J Mol Med (2012) 90:935–944
Expression of GCLC in primary malignant melanoma
We investigated, retrospectively, GCLC expression in pri-mary malignant melanoma samples from 28 stage III–IVpatients (Fig. 5). No or low GCLC was found in 20 of 28(71%) of the cases. Strong expression of GCLC, defined asthe cumulative score >4, was detectable in 8 of 28 (29%) ofthe patients. Expression of GCLC in the primary tissue wasnot associated (data not shown) to any of the availableclinicopathologic parameters (Table 1) or the intensity oflymphocytic infiltration (Supplemental Figure 9). However,5-year survival rate in the GCLChigh cohort (5 of 8, 63%)was higher than in the GCLClow (4 of 20, 20%), andKaplan–Meier analysis showed that GCLC expression wassignificantly (p00.039) associated to the 5-year overall sur-vival (Fig. 5).
Discussion
Oxidative stress is associated with melanomagenesis andfavors melanoma progression [2–4]. It is mediated by ROSthat on the one hand induces pro-oncogenic DNA mutationsand, furthermore, as bona fide second messengers promotemalignant behavior by activating kinases and thereby cellproliferation [14, 20]. Since high levels of oxidative stress,as induced by radiochemotherapy, may also result in celldeath, it is evident that there is a fine balance betweendetrimental and beneficial ROS-mediated effects [21]. Alter-ations in pro- and antioxidants during carcinogenesis as wellas their levels in the melanoma-surrounding/healthy tissuehave been a well-studied subject [4, 8, 22]. Altogether, thevolume of findings suggests that melanoma cells have anincreased inherent oxidative stress but at the same time
Fig. 3 Expression of molecules associated with malignant transforma-tion, metastasis, immunogenicity and in vitro invasiveness. a Differ-ences in the mean values (MFI) of intercellular adhesion molecule-1(ICAM-1), N-/E-cadherin (N-/E-Cad), Integrin αvβ3 (CD51/CD61),and CD44 were evaluated in the cells of the GCLClo and GCLChigroups. b The basal levels of total ERK1/2 and AKT as well as theiractive, phosphorylated forms (pERK1/2 and pAKT) were quantified byWestern Blot analysis for all cells of the GCLClo and GCLChi groups.
c GCLClo/hi cells were placed in the according chambers of a cellinvasion assay (Millipore) and cultured overnight. Invaded cells werecounted semiquantitatively. d Basal expression of MHC-I and MHC-IIon the cells of the GCLC lo/hi groups and the respective upregulation efollowing 24-h stimulation with indicated doses of IFN-γ wereassessed by flow cytometry. Bars indicate standard deviation. AUarbitrary units, μg microgram, ml milliliter. *p<0.05, **p<0.01,***p<0.001
J Mol Med (2012) 90:935–944 941
upregulate, potentially for reasons of self-protection, theirantioxidative machinery. It may be speculated that highROS levels promote muta- and carcinogenesis, while sur-rounding tissue as well as infiltrating immune cells aredamaged. Since antioxidants obviously can have an ambiv-alent role in cancer, we were interested in the significance ofGCLC expression in melanoma. Levels of GSH, the mostabundant intracellular nonenzymatic antioxidant, are strong-ly regulated by the rate-limiting GCLC [23]. We foundGCLC at various levels in different melanoma cell lines,and its increased expression was significantly associatedwith lower cell growth. These results are in line with ourand others' observations that treatment of melanoma cellswith antioxidants can reduce tumor activity, proliferation,
and risk for malignant transformation of naevi [4, 11, 12]. Itis not surprising that GCLC expression and GSH contentwere not positively correlating since GCLC is an enzymeunder allosteric feedback control and it is not self-evidentthat expression levels reflect the actual cellular activity [24].A guanine–adenine–guanine repeat polymorphism in the 5′-untranslated region of the GCLC gene has also been asso-ciated with a varying GCLC activity and GSH production[25]. Furthermore, intracellular transport of cysteine (aGCLC substrate) by the xc− system represents a critical stepfor GSH synthesis and depends on quantity as well asstability of the according transport channels (e.g., the xCTsubunit) [26]. Of course, efflux of intracellular GSH throughamongst others the multidrug resistance protein 1 can
Fig. 5 Expression of GCLC inprimary cutaneous malignantmelanoma. The followingrepresentative photomicrographsa show immunostainings ofprimary malignant melanomaswith (i) weak and (ii) strongimmunoreactivity for the anti-GCLC antibody (originalmagnification, ×100). b TheKaplan–Meier survival curvesillustrate the GCLC expression.A significantly better survivalwas noted for the group with astrong GCLC immunoreactivity(n08) versus low (n020)
Fig. 4 Tumorigenicity of melanoma cells expressing high and lowlevels of GCLC. a Rag2-/-γc-/- mice were injected subcutaneously inthe flank with EST067 (n02), EST081 (n02) (both GCLClo) andEST128 (n02), EST163 (n02) (both GCLChi) cells. a RepresentativeIHC GCLC stainings of the EST081 (GCLClo) and EST163 (GCLChi)
tumor formations using a rabbit polyclonal anti-human GCLC antibody(original magnification, ×100). b Tumor formation was monitored anddata are expressed as the largest average tumor diameter (millimeters)measured at each time point. c Survival of the pooled GCLC (GCLCloversus GCLChi) groups was assessed by log-rank testing
942 J Mol Med (2012) 90:935–944
negatively impact the GSH content. Notably, in murine B16melanoma cells, high GSH efflux has been associated withan increased GCLC expression [27]. Interestingly, levels ofGCLC expression correlated positively with all the testedantioxidants. Consequently, GCLChigh cells exhibited lowerbasal levels of oxidative stress and a higher antioxidativecapacity, suggesting that GCLC may qualify as a surrogatemarker for the redox balance in melanoma cells. The extrin-sic application of enzymatic antioxidants has been efficientin suppressing in vitro and in vivo the malignant phenotypeand progression of melanomas [28, 29]. In addition, mela-nomas produce large amounts of inherent ROS throughoverexpression of the NADPH oxidase (especially thegp91 phox subunit), with substantial impact on the cellcycle [4]. There was no difference in the expression of thetested NADPH subunits among our GCLC groups. Oxida-tive stress is also known to promote cell survival in mam-malian cells by inducing activation of the AKT and ERK1/2pathways [21]. In fact, we observed a lower spontaneous aswell as stress-induced cell death in the GCLClow cells.However, ROS-sensitive AKT and ERK kinases that arestrongly involved in melanoma progression were not differ-entially expressed or activated in the GCLC subgroups [3,30]. As previously mentioned, ROS can act as a secondmessenger augmenting migration and anoikis [5, 21, 31].For metastasis, malignant cells need to adhere to endothelialwalls and to extravasate into the distant site. Indeed, the invitro invasiveness was higher for GCLClow cells. One of themajor adhesion molecules mediating these processes isICAM-1. It has been shown that ROS and antioxidantsaffect its expression in an opposed way resulting in substan-tial consequences for the disease course [32, 33]. Duringtumorigenic transition in melanoma, loss of E-cadherin isaccompanied by gain in N-cadherin. This transition is asso-ciated with a loss of control by keratinocytes, increasedgrowth, and metastatic tendencies [34]. In breast cancer,where a GCLC deficit is associated with a more invasivetumor form, it has also been shown that increased oxidativestress could lead to loss of E-cadherin [15, 35]. Similarly,surface glycoprotein CD44, which has been implicated inprogression and metastasis of melanoma as well as having aprognostic value, can also be regulated by ROS [36, 37]. Inaccordance with the higher ROS levels, we found increasedlevels of N-cadherin and ICAM-1, and decreased E-cadherinlevels in the GCLClow group, further supporting the role ofGCLC as a suitable marker for the redox/malignant melanomaphenotype. We did not observe any differences regardingsurface CD44. Oxidative stress promotes CD44 expression[36, 37], and recent reports suggest that CD44 itself is in-volved in the cellular protection against ROS by regulating thecysteine transport, thereby forming a potential feedback cir-cuit [26]. Several defects in the ROS-sensitive antigen presen-tation machinery have been described for melanoma [38, 39].
We could not detect any alterations in the basal and inducibleMHC-I and MHC-II expression being associated with GCLClevels. Proteomic analyses in murine models have shown thatduring the process in which melanoma cells acquire a moremalignant phenotype, they lose the ability to sufficiently elim-inate ROS, and anti/pro-apoptotic pathways are shifted to-wards survival [6]. Our in vitro data were confirmed in invivo experiments clearly showing an increased malignantpotential in the GCLClow melanomas. In our analysis of asmall cohort of stage III–IV melanoma patients, high expres-sion of GCLC was, similarly to breast cancer and ependy-moma, associated with a significantly higher 5-year overallsurvival [14, 15]. In order to perform accurate multivariateanalyses (with well-established markers) and enable us tobetter estimate the value of GCLC as a prognostic marker,larger patient cohorts will be studied in the future.
Overall, we were able to show for the first time thatGCLC expression may be an appropriate surrogate markerfor the redox phenotype of melanoma cells, a phenotype thatis strongly associated with the malignant behavior. Further-more, our observations (together with several previous stud-ies) suggest that it may be feasible in melanoma to achieve abetter control of progression and recurrence at distant sitesthrough tackling ROS production. Further clinical studiesexploring this therapeutic alternative of low toxicity andcost (as for example introduced with the preventive NACtrial) should be encouraged [12].
Acknowledgments This work was supported by grants from theCancer Society of Stockholm, Karolinska Institutet, Stockholm CityCouncil, Swedish Cancer Society, and Swedish Research Council. D.M. was supported by the German Research Association (DFG) and theInterdisciplinary Centre for Clinical Research (IZKF), Erlangen.
Disclosure The authors declare no competing financial interest.
References
1. Tsao H, Atkins MB, Sober AJ (2004) Management of cutaneous mela-noma. N Engl J Med 351:998–1012
2. Meyskens FL Jr, Farmer PJ, Anton-Culver H (2004) Etiologicpathogenesis of melanoma: a unifying hypothesis for the missingattributable risk. Clin Cancer Res 10:2581–2583
3. Govindarajan B, Sligh JE, Vincent BJ, Li M, Canter JA, Nickoloff BJ,Rodenburg RJ, Smeitink JA, Oberley L, Zhang Y et al (2007) Over-expression of Akt converts radial growth melanoma to vertical growthmelanoma. J Clin Invest 117:719–729
4. Yamaura M, Mitsushita J, Furuta S, Kiniwa Y, Ashida A, Goto Y,Shang WH, Kubodera M, Kato M, Takata M et al (2009) NADPHoxidase 4 contributes to transformation phenotype of melanomacells by regulating G2-M cell cycle progression. Cancer Res69:2647–2654
5. Joosse A, De Vries E, van Eijck CH, Eggermont AM, Nijsten T,Coebergh JW (2010) Reactive oxygen species and melanoma: anexplanation for gender differences in survival? Pigment Cell Melano-ma Res 23:352–364
J Mol Med (2012) 90:935–944 943
6. de Souza GA, Godoy LM, Teixeira VR, Otake AH, Sabino A, RosaJC, Dinarte AR, Pinheiro DG, Silva WA Jr, Eberlin MN et al (2006)Proteomic and SAGE profiling of murine melanoma progressionindicates the reduction of proteins responsible for ROS degradation.Proteomics 6:1460–1470
7. Grammatico P, Maresca V, Roccella F, Roccella M, Biondo L,Catricala C, Picardo M (1998) Increased sensitivity to peroxidizingagents is correlated with an imbalance of antioxidants in normalmelanocytes from melanoma patients. Exp Dermatol 7:205–212
8. Picardo M, Grammatico P, Roccella F, Roccella M, Grandinetti M,Del Porto G, Passi S (1996) Imbalance in the antioxidant pool inmelanoma cells and normal melanocytes from patients with mela-noma. J Invest Dermatol 107:322–326
9. Prasad KN, Edwards-Prasad J (1982) Effects of tocopherol (vita-min E) acid succinate on morphological alterations and growthinhibition in melanoma cells in culture. Cancer Res 42:550–555
10. Yang J, Su Y, Richmond A (2007) Antioxidants tiron and N-acetyl-L-cysteine differentially mediate apoptosis in melanoma cells via areactive oxygen species-independent NF-kappaB pathway. Free RadicBiol Med 42:1369–1380
11. Cotter MA, Thomas J, Cassidy P, Robinette K, Jenkins N, Florell SR,Leachman S, Samlowski WE, Grossman D (2007) N-acetylcysteineprotects melanocytes against oxidative stress/damage and delaysonset of ultraviolet-induced melanoma in mice. Clin Cancer Res13:5952–5958
12. Goodson AG, Cotter MA, Cassidy P, Wade M, Florell SR, Liu T,Boucher KM, Grossman D (2009) Use of oral N-acetylcysteine forprotection of melanocytic nevi against UV-induced oxidativestress: towards a novel paradigm for melanoma chemoprevention.Clin Cancer Res 15:7434–7440
13. Jarvinen K, Soini Y, Kahlos K, Kinnula VL (2002) Overexpressionof gamma-glutamylcysteine synthetase in human malignant meso-thelioma. Hum Pathol 33:748–755
14. Jarvela S, Nordfors K, Jansson M, Haapasalo J, Helen P, PaljarviL, Kalimo H, Kinnula V, Soini Y, Haapasalo H (2008) Decreasedexpression of antioxidant enzymes is associated with aggressivefeatures in ependymomas. J Neurooncol 90:283–291
15. Soini Y, Karihtala P, Mantyniemi A, Turunen N, Paakko P, KinnulaV (2004) Glutamate-L-cysteine ligase in breast carcinomas. Histo-pathology 44:129–135
16. Tatebe S, Unate H, Sinicrope FA, Sakatani T, Sugamura K, MakinoM, Ito H, Savaraj N, Kaibara N, KuoMT (2002) Expression of heavysubunit of gamma-glutamylcysteine synthetase (gamma-GCSh) inhuman colorectal carcinoma. Int J Cancer 97:21–27
17. Tew KD (1994) Glutathione-associated enzymes in anticancerdrug resistance. Cancer Res 54:4313–4320
18. Pawelec G, Marsh SG (2006) ESTDAB: a collection of immuno-logically characterised melanoma cell lines and searchable data-bank. Cancer Immunol Immunother 55:623–627
19. Gadiot J, Hooijkaas AI, Kaiser AD, van Tinteren H, van Boven H,Blank C (2011) Overall survival and PD-L1 expression in metasta-sized malignant melanoma. Cancer 117:2192–2201
20. Burdon RH (1995) Superoxide and hydrogen peroxide in relationto mammalian cell proliferation. Free Radic Biol Med 18:775–794
21. Fruehauf JP, Meyskens FL Jr (2007) Reactive oxygen species: abreath of life or death? Clin Cancer Res 13:789–794
23. Meister A (1995) Glutathione metabolism.Methods Enzymol 251:3–724. Franklin CC, Backos DS,Mohar I,White CC, Forman HJ, Kavanagh
TJ (2009) Structure, function, and post-translational regulation of thecatalytic and modifier subunits of glutamate cysteine ligase. MolAspects Med 30:86–98
25. Walsh AC, Feulner JA, Reilly A (2001) Evidence for functionallysignificant polymorphism of human glutamate cysteine ligase cat-alytic subunit: association with glutathione levels and drug resis-tance in the National Cancer Institute tumor cell line panel. ToxicolSci 61:218–223
26. Ishimoto T, Nagano O, Yae T, Tamada M, Motohara T, Oshima H,Oshima M, Ikeda T, Asaba R, Yagi H et al (2011) CD44 variantregulates redox status in cancer cells by stabilizing the xCT subunitof system xc(-) and thereby promotes tumor growth. Cancer Cell19:387–400
27. Benlloch M, Ortega A, Ferrer P, Segarra R, Obrador E, Asensi M,Carretero J, Estrela JM (2005) Acceleration of glutathione effluxand inhibition of gamma-glutamyltranspeptidase sensitize meta-static B16 melanoma cells to endothelium-induced cytotoxicity. JBiol Chem 280:6950–6959
28. Church SL, Grant JW, Ridnour LA, Oberley LW, Swanson PE,Meltzer PS, Trent JM (1993) Increased manganese superoxidedismutase expression suppresses the malignant phenotype of hu-man melanoma cells. Proc Natl Acad Sci USA 90:3113–3117
29. Ranganathan AC, Nelson KK, Rodriguez AM, Kim KH, TowerGB, Rutter JL, Brinckerhoff CE, Huang TT, Epstein CJ, Jeffrey JJet al (2001) Manganese superoxide dismutase signals matrix met-alloproteinase expression via H2O2-dependent ERK1/2 activation.J Biol Chem 276:14264–14270
30. Cohen C, Zavala-Pompa A, Sequeira JH, Shoji M, Sexton DG,Cotsonis G, Cerimele F, Govindarajan B, Macaron N, Arbiser JL(2002) Mitogen-actived protein kinase activation is an early eventin melanoma progression. Clin Cancer Res 8:3728–3733
31. Albini A, D'Agostini F, Giunciuglio D, Paglieri I, Balansky R, DeFlora S (1995) Inhibition of invasion, gelatinase activity, tumortake and metastasis of malignant cells by N-acetylcysteine. Int JCancer 61:121–129
32. Hamai A, Meslin F, Benlalam H, Jalil A, Mehrpour M, Faure F,Lecluse Y, Vielh P, Avril MF, Robert C et al (2008) ICAM-1 has acritical role in the regulation of metastatic melanoma tumor sus-ceptibility to CTL lysis by interfering with PI3K/AKT pathway.Cancer Res 68:9854–9864
33. ten Kate M, van der Wal JB, Sluiter W, Hofland LJ, Jeekel J, Sonne-veld P, van Eijck CH (2006) The role of superoxide anions in thedevelopment of distant tumour recurrence. Br J Cancer 95:1497–1503
34. Li G, Satyamoorthy K, Herlyn M (2001) N-cadherin-mediated inter-cellular interactions promote survival and migration of melanomacells. Cancer Res 61:3819–3825
35. Parmar H, Melov S, Samper E, Ljung BM, Cunha GR, Benz CC(2005) Hyperplasia, reduced E-cadherin expression, and develop-mental arrest in mammary glands oxidatively stressed by loss ofmitochondrial superoxide dismutase. Breast 14:256–263
36. Dietrich A, Tanczos E, Vanscheidt W, Schopf E, Simon JC (1997)High CD44 surface expression on primary tumours of malignantmelanoma correlates with increased metastatic risk and reducedsurvival. Eur J Cancer 33:926–930
37. Vendrov AE, Madamanchi NR, Niu XL, Molnar KC, Runge M,Szyndralewiez C, Page P, Runge MS (2010) NADPH oxidasesregulate CD44 and hyaluronic acid expression in thrombin-treatedvascular smooth muscle cells and in atherosclerosis. J Biol Chem285:26545–26557
38. Jager E, Ringhoffer M, Altmannsberger M, Arand M, Karbach J,Jager D, Oesch F, Knuth A (1997) Immunoselection in vivo: inde-pendent loss of MHC class I and melanocyte differentiation antigenexpression in metastatic melanoma. Int J Cancer 71:142–147
39. Seifert U, Bialy LP, Ebstein F, Bech-Otschir D, Voigt A, Schroter F,Prozorovski T, Lange N, Steffen J, Rieger M et al (2010) Immuno-proteasomes preserve protein homeostasis upon interferon-inducedoxidative stress. Cell 142:613–624