Histone H1 Recruitment by CHD8 Is Essential for Suppression of theWnt–�-Catenin Signaling Pathway
Masaaki Nishiyama,a,b Arthur I. Skoultchi,c and Keiichi I. Nakayamaa,b
Department of Molecular and Cellular Biology, Medical Institute of Bioregulation, Kyushu University, Fukuoka, Japana; CREST, Japan Science and Technology Agency,Saitama, Japanb; and Department of Cell Biology, Albert Einstein College of Medicine, Bronx, NY, USAc
Members of the chromodomain helicase DNA-binding (CHD) family of proteins are thought to regulate gene expression.Among mammalian CHD proteins, CHD8 was originally isolated as a negative regulator of the Wnt–�-catenin signaling path-way that binds directly to �-catenin and suppresses its transactivation activity. The mechanism by which CHD8 inhibits�-catenin-dependent transcription has been unclear, however. Here we show that CHD8 promotes the association of �-cateninand histone H1, with formation of the trimeric complex on chromatin being required for inhibition of �-catenin-dependenttransactivation. A CHD8 mutant that lacks the histone H1 binding domain did not show such inhibitory activity, indicating thathistone H1 recruitment is essential for the inhibitory effect of CHD8. Furthermore, either depletion of histone H1 or expressionof a dominant negative mutant of this protein resulted in enhancement of the response to Wnt signaling. These observationsreveal a new mode of regulation of the Wnt signaling pathway by CHD8, which counteracts �-catenin function through recruit-ment of histone H1 to Wnt target genes. Given that CHD8 is expressed predominantly during embryogenesis, it may thus con-tribute to setting a threshold for responsiveness to Wnt signaling that operates in a development-dependent manner.
The Wnt–�-catenin signaling pathway plays key roles in devel-opment, specification of cell fate, and adult stem cell prolifer-
ation (2, 21, 27, 44). Abnormal activation of this pathway is asso-ciated with various human cancers, including colorectal cancerand head and neck squamous cell carcinoma. Many genes associ-ated with tumor growth, including those for c-Myc, matrix met-alloproteinases, and cyclin D1, have been identified as Wnt targetgenes, with �-catenin functioning as a transcriptional coactivatorat such genes (11, 16, 22, 35). In the absence of �-catenin binding,the promoters of Wnt target genes are occupied by Tcf/Lef familyproteins, the corepressor Groucho (also known as TLE1), andhistone deacetylase 1 (3, 5, 9, 34). Wnt signaling results in theaccumulation of �-catenin in the cytosol and nucleus, and nuclear�-catenin displaces Groucho from Tcf by binding Tcf at the pro-moters of Wnt target genes. �-Catenin then recruits chromatin-remodeling complexes or other transcriptional coactivators tostimulate gene transcription (2, 12, 21, 27, 43, 44). Several mem-bers of the Snf2 superfamily of ATP-dependent chromatin-remodeling enzymes, including BRG1 (1), Snf2H, and p400 (39),were recently found to be recruited by �-catenin in the inductionof target gene transcription. Although these findings suggest thatATP-dependent chromatin remodeling plays a fundamental rolein the regulation of �-catenin-dependent transcription, it remainsunclear how such enzymes contribute to the regulation of chro-matin at Wnt target genes.
Members of the chromodomain helicase DNA-binding(CHD) family of proteins also belong to the Snf2 superfamily ofATP-dependent chromatin remodelers. Among the nine mam-malian members of this family, CHD1 is thought to play an im-portant role in gene transcription. The tandem chromodomainsof human CHD1 thus specifically recognize and bind to the tri-methylated form of lysine 4 of histone H3 (H3K4me3), a hallmarkof actively transcribed chromatin (7), and mediate the recruit-ment of transcriptional initiation and pre-mRNA splicing factors(40). In mammals, CHD3 and CHD4 are subunits of nucleosome-remodeling and histone deacetylase (NURD) complexes, which
contain histone deacetylases and function as transcriptional re-pressors (47). Mutations in CHD7 result in CHARGE syndrome,a multiple-malformation syndrome in humans for which morethan 40 alleles have been defined (42). Chd7�/� mice recapitulateseveral aspects of this human disease, including inner-ear vestib-ular dysfunction (13). Molecular studies suggest that CHD7 con-tributes to transcriptional activation of tissue-specific genes dur-ing differentiation (38). However, most of the biological functionsmediated by members of the CHD family remain to be elucidated.
CHD8 exists in two isoforms, short (CHD8S) and long(CHD8L), that are generated as a result of alternative mRNA splic-ing. CHD8S (also known as duplin) was originally isolated as anegative regulator of the Wnt–�-catenin signaling pathway (36)and binds directly to the Armadillo repeats of �-catenin (36, 41).The COOH-terminal region of full-length CHD8 (CHD8L) inter-acts with the insulator-binding protein CTCF, with this interac-tion being important for insulator activity (14). CHD8 has alsobeen implicated as a positive or negative transcriptional regulatorof various genes (19, 32, 33, 45, 46). We previously showed thatChd8�/� mice die early during embryogenesis, manifesting wide-spread apoptosis (28), whereas additional deletion of the tumorsuppressor gene p53 ameliorated this developmental arrest (29).Both isoforms of CHD8 bind to p53 and suppress its transactiva-tion activity by recruiting histone H1, with formation of a p53-CHD8-histone H1 trimeric complex on chromatin being requiredfor inhibition of p53-dependent transactivation and apoptosis.These observations led us to examine whether such histone H1recruitment by CHD8 might also contribute to the regulation of
other genes such as Wnt target genes. We now show that CHD8mediates the recruitment of histone H1 to Wnt target genes, re-sulting in suppression of the expression of these genes induced byactivation of Wnt–�-catenin signaling. Our results thus suggestthat CHD8 may target multiple transcriptional activators, includ-ing �-catenin and p53, for suppression of transcriptional activitythrough recruitment of histone H1.
MATERIALS AND METHODSAntibodies and reagents. Antibodies to �-catenin and to Hsp90 wereobtained from BD Biosciences (San Jose, CA); those to FLAG (M2 or M5)
were from Sigma (St. Louis, MO); those to the hemagglutinin (HA)epitope (HA.11) were from Covance (Princeton, NJ); those to Myc (9E10)were from Roche (Indianapolis, IN); those to glutathione S-transferase(GST) were from MBL (Nagoya, Japan); those to histone H1 were fromAbcam (Cambridge, United Kingdom); and those to CHD8 were gener-ated in rabbits by injection with a recombinant fragment of mouse CHD8(residues 484 to 670). Recombinant human �-catenin was obtained fromMillipore (Billerica, MA), and recombinant mouse Wnt3a was obtainedfrom R&D Systems (Minneapolis, MN). Alexa Fluor 488-conjugated goatantibodies to mouse immunoglobulin G (IgG) were obtained from Mo-lecular Probes-Invitrogen (Carlsbad, CA).
FIG 1 CHD8 interacts with and inhibits transactivation by �-catenin. (A) Whole-cell lysates of HEK293T cells were subjected to immunoprecipitation (IP) withmouse antibodies to �-catenin or with control mouse IgG. The resulting precipitates, as well as 1% of the original cell lysates (input), were subjected toimmunoblot analysis (IB) with antibodies to CHD8 or to �-catenin. (B and C) HEK293T cells were incubated in the absence (�) or presence (�) of LiCl (20 mM)(B) or Wnt3a (20 ng/ml) (C) for 4 h and were then subjected to immunoprecipitation with rabbit antibodies to CHD8 or with control rabbit IgG. The resultingprecipitates, as well as 1% of the original cell lysates (input), were subjected to immunoblot analysis with antibodies to �-catenin, to CHD8, or to Hsp90 (loadingand negative control). (D and E) HeLa cells infected with retroviruses encoding FLAG-tagged forms of CHD8S or CHD8L (or with the empty retrovirus) wereincubated in the absence (�) or presence of LiCl or Wnt3a for 4 h. The cells were then subjected to ChIP with antibodies to FLAG, and the precipitated DNA wasquantitated by real-time PCR analysis with primers specific for Wnt-responsive elements (WRE) or a control region (CR) of the axin2 (D) or c-Myc (E) genepromoter or for the p27 gene promoter. The cells were also subjected to immunoblot analysis with antibodies to CHD8 or to Hsp90 (D). (F and G) HeLa cellsinfected with a retrovirus for CHD8S or the empty vector were incubated with LiCl or Wnt3a for 4 h and were then subjected to RT and real-time PCR analysisof axin2 (F) and c-Myc (G) mRNAs. All quantitative data are means � standard deviations (SD) from three independent experiments.
Plasmids. Complementary DNAs encoding wild-type or mutantforms of mouse CHD8S tagged with the FLAG or Myc epitope at the NH2
terminus were subcloned into pcDNA3 (Invitrogen) with the use of theGateway vector conversion system (Invitrogen); those encoding wild-typeor mutant forms of mouse histone H1c with three copies of the FLAGepitope at the NH2 terminus were also subcloned into pcDNA3. A cDNAfor mouse �-catenin tagged at its NH2 terminus with the HA epitope was
subcloned into pCGN. His6-tagged proteins were expressed in Escherichiacoli strain BL21(DE3)pLys(S) (Merck, Darmstadt, Germany).
Cell culture, immunoprecipitation, and immunoblot analysis.HEK293T and HeLa cells were cultured under an atmosphere of 5% CO2
at 37°C in Dulbecco’s modified Eagle’s medium (DMEM) (Wako, Osaka,Japan) supplemented with 10% fetal bovine serum (Invitrogen). NIH 3T3cells were cultured under the same conditions in DMEM supplemented
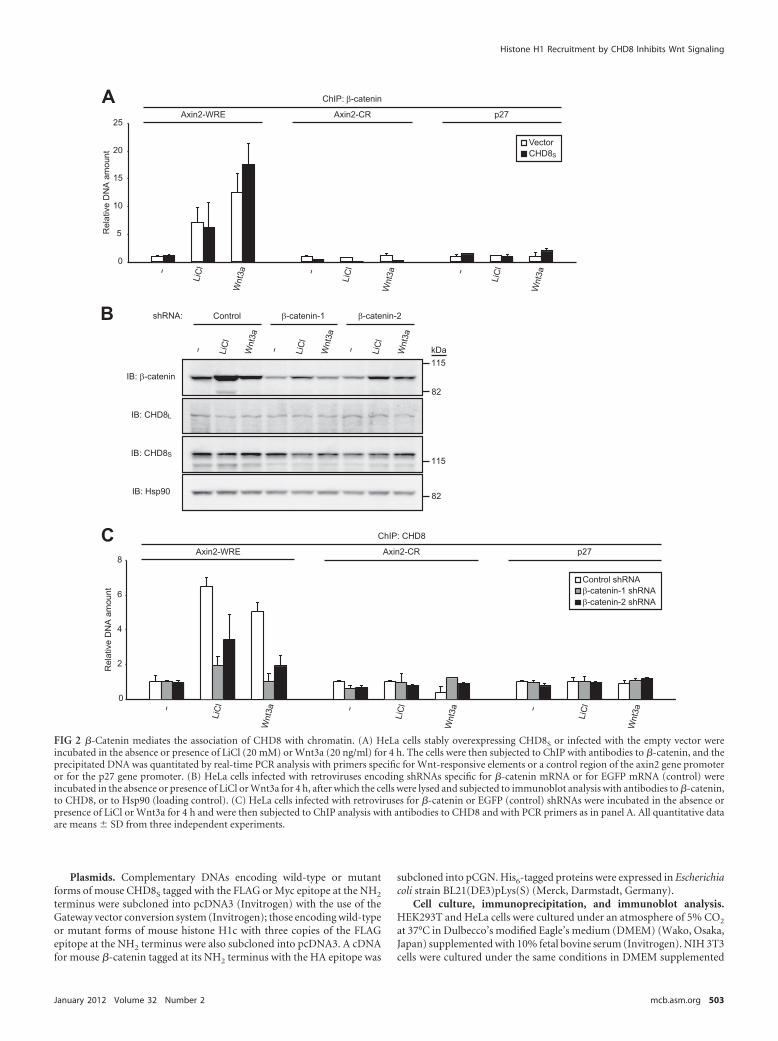
FIG 2 �-Catenin mediates the association of CHD8 with chromatin. (A) HeLa cells stably overexpressing CHD8S or infected with the empty vector wereincubated in the absence or presence of LiCl (20 mM) or Wnt3a (20 ng/ml) for 4 h. The cells were then subjected to ChIP with antibodies to �-catenin, and theprecipitated DNA was quantitated by real-time PCR analysis with primers specific for Wnt-responsive elements or a control region of the axin2 gene promoteror for the p27 gene promoter. (B) HeLa cells infected with retroviruses encoding shRNAs specific for �-catenin mRNA or for EGFP mRNA (control) wereincubated in the absence or presence of LiCl or Wnt3a for 4 h, after which the cells were lysed and subjected to immunoblot analysis with antibodies to �-catenin,to CHD8, or to Hsp90 (loading control). (C) HeLa cells infected with retroviruses for �-catenin or EGFP (control) shRNAs were incubated in the absence orpresence of LiCl or Wnt3a for 4 h and were then subjected to ChIP analysis with antibodies to CHD8 and with PCR primers as in panel A. All quantitative dataare means � SD from three independent experiments.
Histone H1 Recruitment by CHD8 Inhibits Wnt Signaling
with 10% bovine serum (Invitrogen). Embryonic stem (ES) cells werecultured as described previously (25, 26). In some experiments, HeLa orNIH 3T3 cells were cultured in the presence of LiCl (20 mM) or Wnt3a (20ng/ml) for 4 h and ES cells were incubated with LiCl (2.5 mM) or Wnt3a(2.5 ng/ml) for 4 h. Cell lysis, immunoprecipitation, and immunoblotanalysis were performed as described previously (15).
Retrovirus expression system. Complementary DNAs encodingwild-type or mutant forms of mouse CHD8S, human CHD8L, or mousehistone H1c tagged with the FLAG epitope at the NH2 terminus weresubcloned into pMX-puro (kindly provided by T. Kitamura, University ofTokyo, Japan) with the use of the Gateway vector conversion system. Theresulting vectors were used to transfect Plat E packaging cells and therebyto generate recombinant retroviruses (23). NIH 3T3 cells as well as HeLacells stably expressing the mouse ecotropic retrovirus receptor (mCAT-1)were infected with retroviruses produced by Plat E cells and were thencultured in the presence of puromycin (Sigma) at 10 �g/ml.
RNA interference. The pMX-puro II vector was constructed by dele-tion of the U3 portion of the 3= long terminal repeat of pMX-puro. Themouse U6 gene promoter, followed by DNA corresponding to a shorthairpin RNA (shRNA) sequence, was subcloned into the NotI and XhoIsites of pMX-puro II, yielding pMX-puro II-U6/siRNA. The DNA for theshRNA encoded a 21-nucleotide hairpin sequence specific to the mRNAtarget, with a loop sequence (-TTCAAGAGA-) separating the two com-plementary domains, and contained a tract of five T nucleotides to termi-nate transcription. The hairpin sequences specific for human �-catenin(�-catenin-1 and �-catenin-2), human CHD8, and enhanced green fluo-rescent protein (EGFP) mRNAs (Clontech, Mountain View, CA) corre-sponded to nucleotides 1774 to 1794 (�-catenin-1), 501 to 521 (�-catenin-2), 138 to 158 (CHD8), and 126 to 146 (EGFP) of the respectivecoding regions. The resulting vectors were used to transfect Plat E cellsand thereby to generate recombinant retroviruses.
RT-PCR. Total RNA (1 �g) isolated from HeLa, NIH 3T3, or ES cellswith the use of Isogen (Nippon Gene, Tokyo, Japan) was subjected toreverse transcription (RT) with ReverTra Ace � (Toyobo, Osaka, Japan).The resulting cDNA was subjected to PCR with Power SYBR green PCRmaster mix in an ABI-Prism 7000 sequence detection system (AppliedBiosystems, Foster City, CA). The relative amounts of each target mRNAwere calculated as described previously (31). The primer sequences forRT-PCR analysis (sense and antisense, respectively) are 5=-CTGGCTTTGGTGAACTGTTG-3= and 5=-AGTTGCTCACAGCCAAGACA-3= forhAxin-2, 5=-ATCTGTATGTCCTGTCTGCCAGCG-3= and 5=-TGTCCTGATTCCCAAGTCAAGCAC-3= for mAxin-2, 5=-CTACCCTCTCAACGACAGCA-3= and 5=-AGAGCAGAGAATCCGAGGAC-3= for hc-Myc, 5=-TCCTGACGACGAGACCTTCATCAAG-3= and 5=-TGAGAAACCGCTCCACATACAGTCC-3= for mc-Myc, 5=-GCAAATTCCATGGCACCGT-3= and 5=-TCGCCCCACTTGATTTTGG-3= for hGAPDH, and 5=-GGACCCGAGAAGACCTCCTT-3= and 5=-GCACATCACTCAGAATTTCAATGG-3= for mRplp0. Human and mouse genes are indicated by h and m,respectively.
ChIP. Chromatin immunoprecipitation (ChIP) assays were per-formed with a ChIP assay kit (Millipore) and 1 � 106 cells for each reac-tion. Precipitated DNA was quantitated by real-time PCR analysis asdescribed previously (31). The primer sequences for ChIP analysis(sense and antisense, respectively) are 5=-CTGGAGCCGGCTGCGCTT
TGATAA-3= and 5=-CGGCCCCGAAATCCATCGCTCTGA-3= forhAxin-2 (Wnt-responsive element [WRE]), 5=-CTGGAGCCGGCTGCGCTTTGATAA-3= and 5=-TGGCCCCGAAATCCATCGCGAACGG-3= formAxin-2 (WRE), 5=-CTGGCTTTGGTGAACTGTTG-3= and 5=-AGTTGCTCACAGCCAAGACA-3= for hAxin-2 (control region [CR]), 5=-ATCTGTATGTCCTGTCTGCCAGCG-3= and 5=-TGTCCTGATTCCCAAGTCAAGCAC-3= for mAxin-2 (CR), 5=-GCGGGTTACATACAGTGCACTTCA-3= and 5=-TGGAAATGCGGTCATGCACAAA-3= for hc-Myc (WRE),5=-CTACCCTCTCAACGACAGCA-3= and 5=-AGAGCAGAGAATCCGAGGAC-3= for hc-Myc (CR), and 5=-CCGCCGCCGCAACCAATGGAT-3= and 5=-GGAGTCGCAGAGCCGTGAGCA-3= for hp27.
RESULTSBoth forms of CHD8 interact with �-catenin on chromatin in aWnt signaling-dependent manner. Forced expression of CHD8S
or CHD8L in mammalian cells revealed that both isoforms asso-ciated with �-catenin (36, 41). We examined whether endogenousCHD8S and CHD8L also interact with endogenous �-catenin inHEK293T cells. Immunoblot analysis revealed that immunopre-cipitates prepared with antibodies to �-catenin contained bothCHD8S and CHD8L (Fig. 1A). In addition, reciprocal analysis alsoshowed that endogenous �-catenin was present in immunopre-cipitates prepared with antibodies to CHD8 (Fig. 1B and C). Bothendogenous CHD8S and CHD8L thus specifically interacted withendogenous �-catenin in HEK293T cells, and such associationwas markedly increased in cells that were exposed to LiCl (Fig. 1B)or to Wnt3a (Fig. 1C) as activators of the Wnt–�-catenin signalingpathway. Given that CHD8 is localized in the nucleus (29), thebinding between CHD8 and �-catenin may increase with the ac-cumulation and nuclear translocation of �-catenin induced byactivation of Wnt signaling.
We next examined whether CHD8 might be recruited to thepromoter regions of Wnt target genes with the use of ChIP. ChIPwith antibodies to the FLAG epitope revealed that FLAG-taggedforms of both CHD8S and CHD8L were specifically associatedwith Wnt-responsive elements of the axin2 gene promoter inHeLa cells only when the cells were exposed to LiCl or Wnt3a (Fig.1D). Similar results were obtained with the promoter of the c-Mycgene (Fig. 1E). These data thus suggested that both CHD8S andCHD8L are recruited to the promoters of Wnt target genes in aWnt signaling-dependent manner and that CHD8 and �-cateninmay interact with each other but are not located at such genepromoters in unstimulated cells.
We next examined whether CHD8 might affect transcriptionalactivation by �-catenin in cancer cells. Forced expression ofCHD8S in HeLa cells markedly inhibited the LiCl- or Wnt3a-induced upregulation of mRNAs derived from the Wnt targetgenes for axin2 (Fig. 1F) and c-Myc (Fig. 1G). Collectively, theseresults thus suggested that both CHD8S and CHD8L interact with�-catenin and inhibit its transactivation activity on chromatin.
FIG 3 CHD8 recruits histone H1 to the promoters of Wnt target genes. (A) Recombinant GST–�-catenin, HA-CHD8S, and FLAG-histone H1 (HH1) weremixed as indicated and then subjected to immunoprecipitation with antibodies to FLAG. The resulting precipitates, as well as 10% of the original bindingmixtures (input), were subjected to immunoblot analysis with antibodies to GST, to HA, or to FLAG. (B) HEK293T cells expressing HA–�-catenin, Myc-CHD8S,Myc-CHD8S�H1, or 3�FLAG-histone H1 as indicated were subjected to immunoprecipitation with antibodies to FLAG. The resulting precipitates, as well as 3%of the original cell lysates (input), were subjected to immunoblot analysis with antibodies to HA, to Myc, or to FLAG. (C and D) HeLa cells stably overexpressingCHD8S or infected with the empty vector were incubated in the absence or presence of LiCl (20 mM) or Wnt3a (20 ng/ml) for 4 h. The cells were then subjectedto ChIP with antibodies to histone H1, and the precipitated DNA was subjected to real-time PCR analysis with primers specific for Wnt-responsive elements ora control region of the axin2 (C) or c-Myc (D) gene promoter. (E and F) HeLa cells infected with retroviral vectors for CHD8 or EGFP (control) shRNAs weresubjected to immunoblot analysis with antibodies to CHD8 or to Hsp90 (E) as well as to ChIP analysis (F) as in panels C and D. All quantitative data are means �SD from three independent experiments. ��, P � 0.01 (unpaired Student’s t test); NS, not significant (P � 0.05).
Histone H1 Recruitment by CHD8 Inhibits Wnt Signaling
We investigated whether the association of CHD8 with chro-matin at Wnt target genes is dependent on �-catenin. ChIP withantibodies to �-catenin revealed that forced expression of CHD8S
did not substantially affect the association of �-catenin with chro-matin in response to Wnt signaling (Fig. 2A). We also depletedHeLa cells of �-catenin with the use of two independent shRNAstargeting nucleotides 1774 to 1794 (shRNA-1) or nucleotides 501to 521 (shRNA-2) of the coding region (Fig. 2B), with the decreasein the abundance of endogenous �-catenin being greater withshRNA-1 than with shRNA-2. ChIP with antibodies to CHD8revealed that the amount of CHD8 associated with Wnt-responsive elements of the axin2 gene promoter was decreased in�-catenin-depleted cells stimulated with LiCl or Wnt3a and thatthe extent of this decrease was correlated with that in the totalabundance of �-catenin (Fig. 2C). These results thus indicatedthat �-catenin mediates the association of CHD8 with Wnt-responsive elements of the axin2 gene promoter. Collectively, ourobservations thus suggested that �-catenin binds to the axin2 genepromoter in a CHD8-independent manner, whereas CHD8 asso-ciation with this promoter is dependent on �-catenin. They arealso consistent with the results of ChIP analysis showing thatCHD8 was not localized to a substantial extent at Wnt target genesin unstimulated cells (Fig. 1D and E).
CHD8 recruits histone H1 to Wnt target genes. We previ-ously identified histone H1 as a molecule that associates withCHD8 with the use of a “shotgun” proteomics approach (29).Given that histone H1 was shown to be recruited to p53-responsive elements by CHD8 to suppress the transactivation ac-tivity of p53, we examined whether CHD8 also recruits histone H1to Wnt-responsive elements and forms a �-catenin–CHD8 – his-tone H1 trimeric complex. Pulldown assays in vitro with recom-binant glutathione S-transferase (GST)-tagged �-catenin, HA-tagged CHD8S, and FLAG-tagged histone H1 proteins that hadbeen produced in bacteria revealed that histone H1 directly inter-acted with CHD8S but not with �-catenin; histone H1 thus asso-ciated with �-catenin only in the presence of CHD8S (Fig. 3A). Wealso examined the association of 3�FLAG-tagged histone H1 withHA-tagged �-catenin and with Myc epitope-tagged CHD8S orCHD8S�H1, a CHD8S mutant that lacks the histone H1 bindingdomain, in HEK293T cells. Coimmunoprecipitation analysis re-vealed that the interaction between histone H1 and �-catenin wasmarkedly increased by forced expression of CHD8S but not by thatof CHD8S�H1 (Fig. 3B). Collectively, these results indicated that�-catenin directly interacts with CHD8 and that CHD8 directlyinteracts with histone H1, whereas �-catenin indirectly interactswith histone H1 through CHD8.
ChIP with antibodies to histone H1 showed that histone H1 wasindeed recruited specifically to the axin2 gene promoter in HeLa cellsin a manner dependent both on exposure of the cells to LiCl or Wnt3aand on forced expression of CHD8S (Fig. 3C). Similar results wereobtained with the Wnt-responsive elements of the c-Myc gene pro-moter (Fig. 3D). These results thus suggested that upregulation ofCHD8 might antagonize transactivation of Wnt target genes by�-catenin through recruitment of histone H1.
We also depleted HeLa cells of both CHD8S and CHD8L with aspecific shRNA (Fig. 3E) and then performed ChIP analysis withantibodies to histone H1 (Fig. 3F). The amount of histone H1associated with the Wnt-responsive elements of the c-Myc genepromoter was specifically and markedly reduced in the cells de-pleted of CHD8. Together with the results obtained by CHD8
overexpression, these data indicated that CHD8 recruits histoneH1 specifically to Wnt-responsive elements.
Histone H1 recruitment by CHD8 is necessary for suppres-sion of �-catenin function. Our previous study with a series ofdeletion mutants delineated the region of CHD8S required forbinding to histone H1 as that comprising residues 500 to 600 (29).Consistent with the previous finding that �-catenin interacts withrat CHD8 through residues 667 to 749 of the latter (36), whichcorrespond to residues 669 to 751 of mouse CHD8, coimmuno-
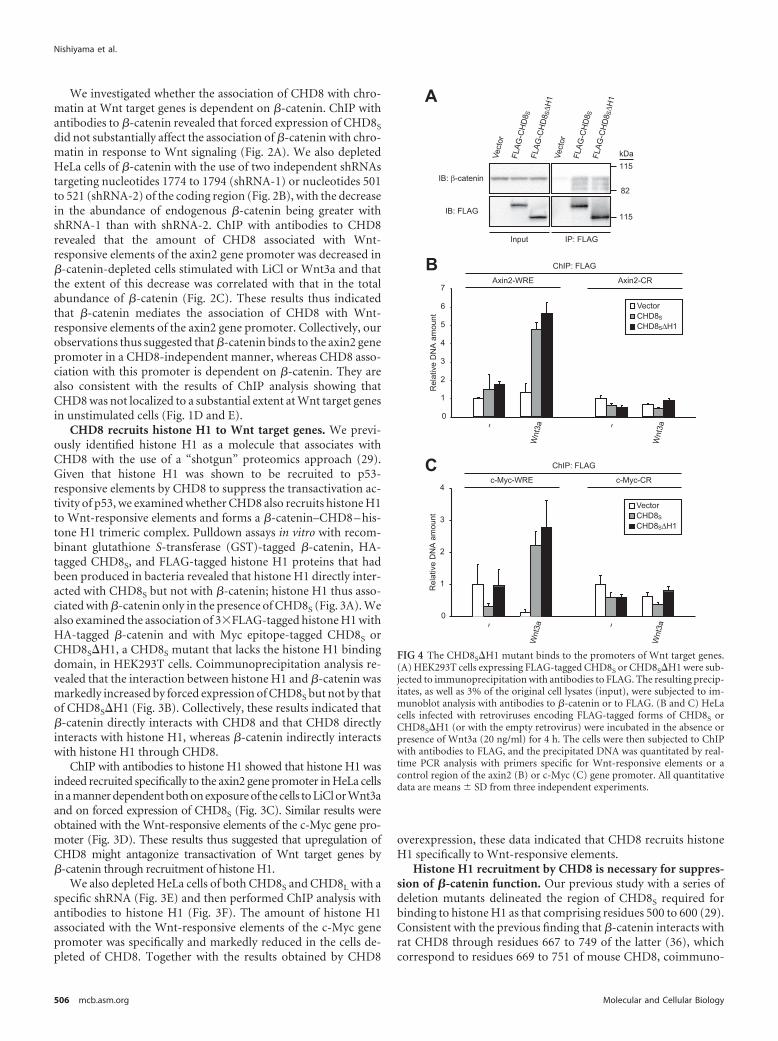
FIG 4 The CHD8S�H1 mutant binds to the promoters of Wnt target genes.(A) HEK293T cells expressing FLAG-tagged CHD8S or CHD8S�H1 were sub-jected to immunoprecipitation with antibodies to FLAG. The resulting precip-itates, as well as 3% of the original cell lysates (input), were subjected to im-munoblot analysis with antibodies to �-catenin or to FLAG. (B and C) HeLacells infected with retroviruses encoding FLAG-tagged forms of CHD8S orCHD8S�H1 (or with the empty retrovirus) were incubated in the absence orpresence of Wnt3a (20 ng/ml) for 4 h. The cells were then subjected to ChIPwith antibodies to FLAG, and the precipitated DNA was quantitated by real-time PCR analysis with primers specific for Wnt-responsive elements or acontrol region of the axin2 (B) or c-Myc (C) gene promoter. All quantitativedata are means � SD from three independent experiments.
precipitation analysis in HEK293T cells revealed that �-catenininteracted with the mouse mutant protein CHD8S�H1 (whichlacks residues 500 to 600) to a similar extent as it did with wild-type CHD8S (Fig. 4A). We also performed ChIP analysis withantibodies to FLAG in HeLa cells expressing FLAG-tagged CHD8S
or CHD8S�H1. Both CHD8S and CHD8S�H1 were recruited tosimilar extents to the Wnt-responsive elements of the axin2 (Fig.4B) or c-Myc (Fig. 4C) gene promoters in a Wnt signaling-dependent manner.
Whereas wild-type CHD8S promoted the recruitment of his-
tone H1 to the Wnt-responsive elements of the axin2 gene pro-moter, CHD8S�H1 failed to do so (Fig. 5A). Similar results wereobtained for the c-Myc gene promoter (Fig. 5B). Furthermore, incontrast to wild-type CHD8S, CHD8S�H1 did not substantiallyinhibit the upregulation of axin2 (Fig. 5C) or c-Myc (Fig. 5D)mRNAs induced by exposure of cells to LiCl or Wnt3a. Theseresults suggested that the inability of CHD8S�H1 to suppresstranscriptional activation by �-catenin is indeed attributable tothe lack of histone H1 binding, rather than to an altered localiza-tion of the mutant CHD8 on chromatin. We therefore conclude
FIG 5 Interaction of CHD8 with histone H1 is necessary for recruitment of histone H1 to the promoters of Wnt target genes. (A and B) HeLa cells stablyexpressing CHD8S or CHD8S�H1 were incubated in the absence or presence of LiCl (20 mM) or Wnt3a (20 ng/ml) for 4 h and then subjected to ChIP assays asin Fig. 3C and D, respectively. (C and D) HeLa cells stably expressing CHD8S or CHD8S�H1 were incubated in the absence or presence of LiCl or Wnt3a for 4h, after which the abundance of axin2 (C) and c-Myc (D) mRNAs was determined by quantitative RT-PCR analysis. All quantitative data are means � SD fromthree independent experiments.
Histone H1 Recruitment by CHD8 Inhibits Wnt Signaling
that CHD8 recruits histone H1 to Wnt-responsive elements andthat such recruitment of histone H1 results in suppression of tran-scriptional activation by �-catenin.
The COOH-terminal domain of histone H1 plays a major role indetermination of linker DNA conformation and chromatin conden-sation (10). We generated deletion mutants of histone H1 that lackthe NH2-terminal, central globular, or COOH-terminal domain (Fig.6A) and tested these mutants for their ability to bind to CHD8 withthe use of coimmunoprecipitation analysis (Fig. 6B). We found thatCHD8 interacted with the COOH-terminal domain of histone H1,suggesting that the biological relevance of this domain relies, at leastin part, on the interaction with CHD8. To examine the effect of de-
letion of the COOH-terminal domain of histone H1 on the suppres-sion of Wnt-mediated transcriptional activation, we forcibly ex-pressed wild-type histone H1 or the mutants lacking the NH2-terminal or COOH-terminal domain in NIH 3T3 cells (Fig. 6C). TheWnt3a-induced increase in the abundance of axin2 mRNA was in-hibited in cells expressing wild-type histone H1 and the mutant lack-ing the NH2-terminal domain compared with that in control cells,whereas it was unaffected in cells expressing the mutant lacking theCOOH-terminal domain. These results also suggested that the asso-ciation of histone H1 with CHD8 is essential for the suppression ofWnt target genes.
Histone H1 is essential for repression of �-catenin-dependent transcription. Our observation that the CHD8S�H1mutant is impaired in the ability to repress �-catenin function ledus to investigate the requirement for histone H1 in such repres-sion. To this end, we adopted two independent approaches. First,we examined the expression of Wnt target genes in HH1c�/�
HH1d�/� HH1e�/� triple-knockout (TKO) mouse ES cells. Inthese mutant ES cells, the abundance of histone H1 is reduced to�50% of that in wild-type ES cells, resulting in marked changes inchromatin structure; microarray analysis revealed that the expres-sion of only a few genes is altered in the mutant cells, however (6).Nevertheless, expression of axin2 (Fig. 7A) and c-Myc (Fig. 7B)genes was markedly increased in the TKO cells by low concentra-tions of LiCl or Wnt3a that do not activate Wnt signaling inwild-type control cells. The poor responsiveness of the wild-type cells to stimulation was likely attributable to the inhibitionof Wnt signaling by endogenous CHD8, the expression ofwhich is much greater in ES cells than in any adult tissuesexamined (29). These results thus suggested that CHD8 regu-lates cellular sensitivity to Wnt–�-catenin signaling in earlydevelopment. Second, we examined the effects both of a dom-inant negative mutant of histone H1 (EGFP-HH1) that reducesthe amount of endogenous histone H1 and of a reciprocal mu-tant (HH1-EGFP) that does not exhibit dominant negative ac-tivity (Fig. 7C and D) (8). ChIP analysis revealed that bothEGFP-HH1 and HH1-EGFP were equally recruited to the Wnt-responsive elements of the axin2 gene promoter (Fig. 7E). Ex-pression of EGFP-HH1 potentiated the stimulatory effects ofLiCl and Wnt3a on expression of the axin2 gene, whereas thatof HH1-EGFP had no such effect (Fig. 7F). Together, these twoapproaches thus supported our conclusion that CHD8 nega-tively regulates the transactivation function of �-catenin byrecruiting histone H1 to the promoters of Wnt target genes(Fig. 8). CHD8 is highly expressed in the early phase of embry-onic development and in cancer cell lines, whereas its expres-sion ceases during late embryogenesis and its abundance isgenerally low in normal adult tissues. CHD8 may thereforeserve to set a threshold for responsiveness to the Wnt signalingpathway in a development-dependent manner.
DISCUSSION
With the use of biochemical and genetic approaches, we haveshown that CHD8 negatively regulates �-catenin function by re-cruiting histone H1 to the promoters of Wnt target genes. Forma-tion of the �-catenin–CHD8 – histone H1 complex requires twoconditions: expression of CHD8 and stabilization of �-catenin byactivation of Wnt signaling. CHD8 is preferentially expressed inembryonic tissues and in cancer cell lines, which are thought toreflect the embryonic state.
FIG 6 The COOH-terminal domain of histone H1 is necessary for repressionof �-catenin-dependent transcription. (A) Schematic representation of full-length (FL) histone H1 (HH1) and its derivatives. The NH2-terminal domain,central globular domain, and COOH-terminal domain are shown. (B)HEK293T cells expressing 3�FLAG-tagged histone H1 derivatives were sub-jected to immunoprecipitation with antibodies to FLAG. The resulting precip-itates, as well as 3% of the original cell lysates (input), were subjected to im-munoblot analysis with antibodies to CHD8 or to FLAG. (C) NIH 3T3 cellsstably expressing HH1 derivatives were incubated in the absence or presence ofWnt3a (20 ng/ml) for 4 h, after which the abundance of axin2 mRNA wasdetermined by quantitative RT-PCR analysis. Data are means � SD from threeindependent experiments.
CHD8S, also known as duplin, was originally isolated as a neg-ative regulator of the Wnt–�-catenin signaling pathway (36). Re-cent studies have indicated that CHD8L interacts directly with�-catenin and negatively regulates �-catenin-dependent gene ex-pression through the ATP-dependent modulation of chromatinstructure (41). On the other hand, the COOH-terminal region ofCHD8L interacts with the insulator-binding protein CTCF, withthis interaction being important for insulator activity (14). How-
ever, our present study suggests that not only CHD8L but alsoCHD8S is able to inhibit Wnt- and �-catenin-dependent transac-tivation. Given that CHD8S contains the chromodomain but lacksthe Snf2 helicase domain and the CTCF binding domain ofCHD8L, neither the ATP-dependent remodeling activity medi-ated by the Snf2 helicase domain nor CTCF binding through theCOOH-terminal region of CHD8L is necessary for this suppres-sive effect of CHD8 on the Wnt–�-catenin signaling pathway. In
FIG 7 Requirement for histone H1 in repression of �-catenin-dependent transcription. (A and B) Wild-type (WT) or HH1c�/� HH1d�/� HH1e�/� (HH1TKO) ES cells were incubated with LiCl (2.5 mM) or Wnt3a (2.5 ng/ml) for 4 h and then subjected to quantitative RT-PCR analysis of axin2 (A) and c-Myc (B)mRNAs. (C) Schematic representation of histone H1 (HH1) tagged at its NH2 terminus (EGFP-HH1) or COOH terminus (HH1-EGFP) with EGFP. Both fusionproteins were also tagged at their NH2 termini with HA. (D) NIH 3T3 cells infected with retroviruses encoding EGFP-HH1 or HH1-EGFP (or with the emptyvector) were then subjected to immunoblot analysis with antibodies to HA, to histone H1, or to Hsp90. (E) NIH 3T3 cells infected as in panel D were incubatedin the absence or presence of Wnt3a (20 ng/ml) for 4 h. The cells were then subjected to ChIP with antibodies to HA, and the precipitated DNA was quantitatedby real-time PCR analysis with primers specific for Wnt-responsive elements or a control region of the axin2 gene promoter. (F) NIH 3T3 cells infected as in panelD were incubated with LiCl (20 mM) or Wnt3a (20 ng/ml) for 4 h and then subjected to quantitative RT-PCR analysis of axin2 mRNA. All quantitative data aremeans � SD from three independent experiments. ��, P � 0.01 versus the corresponding value for cells infected with the empty virus (unpaired Student’s t test).
Histone H1 Recruitment by CHD8 Inhibits Wnt Signaling
contrast, this suppressive activity requires the region present inboth CHD8S and CHD8L that mediates binding to histone H1. ACHD8S mutant that lacks the histone H1 binding domain thus didnot exhibit this inhibitory activity, and either depletion of histoneH1 or expression of a dominant negative mutant thereof increasedthe cellular sensitivity to Wnt signaling. Our results thus indicatethat histone H1 recruitment mediated by CHD8 is fundamental tonegative regulation of the Wnt–�-catenin signaling pathway.
Histone H1 molecules are highly mobile, and the interaction of aspecific H1 molecule with a specific nucleosome is transient (18, 20),suggesting that the molecules are continuously exchanged amongchromatin binding sites according to a “stop-and-go” process inwhich a histone H1 molecule remains at a binding site for a limitedtime before dissociating and moving rapidly to another such site.Most chromatin fibers thus likely always contain histone H1, butthere is a continuous turnover of histone H1 molecules at the level ofthe individual nucleosome. Given that histone H1 is implicated inregulation of the expression of specific genes, we postulated the exis-tence of a factor such as CHD8 that extends the residence time ofhistone H1 at the corresponding specific chromatin sites. It is alsopossible that CHD8 alters the interaction of histone H1 with linkerDNA and represses transcription by enhancing the intrinsic stabilityof nucleosomes. The detailed mechanisms of transcriptional suppres-sion for specific genes remain to be elucidated.
The identification of various proteins that associate with�-catenin has provided insight into its function as a transcrip-tional regulator. Many of these proteins, including componentsof histone acetylation and methylation complexes as well aschromatin-binding proteins, contribute to regulation of chromatin
structure or of RNA polymerase II. Several transcriptional complexesor coactivators, including Bcl-9 (also known as Lgs), Pygopus (Pygo),polymerase-associated factor 1 (Paf1), and SET1 (trithorax), havebeen found to be recruited by �-catenin for target gene regulation(17, 24, 39). At active target genes, Pygo binds through its PHD fingerto methylated H3K4 and, by using Bcl-9 as an adaptor, retains�-catenin near Wnt-responsive elements independently of Tcf (37).Pygo-dependent binding of �-catenin to methylated histones freesTcf to recruit its corepressors, such as Groucho, that counteract theactivating �-catenin-induced chromatin-remodeling processes. Wehave now uncovered another mode of �-catenin regulation mediatedby the CHD8-dependent recruitment of histone H1 to the promotersof Wnt target genes.
We previously showed that apoptosis mediated by p53-dependent transactivation is also suppressed by histone H1 re-cruited by CHD8 (29). Loss of CHD8 induced hyperactivation ofp53, resulting in apoptosis, which was prevented by the additionaldepletion of p53. Mice deficient in CHD8 die during early em-bryogenesis (around embryonic day 7.5) as a result of aberrantapoptosis induced by the unscheduled activation of p53 (28, 29).It is likely that such apoptosis dominates and masks possible phe-notypes induced by hyperactivation of Wnt signaling. Additionaldeletion of p53 in CHD8-deficient mice ameliorated the develop-mental arrest, resulting in extension of survival until embryonicday 10.5. Death of the Chd8�/� p53�/� mice is associated withsevere hemorrhage, probably as a result of a defect in cardiovas-cular development (M. Nishiyama and K. I. Nakayama, unpub-lished observations). Given that mice homozygous for a mutantApc allele encoding a product truncated at position 716 (Apc�716)
FIG 8 Model for CHD8 function. CHD8 interacts with �-catenin and subsequently facilitates recruitment of histone H1, resulting in chromatin compaction andtranscriptional silencing.
die in utero as a result of the unscheduled activation of Wnt sig-naling (30) and that Wnt signaling plays an essential role in car-diovascular development during embryogenesis (4), the embry-onic death of Chd8�/� p53�/� mice may be attributable to a defectin the cardiovascular system resulting from unscheduled activa-tion of Wnt signaling.
The mode of action of CHD8 in antagonism of Wnt–�-cateninsignaling appears almost identical to that for p53 inhibition. Thehistone H1 binding domain that is present in both CHD8S andCHD8L is thus indispensable for CHD8-mediated inhibition oftransactivation by p53 or �-catenin. Furthermore, genetic evi-dence from cells depleted of histone H1 also supports this similar-ity. Given that CHD8 is preferentially expressed in embryonictissues and in cancer cell lines, which are thought to reflect theembryonic state, CHD8 may counteract p53 and Wnt–�-cateninfunction through recruitment of histone H1 during early embryo-genesis to set a threshold for induction of apoptosis, cell prolifer-ation, and axis formation. It is also possible that CHD8 may bindto transcriptional factors other than p53 and �-catenin and act asa more general inhibitor of transactivation activity through re-cruitment of histone H1.
ACKNOWLEDGMENTS
We thank T. Kitamura for pMX-puro; J. M. Cunningham and K. Hanadafor the mCAT-1 plasmid; F. Ishikawa and R. Funayama for plasmids en-coding the EGFP-HH1 and HH1-EGFP mutants of histone H1; A. Kiku-chi for Wnt3a; M. Sato, H. Takeda, C. Mitai, N. Nishimura, and K. Oy-amada for technical assistance; A. Ohta for help in preparation of themanuscript; and members of our laboratories for discussion.
A.I.S. was supported by NIH grant CA79057.
REFERENCES1. Barker N, et al. 2001. The chromatin remodelling factor Brg-1 interacts
with beta-catenin to promote target gene activation. EMBO J. 20:4935– 4943.
2. Bienz M, Clevers H. 2000. Linking colorectal cancer to Wnt signaling.Cell 103:311–320.
3. Billin AN, Thirlwell H, Ayer DE. 2000. �-Catenin-histone deacetylaseinteractions regulate the transition of LEF1 from a transcriptional repres-sor to an activator. Mol. Cell. Biol. 20:6882– 6890.
4. Cohen ED, Tian Y, Morrisey EE. 2008. Wnt signaling: an essentialregulator of cardiovascular differentiation, morphogenesis and progeni-tor self-renewal. Development 135:789 –798.
6. Fan Y, et al. 2005. Histone H1 depletion in mammals alters global chro-matin structure but causes specific changes in gene regulation. Cell 123:1199 –1212.
7. Flanagan JF, et al. 2005. Double chromodomains cooperate to recognizethe methylated histone H3 tail. Nature 438:1181–1185.
8. Funayama R, Saito M, Tanobe H, Ishikawa F. 2006. Loss of linkerhistone H1 in cellular senescence. J. Cell Biol. 175:869 – 880.
9. Graham TA, Weaver C, Mao F, Kimelman D, Xu W. 2000. Crystalstructure of a �-catenin/Tcf complex. Cell 103:885– 896.
10. Happel N, Doenecke D. 2009. Histone H1 and its isoforms: contributionto chromatin structure and function. Gene 431:1–12.
11. He TC, et al. 1998. Identification of c-MYC as a target of the APC path-way. Science 281:1509 –1512.
12. He X, Semenov M, Tamai K, Zeng X. 2004. LDL receptor-relatedproteins 5 and 6 in Wnt/�-catenin signaling: arrows point the way. Devel-opment 131:1663–1677.
13. Hurd EA, et al. 2007. Loss of Chd7 function in gene-trapped reportermice is embryonic lethal and associated with severe defects in multipledeveloping tissues. Mamm. Genome 18:94 –104.
14. Ishihara K, Oshimura M, Nakao M. 2006. CTCF-dependent chromatininsulator is linked to epigenetic remodeling. Mol. Cell 23:733–742.
15. Kamura T, et al. 2004. VHL-box and SOCS-box domains determinebinding specificity for Cul2-Rbx1 and Cul5-Rbx2 modules of ubiquitinligases. Genes Dev. 18:3055–3065.
16. Korinek V, et al. 1997. Constitutive transcriptional activation by a�-catenin-Tcf complex in APC�/� colon carcinoma. Science 275:1784 –1787.
17. Kramps T, et al. 2002. Wnt/wingless signaling requires BCL9/legless-mediated recruitment of pygopus to the nuclear �-catenin-TCF complex.Cell 109:47– 60.
18. Lever MA, Th’ng JP, Sun X, Hendzel MJ. 2000. Rapid exchange ofhistone H1.1 on chromatin in living human cells. Nature 408:873– 876.
19. Menon T, Yates JA, Bochar DA. 2010. Regulation of androgen-responsive transcription by the chromatin remodeling factor CHD8. Mol.Endocrinol. 24:1165–1174.
20. Misteli T, Gunjan A, Hock R, Bustin M, Brown DT. 2000. Dynamicbinding of histone H1 to chromatin in living cells. Nature 408:877– 881.
21. Moon RT, Kohn AD, De Ferrari GV, Kaykas A. 2004. WNT and�-catenin signalling: diseases and therapies. Nat. Rev. Genet. 5:691–701.
22. Morin PJ, et al. 1997. Activation of �-catenin-Tcf signaling in coloncancer by mutations in �-catenin or APC. Science 275:1787–1790.
23. Morita S, Kojima T, Kitamura T. 2000. Plat-E: an efficient and stablesystem for transient packaging of retroviruses. Gene Ther. 7:1063–1066.
24. Mosimann C, Hausmann G, Basler K. 2006. Parafibromin/Hyrax acti-vates Wnt/Wg target gene transcription by direct association with�-catenin/Armadillo. Cell 125:327–341.
25. Nakayama K, et al. 1996. Mice lacking p27Kip1 display increased body size,multiple organ hyperplasia, retinal dysplasia, and pituitary tumors. Cell85:707–720.
26. Nakayama K, et al. 2000. Targeted disruption of Skp2 results in accumu-lation of cyclin E and p27Kip1, polyploidy and centrosome overduplica-tion. EMBO J. 19:2069 –2081.
27. Nelson WJ, Nusse R. 2004. Convergence of Wnt, �-catenin, and cadherinpathways. Science 303:1483–1487.
28. Nishiyama M, et al. 2004. Early embryonic death in mice lacking the�-catenin-binding protein Duplin. Mol. Cell. Biol. 24:8386 – 8394.
29. Nishiyama M, et al. 2009. CHD8 suppresses p53-mediated apoptosisthrough histone H1 recruitment during early embryogenesis. Nat. CellBiol. 11:172–182.
30. Oshima M, et al. 1995. Loss of Apc heterozygosity and abnormal tissuebuilding in nascent intestinal polyps in mice carrying a truncated Apcgene. Proc. Natl. Acad. Sci. U. S. A. 92:4482– 4486.
31. Pfaffl MW. 2001. A new mathematical model for relative quantification inreal-time RT-PCR. Nucleic Acids Res. 29:e45.
32. Rodenberg JM, et al. 2010. Regulation of serum response factor activityand smooth muscle cell apoptosis by chromodomain helicase DNA-binding protein 8. Am. J. Physiol. Cell Physiol. 299:C1058 –1067.
33. Rodriguez-Paredes M, Ceballos-Chavez M, Esteller M, Garcia-DominguezM, Reyes JC. 2009. The chromatin remodeling factor CHD8 interacts withelongating RNA polymerase II and controls expression of the cyclin E2 gene.Nucleic Acids Res. 37:2449–2460.
34. Roose J, et al. 1998. The Xenopus Wnt effector XTcf-3 interacts withGroucho-related transcriptional repressors. Nature 395:608 – 612.
35. Rubinfeld B, et al. 1997. Stabilization of �-catenin by genetic defects inmelanoma cell lines. Science 275:1790 –1792.
36. Sakamoto I, et al. 2000. A novel �-catenin-binding protein inhibits beta-catenin-dependent Tcf activation and axis formation. J. Biol. Chem. 275:32871–32878.
37. Sampietro J, et al. 2006. Crystal structure of a �-catenin/BCL9/Tcf4complex. Mol. Cell 24:293–300.
38. Schnetz MP, et al. 2009. Genomic distribution of CHD7 on chromatintracks H3K4 methylation patterns. Genome Res. 19:590 – 601.
39. Sierra J, Yoshida T, Joazeiro CA, Jones KA. 2006. The APC tumorsuppressor counteracts �-catenin activation and H3K4 methylation atWnt target genes. Genes Dev. 20:586 – 600.
40. Sims RJ, 3rd, et al. 2007. Recognition of trimethylated histone H3 lysine4 facilitates the recruitment of transcription postinitiation factors andpre-mRNA splicing. Mol. Cell 28:665– 676.
41. Thompson BA, Tremblay V, Lin G, Bochar DA. 2008. CHD8 is anATP-dependent chromatin remodeling factor that regulates �-catenintarget genes. Mol. Cell. Biol. 28:3894 –3904.
42. Vissers LE, et al. 2004. Mutations in a new member of the chromodomaingene family cause CHARGE syndrome. Nat. Genet. 36:955–957.
Histone H1 Recruitment by CHD8 Inhibits Wnt Signaling
43. Willert K, et al. 2003. Wnt proteins are lipid-modified and can act as stemcell growth factors. Nature 423:448 – 452.
44. Willert K, Jones KA. 2006. Wnt signaling: is the party in the nucleus?Genes Dev. 20:1394 –1404.
45. Yates JA, Menon T, Thompson BA, Bochar DA. 2010. Regulation ofHOXA2 gene expression by the ATP-dependent chromatin remodelingenzyme CHD8. FEBS Lett. 584:689 – 693.
46. Yuan CC, et al. 2007. CHD8 associates with human Staf and contributesto efficient U6 RNA polymerase III transcription. Mol. Cell. Biol. 27:8729 – 8738.
47. Zhang Y, LeRoy G, Seelig HP, Lane WS, Reinberg D. 1998. Thedermatomyositis-specific autoantigen Mi2 is a component of a complexcontaining histone deacetylase and nucleosome remodeling activities. Cell95:279 –289.