tion, a number of workers have reported that the degree ofphosphonylation of histones exerts a specific influence onthe processes of differentiation and development in varioussystems (16, 45).
The number of systems available for the in vivo study ofthe biological significance of histone phosphomylation islimited. The majority of such investigations have been canned out with regenerating rat liver, but this system has aninherent difficulty, in that a series of overlapping peaks inDNA synthesis is observed, making the correlation of onebiochemical event with another extremely difficult. In acontinuing series of experiments designed to elucidate themechanism of action of the tumor promoter TPA,2 it wasnoted that treatment of mouse epidermis with this agentinduces a wave of cell proliferation that is followed by adistinct wave of keratinization. The wave of differentiationhas been characterized momphologically (2, 38) as well asbiochemically by demonstrating the synthesis of differentiation-specific epidermal proteins (8). Furthermore, with asuitably low dose of TPA, only a single wave of DNA synthesis is induced, which is sharp and completely reproducible(10, 29). In previous studies on the mechanism of action ofTPA, a wide variety of doses and conditions of applicationhas been used by various ‘authors.The results reported onthe kinetics of the stimulation of nucleic acid synthesis, forexample, are consequently confusing and contradictory.The present experiments were therefore designed to investigate the temporal relationship among the various biochemical events during the discrete waves of proliferationand differentiation that are observed in the epidermis aftertreatment with this low dose of TPA.
In a series of publications (13, 26), Boutwell et at. havedeveloped the hypothesis that tumor promotion involves thespecific activation of genes that had been critically alteredby prior interaction with an “initiating agent.―A recentreport on stimulation of the phosphonylation of epidemmalhistones by TPA was interpreted by those authors as beingevidence for the role of specific gene activation in theprocess of tumor formation (39). In these investigations,however, the histones were characterized only as “rapidly
I Present address: The Beatson Institute for Cancer Research, Wolfson
Histone Phosphorylation and Synthesis of DNA and RNA duringPhases of Proliferation and Differentiation Induced inMouse Epidermis by the Tumor Promoter 12-0-Tetradecanoyl-phorbol-13-acetate
Allan Balmain,1 Angel Alonso, and JUrgen Fischer
Institute for ExperimentalPathology,GermanCancerResearchCenter,6900Heidelberg,Im NeuenheimerFeld280,Germany
SUMMARY
The tumor promoter 12-O-tetradecanoyl-phonbol-1 3-acetate (TPA) induces discrete waves of proliferation and differentiation when applied to mouse epidermis. The aim of thisstudy was to elucidate the temporal relationship betweenhistone phosphonylation and synthesis of DNA and RNA inmouse epidermis during the process of tumor promotion.This investigation was facilitated by choosing a low butnevertheless strongly tumor-promoting dose of TPA (0.002
@mole/mouse),which induced the epidemmal cells to gothrough only one round of DNA synthesis and cell division.Histones were isolated from mouse epidermis, and the ratesof phosphomylation of the individual histone species weredetermined at different times after treatment with TPA. Theresults demonstrated that the phosphorylation of Hi histone was initiated at about the same time as the synthesis ofDNA but continued past the S phase and reached a maximum simultaneously with the maximum in the epidermalmitotic rate. The only other histone that phosphorylated toany significant extent was Histone H2A. From the resultsobtained, it was concluded that histone phosphorylation inthe epidermis is related to the processes of DNA replicationand mitosis after stimulation with TPA. Positive evidence forthe activation of specific genes, which has been proposedby other authors to be important in the promotion of epidermal tumors, was not found in this investigation.
INTRODUCTION
The role of histone phosphomylation in the control ofcellular proliferation and differentiation has been the subject of innumerable investigations oven the past few years.Earlier studies linked histone phosphonylation to specificgene activation (28, 31), cytoplasmic-nucleam transport ofhistones (36), or DNA replication (5, 6). More recent evidence has suggested a central molefor Hi-histone phosphonylation in the condensation of chromosomes in preparationfor mitosis (12, 15, 34), although this conclusion does notseem to be applicable to all systems studied (22). In addi
migrating―or “slowlymigrating―fractions, and no controlled comparison was carried out with the kinetics of thesynthesis of other macromolecules.
The main purpose of these experiments was therefore: (a)to study the temporal relationship among the stimulation ofnucleic acid synthesis, histone phosphorylation, and mitosis during waves of proliferation and differentiation in vivoand (b) to attempt to determine whether histone prolifemation is important in tumor promotion as a result of specificgene activation or because of its inherent molein the proliferation and division of epidemmal cells.
MATERIALS AND METHODS
Radiochemicals. 32P1(specific activity, 1250 mCi/mmole)in sterile water was obtained from New England Nuclear,Boston, Mass., as was [G-3H]umidine (15 Ci/mmole).[methyt-3HjThymidine (15 to 30 mCi/mmole) was ot@tainedfrom The Radiochemical Centre, Amersham, England.
Animals. Seven- to 8-week-old female NMRI mice wereused in all experiments. Animals were maintained in climatized rooms with an artificially invensed day-night rhythm.The back skins of the mice were shaved 48 hr prior to use,and any animals showing evidence of wounds or hair megrowth were excluded from experiments. Mice were treatedwith the appropriate dose of TPA (normally 0.002 @mole)dissolved in 0.1 ml acetone by pipeting the solution onto theback skin. Controls were treated in the same way withacetone alone. Injection with radioactive precursor was canned out 1 hr before killing in the case of 32P1and 30 mmbefore killing for [3H]umidineand [3H]thymidine. Skins wereremoved from the dead animals and were immediately depilated at 4°with hair-removing cream (Pilca) for 3 mm. Theskins were then thoroughly washed in ice water and pinneddermis side down onto cork plates; the epidemmal fractionswere removed by gentle scraping with a rounded scalpelblade. This method provides a good separation of dermisfrom epidermis with a minimum of demmal contamination(9).
Isolation of Nuclei and Preparation of Histones. In someexperiments, up to 12 mice were taken at each time point,but in most cases 6 mice provided enough epidemmal matemialfor repeat analyses of histone-specific activities.
Epidermal fractions were homogenized with 10 to 15strokes in a Potter-type homogenizer. This and all subsequent operations were carried out at 4°.The homogenizingmedium (1 mI/mouse epidermis) was TKMS. The homogenate was filtered through fine gauze, and a crude nuclearpellet was prepared by centnifugation at 3000 rpm for 15 mm(Sorvall centrifuge, SM 34 motor). The crude pellet wasresuspended by gentle hand homogenization in TKMS (6ml) and layered onto TKMS in which the sucrose concentration was increased to 1.5 M (6 ml). Fractions were thencentrifuged at 35,000 rpm for 2 hr in a Beckman ultmacentnifuge (SW 40 rotor). The resulting nuclear pellets werewashed successively with 0.14 M NaCI and 80% ethanol; thetotal histones were extracted with 0.4 M H2S04 (3 ml) at 0°for 30 mm. After centnifugation (10,000 rpm, 15 mm), theextraction was repeated for 10 mm at 0°.The pooled supernatants containing the histones were then dialyzed over
night against 2 changes of 0.1 N acetic acid. The fractionswere subsequently freeze-dried and further prepared foracid-urea gel electmophoresis according to the method ofPanyirn and Chalkley (37) or for SDS-gel electrophoresisusing a method essentially similar to that of Laernrnli (30).
The above method for the preparation of nuclei providedfractions contaminated mainly by fragments of insolublekeratin. The nuclear sediment consisted of approximately70% nuclei as estimated from microscopic investigation.
However, no appreciable contamination of the histones wasobserved after separation by acidic and basic gel electrophoresis. Nuclei that were morphologically intact and vimtually free from contamination could be prepared by a procedume involving treatment with dilute citric acid. Thesecould not be used for the present investigations since analysis by gel electrophomesis showed the Hi histone to bemissing, an observation made previously by other workers(32).
Determination of Specific Activity of Histone Fractions.Acid-urea gels of 32P-labeled histones prepared at varioustimes after treatment with TPA were stained with 1% Arnidoblack in 0.7% acetic acid and destained by diffusion in 0.7%acetic acid. The gels were scanned at 550 nm with a Gilfordrecording spectrophotornetem equipped with a gel-scanningdevice. The area under each peak was determined by tracing the scan onto heavy transparent paper, cutting out eachpeak separately, and weighing. Since the peaks obtainedfor Histones H3 and H2B were not adequately separated,these 2 fractions were cut out together, and an averagevalue for their specific activities was obtained . For measurement of the incorporated radioactivity, gels were frozen onsolid CO2,and each band was cut out with a razor blade. Gelslices were incubated overnight in 30% H@O2at 60°(11),diluted with toluene-dioxane scintillatom, and counted in aliquid scintillation counter (Nuclear-Chicago Mark II). Possible differences in quench between samples were checkedby the use of external standards. Relative values for thespecific activity of each histone fraction could then be calculated.
Extraction and Characterization of Phosphorylated HIHistone. Pure Hi histone was isolated from epidermis andliver essentially according to the method of Johns (27).Washed nuclei were extracted twice with 0.74 N perchlomicacid (2 ml) for 15 mm at 0°.Eight hundred mg of solidtrichlomoacetic acid were added, and the mixture was alowed to stand overnight at 0°.The precipitate of Hi histone
was collected by centnifugation at 20,000 rpm for 30 mm(Beckman ultmacentmifuge,SW 40 rotor), washed once withacetone containing 0.05% HCI and once with acetonealone, and dried under vacuum. The isolated Hi fractionwas subjected to preparative acid-urea gel electmophoresis;the Hi band was localized by comparison with a stained gelrun under identical conditions, cut from the gel with a razorblade, and eluted with distilled water. After freeze-drying,the Hi histone was digested with trypsin essentially according to the method of Breindl and Gallwitz (17). Treatmentwith trypsin was carried out for 3.5 hr at 40°.The resultingtnyptic peptides were separated by high-voltage paper electrophoresis and stained with ninhydnin. For comparison,the exact procedure was carried out with the Hi histone isolated from mouse liver.
‘.FordemonstrationofthefactthatphosphorylationoftheHi histone was really taking place in vivo and exclusion ofthe possibility of unspecific adsorption of 32Pon the gels,aliquotsof the 32P-Iabeled purified Hi histone were hydmolyzed In 6 N HCI.for6 hrat liO°asdescribed byAllenton andPenman (1). The hydmolysate was used for high-voltagepaper.electmophoresisin the presence of phosphosemineand'@ phosphothmeonine standards. The amino acids weredetected by spraying with ninhydnin. The paper was thencut into 2-mm-broad strips, which were immersed in scintillatomand counted.
Determination of the Specific Activity of DNA and RNA.Epidemrnal fractions labeled with either [3H]umidine or[3H]thyrnidine were prepared from groups of 4 to 6 mice atdifferent times after treatment with TPA. Homogenizationwas carried out in double-distilled water, the homogenatewas filtered through gauze, and the total nucleic acids wereprecipitated at 0°by addition of TCA to a final concentrationof 10%. The homogenate was centrifuged at 3000 rpm for 10mm (Sorvall, SM 34 motor), and the pellet was washed 3times with 5% TCA. Lipids were removed by extraction withethanol and ether, and the RNA was hydrolyzed by incubation at 37°for 3 hr in 0.3 N KOH. DNA and protein wereprecipitated at 0°by acidification with TCA to a concentration of 10%. Aliquots of the supemnatant were used for thedetermination of [3H]unidine incorporated into RNA by scmtillation counting and for the estimation of RNA by theorcinol reaction (40). The specific activity of the RNA atdifferent times after treatment of the epidermis with TPAcould then be calculated. In some experiments, RNA wasprepared by a phenol-chloroform extraction procedure, andthe specific activity was determined by scintillation countin9 and by measurement of absombance at 260 nm. “Acidsoluble activity―is defined as the amount of radioactivityfound in the supemnatant obtained by centnifugation of theacidified homogenate. This was related to the amount ofnucleic acid found in the corresponding sediment, and theresults were expressed as cpm/@g DNA.
DNA was estimated by heating the RNA- and lipid-freepellet in 5% TCA at 100°for 5 mm. Aliquots of the supemnatant were used for DNA determination by the method ofBurton (18) or for radioactivity measurements.
Estimation of Mitotic Counts. Mitotic figures werecounted in interfollicular epidermis in stained, paraffinembedded sections of mouse skin. Mice were killed at 3-hrintervals after treatment with TPA. Mitoses were countedwithout previous injection of metaphase-amresting agentssuch as colchicine on vincnistine. This was necessary tomaintain the exact conditions that were observed during thehistone phosphomylation experiments. The number of mitotic figures was counted per 1000 basal cells. Three sections were counted from each mouse, with 4 mice beingkilled at each time point. A minimum of 12,000 basal cellswas therefore counted at each time point.
RESULTS
The histones isolated from mouse epidermis are verysimilar to mouse liver histones by comparison on acid-ureaand SDS-polyacrylarnide gel systems (21) (Fig. 1). Fig. lb
1550 CANCER RESEARCHVOL. 37
•@11@,H3@ dimy•
ab
H4-―
CFig. 1. Histone patterns obtained from mouse liver and epidermis. a, acid
urea gel of liver histones; b, acid-urea gel of epidermal histones; c, SDS-gelof epidermal histones (15% polyacrylamide).
shows the incompletely reduced epidemmal histone fraction,in which the dimen of Histone H3 is visible. When gels wereto be quantitated for the calculation of specific activities,the total histone fraction was reduced by treatment with 2-mercaptoethanol, which resulted in the disappearance ofthe H3 dimem. Chart 1 shows the degree of peak separationobtained when the stained gels were scanned. Similam histone patterns have previously been obtained for wholemouse skin (41) and isolated epidermis (43).
The results of the phosphomylation experiments areshown in Chart 2. Animals were treated at Time Point 0 withTPA (0.002 @rnoIe),and groups of 6 to 12 mice were giveninjections of 32p1@ hr prior to being killed at the time pointsshown. The highest degree of phosphorylation was observed in the Hi fraction, with a maximum phosphorylationmateattained about 18 hr after treatment. A small increase inthe rate of phosphorylation of the Hi histone was observedas soon as 3 hr after application, but this was not verysignificant and could be due to an early influx of 32p1into theepidemmal cells (9).
The only other histone for which a significant increase inthe rate of phosphorylation was observed was histone H2A.The phosphonylation of histones H3 plus H2B and histoneH4 was not reproducibly or significantly increased over thecontrol values.
It was necessary to demonstrate that the incorporationobserved was due to the phosphomylation of amino acidresidues of the Hi histone. For this purpose, Hi was isolated from epidermis labeled with 32p1between 17 and 18 hrafter treatment with TPA. The Hi fraction was purified bypreparative acid-urea gel electrophoresis and comparedwith the Hi histone isolated from mouse liver by gel electro
on separation of the tryptic peptides from the liver Hi histone.
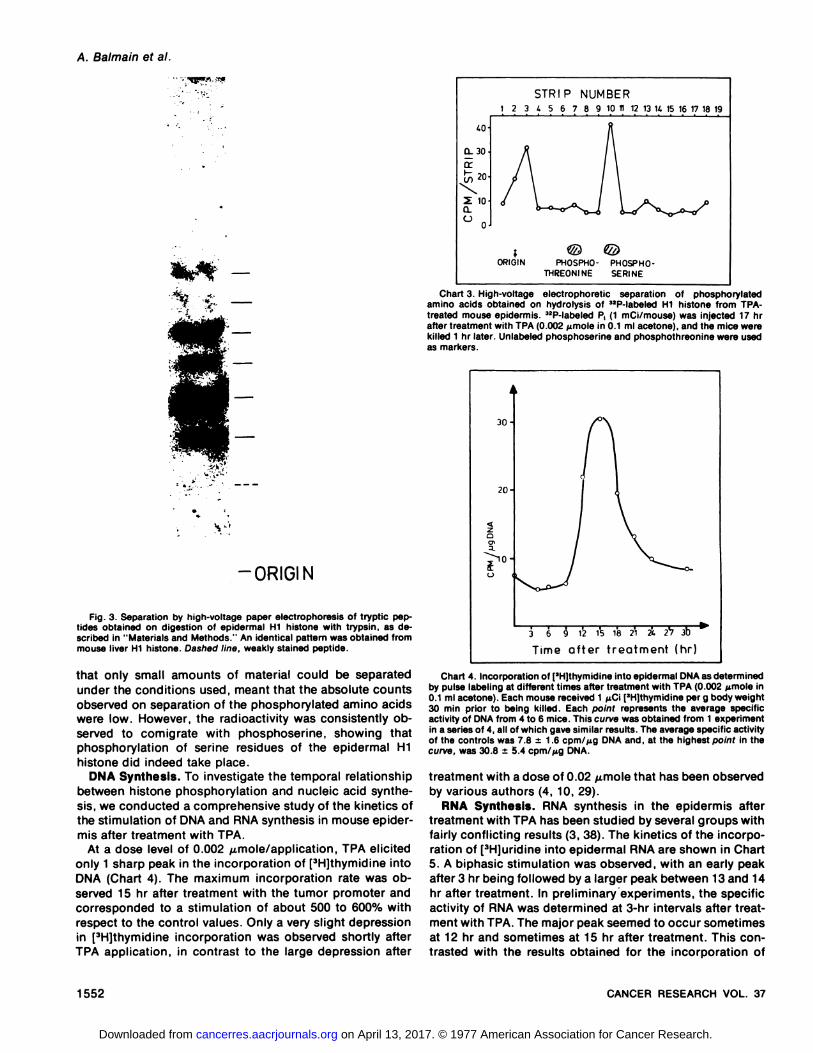
Aliquots of the phosphorylated Hi fraction after elutionfrom the gels were subjected to hydrolysis in 6 N HCI asdescribed in “Materialsand Methods.―High-voltage electrophomesis of the hydmolysate together with standards ofphosphosenine and phosphothreon me demonstrated thatthe bulk of the radioactivity comigrated with phosphosenine(Chart 3). Some activity was consistently observed at thestart, presumably due to the presence of incompletely hydrolyzed material. The nature of these in vivo experimentswas such that the incorporation matesattained were relativelylow,inspiteofthefactthathighamounts ofmadioactivity were injected (1 mCi/mouse). This, along with the fact
H2B
H2A
H4
Hi
Chart I . Scan of epidermal histones after reduction with 2-mercaptoethanol and separation on 15% acid-urea polyacrylamide gels.
3 6 9 12
Time after treatment (hr)
Chart 2. Phosphorylation of epidermal histone fractions at different timesafter treatment with TPA (0.002 @moIein 0.1 ml acetone). 0, Hi ; 0, H2A; x,H2B plus H3; •,H4. Each mouse received I mCi 32P1by p. injection 1 hrbefore being killed. The values obtained for Hi phosphorylation are theaverages from 3 separate experiments, with 6 to 12 mice being used at eachtime point in each experiment. The values forthe remaining histone fractionsare the averages from 2 separate experiments.
phomesis and tryptic digestion. The epidemmal and liver Hihistone fractions showed identical patterns on polyacrylamide gels (Fig. 2). Digestion of epidemmal Hi with trypsinand subsequent separation of the resulting tryptic peptidesby high-voltage paper electrophomesis showed that at least 7peptides were formed (Fig. 3). Similar results were obtained
ab
Fig. 2. Isolated Hi histone fractions from mouse liver (a) and epidermis (b)(15% acid-urea polyacrylamide gels).
Fig. 3. Separation by high-voltage paper electrophoresis of tryptic peptides obtained on digestion of epidermal Hi histone with trypsin, as describedin “Materialsand Methods.―An identicalpatternwasobtainedfrommouse liver Hi histone. Dashed line, weakly stained peptide.
that only small amounts of material could be separatedunder the conditions used, meant that the absolute countsobserved on separation of the phosphorylated amino acidswere low. However, the radioactivity was consistently observed to cornignate with phosphosemine, showing thatphosphomylation of senine residues of the epidemmal Hihistone did indeed take place.
DNA Synthesis. To investigate the temporal relationshipbetween histone phosphorylation and nucleic acid synthesis, we conducted a comprehensive study of the kinetics ofthe stimulation of DNA and RNA synthesis in mouse epidemmis after treatment with TPA.
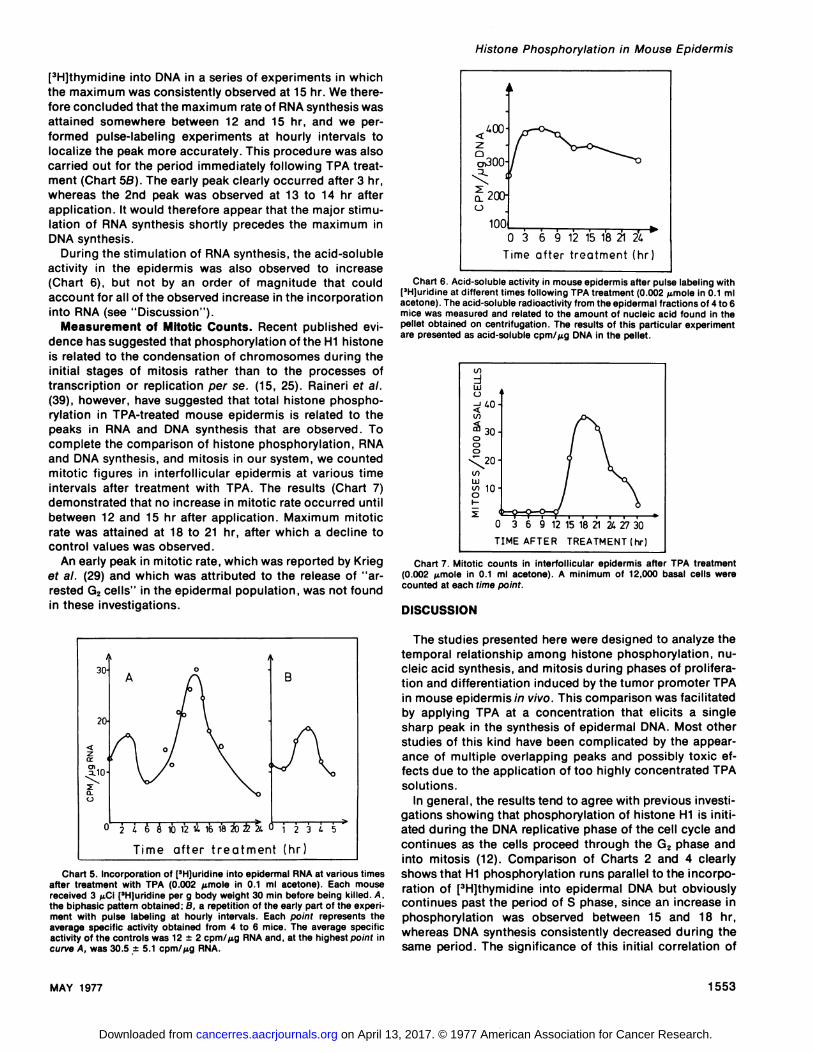
At a dose level of 0.002 @mole/application, TPA elicitedonly 1 sharp peak in the incorporation of [3H]thymidmneintoDNA (Chart 4). The maximum incorporation mate was observed 15 hr after treatment with the tumor promoter andcorresponded to a stimulation of about 500 to 600% withrespect to the control values. Only a very slight depressionin [3H@thyrnidmneincorporation was observed shortly afterTPA application, in contrast to the large depression after
60
a. 30
I—.20
@ 100.C-)
;ORIGIN PHOSPHO- PHOSPHO
THREONINE SERINE
Chart 3. High-voltage electrophoretic separation of phosphorylatedamino acids obtained on hydrolysis of 32P-labeled Hi histone from TPAtreated mouse epidermis. 32P-labeled P (1 mCi/mouse) was injected 17 hrafter treatment with TPA (0.002 @.tmoIein 0.i ml acetone), and the mice werekilled 1 hr later. Unlabeled phosphoserine and phosphothreonine were usedas markers.
Chart 4. Incorporation of[3H)thymidine into epidermal DNA as determinedby pulse labeling at different times after treatment with TPA (0.002 @moIein0.i ml acetone). Each mouse received 1 @Ci[3H)thymidine per g body weight30 mm prior to being killed. Each point represents the average specificactivity of DNA from 4 to 6 mice. This curve was obtained from 1 experimentin a series of 4, all of which gave similar results. The average specific activityof the controls was 7.8 ±i .6 cpm/@g DNA and, at the highest point in thecurve, was 30.8 ±5.4 cpm/@g DNA.
treatment with a dose of 0.02 @molethat has been observedby various authors (4, 10, 29).
RNA Synthesis. RNA synthesis in the epidermis aftertreatment with TPA has been studied by several groups withfairly conflicting results (3, 38). The kinetics of the incorporation of [3H]unidmneinto epidemmal RNA are shown in Chart5. A biphasic stimulation was observed, with an early peakafter 3 hr being followed by a larger peak between 13 and 14hr after treatment. In preliminaryexpeniments, the specificactivity of RNA was determined at 3-hr intervals after treatment with TPA. The major peak seemed to occur sometimesat 12 hr and sometimes at 15 hr after treatment. This contrasted with the results obtained for the incorporation of
[3H]thymidmne into DNA in a series of experiments in whichthe maximum was consistently observed at 15 hr. We therefore concluded that the maximum mateof RNA synthesis wasattained somewhere between 12 and 15 hr, and we penformed pulse-labeling experiments at hourly intervals tolocalize the peak more accurately. This procedure was alsocarried out for the period immediately following TPA treatment (Chart SB). The early peak clearly occurred after 3 hn,whereas the 2nd peak was observed at i 3 to i 4 hr afterapplication. It would therefore appear that the major stimulation of RNA synthesis shortly precedes the maximum inDNA synthesis.
During the stimulation of RNA synthesis, the acid-solubleactivity in the epidermis was also observed to increase(Chart 6), but not by an order of magnitude that couldaccount for all of the observed increase in the incorporationinto RNA (see “Discussion―).
Measurement of Mitotic Counts. Recent published evidence has suggested that phosphomylation of the Hi histoneis related to the condensation of chromosomes during theinitial stages of mitosis rather than to the processes oftranscription or replication per se. (15, 25). Rainemiet at.(39), however, have suggested that total histone phosphorylation in TPA-tmeated mouse epidermis is related to thepeaks in RNA and DNA synthesis that are observed. Tocomplete the Comparison of histone phosphorylation, RNAand DNA synthesis, and mitosis in our system, we countedmitotic figures in interfolliculam epidermis at various timeintervals after treatment with TPA. The results (Chart 7)demonstrated that no increase in mitotic mateoccurred untilbetween 12 and 15 hr after application. Maximum mitoticmatewas attained at 18 to 21 hn, after which a decline tocontrol values was observed.
An early peak in mitotic rate, which was reported by Knieget at. (29) and which was attributed to the release of “amrested G2cells―in the epidemmal population, was not foundin these investigations.
Chart 5. Incorporation of [3H]uridine into epidermal RNA at various timesafter treatment with TPA (0.002 @moIein 0.i ml acetone). Each mousereceived 3 pCi [3H)uridine per g body weight 30 mm before being killed. A,the biphasic pattern obtained; B, a repetition of the early part of the experiment with pulse labeling at hourly intervals. Each point represents theaverage specific activity obtained from 4 to 6 mice. The average specificactivity of the controls was i2 ±2 cpm/@g RNA and, at the highest point incurveA, was30.5 ±5.1cpm/@igRNA.
@20O
1000 3 6 9 12 151'8@12@
Time after treatment (hr)
Chart 6. Acid-soluble activity in mouse epidermis after pulse labeling with[3Hjuridine at different times following TPA treatment (0.002 @molein 0.1 mlacetone). The acid-soluble radioactivity from the epidermal fractions of 4 to 6mice was measured and related to the amount of nucleic acid found in thepellet obtained on centrifugation. The results of this particular experimentare presented as acid-soluble cpm/@g DNA in the pellet.
Chart 7. Mitotic counts in interfollicular epidermis after TPA treatment(0.002 .tmole in 0.1 ml acetone). A minimum of 12,000 basal cells werecounted at each time point.
DISCUSSION
The studies presented here were designed to analyze thetemporal relationship among histone phosphorylation, nucleic acid synthesis, and mitosis during phases of prolifemation and differentiation induced by the tumor promoter TPAin mouse epidermis in vivo. This comparison was facilitatedby applying TPA at a concentration that elicits a singlesharp peak in the synthesis of epidenmal DNA. Most otherstudies of this kind have been complicated by the appearance of multiple overlapping peaks and possibly toxic effects due to the application of too highly concentrated TPAsolutions.
In general, the results tend to agree with previous investigations showing that phosphonylation of histone Hi is initiated during the DNA replicative phase of the cell cycle andcontinues as the cells proceed through the G2 phase andinto mitosis (12). Comparison of Charts 2 and 4 clearlyshows that Hi phosphomylation runs parallel to the incorporation of [3H]thymidine into epidemmal DNA but obviouslycontinues past the period of S phase, since an increase inphosphorylation was observed between 15 and 18 hm,whereas DNA synthesis consistently decreased during thesame period. The significance of this initial correlation of
Fambrough et at. (19) have reported that the proportion ofthe Hi histone in developing pea seeds increases duringmaturation. Recently, Fukuyarna et a!. (20) found that thechromatin of the basal cells of newborn matepidermis contains less of the Hi histone than that of the differentiatedcells of the upper layers.For these reasonsit was of interestto examine any quantitative chan@èsin histone patternsthat might occur in the epidermis during the phases ofproliferation and differentiation. Integration of the stainedpolyacrylamide gels of histone fractions obtained from theepidermis at different times after treatment with TPAshowed that no significant differences occurred. In general,the present results therefore support the conclusions ofother workers that little or no quantitative differences occurin the proportions of the various histones during the prolifemation and differentiation of mammalian cells (44).
Raineni et at. (39) studied the phosphomylation of epidermal histones after TPA treatment and concluded that theobservation of 2 peaks at roughly the same times as peaksin RNA and DNA synthesis provided evidence for the melevance of gene activation to the phenomenon of tumor promotion in mouse skin. It was further claimed that a comrelation exists between the ability of TPA to stimulate histonephosphorylation and its ability to promote tumors. The pattern of histone phosphomylation observed in our experiments is very similar to that in other systems in which theincreased phosphorylation was considered to be directlyrelated to the stimulation of DNA replication and mitoticactivity.
Since the biochemical evidence for specific gene activation by tumor promoters is very sparse, we consider itpremature to conclude that a “conversion―process (13, 26)is necessary for the development of macroscopic tumorsfrom initiated skin. Our data and those of other workers arecompatible with the interpretation that TPA is a substancethat combines the property of being highly hyperplasiogenic for mouse epidermis with a minimum of nonspecifictoxic effects.
ACKNOWLEDGMENTS
We thank Professor Dr. E. Hecker for supplying TPA and Professor Dr. K.Goerttler for his encouragement. We are grateful to Dr. V. Kinzel for assistance with high-voltage electrophoresis.
REFERENCES
1 . Allerton, S. E., and Perlman, G. E. Chemical Characterization of thePhosphoprotein Phosvitin. J. Biol. Chem., 240: 3892-3898, 1965.
2. Bach, H., and Goerttler, K. Morphologische Untersuchungenzur hyperplasiogenen Wirkung des biologisch aktiven Phorbolesters A,. VirchowsArch. Abt. B. Zellpathol., 8: i96-205, i97i.
3. Baird,W. M., Melera,P.W.,andBoutwell,R.K.AcrylamideGelElectrophoresis Studies of the Incorporation of Cytidmne-3Hinto Mouse SkinRNAat Early Times afterTreatment with Phorbol Esters. Cancer Res., 32:781-788, i972.
4. Baird,W.M.,Sedgwick,J. A., andBoutwell,R.K.Effectsof PhorbolandFour Diesters of Phorbol on the Incorporation of Tritiated Precursorsinto DNA, RNA, and Protein in Mouse Epidermis. Cancer Res., 31: i434-1439,1971.
5. Balhom, R., Balhom, M., Morris, H. P., and Chalkley, R. ComparativeHigh-Resolution Electrophoresis of Tumor Histones: Variation in Phosphorylation as a Function of Cell Replication Rate. Cancer Res., 32:1775-1784, 1972.
6. Balhom, R., Bordwell, F., Sellers, L., Granner, D., and Chalkley, R.
Hi phosphorylation with DNA synthesis has, however, beenthe subject of some controversy. In particular, Bmadbury etat. (15) have claimed that the high mateof phosphorylation ofHi observed, for example, during the S phase of regenemating liver (7) may be an artifact and that the true peak inphosphorylation occurs in the late G2 phase. Bmadbuny'sinvestigations were carried out with the slime mold Physarum potycephalum, but similar conclusions have beenreached from experiments with mammalian cells (25). Thepresent results do not provide any definitive solution to thisproblem, since such in vivo systems are not amenable tosynchronization or manipulation in the same way as cellcultures. It is, however, obvious that phosphorylation is notsolely a replication-related phenomenon but may indeed bea prerequisite for the condensation of chromosomes immediately prior to mitosis, as has been suggested (14).
The only other histone that was observed to be phosphorylated to any significant extent in the epidermis was histone H2A, which exhibited a fairly broad phosphorylationpeakfrom 6 to 12 hr after treatment. Interestingly, histonesHi and H2A were reported by Sherod et at. (42) to be theonly histones phosphorylated in mouse tissues. In addition,Gumleyand Walters (23) and Chalkley's group (6) obtainedsimilar results using synchronized Chinese hamster cells orMorris hepatorna 7288C cells in culture. The different kinetics of phosphomylation of Hi and H2A histones during thecell cycle led Gumleyet at. (24) to conclude that the phosphorylation of H2A may be related to processes other thanthe replication of DNA or mitosis.
The biphasic stimulation of [3H]umidineincorporation intoepidermal RNA after treatment with TPA is not exactly cornpatible with the results of Baird et at. (3) and Raick (38).These authors, however, applied higher doses of TPA todifferent strains of mice, and direct comparison with theresults of the present investigations is therefore questionable. The early peak in [3Hjumidine incorporation into RNA 3hr after application of TPA may be in part due to pemrneability changes, which result in fluctuations in the specificactivity of the intracellular UTP pool. A similar early “RNAsynthesis―peak in the estrogen-stimulated uterus was latershown by Means and Hamilton (35) to be attributable tosuch pool changes. However, this would not seem to be thesole factor operating here, since a more accurate analysisof the newly synthesized RNA by Baird et a!. (3) suggestedthat the matesof synthesis of the various RNA species werenot all stimulated to the same extent. This would not havebeen the case if pool changes alone were responsible forthe early peak in incorporation. The major peak in[3H]umidine incorporation (Chart 5) would appear to be dueto de novo RNA synthesis, particularly since electron microscopic observations have demonstrated a large stimulationof nibosome production during the premeplicative phase following TPA treatment (38). This peak does indeed shortlyprecede the maximum in DNA synthesis. Similar resultswere obtained with synchronized cell cultures (ii).
The proliferative phase resulting from treatment of theepidermis with TPA is followed by a distinct wave of differentiation that has been characterized morphologically andbiochemically. Several reports have appeared in the literatune to the effect that the complement of histones changeswith development and differentiation (33). In particular,
Histone Phosphorylation and DNA Synthesis Are Linked in SynchronousCultures of HTC Cells. Biochem. Biophys. Res. Commun., 46: 1326-1333, 1972.
7. Balhom, R., Riecke, 0., and Chalkley, R. Rapid Electrophoretic Analysisfor Histone Phosphorylation. A Reinvestigation of Phosphorylation ofLysine-Rich Histone during Rat Liver Regeneration. Biochemistry, 10:3952-3959,1971.
8. Balmain, A. The synthesisof Specific Proteinsin Mutt Mouse Epidermisduring Phases of Proliferation and Differentiation Induced by the TumorPromoter TPA, and in Basal and Differentiating Layers of NeonatalMouse Epidermis. J. Invest. Dermatol., 67: 246-253, 1976.
9. Balmain,A., andHecker,E.OnTheBiochemicalMechanismof Tumorigenesisin MouseSkin. VI. EarlyEffectsof Growth-StimulatingPhorbolEsters on Phosphate Transport and Phospholipid Synthesis in MouseEpidermis. Biochim. Biophys. Acta, 362: 457-468, 1974.
10. Balmain, A., and Hecker, E. On The Biochemical Mechanism of Tumorigenesis in Mouse Skin. VII. The Effects of Tumor Promoters on 3H-Choline and 3H-Glycerol Incorporation into Mouse Epidermal Phosphatidylcholine in Relation to Their Effects on 3H-Thymidine Incorporationinto DNA. Z. Krebsforsch., 86: 251-261 , 1976.
11. Benjamin, W. B. Selective in vitro Methylation of Rat Chromatin Associated Histone after Partial Hepatectomy. Nature New Biol., 234: 18-20,1971.
12. Borun, T. W., Paik, W. K., and Marks, D. Histone Phosphorylation andthe Control of Cellular Proliferation. In: M. A. Mehlman and R. W.Hanson (eds.), Control Processes in Neoplasia, pp. i87-2i9. New York:Acedemic Press, Inc., 1974.
13. Boutwell, R. K. Function and Mechanism of Promoters of Carcinogenssis. CRC Critical Rev. Toxicol., 2: 419-442, 1974.
14. Bradbury, E. M., Inglis, R. J., and Matthews, H. R. Control of CellDivision by Very Lysine Rich Histone (Fl) Phosphorylation. Nature, 247:257-261.1974.
15. Bradbury, E. M., Inglis, A. J., and Matthews, H. R. Molecular Basis ofControl of Mitotic Cell Division in Eucaryotes. Nature, 249: 553-556,1974.
16. Brasch, K., Adams, G. H. M., and Neelin, J. M. Evidence for ErythrocyteSpecific Histone Modification and Structural Changes in Chromatin during Goose Erythrocyte Maturation. J. Cell Sci., 15: 659-677, 1974.
17. Breindl, M., and Gallwitz, D. Identification of Histone Messenger RNAfrom HeLa Cells. European J. Biochem., 32: 381-391 , 1973.
18. Burton, K. A Study of the Conditions and Mechanism of the Diphenylamine Reaction for the Colorimetnic Estimation of Deoxyribonucleic Acid.Biochem.J.,62:315-323,1956.
19. Fambrough, D. M., Fujimura, F., and Bonner, J. Quantitative DistributionofHistoneComponentsinthePeaPlant.Biochemistry,7:575-585,1968.
20. Fukuyama, K., 5eki, N., Nishita, K., and Epstein, W. L. Studiesof Histonein Differentiated and Undifferentiated Epidermal Cells. Joint Meeting,Society for Investigative Dermatology and European Society for Dermatological Research, Amsterdam, June 9-13, 1975, Abstracts, p. 16.
21. Gordon, A. H. Electrophoresisof Proteins in Polyacrylamideand StarchGels. In: T. 5. Work and E. Work (ads.), Laboratory Techniques in Biochemistry and Molecular Biology, pp. 26-27. Amsterdam/New York:North Holland/American Elsevier, I 969.
22. Gorovsky,M. A., and Keevert,J. B. Absenceof HistoneFl in a MitoticallyDividing, Genetically Inactive Nucleus. Proc. NatI. Acad. Sci. U. S., 72:2672-2676,1975.
23. Gurley, L. R., and Walters, R. A. Response of Histone Turnover andPhosphorylation to X-Irradiation. Biochemistry, 10: 1588-1593, i97i.
24. Gurley, L. R., Wafters, R. A., and Tobey, R. A. The Metabolismof HistoneFractions. VI. Differences in the Phosphorylatlon of Histone Fractions
during the Cell Cycle. Arch. Biochem. Biophys., 154: 212-218, i972.25. Gurley, L. R., Walters, A. A., and Tobey, R. A. Histone Phosphorylatlon
in Late Interphase and Mitosis. Biochem. Biophys. Res. Commun., 50:744—750,i973.
26. Hennings,H., and Boutwell, A. K. Studieson the Mechanismof SkinTumor Promotion. Cancer Res., 30: 312-320, 1970.
27. Johns, E. W. Studies on Histones. Biochem. J., 92: 55-59, 1964.28. Kleinsmith, L. G., Allfrey, V. G., and Mirsky, A. E. Phosphoprotein
Metabolism in Isolated Lymphocyte Nuclei. Proc. NatI. Acad. Sci. U. S.,55: 1182-1189, 1966.
29. Krieg, L., Ki]hlmann, I., and Marks, F. Effects of Tumor-promoting Phorbol Esters and of Acetic Acid on Mechanisms Controlling DNA Synthesisand Mitosis (Chalones) and on the Biosynthesis of Histidine-rich Proteinin Mouse Epidermis. Cancer Res., 34: 3135-3146, 1974.
30. Laemmli, U. K. Cleavage of Structural Proteins during the Assembly ofthe Head of Bacteriophage T4. Nature, 227: 680-685, 1970.
31. Langan, T. A. Histone Phosphorylation: Stimulation of Adenosine 3'S'-Monophosphate. Science, 162: 579-580, 1968.
32. MacGillivray, A. J., Cameron, A., Krauze, R. J., Rickwood, D., and Paul,J. The Non-histone Proteins of Chromatin. Their Isolation and Composition in a Number of Tissues. Biochim. Biophys. Acta, 277: 384—402,1972.
33. MacGillivray, A. J., and Rickwood, D.The RoleofChromosomal Proteinsas Gene Regulators. In: J. Paul (ed), Biochemistry of Cell Differentiation, pp. 301-361 . London: Butterworths, 1974.
34. Marks, D. B., Paik, W. K., and Borun, T. W. The Relationship of HistonePhosphorylation to Deoxyribonucleic Acid Replication and Mitosis during the HeLa 5-3 Cell Cycle. J. Biol. Chem., 248: 5660-5667, i973.
35. Means, A. R., and Hamilton, T. H. Early Estrogen Action: ConcomitantStimulations within Two Minutes of Nuclear RNA Synthesis and Uptakeof RNA Precursor by the Uterus. Proc. NatI. Acad. Sci. U.S., 56: 1594-1598, 1966.
36. Oliver, D., Balhorn, D., Granner, D., and Chalkley, R. Molecular Natureof Fl Histone Phosphorylation in Cultured Hepatoma Cells. Biochemistry, 11: 3921-3925, 1972.
37. Panyim, S., and Chalkley, R. High Resolution Acrylamide Gel Electrophoresis of Histones. Arch. Biochem. Biophys., 130: 337-346, 1969.
38. Raick, A. N. Ultrastructural, Histological, and Biochemical AlterationsProduced by i2-O-Tetradecanoyl-phorbol-13-acetate on Mouse Epidermis and Their Relevance to Skin Tumor Promotion. Cancer Res., 33:269-286, i973.
39. Raineri, R., Simsiman, R. C., and Boutwell, R. K. Stimulation of thePhosphorylatlon of Mouse Epidermal Histones by Tumor-promotingAgents. Cancer Res., 33: 134-139, 1973.
40. SchneIder, W. C. Determination of Nucleic Acids in Tissues by PentoseAnalysis. Methods Enzymol., 3: 680-684, 1957.
41. Segal, A., Schroeder, M., Bamett, P., and Van Duuren, B. L. Studies ofthe Effects in Vitro of $-Propiolactone and @-Propiolact-('@C)-oneonWhole Mouse Skin Chromatin. Biochem. Pharmacol. ,23: 937-946. 1974.
42. Sherod, D., Johnson, G., and Chalkley, R. Phosphorylationof MouseAscites Tumor Cell Lysine-Rich Histone. Biochemistry, 9: 4611-4615,1970.
43. Slaga, T. J., Des, S. B., Rice, J. M., and Thomson, S. Fractionation ofMouse Epidermal Chromatin Components. J. Invest. Dermatol., 63: 343-349, 1974.
44. Stellwagen, A. H., and Cole, R. D. Comparison of Histones Obtainedfrom Mammary Gland at Different Stages of Development and Lactation.J. Biol. Chem., 243: 4456-4462, 1968.
45. Trevithick, J. R. Patterns of Incorporation of 32@Orthophosphate and 3HAcetate into Histones of Rainbow Trout during Early Development. Can.J. Biochem., 52: 399-405, 1974.
1977;37:1548-1555. Cancer Res Allan Balmain, Angel Alonso and Jürgen Fischer -Tetradecanoyl-phorbol-13-acetate
OEpidermis by the Tumor Promoter 12-Phases of Proliferation and Differentiation Induced in Mouse Histone Phosphorylation and Synthesis of DNA and RNA during