High Glucose Inhibits Nitric Oxide Production in Cultured Rat Mesangial Cells HOWARD TRACHTMAN, STEPHEN FUTTERWEIT, and DAN L. CRIMMINS Division of Nephrology, Schneider Children ‘s Hospital, Long Island Jewish Medical Center, Long island Campus for Albert Einstein College of Medicine, New Hyde Park, New York; and Department of Molecular Biology and Pharmacology. and Protein and Nucleic Acid Chemistry Laboratory, Washington University School of Medicine, St. Louis, Missouri. Abstract. Hyperglycemia directly contributes to the develop- ment of diabetic nephropathy. A high-serum glucose concen- tration alters intraglomerular hemodynamics and promotes deposition of extracellular matrix in the kidney. Nitric oxide (NO) is a short-lived messenger molecule that participates in the regulation of renal blood flow, GFR, and mesangial matrix accumulation. Therefore, in this study it was tested whether high glucose directly modulates NO synthesis by rat mesangial cells in vitro by measuring the accumulation of nitrite, the stable metabolite of NO, in the incubation media. Raising the external glucose concentration to 33.3 mM for 24 to 72 h reduced nitrite levels in cell supernatants in a time-dependent manner to a nadir of 14 ± 3% of the amount in normal glucose media (5.6 mM) (P < 0.01). The decline in NO synthesis in high glucose media was paralleled by decreased cyclic guanosine monophosphate generation; however, there was no alteration in rat mesangial cell expression of inducible NO synthase protein. The suppressive effect of high glucose on NO production by mesangial cells was not modified by inhibition of protein kinase C (H-7), the addition of antioxidants (vitamin E or superoxide dismutase), or a pan-specific anti-transforming growth factor-a antibody. An elevated ambient glucose caused a time-dependent reduction in mesangial cell L-arginine con- tent. Addition of L-arginine (10 to 20 mM) to external media partially reversed the inhibitory effect of high glucose on mesangial cell NO production in a dose-dependent manner. The highest dose of L-arginine (20 mM) increased mesangial cell L-arginine content to comparable levels in normal and high glucose media. These results indicate that high glucose causes depletion of L-arginine in mesangial cells and compromises NO synthesis. Limitation in the metabolic precursor and other, as yet unidentified, factors act to reduce NO production by mesangial cells in the presence of an elevated ambient glucose level, a change that may play a role in the development of diabetic gbomerulosclerosis. (J Am Soc Nephrol 8: 1276-1282, 1997) Hyperglycemia is a major factor in the pathogenesis of target organ complications of diabetes including nephropathy (1). An elevated glucose concentration enhances single nephron plasma flow and GFR (2), causes oxidative injury to mesangial cells (3). and enhances the production of extracellular matrix (ECM) components by renal cells (4,5). Nitric oxide (NO) is a volatile, cell-signaling molecule that is synthesized from the guanidino nitrogen of L-arginine and dioxygen by at least three different isoforms of the enzyme NO synthase (NOS) (6). NO participates in the regulation of affer- ent arteniolar tone in the glomerular circulation (7), tubular handling of sodium and salt balance (8,9), and mesangial cell proliferation (10). Mesangial cells and renal tubular epithelial cells possess inducible NOS (iNOS) and synthesize NO in response to various cytokines (I 1,12). There are conflicting data concerning the effect of experi- Received April 18, 1996. Accepted March 17, 1997. Correspondence to Dr. Howard Trachtman. Schneider Children’s Hospital, Division of Nephrology. 269-01 76th Avenue, New Hyde Park, NY I 1040. l()46-6673/0808- I 276$03.00/0 Journal of the American Society of Nephrology Copyright U 1997 by the American Society of Nephrology mental diabetes on renal NO production. Some reports indicate increased NO synthesis (13,14), whereas others have docu- mented reduced NO production and decreased NO-dependent cyclic guanosine monophosphate (cGMP) generation by gb- meruli isolated from diabetic rats (15-17). To clarify the role of NO in the pathogenesis of diabetic nephropathy, we exam- med the effect of high glucose on NO production in cultured rat mesangial cells (RMC) by measuring the accumulation of nitrite, the stable metabolite of NO, in the incubation media. Materials and Methods Cells RMC were obtained from primary glomerular explants and used between passages 6 and 10. Using phase-contrast microscopy, the identity of mesangial cells was confirmed by their elongated, stellate, or fusiform appearance: in addition, there was no inhibitory effect of puromycin aminonucleoside or d-valine on cell growth (18). Cells were plated in 96-well plates, 25 X I0 cells/mb, to assay nitrite production, an index of NO synthesis ( 19). RMC were grown in 75-cm2 flasks for determination of L-arginine content and Western analysis of iNOS protein. Cells were incubated at 37#{176}C in a humid- ified atmosphere of 10% C02-90% air and maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 1(X) g/ml streptomycin, 100 pg/ml penicillin, and 10% fetal bovine serum.
Transcript
High Glucose Inhibits Nitric Oxide Production in Cultured
Rat Mesangial Cells
HOWARD TRACHTMAN, STEPHEN FUTTERWEIT, and DAN L. CRIMMINSDivision of Nephrology, Schneider Children ‘s Hospital, Long Island Jewish Medical Center, Long island
Campus for Albert Einstein College of Medicine, New Hyde Park, New York; and Department of Molecular
Biology and Pharmacology. and Protein and Nucleic Acid Chemistry Laboratory, Washington University
School of Medicine, St. Louis, Missouri.
Abstract. Hyperglycemia directly contributes to the develop-
ment of diabetic nephropathy. A high-serum glucose concen-
tration alters intraglomerular hemodynamics and promotes
deposition of extracellular matrix in the kidney. Nitric oxide
(NO) is a short-lived messenger molecule that participates in
the regulation of renal blood flow, GFR, and mesangial matrix
accumulation. Therefore, in this study it was tested whether
high glucose directly modulates NO synthesis by rat mesangial
cells in vitro by measuring the accumulation of nitrite, the
stable metabolite of NO, in the incubation media. Raising the
external glucose concentration to 33.3 mM for 24 to 72 h
reduced nitrite levels in cell supernatants in a time-dependent
manner to a nadir of 14 ± 3% of the amount in normal glucose
media (5.6 mM) (P < 0.01). The decline in NO synthesis in
high glucose media was paralleled by decreased cyclic
guanosine monophosphate generation; however, there was no
alteration in rat mesangial cell expression of inducible NO
synthase protein. The suppressive effect of high glucose on NO
production by mesangial cells was not modified by inhibition
of protein kinase C (H-7), the addition of antioxidants (vitamin
E or superoxide dismutase), or a pan-specific anti-transforming
growth factor-a antibody. An elevated ambient glucose caused
a time-dependent reduction in mesangial cell L-arginine con-
tent. Addition of L-arginine (10 to 20 mM) to external media
partially reversed the inhibitory effect of high glucose on
mesangial cell NO production in a dose-dependent manner.
The highest dose of L-arginine (20 mM) increased mesangial
cell L-arginine content to comparable levels in normal and high
glucose media. These results indicate that high glucose causes
depletion of L-arginine in mesangial cells and compromises
NO synthesis. Limitation in the metabolic precursor and other,
as yet unidentified, factors act to reduce NO production by
mesangial cells in the presence of an elevated ambient glucose
level, a change that may play a role in the development of
diabetic gbomerulosclerosis. (J Am Soc Nephrol 8: 1276-1282,
1997)
Hyperglycemia is a major factor in the pathogenesis of target
organ complications of diabetes including nephropathy (1). An
elevated glucose concentration enhances single nephron
plasma flow and GFR (2), causes oxidative injury to mesangial
cells (3). and enhances the production of extracellular matrix
(ECM) components by renal cells (4,5).
Nitric oxide (NO) is a volatile, cell-signaling molecule that
is synthesized from the guanidino nitrogen of L-arginine and
dioxygen by at least three different isoforms of the enzyme NO
synthase (NOS) (6). NO participates in the regulation of affer-
ent arteniolar tone in the glomerular circulation (7), tubular
handling of sodium and salt balance (8,9), and mesangial cell
proliferation (10). Mesangial cells and renal tubular epithelial
cells possess inducible NOS (iNOS) and synthesize NO in
response to various cytokines (I 1,12).
There are conflicting data concerning the effect of experi-
Received April 18, 1996. Accepted March 17, 1997.
Correspondence to Dr. Howard Trachtman. Schneider Children’s Hospital,
Division of Nephrology. 269-01 76th Avenue, New Hyde Park, NY I 1040.
l()46-6673/0808- I 276$03.00/0
Journal of the American Society of Nephrology
Copyright U 1997 by the American Society of Nephrology
mental diabetes on renal NO production. Some reports indicate
increased NO synthesis (13,14), whereas others have docu-
mented reduced NO production and decreased NO-dependent
cyclic guanosine monophosphate (cGMP) generation by gb-
meruli isolated from diabetic rats (15-17). To clarify the role
of NO in the pathogenesis of diabetic nephropathy, we exam-
med the effect of high glucose on NO production in cultured rat
mesangial cells (RMC) by measuring the accumulation of
nitrite, the stable metabolite of NO, in the incubation media.
Materials and MethodsCells
RMC were obtained from primary glomerular explants and used
between passages 6 and 10. Using phase-contrast microscopy, the
identity of mesangial cells was confirmed by their elongated, stellate,
or fusiform appearance: in addition, there was no inhibitory effect of
puromycin aminonucleoside or d-valine on cell growth (18). Cells
were plated in 96-well plates, 25 X I0� cells/mb, to assay nitrite
production, an index of NO synthesis ( 19). RMC were grown in
75-cm2 flasks for determination of L-arginine content and Western
analysis of iNOS protein. Cells were incubated at 37#{176}Cin a humid-
ified atmosphere of 10% C02-90% air and maintained in Dulbecco’s
modified Eagle’s medium (DMEM) supplemented with 1(X) �g/ml
streptomycin, 100 p�g/ml penicillin, and 10% fetal bovine serum.
High Glucose Inhibits NO Production in Cultured RMC 1277
Experimental Conditions
At 24 h after plating RMC into 96-well plates or when the cells
reached confluence in 75-cm2 flasks (usually within I wk), cells were
randomly assigned to one of the following two experimental condi-
tions: (1) Control: DMEM containing 5.6 mM glucose; (2) High
glucose: DMEM supplemented with 27.8 mM glucose to a final
concentration of 33.3 mM. Lipopolysacchanide (LPS) ( 10 pg/ml) and
interferon (IFN)-y (50 U/mI) were added to both test media because
NO production was negligible in the absence of the cytokine combi-
nation. A hyperosmolality control medium consisted of DMEM +
27.8 mM mannitol.
Measurement of Nitrite and cGMP Production
Nitrite production was measured using the Griess assay (19).
Briefly, 125 �tl of a solution containing 1% sulfanilamide, 0.1%
naphthylethylene diamine dihydnochloride, and 2.5% phosphoric acid
was added to I 25 j.tl of conditioned media. Samples were incubated at
25#{176}Cfor 10 mm, and absorbance was measured at 550 nm. Nitrite
production was normalized to the number of viable cells and ex-
pressed as a percentage of the value in control media containing a
normal glucose concentration. RMC were grown in 25-cm2 flasks,
and cGMP in the supennatants was measured by enzyme immunoas-
say after acetylation. using a commercially available kit (Cayman
Chemical Co., Ann Arbor, MI).
Cell Proliferation Assay
A colonimetnic method was used to determine the number of viable
RMC in these experiments. A total of 100 jd of phenazine methosul-
fate was added to 2 ml of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxy-
methoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium. inner salt in the
dark. Twenty-five microliters of this mixture was added to each wellin 96-well plates. Plates were wrapped in foil and incubated for I h at
37#{176}Cin the 10% CO,-90% air atmosphere. Absorbance was read at
490 nm in samples and solution blanks.
HPLC Assay for L-Argimne
After incubation in normal and high glucose media that did not
contain LPS and IFN-y, cell monolayers were rinsed twice with
phosphate-buffered saline (PBS). RMC were removed, and the sus-
pension was spun at 10(X) rpm for 5 mm to sediment the cells. They
were resuspended in 500 �l of PBS, and then 1000 �l of 100%
methanol was added. Samples were spun at 10,000 X g for 10 mm,
and the cleared supernatants were analyzed by HPLC.
Vacuum-dried extracts were derivatized with 6-aminoquinolyl-N-
hydroxysuccinimidyl carbamate, using the Waters kit (Millipore, Mil-
fond, MA) with slight modifications (20). The most important change
was reducing the final volume of derivatized sample from 100 to 50�l comprising 10 pJ of 20 mM HCI, 30 �l of borate buffer, and 10 �l
of 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate reagent. A
maximum of 20 M1 of the denivatized sample can be injected onto the
HPLC column without obliterating the first one-third to one-half of
the chromatogram. Therefore, this modification allows up to 40% of
the sample to be analyzed. Mobile phase and gradient development
were adapted from Liu (2 1 ) for ultraviolet detection at 248 nm.
Calibration was performed with 250-pmol injections of standard H
(Pierce Chemical Co., Rockford. IL), and a-amino butynic acid was
the internal standard. Day-to-day system performance was reproduc-
ible to <4% variation.
Western Analysis
RMC were harvested in PBS and centrifuged at 1000 rpm for 5 mm
to sediment cells. The pellet was dissolved in 100 �l of lysis buffer
(50 mM Tnis/HC1, pH 7.6, 100 mM NaCl, 2 mM ethylenediami-
netetra-acetic acid, 2 mM ethyleneglycol-bis(�3-aminoethyI ether)-
N,N’-tetra-acetic acid, 1 mM dithiothreitol, I mM phenylmethylsul-
fonyl fluoride, and 1% Triton X-l00). The suspension was freeze-
thawed three times, and equal aliquots ofthe lysates (25 p�g of protein)
were loaded onto a 7.5% acrylamide-sodium dodecyl sulfate gel. Gels
were run at 200 V and 70 mA (milliamps) for 45 mm, and protein was
then transferred from the gel onto a nitroceblulose membrane electro-
phonetically. After blocking the membrane with buffer containing
0.25% gelatin and 0.05% Tween 20, the membrane was exposed to a
primary murine monoclonal antibody to iNOS (Transduction Labo-
ratonies, Lexington, KY) followed by a secondary antibody (horse-
radish peroxidase-binked anti-mouse IgG). Immunoblots of the iNOS
protein were visualized with enhanced chemibuminescence (Amen-
sham, Arlington Heights. IL).
Protein Assay
The protein contents of the cell homogenates were determinedusing a Coomassie blue reagent (BioRad, Richmond, CA).
Reagents
A pan-specific. polyclonal neutralizing anti-transforming growth
factor (TGF)-� antibody, reactive with TGF-� I ,TGF-�2, TGF-� I .2,TGF-/33, and TGF-�5, was purchased from R&D Systems (Minne-
apolis, MN). All other reagents were purchased from Sigma Chemical
Co. (St. Louis, MO).
Statistical Analyses
Results are presented as mean ± SEM. The means between the
experimental groups were compared using ANOVA; post hoc inter-
group comparisons were made using the Bonferroni correction.
ResultsNO Production
There was no measurable NO production by RMC, indicated
by undetectable nitrite levels, in media that did not contain
IFN-y and LPS. In the presence of the cytokine combination,
exposure of RMC to 33.3 mM glucose led to a time-dependent
inhibition of NO production (Figure 1). Thus, after 24 h of
incubation in high glucose media, nitrite accumulation (0.5 to
I .0 nmol/well) per unit number of viable cells was 88 ± 3% of
the amount in normal glucose media (P < 0.05). More pro-
longed exposure to high glucose conditions resulted in a pro-
gressively greater effect, i.e., a decline to 52 ± 4 and 14 ± 3%
of the nitrite levels (1 to 4 nmol/well) observed in control
media after 48 and 72 h, respectively (P < 0.01). Cell viability
was consistently greater in the high glucose media at 24, 48,
and 72 h, i.e., 108 ± 6, 152 ± 8, and 186 ± 11% of the value
in normal glucose media, respectively (P < 0.05). Thus, the
effect of high glucose on nitrite levels was the result of a
combined effect on NO production and improved cell viability.
NO production and cell viability were comparable in the dif-
ferent cell lines and passages used in these experiments.
To determine whether high glucose altered the partitioning
of NO metabolites between nitrate and nitrite, the nitrite level
-J0
I-z00
U,-Jw>U-J
UI-
1-
z
0
,;;- 1 00
0
a,
.n 75a>
L
I �#{176}
� 25
0
E%E.o
*
**
I a
a The results are mean ± SEM. it = 4 pained experiments foreach test condition. Nitrite levels (nmol/well) were measured in
RMC supernatants after 48 h of exposure to test media withindicated additive. Samples were analyzed before incubation with
Escherichia co/i reductase (NO, ) or after incubation withEsc/ierichia co!i reductase (NO, + NO� ). RMC, rat mesangialcells.
1278 Journal of the American Society of Nephrology
24H 48H 72H
Figure 1. Effect of high glucose on rat mesangial cell (RMC) nitrite
levels (nmol/unit number of viable cells). Results are expressed as a
percentage of paired normal glucose media (n = 10 for each time
point). */) < 0.05 versus pained normal glucose media. **� < 0.01versus paired normal glucose media.
in conditioned RMC media was determined in normal and high
glucose media, before and after incubation of samples with
Escher#{252}hia co/i reductase ( I 9). The difference between these
two values represents the amount of nitrate in the supernatant.
The ratio of NO,/NO, + NO1 was the same in normal and
high glucose media and was unaffected by supplementation
with 20 mM L-arginine (Table 1).
The inhibition of NO production by RMC during exposure
to high glucose media was paralleled by a reduction in cGMP
generation. Thus, after 24 h, cGMP accumulation in the media
(expressed as pmollnil supernatant per unit number of viable
cells) declined from 3. 1 5 ± 0.30 to 2.9 1 ± 0.43. After 48 h of
incubation in high glucose media, cGMP accumulation was
further reduced by 47%, from 2.71 ± 0.29 in normal glucose
media to 1 .44 ± 0. 13 in high glucose media (ii = 3) (P <
0.05).
Hyperosmolal media containing 27.8 mM mannitol did not
alter RMC NO production (data not shown). There was no
evidence of cytotoxicity as a result of incubation with high
Table 1. N0/NO + NO� ratio in cultured RMC: effect
of normal and high glucose media�’
Group Norma 1 Glucose High Glucose
No additives 0.38 ± 0.05 0.41 ± 0.03
+2 0 mM L-arginine 0.34 ± 0.07 0.35 ± 0.04
glucose media, based on the amount of lactate dehydrogenase
detectable in the conditioned media (data not shown).
The diabetic state is associated with activation of protein
kinase C (PKC) (22). This enzyme can phosphorylate the
neuronal and endothelial isoforms of NOS and reduce their
activity (23,24). Therefore, we examined whether addition of
H-7 (50 �sM), an inhibitor of PKC, could normalize NO syn-
thesis by RMC incubated in high glucose media for 48 h.
Instead, we found that H-7 caused a profound reduction in NO
synthesis per unit number of viable cells to 9 ± 3 and 7 ± 2%
of the control value under normal glucose and high glucose
conditions, respectively (Table 2). The inhibitory effect also
was observed after 24 h of exposure to H-7. Despite the lack of
specificity of H-7 as a PKC inhibitor, the data do not support
a role of PKC activation in reducing NO production by RMC
in high glucose media.
High glucose stimulates gene expression and release of
TGF-/3 by renal cells (25). This cytokine destabilizes iNOS
mRNA, reduces translational efficiency, and enhances iNOS
protein degradation; these changes result in decreased NO
synthesis (26). Therefore, we tested whether a pan-specific,
polyclonal anti-TGF-3 antibody (35 to 200 j�g/ml) would
restore mesangial cell NO synthesis to normal. In fact, there
was no significant effect of anti-TGF-�3 antibody on NO syn-
thesis by RMC that were maintained in normal glucose media
for 48 h. In contrast, RMC exposed to a high, ambient glucose
level for 48 h manifested a dose-dependent inhibition of NO
synthesis by anti-TGF-f3 antibody (Table 2). Nonimmune IgG
(200 pg/ml) had no effect on RMC NO production (data not
shown).
An elevated, ambient glucose exerts a direct oxidant stress
on cultured RMC (3). We examined the effect of the antioxi-
dants vitamin E (100 p�M) and superoxide dismutase (250
U/mb) on NO synthesis by RMC. These agents failed to reverse
Table 2. Nitrite levels in cultured RMC: effect of H-7,
anti-TGF-3 antibody, and antioxidantsa
Group Normal Glucose Hig h Glucose
No additives I 00 ( 12)” 48 ± 4 (12)”
H-7 (50 �M) 9 ± 3 (6)” 7 ± 2 (6)”
Anti-TGF-� antibody
35 p.g/ml 92 (2) 43 (2)
100 �tg/ml 102 (2) 31 (2)
200�g/ml 78(2) 6(2)
Vitamin E ( 100 �M) 90 ± 5 (4) 49 ± 3 (4)�)
SOD (250 U/ml) 66 ± 5 (4)” 35 ± 5 (4)b.c
a The results are mean ± SEM. The numbers in parentheses
indicate number of paired samples. Nitrite levels (nmol/unit numberof viable cells) after 48 h of exposure to the test media withindicated additive is expressed as a percentage of control normalglucose medium containing interferon-y and lipopolysacchanide andno further additives. TGF, transforming growth factor; SOD,superoxide dismutase.
b p < 0.01 versus normal glucose, no additives.
C p < 0.05 versus normal glucose plus SOD.
z�-I- Cl
zo
0a)
<a
.L�0
00 -
75
50
25
V
I*
I24H 48H 72H
the suppressed NO synthesis by RMC incubated in high glu-
cose media to the bevel observed in normal glucose conditions
(Table 2).
Addition of L-arginine caused a dose-dependent increase in
NO synthesis by RMC maintained in normal glucose media for
48 h. Thus, there was a 125 ± 7 and 197 ± 20% increase in
NO synthesis after addition of 10 and 20 mM L-arginine,
respectively, to the external media (P < 0.01) (Figure 2). In
contrast, in the presence of an elevated, external glucose con-
centration, 10 mM L-arginine had no effect on RMC NO
synthesis, whereas 20 mM L-argrnine enhanced NO production
to I 17 ± 2 1% of the value in control normal glucose media
(P < 0.01) (Figure 2). The increased NO synthesis by RMC
exposed to supplemental L-arginine was reduced nearly 80%
after the addition of NG�nitro�L�arginine methyl ester
(L-NAME) (I mM) to normal and high glucose media.
HPLC Analysis of RMC Amino Acid Content
In these experiments, LPS and IFN-y were not present in the
test media. There was no significant change in L-arginine
content in RMC after 24 h of exposure to high glucose (n = 3)
(Figure 3). L-Arginine content varied widely in RMC that were
exposed to high glucose (range, 0.2 to 2.5 nmol/unit number of
viable cells) or normal glucose media (range, 0.5 to 5.6 nmoll
unit number of viable cells) for 48 to 72 h because of the use
of RMC of different passage. Therefore, we expressed
L-arginine content in cells exposed to high glucose as a per-
centage of the paired sample maintained in normal glucose
media. There was a time-dependent decrease in RMC L-argi-
nine content by 34 and 68% after 48 and 72 h exposure to high
glucose, respectively (P < 0.05) (n 3 for each time point).
The reduction in RMC L-arginine content paralleled the decline
in NO production observed in the presence of an elevated,
ambient glucose and the two cytokines (Figures 1 and 3).
_J
0�!�
� a,2000�
>tI) 150_J I-
U �>-0WE-J �iooU
1-�
I- j 50
0
Figure 2. Effect of L-arginine on RMC nitrite levels after 48 h
(nmol/unit number of viable cells). Results are expressed as a per-
centage of paired normal glucose media (it = I 0 for each time point).
Open bars, normal glucose; hatched bars. high glucose. *� < 0.01
versus control normal glucose media, no added L-arginine. **�P <
0.01 versus high glucose media, no added L-argiflifle.
0
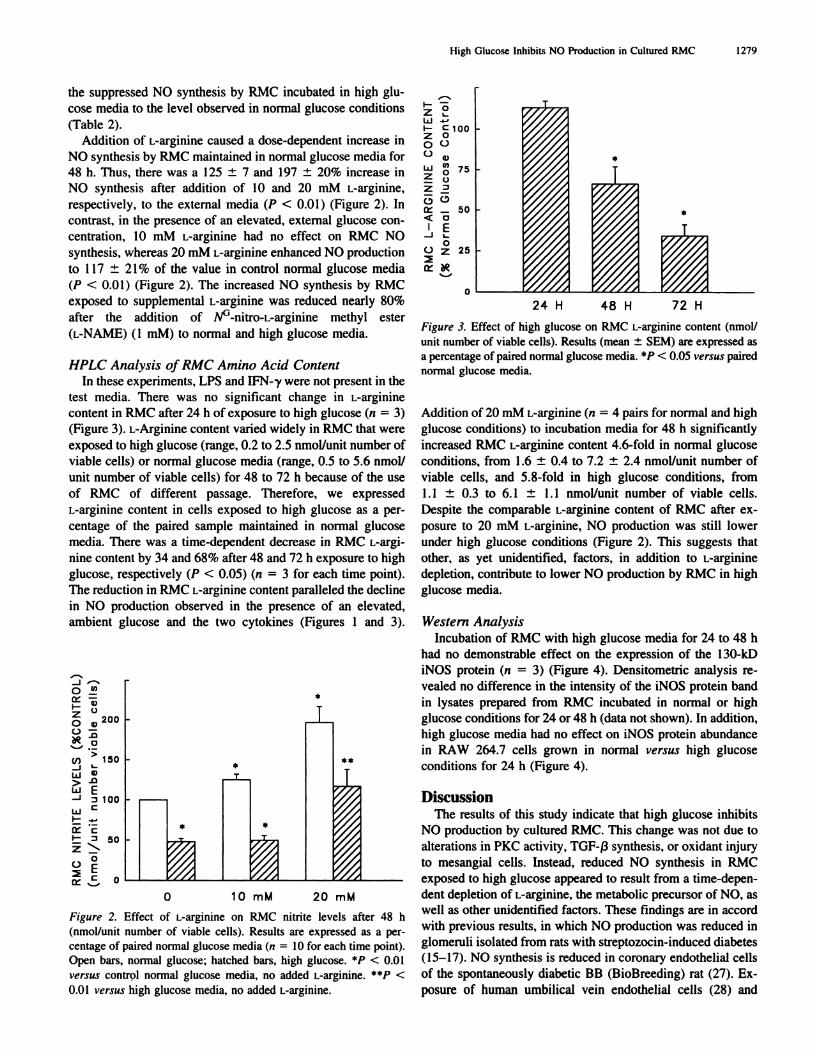
Figure 3. Effect of high glucose on RMC L-arginine content (nmol/
unit number of viable cells). Results (mean ± SEM) are expressed asa percentage of paired normal glucose media. *P < 0.05 versus paired
normal gbucose media.
Addition of 20 mM L-arginine (ii = 4 pairs for normal and high
glucose conditions) to incubation media for 48 h significantly
increased RMC L-arginine content 4.6-fold in normal glucose
conditions, from I .6 ± 0.4 to 7.2 ± 2.4 nmol/unit number of
viable cells, and 5.8-fold in high glucose conditions, from
1.1 ± 0.3 to 6.1 ± 1.1 nmol/unit number of viable cells.
Despite the comparable L-arginine content of RMC after ex-
posure to 20 mM L-arginine, NO production was still lower
under high glucose conditions (Figure 2). This suggests that
other, as yet unidentified, factors, in addition to L-arginine
depletion. contribute to lower NO production by RMC in high
glucose media.
Western Analysis
Incubation of RMC with high glucose media for 24 to 48 h
had no demonstrable effect on the expression of the 1 30-kD
iNOS protein (n = 3) (Figure 4). Densitometric analysis re-
vealed no difference in the intensity of the iNOS protein band
in lysates prepared from RMC incubated in normal or high
glucose conditions for 24 or 48 h (data not shown). In addition,
high glucose media had no effect on iNOS protein abundance
in RAW 264.7 cells grown in normal versus high glucose
conditions for 24 h (Figure 4).
DiscussionThe results of this study indicate that high glucose inhibits
NO production by cultured RMC. This change was not due to
alterations in PKC activity, TGF-f3 synthesis, or oxidant injury
to mesangial cells. Instead, reduced NO synthesis in RMC
exposed to high glucose appeared to result from a time-depen-
dent depletion of L-arginine, the metabolic precursor of NO, as
well as other unidentified factors. These findings are in accord
with previous results, in which NO production was reduced in
gbomeruli isolated from rats with streptozocin-induced diabetes
(15-17). NO synthesis is reduced in coronary endothelial cells
of the spontaneously diabetic BB (BioBreeding) rat (27). Ex-
posure of human umbilical vein endothelial cells (28) and
**
0 10mM 20mM
High Glucose Inhibits NO Production in Cultured RMC 1279
1280 Journal of the American Society of Nephrology
C I II lilly vvIcFigure 4. Effect of exposure to high glucose for 24 h on inducible NO synthase (iNOS) protein content assayed by Western analysis. Lane C.
iNOS control specimen; lane I. normal glucose, 24 h: lane II, high glucose. 24 h: lane III. normal glucose, 48 h: lane IV, high glucose. 48 h:
lane V. RAW 264.7 cells. normal glucose, 24 h; lane VI, RAW 264.7 cells, high glucose, 24 h. Arrow indicates the location of iNOS protein
at 130 kD.
SH-SY5Y human neuroblastonla cells to high glucose (29)
inhibits NO production. Finally. a recent study indicates that
high glucose downregulates NO production and iNOS expres-
sion in cultured rat aortic vascular smooth muscle cells, an
effect that was reversed by addition of a PKC inhibitor (30).
However, high glucose does not uniformly inhibit NO produc-
the synthesis of extracellular matrix proteins in cultured rat
mesangial cells. Biochein Bioph’�’s Res Commun 207: 120-125,
1995
40. Trachtman H, Futterweit 5, Garg P. Reddy K, Singhal PC: Nitricoxide stimulates the activity of a 72-kD neutral matrix metallo-proteinase in cultured rat mesangial cells. Biochem Biophys Res